Explore
Explore Validate
Validate Learn
Learn Western blot
Western blotAntibody data
- Antibody Data
- Antigen structure
- References [54]
- Comments [0]
- Validations
- Western blot [2]
- Other assay [30]
Submit
Validation data
Reference
Comment
Report error
- Product number
- PA1-987 - Provider product page

- Provider
- Invitrogen Antibodies
- Product name
- GAPDH Polyclonal Antibody
- Antibody type
- Polyclonal
- Antigen
- Synthetic peptide
- Description
- PA1-987 detects rat GAPDH. Shares 100% sequence homology with human and mouse.
- Concentration
- 1 mg/mL
Submitted references RIP3 Contributes to Cardiac Hypertrophy by Influencing MLKL-Mediated Calcium Influx.
Temporal and Sex-Linked Protein Expression Dynamics in a Familial Model of Alzheimer's Disease.
MAD2L2 promotes replication fork protection and recovery in a shieldin-independent and REV3L-dependent manner.
Time-limited alterations in cortical activity of a knock-in mouse model of KCNQ2-related developmental and epileptic encephalopathy.
C/D box snoRNA SNORD113-6/AF357425 plays a dual role in integrin signalling and arterial fibroblast function via pre-mRNA processing and 2'O-ribose methylation.
Neuropilin-1, a myeloid cell-specific protein, is an inhibitor of HIV-1 infectivity.
An Orally Bioavailable and Highly Efficacious Inhibitor of CDK9/FLT3 for the Treatment of Acute Myeloid Leukemia.
Membrane Protein OTOF Is a Type I Interferon-Induced Entry Inhibitor of HIV-1 in Macrophages.
Retinal degeneration-3 protein attenuates photoreceptor degeneration in transgenic mice expressing dominant mutation of human retinal guanylyl cyclase.
Zinc oxide nanosphere for hydrogen sulfide scavenging and ferroptosis of colorectal cancer.
Aging lens epithelium is susceptible to ferroptosis.
Low-density lipoprotein receptor-related protein 1 (LRP1) is a novel receptor for apolipoprotein A4 (APOA4) in adipose tissue.
Vpr counteracts the restriction of LAPTM5 to promote HIV-1 infection in macrophages.
Depletion of Retinal Dopaminergic Activity in a Mouse Model of Rod Dysfunction Exacerbates Experimental Autoimmune Uveoretinitis: A Role for the Gateway Reflex.
Small molecule splicing modifiers with systemic HTT-lowering activity.
Mitotic Activation Around Wound Edges and Epithelialization Repair in UVB-Induced Capsular Cataracts.
Tonic prime-boost of STING signalling mediates Niemann-Pick disease type C.
Cellular FLICE-like inhibitory protein (cFLIP) critically maintains apoptotic resistance in human lens epithelial cells.
S-Nitrosylation of G protein-coupled receptor kinase 6 and Casein kinase 2 alpha modulates their kinase activity toward alpha-synuclein phosphorylation in an animal model of Parkinson's disease.
Serum Exosomal Proteins F9 and TSP-1 as Potential Diagnostic Biomarkers for Newly Diagnosed Epilepsy.
Tau Oligomers and Fibrils Exhibit Differential Patterns of Seeding and Association With RNA Binding Proteins.
Inflammasome Sensor NLRP1 Confers Acquired Drug Resistance to Temozolomide in Human Melanoma.
miR-19b targets pulmonary endothelial syndecan-1 following hemorrhagic shock.
Skeletal Muscle Anti-Atrophic Effects of Leucine Involve Myostatin Inhibition.
Stat3 activation induces insulin resistance via a muscle-specific E3 ubiquitin ligase Fbxo40.
RNA Sequence Analyses throughout the Course of Mouse Cardiac Laminopathy Identify Differentially Expressed Genes for Cell Cycle Control and Mitochondrial Function.
lncRNA ZEB1-AS1 Mediates Oxidative Low-Density Lipoprotein-Mediated Endothelial Cells Injury by Post-transcriptional Stabilization of NOD2.
Enantioselective Synthesis of 8-Hydroxyquinoline Derivative, Q134 as a Hypoxic Adaptation Inducing Agent.
Staphylococcus aureus Exploits the Host Apoptotic Pathway To Persist during Infection.
Identification of Novel Inhibitors of DLK Palmitoylation and Signaling by High Content Screening.
Increased expression of FAK isoforms as potential cancer biomarkers in ovarian cancer.
lncRNA ZEB1-AS1 promotes migration and metastasis of bladder cancer cells by post-transcriptional activation of ZEB1.
Activation of LncRNA TINCR by H3K27 acetylation promotes Trastuzumab resistance and epithelial-mesenchymal transition by targeting MicroRNA-125b in breast Cancer.
miRNA‑183‑5p.1 promotes the migration and invasion of gastric cancer AGS cells by targeting TPM1.
lncRNA PLAC2 activated by H3K27 acetylation promotes cell proliferation and invasion via the activation of Wnt/β‑catenin pathway in oral squamous cell carcinoma.
Transgenic overexpression of furin increases epileptic susceptibility.
Metabolomics reveals critical adrenergic regulatory checkpoints in glycolysis and pentose-phosphate pathways in embryonic heart.
Rapid estrogen receptor-α signaling mediated by ERK activation regulates vascular tone in male and ovary-intact female mice.
Exploration of the regulation and control mechanisms of miR-145 in trophoblast cell proliferation and invasion.
Secreted Giardia intestinalis cysteine proteases disrupt intestinal epithelial cell junctional complexes and degrade chemokines.
CRISPR-Cas9-enabled genetic disruptions for understanding ethanol and ethyl acetate biosynthesis in Kluyveromyces marxianus.
Impact of R264C and R264H polymorphisms in human aromatase function.
Chlorogenic acid prevents inflammatory responses in IL‑1β‑stimulated human SW‑1353 chondrocytes, a model for osteoarthritis.
Repression of Stress-Induced LINE-1 Expression Protects Cancer Cell Subpopulations from Lethal Drug Exposure.
Uterotonic Neuromedin U Receptor 2 and Its Ligands Are Upregulated by Inflammation in Mice and Humans, and Elicit Preterm Birth.
Curcumin eluting nanofibers augment osteogenesis toward phytochemical based bone tissue engineering.
The pregnane X receptor down-regulates organic cation transporter 1 (SLC22A1) in human hepatocytes by competing for ("squelching") SRC-1 coactivator.
Human eIF3b and eIF3a serve as the nucleation core for the assembly of eIF3 into two interconnected modules: the yeast-like core and the octamer.
An animal model for chorioamnionitis at term.
Wnt5a and Wnt11 inhibit the canonical Wnt pathway and promote cardiac progenitor development via the Caspase-dependent degradation of AKT.
DAAM1 and DAAM2 are co-required for myocardial maturation and sarcomere assembly.
Intestinal fatty acid binding protein (fabp2) in Atlantic salmon (Salmo salar): Localization and alteration of expression during development of diet induced enteritis.
Impaired β-adrenergic responsiveness accentuates dysfunctional excitation-contraction coupling in an ovine model of tachypacing-induced heart failure.
An antibody-drug conjugate targeting the endothelin B receptor for the treatment of melanoma.
Xue H, Shi H, Zhang F, Li H, Li C, Han Q
Oxidative medicine and cellular longevity 2022;2022:5490553
Oxidative medicine and cellular longevity 2022;2022:5490553
Temporal and Sex-Linked Protein Expression Dynamics in a Familial Model of Alzheimer's Disease.
Blasco Tavares Pereira Lopes F, Schlatzer D, Wang R, Li X, Feng E, Koyutürk M, Qi X, Chance MR
Molecular & cellular proteomics : MCP 2022 Sep;21(9):100280
Molecular & cellular proteomics : MCP 2022 Sep;21(9):100280
MAD2L2 promotes replication fork protection and recovery in a shieldin-independent and REV3L-dependent manner.
Paniagua I, Tayeh Z, Falcone M, Hernández Pérez S, Cerutti A, Jacobs JJL
Nature communications 2022 Sep 8;13(1):5167
Nature communications 2022 Sep 8;13(1):5167
Time-limited alterations in cortical activity of a knock-in mouse model of KCNQ2-related developmental and epileptic encephalopathy.
Biba-Maazou N, Becq H, Pallesi-Pocachard E, Sarno S, Granjeaud S, Montheil A, Kurz M, Villard L, Milh M, Santini PL, Aniksztejn L
The Journal of physiology 2022 May;600(10):2429-2460
The Journal of physiology 2022 May;600(10):2429-2460
C/D box snoRNA SNORD113-6/AF357425 plays a dual role in integrin signalling and arterial fibroblast function via pre-mRNA processing and 2'O-ribose methylation.
van Ingen E, van den Homberg DAL, van der Bent ML, Mei H, Papac-Milicevic N, Kremer V, Boon RA, Quax PHA, Wojta J, Nossent AY
Human molecular genetics 2022 Mar 31;31(7):1051-1066
Human molecular genetics 2022 Mar 31;31(7):1051-1066
Neuropilin-1, a myeloid cell-specific protein, is an inhibitor of HIV-1 infectivity.
Wang S, Zhao L, Zhang X, Zhang J, Shang H, Liang G
Proceedings of the National Academy of Sciences of the United States of America 2022 Jan 11;119(2)
Proceedings of the National Academy of Sciences of the United States of America 2022 Jan 11;119(2)
An Orally Bioavailable and Highly Efficacious Inhibitor of CDK9/FLT3 for the Treatment of Acute Myeloid Leukemia.
Anshabo AT, Bantie L, Diab S, Lenjisa J, Kebede A, Long Y, Heinemann G, Karanjia J, Noll B, Basnet SKC, Li M, Milne R, Albrecht H, Wang S
Cancers 2022 Feb 22;14(5)
Cancers 2022 Feb 22;14(5)
Membrane Protein OTOF Is a Type I Interferon-Induced Entry Inhibitor of HIV-1 in Macrophages.
Ding H, Zhang X, Zhu Z, Wang S, Xiong Y, Shang H, Liang G
mBio 2022 Aug 30;13(4):e0173822
mBio 2022 Aug 30;13(4):e0173822
Retinal degeneration-3 protein attenuates photoreceptor degeneration in transgenic mice expressing dominant mutation of human retinal guanylyl cyclase.
Peshenko IV, Olshevskaya EV, Dizhoor AM
The Journal of biological chemistry 2021 Oct;297(4):101201
The Journal of biological chemistry 2021 Oct;297(4):101201
Zinc oxide nanosphere for hydrogen sulfide scavenging and ferroptosis of colorectal cancer.
Pan X, Qi Y, Du Z, He J, Yao S, Lu W, Ding K, Zhou M
Journal of nanobiotechnology 2021 Nov 27;19(1):392
Journal of nanobiotechnology 2021 Nov 27;19(1):392
Aging lens epithelium is susceptible to ferroptosis.
Wei Z, Hao C, Huangfu J, Srinivasagan R, Zhang X, Fan X
Free radical biology & medicine 2021 May 1;167:94-108
Free radical biology & medicine 2021 May 1;167:94-108
Low-density lipoprotein receptor-related protein 1 (LRP1) is a novel receptor for apolipoprotein A4 (APOA4) in adipose tissue.
Qu J, Fourman S, Fitzgerald M, Liu M, Nair S, Oses-Prieto J, Burlingame A, Morris JH, Davidson WS, Tso P, Bhargava A
Scientific reports 2021 Jun 24;11(1):13289
Scientific reports 2021 Jun 24;11(1):13289
Vpr counteracts the restriction of LAPTM5 to promote HIV-1 infection in macrophages.
Zhao L, Wang S, Xu M, He Y, Zhang X, Xiong Y, Sun H, Ding H, Geng W, Shang H, Liang G
Nature communications 2021 Jun 17;12(1):3691
Nature communications 2021 Jun 17;12(1):3691
Depletion of Retinal Dopaminergic Activity in a Mouse Model of Rod Dysfunction Exacerbates Experimental Autoimmune Uveoretinitis: A Role for the Gateway Reflex.
Stofkova A, Zloh M, Andreanska D, Fiserova I, Kubovciak J, Hejda J, Kutilek P, Murakami M
International journal of molecular sciences 2021 Dec 31;23(1)
International journal of molecular sciences 2021 Dec 31;23(1)
Small molecule splicing modifiers with systemic HTT-lowering activity.
Bhattacharyya A, Trotta CR, Narasimhan J, Wiedinger KJ, Li W, Effenberger KA, Woll MG, Jani MB, Risher N, Yeh S, Cheng Y, Sydorenko N, Moon YC, Karp GM, Weetall M, Dakka A, Gabbeta V, Naryshkin NA, Graci JD, Tripodi T Jr, Southwell A, Hayden M, Colacino JM, Peltz SW
Nature communications 2021 Dec 15;12(1):7299
Nature communications 2021 Dec 15;12(1):7299
Mitotic Activation Around Wound Edges and Epithelialization Repair in UVB-Induced Capsular Cataracts.
Wei Z, Hao C, Srinivasagan R, Wu H, Chen JK, Fan X
Investigative ophthalmology & visual science 2021 Dec 1;62(15):29
Investigative ophthalmology & visual science 2021 Dec 1;62(15):29
Tonic prime-boost of STING signalling mediates Niemann-Pick disease type C.
Chu TT, Tu X, Yang K, Wu J, Repa JJ, Yan N
Nature 2021 Aug;596(7873):570-575
Nature 2021 Aug;596(7873):570-575
Cellular FLICE-like inhibitory protein (cFLIP) critically maintains apoptotic resistance in human lens epithelial cells.
Huangfu J, Hao C, Wei Z, Wormstone IM, Yan H, Fan X
Cell death & disease 2021 Apr 9;12(4):386
Cell death & disease 2021 Apr 9;12(4):386
S-Nitrosylation of G protein-coupled receptor kinase 6 and Casein kinase 2 alpha modulates their kinase activity toward alpha-synuclein phosphorylation in an animal model of Parkinson's disease.
Wu W, Sung CC, Yu P, Li J, Chung KKK
PloS one 2020;15(4):e0232019
PloS one 2020;15(4):e0232019
Serum Exosomal Proteins F9 and TSP-1 as Potential Diagnostic Biomarkers for Newly Diagnosed Epilepsy.
Lin Z, Gu Y, Zhou R, Wang M, Guo Y, Chen Y, Ma J, Xiao F, Wang X, Tian X
Frontiers in neuroscience 2020;14:737
Frontiers in neuroscience 2020;14:737
Tau Oligomers and Fibrils Exhibit Differential Patterns of Seeding and Association With RNA Binding Proteins.
Jiang L, Zhao J, Cheng JX, Wolozin B
Frontiers in neurology 2020;11:579434
Frontiers in neurology 2020;11:579434
Inflammasome Sensor NLRP1 Confers Acquired Drug Resistance to Temozolomide in Human Melanoma.
Zhai Z, Samson JM, Yamauchi T, Vaddi PK, Matsumoto Y, Dinarello CA, Ravindran Menon D, Fujita M
Cancers 2020 Sep 4;12(9)
Cancers 2020 Sep 4;12(9)
miR-19b targets pulmonary endothelial syndecan-1 following hemorrhagic shock.
Wu F, Wang JY, Chao W, Sims C, Kozar RA
Scientific reports 2020 Sep 25;10(1):15811
Scientific reports 2020 Sep 25;10(1):15811
Skeletal Muscle Anti-Atrophic Effects of Leucine Involve Myostatin Inhibition.
Cruz A, Ferian A, Alves PKN, Silva WJ, Bento MR, Gasch A, Labeit S, Moriscot AS
DNA and cell biology 2020 Nov 2;
DNA and cell biology 2020 Nov 2;
Stat3 activation induces insulin resistance via a muscle-specific E3 ubiquitin ligase Fbxo40.
Zhang L, Chen Z, Wang Y, Tweardy DJ, Mitch WE
American journal of physiology. Endocrinology and metabolism 2020 May 1;318(5):E625-E635
American journal of physiology. Endocrinology and metabolism 2020 May 1;318(5):E625-E635
RNA Sequence Analyses throughout the Course of Mouse Cardiac Laminopathy Identify Differentially Expressed Genes for Cell Cycle Control and Mitochondrial Function.
Shao Z, Koh W, Ni Y, Li W, Agatisa-Boyle B, Merkurjev D, Tang WHW
Scientific reports 2020 Apr 20;10(1):6632
Scientific reports 2020 Apr 20;10(1):6632
lncRNA ZEB1-AS1 Mediates Oxidative Low-Density Lipoprotein-Mediated Endothelial Cells Injury by Post-transcriptional Stabilization of NOD2.
Xu X, Ma C, Duan Z, Du Y, Liu C
Frontiers in pharmacology 2019;10:397
Frontiers in pharmacology 2019;10:397
Enantioselective Synthesis of 8-Hydroxyquinoline Derivative, Q134 as a Hypoxic Adaptation Inducing Agent.
Hackler L Jr, Gyuris M, Huzián O, Alföldi R, Szebeni GJ, Madácsi R, Knapp L, Kanizsai I, Puskás LG
Molecules (Basel, Switzerland) 2019 Nov 23;24(23)
Molecules (Basel, Switzerland) 2019 Nov 23;24(23)
Staphylococcus aureus Exploits the Host Apoptotic Pathway To Persist during Infection.
Winstel V, Schneewind O, Missiakas D
mBio 2019 Nov 12;10(6)
mBio 2019 Nov 12;10(6)
Identification of Novel Inhibitors of DLK Palmitoylation and Signaling by High Content Screening.
Martin DDO, Kanuparthi PS, Holland SM, Sanders SS, Jeong HK, Einarson MB, Jacobson MA, Thomas GM
Scientific reports 2019 Mar 6;9(1):3632
Scientific reports 2019 Mar 6;9(1):3632
Increased expression of FAK isoforms as potential cancer biomarkers in ovarian cancer.
Nolasco-Quiroga M, Rosas-Díaz M, Moreno J, Godínez-Aguilar R, López-Ibarra MJ, Piña-Sánchez P, Alvarado-Cabrero I, Vázquez-Gómez G, Rocha-Zavaleta L, Arenas-Aranda D, Salamanca-Gómez F
Oncology letters 2019 Jun;17(6):4779-4786
Oncology letters 2019 Jun;17(6):4779-4786
lncRNA ZEB1-AS1 promotes migration and metastasis of bladder cancer cells by post-transcriptional activation of ZEB1.
Zhao X, Wang D, Ding Y, Zhou J, Liu G, Ji Z
International journal of molecular medicine 2019 Jul;44(1):196-206
International journal of molecular medicine 2019 Jul;44(1):196-206
Activation of LncRNA TINCR by H3K27 acetylation promotes Trastuzumab resistance and epithelial-mesenchymal transition by targeting MicroRNA-125b in breast Cancer.
Dong H, Hu J, Zou K, Ye M, Chen Y, Wu C, Chen X, Han M
Molecular cancer 2019 Jan 8;18(1):3
Molecular cancer 2019 Jan 8;18(1):3
miRNA‑183‑5p.1 promotes the migration and invasion of gastric cancer AGS cells by targeting TPM1.
Lin J, Shen J, Yue H, Cao Z
Oncology reports 2019 Dec;42(6):2371-2381
Oncology reports 2019 Dec;42(6):2371-2381
lncRNA PLAC2 activated by H3K27 acetylation promotes cell proliferation and invasion via the activation of Wnt/β‑catenin pathway in oral squamous cell carcinoma.
Chen F, Qi S, Zhang X, Wu J, Yang X, Wang R
International journal of oncology 2019 Apr;54(4):1183-1194
International journal of oncology 2019 Apr;54(4):1183-1194
Transgenic overexpression of furin increases epileptic susceptibility.
Yang Y, He M, Tian X, Guo Y, Liu F, Li Y, Zhang H, Lu X, Xu D, Zhou R, Ma Y, Wang W, Chen G, Hu Y, Wang X
Cell death & disease 2018 Oct 17;9(11):1058
Cell death & disease 2018 Oct 17;9(11):1058
Metabolomics reveals critical adrenergic regulatory checkpoints in glycolysis and pentose-phosphate pathways in embryonic heart.
Peoples JNR, Maxmillian T, Le Q, Nadtochiy SM, Brookes PS, Porter GA Jr, Davidson VL, Ebert SN
The Journal of biological chemistry 2018 May 4;293(18):6925-6941
The Journal of biological chemistry 2018 May 4;293(18):6925-6941
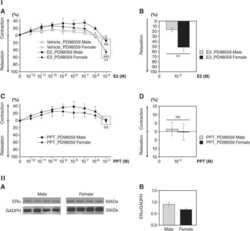
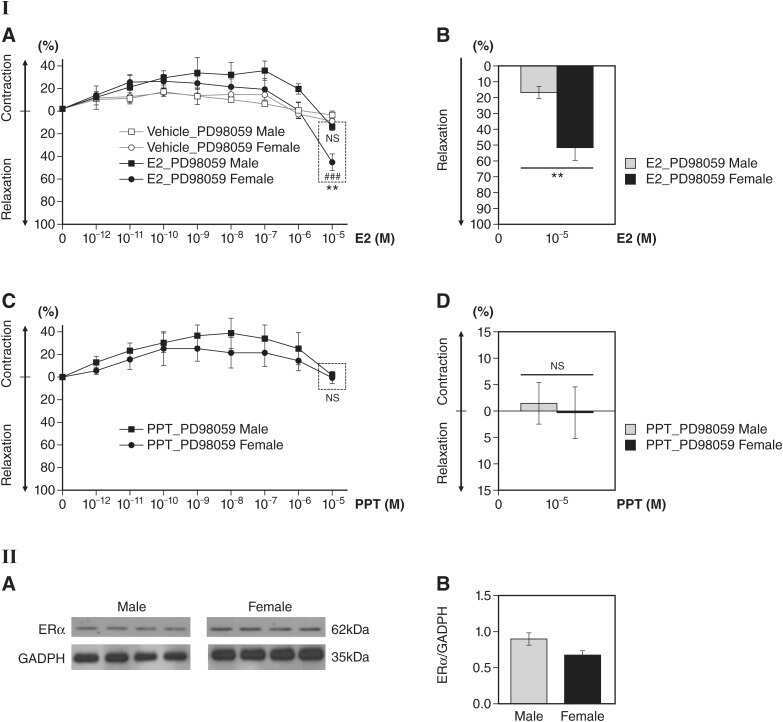
Rapid estrogen receptor-α signaling mediated by ERK activation regulates vascular tone in male and ovary-intact female mice.
Kim SC, Boese AC, Moore MH, Cleland RM, Chang L, Delafontaine P, Yin KJ, Lee JP, Hamblin MH
American journal of physiology. Heart and circulatory physiology 2018 Feb 1;314(2):H330-H342
American journal of physiology. Heart and circulatory physiology 2018 Feb 1;314(2):H330-H342
Exploration of the regulation and control mechanisms of miR-145 in trophoblast cell proliferation and invasion.
Chi Z, Zhang M
Experimental and therapeutic medicine 2018 Dec;16(6):5298-5304
Experimental and therapeutic medicine 2018 Dec;16(6):5298-5304
Secreted Giardia intestinalis cysteine proteases disrupt intestinal epithelial cell junctional complexes and degrade chemokines.
Liu J, Ma'ayeh S, Peirasmaki D, Lundström-Stadelmann B, Hellman L, Svärd SG
Virulence 2018 Dec 31;9(1):879-894
Virulence 2018 Dec 31;9(1):879-894
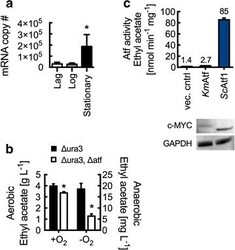
CRISPR-Cas9-enabled genetic disruptions for understanding ethanol and ethyl acetate biosynthesis in Kluyveromyces marxianus.
Löbs AK, Engel R, Schwartz C, Flores A, Wheeldon I
Biotechnology for biofuels 2017;10:164
Biotechnology for biofuels 2017;10:164
Impact of R264C and R264H polymorphisms in human aromatase function.
Baravalle R, Di Nardo G, Bandino A, Barone I, Catalano S, Andò S, Gilardi G
The Journal of steroid biochemistry and molecular biology 2017 Mar;167:23-32
The Journal of steroid biochemistry and molecular biology 2017 Mar;167:23-32
Chlorogenic acid prevents inflammatory responses in IL‑1β‑stimulated human SW‑1353 chondrocytes, a model for osteoarthritis.
Liu CC, Zhang Y, Dai BL, Ma YJ, Zhang Q, Wang Y, Yang H
Molecular medicine reports 2017 Aug;16(2):1369-1375
Molecular medicine reports 2017 Aug;16(2):1369-1375
Repression of Stress-Induced LINE-1 Expression Protects Cancer Cell Subpopulations from Lethal Drug Exposure.
Guler GD, Tindell CA, Pitti R, Wilson C, Nichols K, KaiWai Cheung T, Kim HJ, Wongchenko M, Yan Y, Haley B, Cuellar T, Webster J, Alag N, Hegde G, Jackson E, Nance TL, Giresi PG, Chen KB, Liu J, Jhunjhunwala S, Settleman J, Stephan JP, Arnott D, Classon M
Cancer cell 2017 Aug 14;32(2):221-237.e13
Cancer cell 2017 Aug 14;32(2):221-237.e13
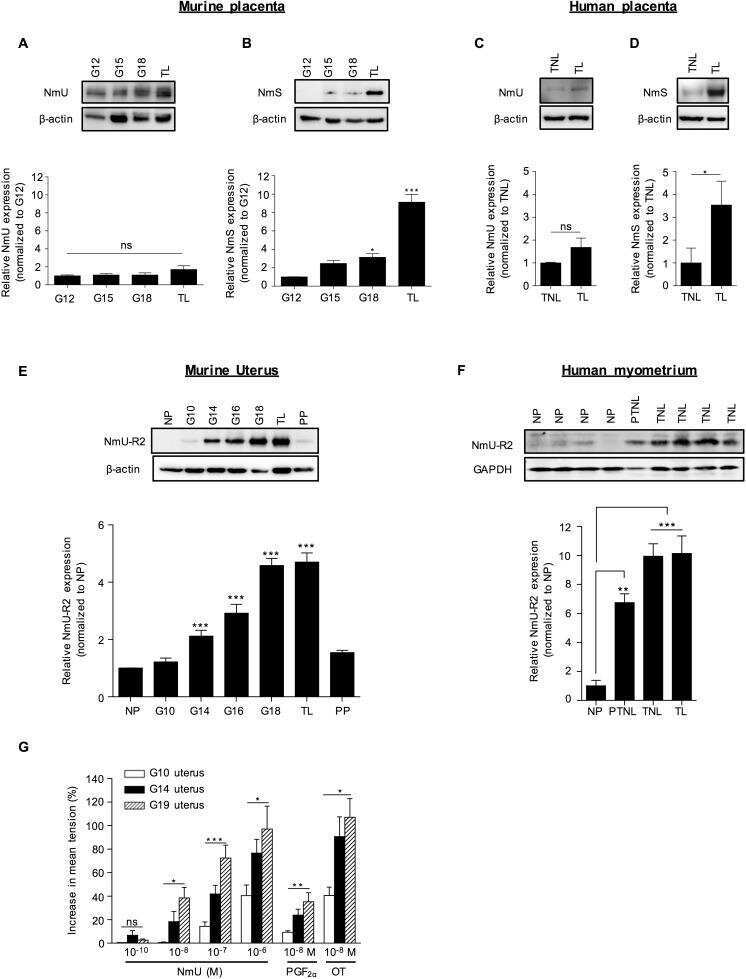
Uterotonic Neuromedin U Receptor 2 and Its Ligands Are Upregulated by Inflammation in Mice and Humans, and Elicit Preterm Birth.
Nadeau-Vallée M, Boudreault A, Leimert K, Hou X, Obari D, Madaan A, Rouget R, Zhu T, Belarbi L, Brien MÈ, Beaudry-Richard A, Olson DM, Girard S, Chemtob S
Biology of reproduction 2016 Sep;95(3):72
Biology of reproduction 2016 Sep;95(3):72
Curcumin eluting nanofibers augment osteogenesis toward phytochemical based bone tissue engineering.
Jain S, Krishna Meka SR, Chatterjee K
Biomedical materials (Bristol, England) 2016 Oct 6;11(5):055007
Biomedical materials (Bristol, England) 2016 Oct 6;11(5):055007
The pregnane X receptor down-regulates organic cation transporter 1 (SLC22A1) in human hepatocytes by competing for ("squelching") SRC-1 coactivator.
Hyrsova L, Smutny T, Carazo A, Moravcik S, Mandikova J, Trejtnar F, Gerbal-Chaloin S, Pavek P
British journal of pharmacology 2016 May;173(10):1703-15
British journal of pharmacology 2016 May;173(10):1703-15
Human eIF3b and eIF3a serve as the nucleation core for the assembly of eIF3 into two interconnected modules: the yeast-like core and the octamer.
Wagner S, Herrmannová A, Šikrová D, Valášek LS
Nucleic acids research 2016 Dec 15;44(22):10772-10788
Nucleic acids research 2016 Dec 15;44(22):10772-10788
An animal model for chorioamnionitis at term.
Dell'Ovo V, Rosenzweig J, Burd I, Merabova N, Darbinian N, Goetzl L
American journal of obstetrics and gynecology 2015 Sep;213(3):387.e1-10
American journal of obstetrics and gynecology 2015 Sep;213(3):387.e1-10
Wnt5a and Wnt11 inhibit the canonical Wnt pathway and promote cardiac progenitor development via the Caspase-dependent degradation of AKT.
Bisson JA, Mills B, Paul Helt JC, Zwaka TP, Cohen ED
Developmental biology 2015 Feb 1;398(1):80-96
Developmental biology 2015 Feb 1;398(1):80-96
DAAM1 and DAAM2 are co-required for myocardial maturation and sarcomere assembly.
Ajima R, Bisson JA, Helt JC, Nakaya MA, Habas R, Tessarollo L, He X, Morrisey EE, Yamaguchi TP, Cohen ED
Developmental biology 2015 Dec 1;408(1):126-39
Developmental biology 2015 Dec 1;408(1):126-39
Intestinal fatty acid binding protein (fabp2) in Atlantic salmon (Salmo salar): Localization and alteration of expression during development of diet induced enteritis.
Venold FF, Penn MH, Thorsen J, Gu J, Kortner TM, Krogdahl A, Bakke AM
Comparative biochemistry and physiology. Part A, Molecular & integrative physiology 2013 Jan;164(1):229-40
Comparative biochemistry and physiology. Part A, Molecular & integrative physiology 2013 Jan;164(1):229-40
Impaired β-adrenergic responsiveness accentuates dysfunctional excitation-contraction coupling in an ovine model of tachypacing-induced heart failure.
Briston SJ, Caldwell JL, Horn MA, Clarke JD, Richards MA, Greensmith DJ, Graham HK, Hall MC, Eisner DA, Dibb KM, Trafford AW
The Journal of physiology 2011 Mar 15;589(Pt 6):1367-82
The Journal of physiology 2011 Mar 15;589(Pt 6):1367-82
An antibody-drug conjugate targeting the endothelin B receptor for the treatment of melanoma.
Asundi J, Reed C, Arca J, McCutcheon K, Ferrando R, Clark S, Luis E, Tien J, Firestein R, Polakis P
Clinical cancer research : an official journal of the American Association for Cancer Research 2011 Mar 1;17(5):965-75
Clinical cancer research : an official journal of the American Association for Cancer Research 2011 Mar 1;17(5):965-75
No comments: Submit comment
Supportive validation
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
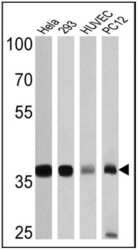
- Western blot analysis of GAPDH was performed by loading 25 µg of Hela (lane 1), 293 (lane 2), HUVEC (lane 3) and PC12 (lane 4) cell lysates onto an SDS polyacrylamide gel. Proteins were transferred to a PVDF membrane and blocked at 4ºC overnight. The membrane was probed with a GAPDH polyclonal antibody (Product # PA1-987) at a dilution of 1:3000 overnight at 4°C, washed in TBST, and probed with an HRP-conjugated secondary antibody for 1 hr at room temperature in the dark. Chemiluminescent detection was performed using Pierce ECL Plus Western Blotting Substrate (Product # 32132). Results show a band at ~36 kDa.

- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
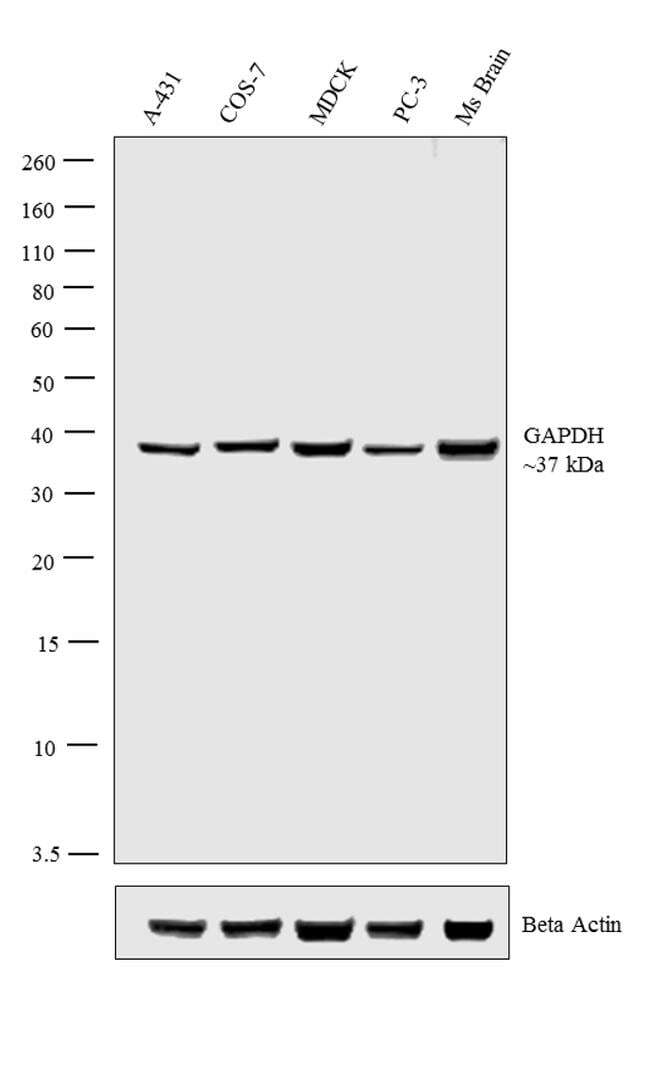
- Western blot analysis was performed on whole cell extracts (30 µg lysate) of A-431 (Lane 1), COS-7 (Lane 2), MDCK (Lane 3), PC-3 (Lane 4) and tissue extract (30 µg lysate) of Ms Brain (Lane 5). The blot was probed with Anti-GAPDH Polyclonal Antibody (Product # PA1-987, 1:1,000 dilution) and detected by chemiluminescence using Goat anti-Rabbit IgG (H+L) Superclonal™ Secondary Antibody, HRP conjugate (Product # A27036, 0.25 µg/mL, 1:4000 dilution). A 37 kDa band corresponding to GAPDH was observed across the cell lines tested. Known quantity of protein samples were electrophoresed using Novex® NuPAGE® 10% Bis-Tris gel (Product # NP0302BOX), XCell SureLock™ Electrophoresis System (Product # EI0002) and Novex® Sharp Pre-Stained Protein Standard (Product # LC5800). Resolved proteins were then transferred onto a nitrocellulose membrane with iBlot® 2 Dry Blotting System (Product # IB21001). The membrane was probed with the relevant primary and secondary Antibody following blocking with 5% skimmed milk. Chemiluminescent detection was performed using Pierce™ ECL Western Blotting Substrate (Product # 32106).
Supportive validation
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL

- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
- Figure 3. The octameric right leg subunits eIF3k and eIF3l impact their mutual expression and are dispensable for the integrity of the rest of eIF3, and the octameric right arm subunit eIF3e stabilizes binding of the right leg subunits (k, l) and eIF3d to the octamer, as well as the octamer attachment to the YLC. Protein levels of all eIF3 subunits and other eIFs upon knock down of either eIF3k, l or e were determined by Western blotting ( A, C, E , respectively). GAPDH was used as a loading control. To assess the integrity of eIF3, the anti-eIF3b co-immunoprecipitation experiments were carried out for each knock-down and the immunoprecipitated eIF3 subunits along with eIF3b were detected by Western blotting ( B, D, F ). The octameric eIF3 subunits are arranged at the left-hand side of each panel and non-octameric subunits are at the right-hand side. The YLC subunits are grouped and highlighted in bold. Quantifications of at least five independent experiments with standard deviations are shown in Supplementary Figure S4 and Table 1 .

- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
- Figure 5. G. intestinalis secreted cysteine proteases have effects on epithelial barrier proteins. A, B, C. Levels of tight junction and adherence junction proteins in Caco-2 whole cell extracts were examined after 24 h of recombinant cysteine proteases treatment (A, 14019; B, 16779; C, 16160), respectively, by Western blot analysis, with GAPDH used as a loading control. Barrier proteins were detected with a mouse or rabbit anti-barrier protein antibody and then incubated with a HR- conjugated mouse or rabbit antibody. D E, F. The localization of occludin in Caco-2 cells visualized by immunofluorescence microscopy. Caco-2 monolayers were incubated with 2.5 ug/ml of 14019 (D), 16779 (E), 16160 (F) in the absence or presence of E64 (10 uM) for 24 h, followed by fixation of PFA. Occludin was detected with a mouse anti-occludin antibody and then incubated with an Alexa Fluor 488-conjugated mouse antibody. Images were taken by a Zeiss Axioplan II Imaging fluorescence microscope and analyzed by ZEN 2.1 software. Bar = 50 um.

- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
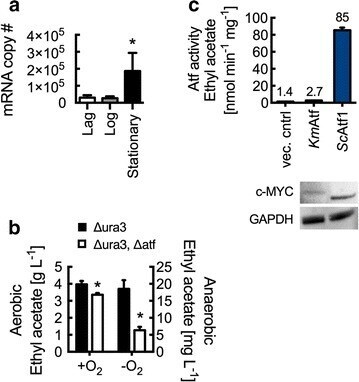
- Fig. 4 Ethyl acetate biosynthesis in K. marxianus by alcohol- O -acetyltransferase. a RT-qPCR analysis of wild-type K. marxianus ATF transcript levels at lag (5 h), log (10 h), and stationary (18 h) phases. mRNA copy number represents total number of transcripts for 5 ng of isolated RNA. b Km ATF disruption reduces ethyl acetate production in both aerobic and anaerobic conditions. Cultures were grown at 37 degC for 10 h (aerobic) and 14 h (anaerobic). c Lysate Atf activity from S. cerevisiae with overexpressed Km ATF and Sc ATF1. Rates are reported per mg of lysate protein. Enzyme expression was confirmed by Western blot analysis using anti-c-MYC and anti-GAPDH antibodies. Bars and error bars represent the arithmetic mean and standard deviation of triplicate biological samples. Statistical significance ( P < 0.05) is indicated by asterisk and was determined using a T test
- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
- Fig. 7 Hemiacetal oxidation activity of K. marxianus alcohol dehydrogenases. Km ADH1-7 were heterologously overexpressed in S. cerevisiae BY 4742. Cell lysate assays showed that Km Adh7 exhibits activity toward synthesis of ethyl acetate from hemiacetal. Enzyme expression was confirmed by Western blot using anti-c-MYC and anti-GAPDH antibodies. Bars and error bars represent the arithmetic mean and standard deviation of triplicate samples

- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
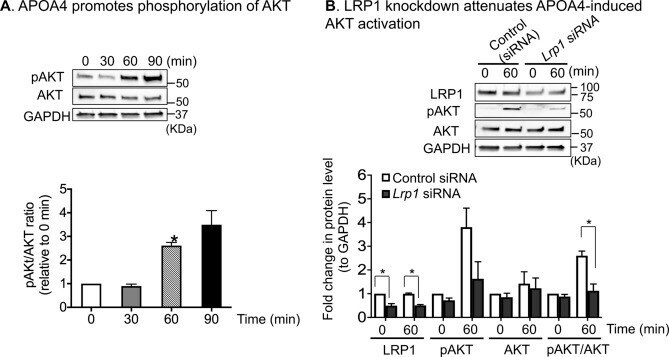
- Figure 5 APOA4 activation of PI3K-AKT pathway is partly LRP1-dependent. ( A ) APOA4 promoted phosphorylation of AKT (pAKT) in a time-dependent manner. Representative western blot images are shown. After transfer of proteins, the membranes were cut into three sections before probing with specific antibodies for western blots. The top section (> 75kD) was used for LRP1 detection, the middle sections (50-75 kD) for pAKT and total AKT detection, and the bottom sections (25-50 kD) for GAPDH detection. This was necessary as stripping and re-probing compromises quantification and running separate gels would not allow for quantification. Bar graphs depicting normalized pAKT and AKT levels to GAPDH. Compared to 0 min, the pAKT/AKT ratio was significantly increased by 2.6-fold at 60 min. ( B ) Representative western blot images showing LRP1, pAKT, AKT, and GAPDH. After pAKT detection, the blot was stripped and used for detecting total AKT. At 60 min post APOA4 treatment, when LRP1 levels were reduced by ~ 50% compared with controls, the pAKT/AKT ratio decreased by ~ 58%, suggesting that APOA4-stimulated increases in pAKT are in part, LRP1-dependent (n = 3/group).
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
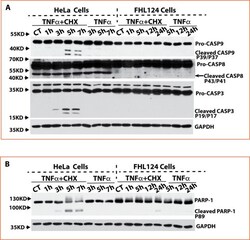
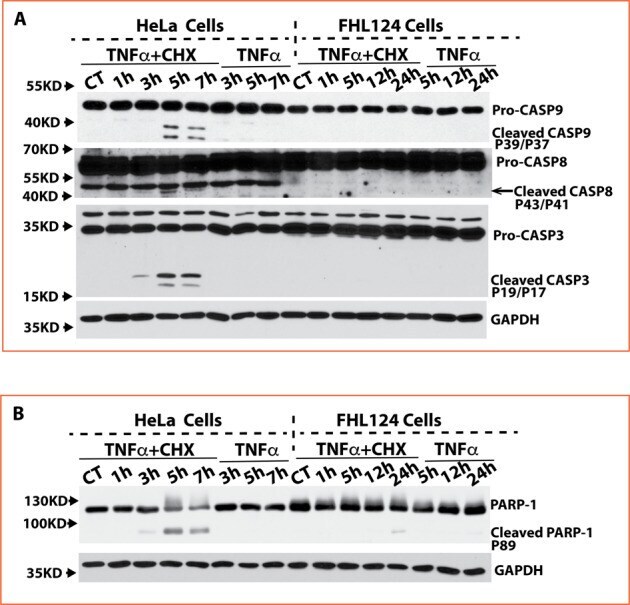
- Fig. 3 FHL124 cells block caspase 8, 9, and 3 activation and PARP-1 cleavage. A Activated caspase-8, caspase-9, and caspase-3 were detected in HeLa cells starting at 3 h and further increased at 5 h and 7 h after TNFalpha and CHX stimulation. No activated caspase-8, caspase-9, and caspase-3 were detectable in FHL124 cells spanning 24 h of treatment with TNFalpha and CHX. A higher pro-caspase-9 quantity (around 2-fold) was seen in HeLa cells than in FHL124 cells, but a higher pro-caspase-3 quantity (around 3-fold) was seen in FHL124 compared to that in HeLa cells. B Cleavage PARP-1 was detected only in HeLa cells but not in FHL124 cells after TNFalpha and CHX stimulation.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
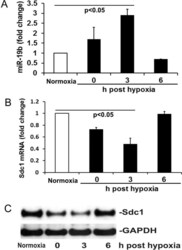
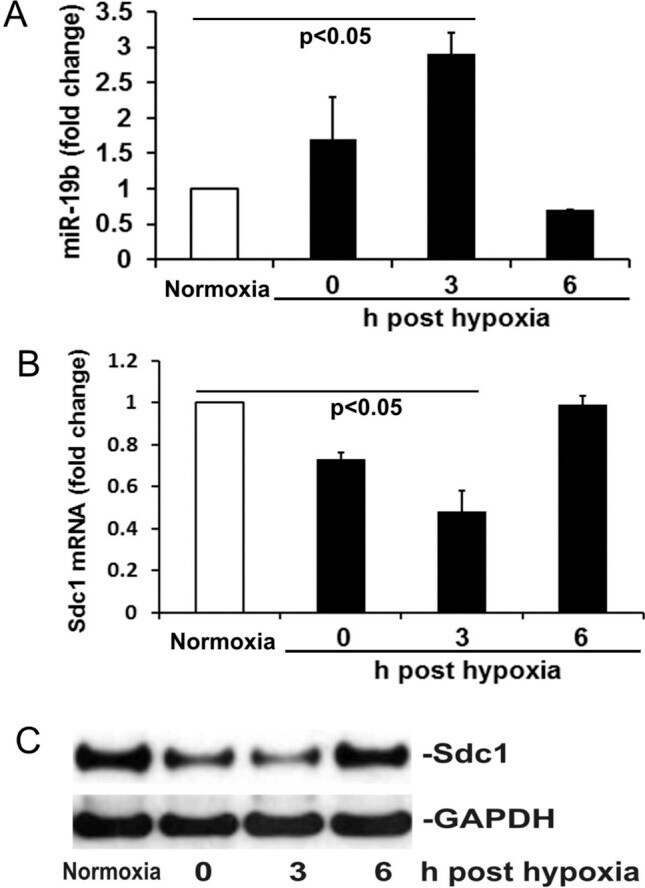
- Figure 2 Hypoxia/reoxygenation (H/R) induces miR-19b expression and Sdc1 inhibition. ( A , B ) The levels of miR-19b and Sdc1 mRNA were measured by RT-qPCR in HLMECs exposed to either normoxia or hypoxia then reoxygenation at 0, 3 and 6 h. N = 4/group; mean +- SD; ANOVA with Bonferroni. ( C ) Expression of Sdc1 and GAPDH was detected by Western blot.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
- Fig. 1 The overproduction of H 2 S in human CRC. A , B H 2 S production measured in human colorectal cancer specimens (* P < 0.05 vs. normal mucosa) and in colorectal cancer cell lines (* P < 0.05 vs. NCM460 cells; n.s. : no significant difference v.s. MCF-10A) by the H 2 S microsensor. (T: tumor tissue; A: aside colorectal tumor tissue; N: normal colorectal tissue). C H 2 S production in different stage of human colorectal cancer specimens measured by the H 2 S microsensor. D Western Blot of CBS protein expression in human colorectal cancer specimens, paired with the adjacent tissue and corresponding normal mucosa tissues. Membranes are re-probed for GAPDH expression to show that similar amounts of protein were loaded in each lane. E Immunohistochemistry of CBS protein expression in human colorectal cancer specimens, paired with the adjacent tissue and corresponding normal mucosa tissues. F Western Blot of CBS protein expression in colon cancer cell lines (vs. NCM460). G H 2 S production is measured in colon cancer cell lines treated by AOAA. H Cell viability of the HCT116 and CT26 treated with AOAA for 24 h. I Cell growth of the HCT116 and CT26 treated with AOAA within 6 days. (* P < 0.05), (n.s. : No significant difference, P value > 0.05)

- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- Fig. 3 The H 2 S scavenging and tumor ferroptosis induced by VZnO in CRC. A The GSH detected by total glutathione assay kit in HCT116 and CT26 after VZnO and ID treatments. B Cell viability by MTT assay in HCT116 and CT26 after VZnO, ID, GSH and Fer-1 treated. C Intracellular ROS levels measured by DCFH-DA assay kit in HCT116 and CT26 with or without VZnO and ID treated. D Colocalization of oxidized lipid and nucleus co-stained by BODIPY C11(green) and DAPI (blue) in HCT116 and CT26 cells. E Representative picture of the ultrastructure of mitochondria in HCT116 and CT26 after VZnO and ID treated. F Average number of mesenchymal cristae per mitochondrion in VZnO-treated cells (statistics of 20 mitochondria) G Representative GPX4, COX2, NOX1 and TFR1 protein expression by Western Blot in HCT116 and CT26 with or without VZnO and ID treated. Membranes were re-probed for GAPDH expression to show that similar amounts of protein were loaded in each lane. (* P < 0.05), (n.s. : No significant difference, P value > 0.05)
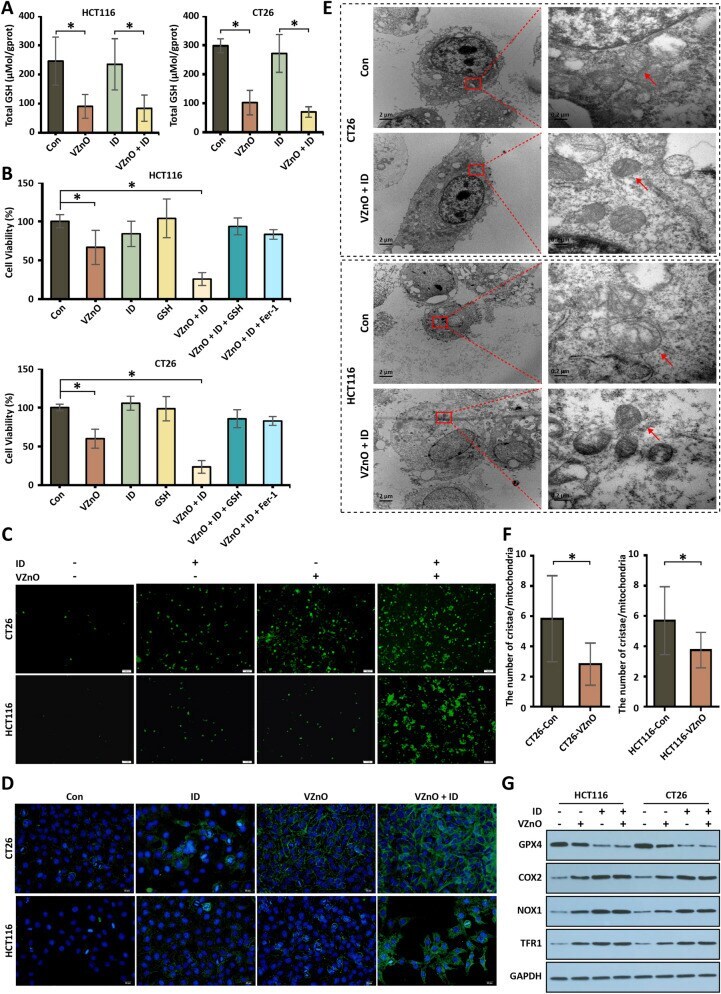
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
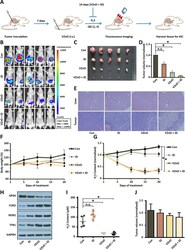
- Fig. 5 Anti-tumor activity of VZnO in orthotopic CRC model. A Experimental timeline of the orthotopic CRC model. B Bioluminescence imaging of orthotopic colon cancer mouse injected with different treatments on day 14. C , D Photograph and volume of the tumors with different treatments on day 14. E Representative H&E stained of liver and tumor tissue on day 14. F Body weights of orthotopic colon cancer model. G The continuously H 2 S production in mouse model. H Representative GPX4, COX2, NOX1 and TFR1 protein expression by Western Blot in tumor on day 14. Membranes were re-probed for GAPDH expression to show that similar amounts of protein were loaded in each lane. I) H 2 S content measured on day 14. J The volume of the tumors in the orthotopic breast model on day 14. (* P < 0.05), (n.s. : No significant difference, P value > 0.05)
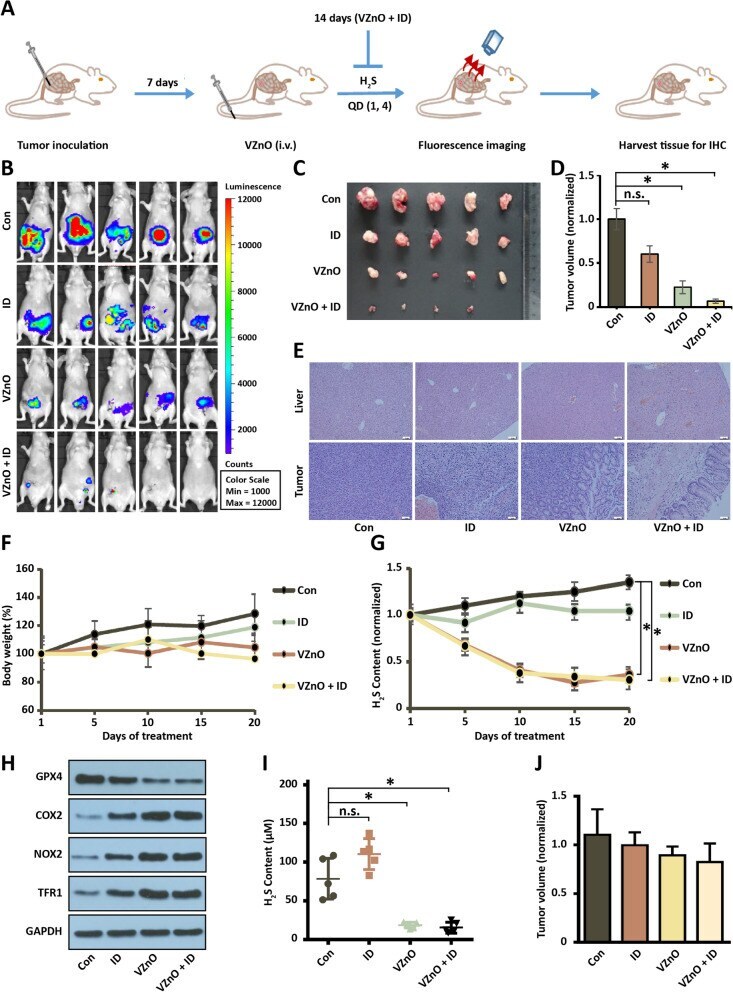
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
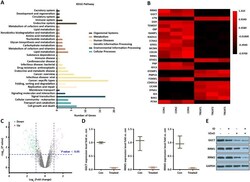
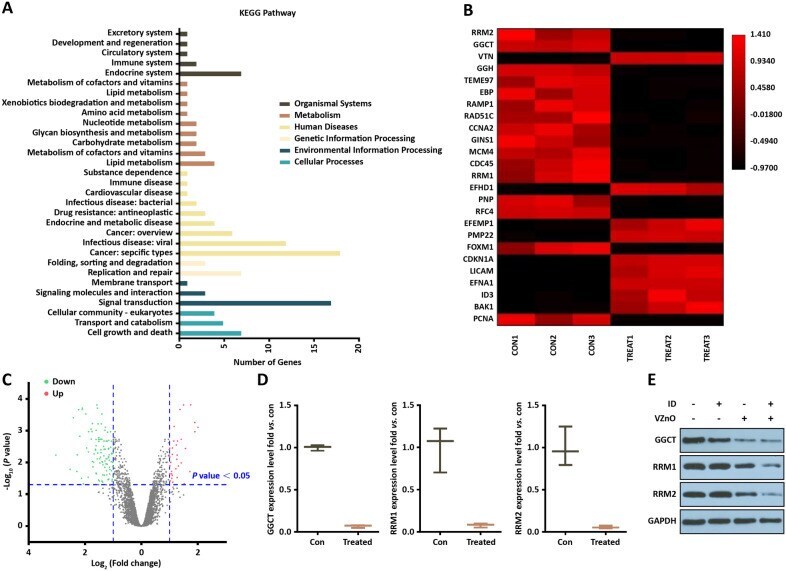
- Fig. 6 Transcriptome analysis in VZnO-treated HCT116. A KEGG pathway analysis based on the RNA-seq results in HCT116. B Representative heatmap of gene expression levels. C Representative scatter plot of 140 significant genes (33 unregulated genes marked in red and 107 downregulated genes marked in green) for Treat vs. Con. D mRNA levels of GGCT, RRM1 and RRM2 by RNA-seq. E Representative Western Blot result of GGCT, RRM1 and RRM2. Membranes were re-probed for GAPDH expression to show that similar amounts of protein were loaded in each lane
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
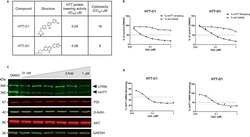
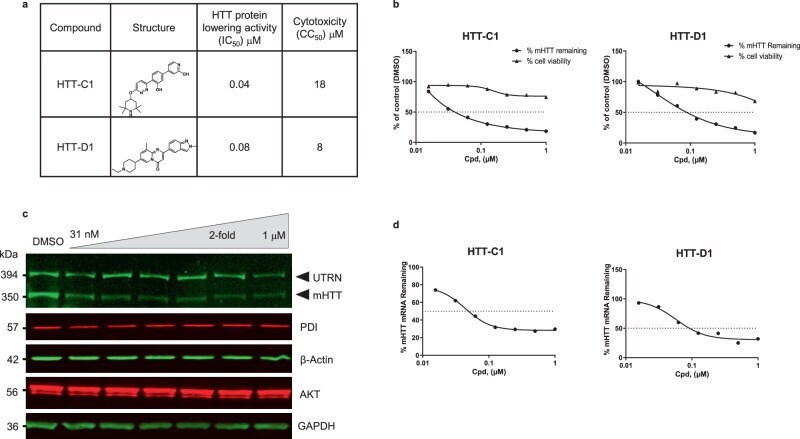
- Fig. 1 Huntingtin (HTT)-lowering activity in vitro. a Chemical structures of HTT-C1 and HTT-D1. b Electrochemiluminescence (ECL) analysis of mutant HTT protein from fibroblasts isolated from a homozygous patient with Huntington's disease (HD) (GM04857) after 96 h of continuous treatment with HTT-C1 and HTT-D1 (0.01-1.0 muM). Representative graphs show percent HTT remaining relative to the dimethyl sulphoxide (DMSO) control. Cell viability assays were performed in parallel. Data represent mean of two ( n = 2) biologically independent samples per data point from one dose-response experiment. c Western blot of HTT protein and housekeeping proteins, oxidoreductase-protein disulphide isomerase (PDI), beta-actin, alpha serine/threonine-protein kinase (AKT) and glyceraldehyde-3-phosphate dehydrogenase (GAPDH) in HD fibroblasts after 96 h of continuous treatment with HTT-C1 (0.015-1.0 muM). Utrophin (UTRN) was also used as a loading control. The western blot data used a representative splicing modifier (tested at multiple concentrations) from a single experiment. Multiple splicing modifiers from the same class were tested and evaluated by western blot analyses. d Reverse transcription-quantitative polymerase chain reaction (RT-qPCR) analysis of HTT mRNA in patient fibroblasts after 24 h of treatment with HTT-C1 and HTT-D1 (0.01-1.0 muM). Representative graphs show percent HTT mRNA remaining relative to DMSO control; normalised to housekeeping gene, TATA-box binding protein. Data repr
- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
- Fig. 1. NRP-1 is incorporated into HIV-1 virion and is mainly expressed in macrophages and DCs. ( A ) The schematic representation of the experimental design used to identify NRP-1 in HIV-1 virions. ( B and C ) Total RNA was extracted from primary monocytes, MDMs, MDDCs, THP-1, PMA-treated THP-1, stimulated or resting CD4 + T cells, established cell lines 293T cells, HeLa cells, and Jurkat cells. NRP-1 transcript levels were measured using qPCR and then normalized to GAPDH levels ( B ). Meanwhile, Western blotting was conducted to assess the level of NRP-1 and GAPDH ( C ). ( D ) The 293T cells exogenously expressing the vector encoding NRP-1-GFP were visualized by microscopy. (Scale bar, 10 mum.) ( E ) MDMs or MDDCs were lysed, and membrane-bound and cytosolic proteins were separated to assess the levels of NRP-1, Caveolin-1, and GAPDH using Western blotting with specific antibodies. All blotting data are representative of three independent experiments.

- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
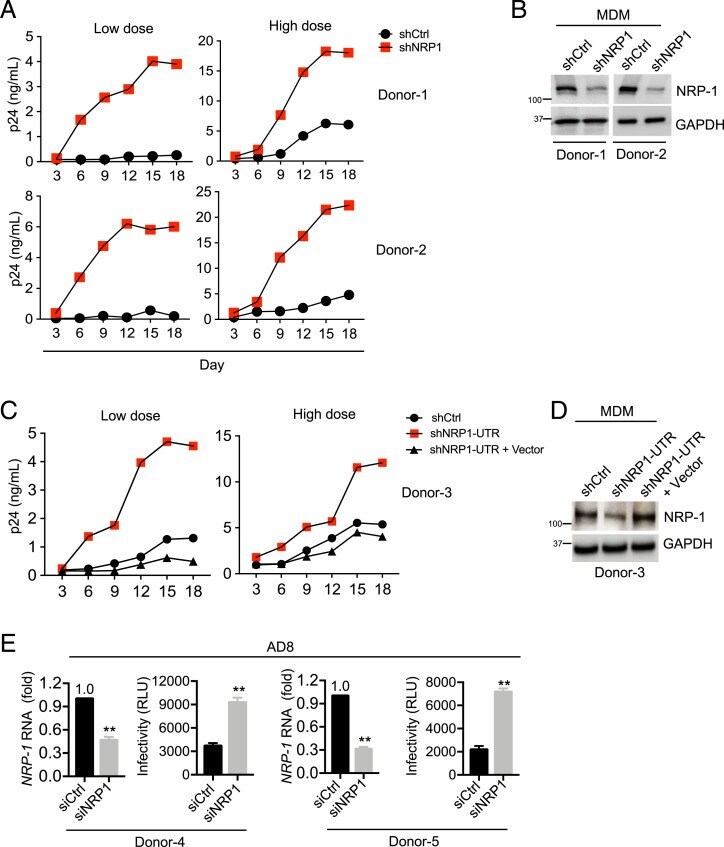
- Fig. 3. NRP-1 inhibits HIV-1 infection in macrophages. ( A and B ) Lentiviral shRNA-transduced MDMs were infected with 10 (low-dose) or 100 (high-dose) ng of HIV-1 AD8 for 18 d. Viral production was measured using p24 ELISA at the indicated time points ( A ). The aliquoted cells were lysed for Western blotting to assess the levels of NRP-1 and GAPDH ( B ). ( C and D ) MDMs were transduced with lentiviral shRNA targeting the 3'-UTR of the NRP-1 or control transcript, and the shNRP-1 -depleted MDMs were then transduced with or without a lentiviral expression vector encoding an untagged NRP-1. Next, the MDMs were infected with 10 (low-dose) or 100 (high-dose) ng HIV-1 AD8 for 18 d, and viral production was measured by p24 ELISA at the indicated time points ( C ). The shRNA-transduced MDMs were lysed for Western blotting to assess the levels of NRP-1 and GAPDH ( D ). ( E ) The MDMs transfected with siRNA targeting NRP-1 or the control were infected with HIV-1 AD8 (multiplicity of infection = 1) and washed twice with PBS to remove the input viruses 6 h after infection. At 3 d after infection, 20 ng of the produced HIV-1 virions in the supernatants were used to infect TZM-bl indicator cells to measure viral progeny infectivity. Total RNA was extracted from the MDMs for qPCR to measure NRP-1 transcript levels normalized to GAPDH levels. ** P < 0.01.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
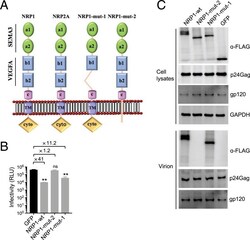
- Experimental details
- Fig. 4. NRP-1 is incorporated into virions to restrict HIV-1 infection. ( A ) The schematic representation of NRP-1 domains. The full-length NRP-1 possesses a ligand-binding ectodomain consisting of a1/a2 and b1/b2 domains, followed by a membrane-proximal c domain involved in oligomerization. The remainder of the protein comprises a short membrane-spanning region and cytoplasmic tail (cyto). ( B and C ) NRP-1 is incorporated into viral particles to inhibit infectivity. The 293T cells were cotransfected with constructs encoding FLAG-tagged NRP-1 or its mutants and GFP, along with NL4-3 proviral vectors. At 2 d after cotransfection, 100 ng of the produced HIV-1 virions in the supernatants were used to infect TZM-bl indicator cells to measure viral progeny infectivity ( B ), and the cells were lysed for Western blotting to assess the levels of NRP-1, its mutants, Gag, gp120, and GAPDH with their specific antibodies. The produced virion particles in the cell supernatants were enriched by sucrose cushion ultracentrifugation and subsequently isolated using anti-CD44 microbeads with a magnetic-based method. The purified virion was lysed for Western blotting to assess the NRP-1 or its mutants, CA (p24), and gp120 levels ( C ). All blotting data are representative of three independent experiments. ** P < 0.01; ns, not significant.

- Submitted by
- Invitrogen Antibodies (provider)
- Main image
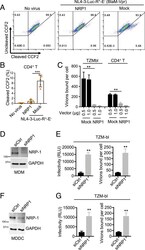
- Experimental details
- Fig. 5. NRP-1 inhibits HIV-1 virion attachment to target cells. ( A and B ) The BlaM-Vpr-based viral entry assay using a VSV-G-pseudotyped HIV-1. The 293T cells were cotransfected with a construct encoding FLAG-tagged NRP-1 or a mock expression construct with or without pNL4-3.GFP.R + E - reporter vectors (VSV-G) combined with an expression construct encoding BlaM-Vpr. Two days after transfection, 50 ng of the produced virions in the supernatants were used to infect stimulated CD4 + T cells. The fluorescence-activated cell sorter dot plots are representative of five independent experiments ( A ). The data of five independent experiments are plotted as mean +- SEM ( B ). *** P < 0.001. ( C ) The HIV-1 attachment assay. The 293T cells were cotransfected with a construct encoding FLAG-tagged NRP-1 or a mock expression construct at different doses, along with HIV-1 NL4-3 proviral vectors. Two days after transfection, TZM-bl reporter cells or stimulated CD4 + T target cells were spinoculated with 500 ng of the produced HIV-1 particles at 25 degC for 2 h and washed with PBS twice to remove the input virus after spinoculation. The cells were further lysed for the p24 ELISA to quantify the bound viral particles. ( D - G ) MDMs ( D and E ) or MDDCs ( F and G ) were cotransfected with siRNA targeting NRP-1 or the control along with HIV-1 AD8 proviral vectors. At 3 d after cotransfection, the cells were lysed for Western blotting to assess the levels of NRP-1 and GAPDH, and 20 ng of the
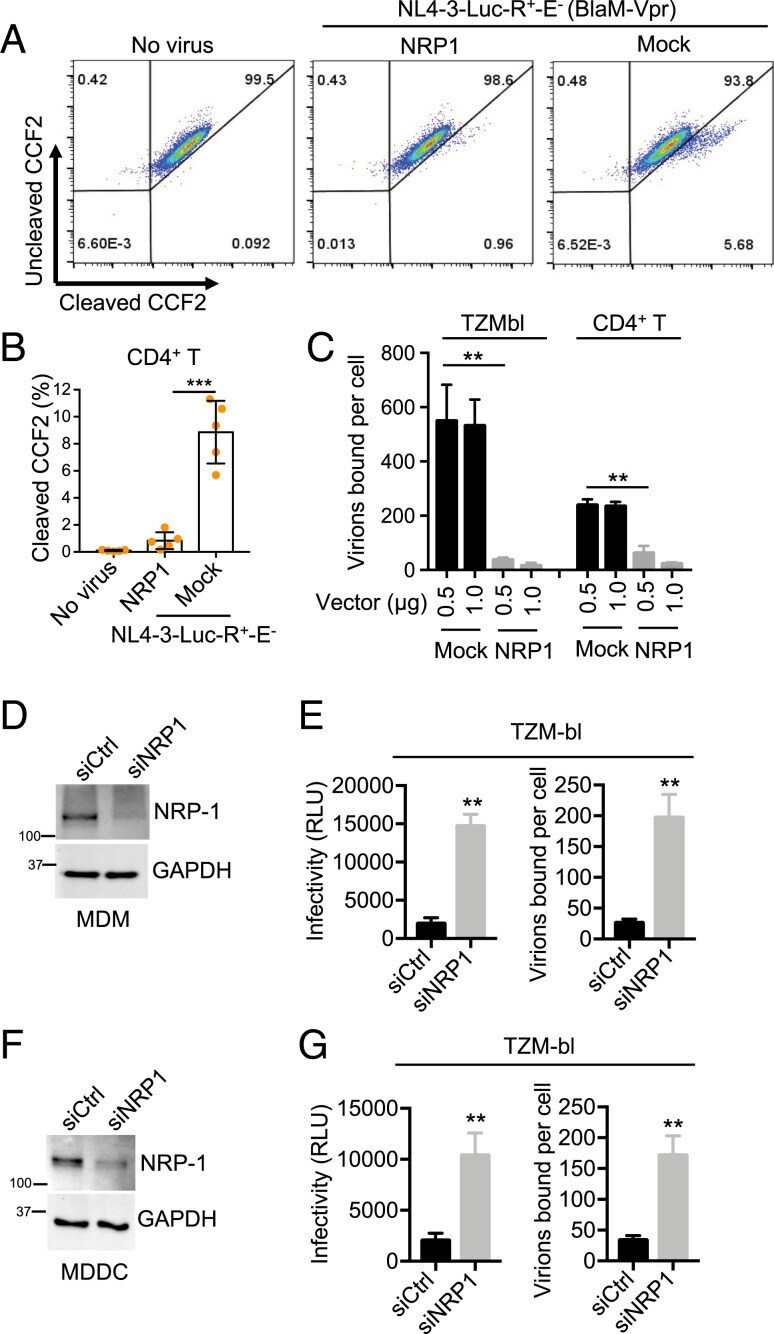
- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
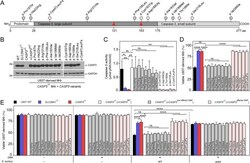
- 10 Figure Kv7.2 and Kv7.3 subunits expression in superficial and deep layers in the motor cortex of developing KCNQ2 WT/WT and KCNQ2 WT/T274M mice A , western blots of Kv7.2 and glyceraldehyde 3-phosphate dehydrogenase (GAPDH) at postnatal day (PND) 7, 21 and 30 in superficial layers ( Aa ) and deep layers ( Ab ). B , boxplots quantifying the Kv7.2 subunit relative to the housekeeping protein GAPDH in superficial ( Ba ) and deep layers ( Bb ) of the motor cortex from KCNQ2 WT/WT (black dots) and KCNQ WT/T274M mice (blue dots and purple dots) aged 7, 21 and 30 days. Each dot corresponds to quantification in one mouse. C , western blots of Kv7.3 and of GAPDH in both superficial ( Ca ) and deep layers ( Cb ). D , boxplots quantifying the Kv7.3 subunit relative to GAPDH in superficial ( Da ) and deep layers ( Db ) of the motor cortex from KCNQ2 WT/WT (black dots) and KCNQ WT/T274M mice (blue dots and purple dots) aged 7, 21 and 30 days. Statistical analysis for the effect of the variant across age was by two-way ANOVA. The two factors were the genotype and the age. For Ba , interaction P = 0.32, genotype P = 0.38 and age P = 0.005. For Bb , interaction P = 0.83, genotype P = 0.21 and age P = 0.0001. For Da , interaction P = 0.17, genotype P = 0.7 and age P = 0.23. For Db , interaction P = 0.06, genotype P = 0.06 and age P = 0.005. Statistical analysis for the comparisons of wild-type with knock-in (KI) mice at PND 7, 21 or 30 was by Student's unpaired t test and the Mann-Whitney

- Submitted by
- Invitrogen Antibodies (provider)
- Main image
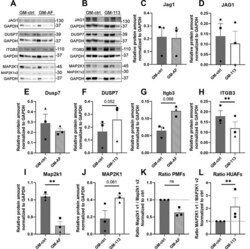
- Experimental details
- Protein expression in murine and human primary fibroblasts. ( A ) Western blots for JAG1, DUSP7, ITGB3 and MAP2K1, each with GAPDH, of three independent replicates (each band represents a pool of three technical triplicates) in PMFs. ( B ) Western blots for JAG1, DUSP7, ITGB3 and MAP2K1, with GAPDH of three independent replicates (each band represents a pool of three technical triplicates) in HUAFs. ( C - L ) Quantifications of protein band intensities, normalized to GAPDH. Data are represented as mean +- SEM. * * P < 0.01, compared with GM-ctrl.

- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- The protein level of RIP3 is elevated in hypertrophic hearts. (a) Representative Western blots and quantitation of RIP3 expression in heart tissue from patients with dilated cardiomyopathy (DCM) and normal controls. (b) (Up) Western blots of RIP3, ANP, beta -MHC, and GAPDH and (down) relative quantitation of RIP3 expression in heart tissue from rats after sham or AB operation. (c) (Up) Western blots of RIP3, ANP, beta -MHC, and GAPDH and (down) relative quantitation of RIP3 in NRCMs stimulated with Ang-II (1 mu M) or PBS. (d) (up) Western blots of RIP3, ANP, beta -MHC, and GAPDH and (down) relative quantitation of RIP3 in NRCMs stimulated with PE (50 mu M) or PBS. kDa: kilo-Dalton; * P < 0.05, ** P < 0.01, and *** P < 0.001.

- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
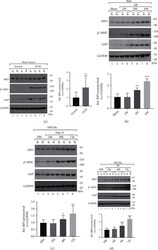
- FIG 1 IFN-alpha treatment induces OTOF in macrophages. (A) OTOF was induced in the PBMCs of untreated patients with HIV infection. Volcano plot of genes differentially expressed in PBMCs isolated from five independent healthy donors and five independent untreated patients with HIV-1 infection. The log 2 fold change difference is presented on the x axis, and the negative log of the false-discovery rate (FDR) is presented on the y axis. Each point represents one gene, which had detectable expression in both groups (five untreated patients with HIV-1 infection versus five healthy donors). The significantly differentially expressed genes are plotted in red with upregulated genes on the right side and downregulated genes on the left side. The nonsignificantly differentially expressed genes are shown as black points. (B) Relative expression of OTOF in the PBMCs of healthy donors ( n = 16), patients with HIV-1 who did not receive ART ( n = 26), and patients with HIV-1 who received ART ( n = 20; viral loads of

- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
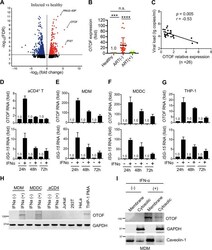
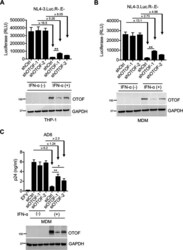
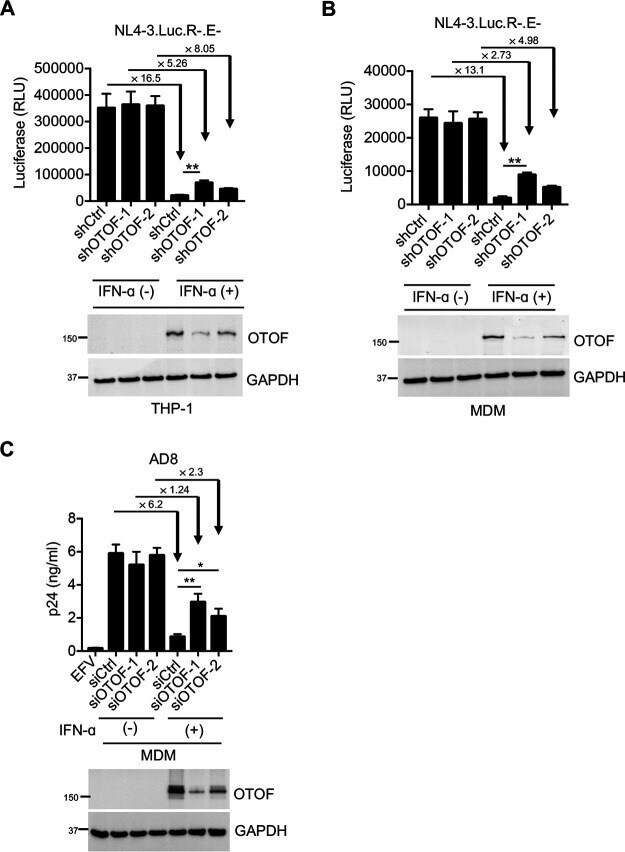
- FIG 2 IFN-alpha induces OTOF to restrict HIV-1 infection in macrophages. (A and B) Lentiviral shRNA-transduced THP-1 cells (A) and MDMs (B) were pretreated with IFN-alpha (1,000 U/mL). At 24 h after treatment, cells were washed using culture medium and subsequently infected with 100 ng of HIV-1 NL4-3.Luc.R-.E- (VSV-G). At 24 h after infection, cells were lysed to measure luciferase reporter activity and for Western blotting to assess the levels of OTOF and GAPDH. RLU, relative luminescence units. (C) MDMs were transfected with siRNA against OTOF or control siRNA, and cells were treated with IFN-alpha (1,000 U/mL) 6 h after transfection. At 24 h after treatment, cells were washed and further infected with 50 ng of replication-competent HIV-1 AD8 . At 72 h after infection, viral load in culture supernatants was measured via p24 ELISA; cells were lysed for Western blotting to assess the levels of OTOF and GAPDH. *, P < 0.05; **, P < 0.01 (two-tailed, unpaired Student's t test). EFV, efavirenz. Data are plotted as the means +- standard deviations for three triplicates and representative of three independent experiments. All Western blot data are representative of three independent experiments. Numbers at left of blots are molecular masses in kilodaltons.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
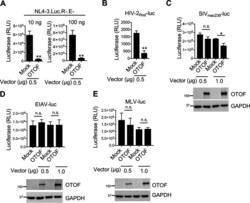
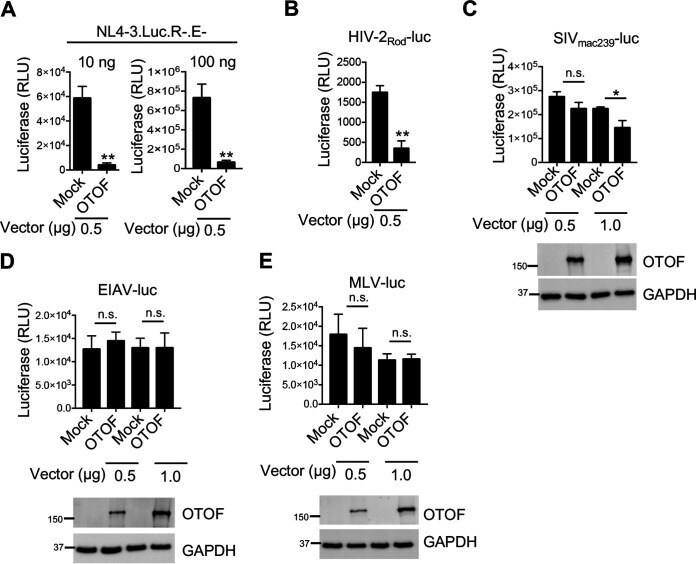
- FIG 3 OTOF suppresses HIV-1 and HIV-2 infection and slightly suppresses SIV infection but does not suppress EIAV and MLV infection. (A) 293T cells were transfected with a construct encoding FLAG-tagged OTOF or a mock expression construct at the indicated doses. At 24 h after transfection, cells were infected with 10 or 100 ng of HIV-1 NL4-3.Luc.R-.E- (VSV-G). At 24 h after infection, cells were lysed to measure luciferase reporter activity. (B to E) 293T cells were transfected with a construct encoding FLAG-tagged OTOF or a mock expression construct at the indicated doses. At 24 h after transfection, cells were infected with VSV-G pseudotyped luciferase reporter viruses: HIV-2 Rod -luc (B), SIV mac239- luc (C), EIAV-luc (D), or MLV-luc (E). At 24 h after infection, cells were lysed to measure luciferase reporter activity and for Western blotting to assess the levels of OTOF and GAPDH using specific antibodies. **, P < 0.01; *, P < 0.015; n.s., not significant (two-tailed, unpaired Student's t test). Data are plotted as the means +- standard errors of the means for three independent experiments. All Western blot data are representative of three independent experiments. Numbers at left of blots are molecular masses in kilodaltons.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
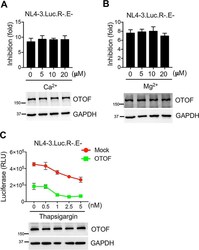
- FIG 4 Antiviral activity of OTOF is independent of Ca 2+ . (A and B) 293T cells were transfected with a construct encoding FLAG-tagged OTOF or a mock expression construct. At 24 h after transfection, cells were infected with 100 ng of HIV-1 NL4-3.Luc.R-.E- (VSV-G) in the presence or absence of Ca 2+ (A) or Mg 2+ (B) at the indicated doses. At 24 h after infection, cells were lysed to measure luciferase reporter activity and for Western blotting to assess the levels of OTOF and GAPDH using specific antibodies. The inhibition of viral infection was accordingly calculated for OTOF and mock. Data are plotted as the means +- standard errors of the means from three independent experiments. (C) 293T cells were transfected with a construct encoding FLAG-tagged OTOF or a mock expression construct. At 24 h after transfection, cells were infected with 100 ng of HIV-1 NL4-3.Luc.R-.E- (VSV-G) in the presence or absence of thapsigargin at the indicated doses. At 24 h after infection, cells were lysed to measure luciferase reporter activity and for Western blotting to assess the levels of OTOF and GAPDH using specific antibodies. Data are plotted as the means +- standard deviations from three triplicates. All Western blot data are representative of three independent experiments. Numbers at left of blots are molecular masses in kilodaltons.

- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
- FIG 1 Caspase-3 is required for deoxyadenosine-induced killing of macrophages. (A) U937-derived macrophages (MPhi) were treated with (+) or without (-) dAdo, and cell lysates were analyzed for caspase-3 activity using a colorimetric assay. As controls, lysates were treated with the caspase-3 inhibitor Ac-DEVD-CHO (+ Inhib.). (B) Survival of MPhi exposed to dAdo and increasing concentrations of Z-DEVD-FMK (0 to 5 muM), an inhibitor of caspase-3. (C) Diagram illustrating the position of CASP3 on chromosome 4 and exons 1 to 8 of CASP3 mRNA. Sequencing results for mutated exon 5 alleles (red box) cloned from CASP3 -/- cells are shown. (D) Immunoblotting of lysates from wild-type (WT) MPhi and their CASP3 -/- and complemented CASP3 -/- variants (+ CASP3 WT ) with caspase-3 and GAPDH-specific antibodies (alpha-CASP3 and alpha-GAPDH, respectively). GAPDH was used as a loading control. Numbers to the left of blots indicate the migration of molecular weight markers in kilodaltons. (E and F) Survival of MPhi (black bars) and their SLC29A1 -/- (blue bars), CASP3 -/- (red bars), and complemented CASP3 -/- variants (+ CASP3 WT , white bars) after treatment with dAdo (E) or after treatment with culture medium (RPMI 1640) that had been conditioned by incubation with either wild-type S. aureus Newman or its adsA mutant in the presence of host DNA, as indicated by + and - signs (F). All samples received adenosine deaminase inhibitor (50 muM dCF). Data are the mean (+- standard deviation [SD])

- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
- FIG 3 Caspase-3 activity suppresses macrophage infiltration into staphylococcal abscesses. (A to P) Immunohistochemical analysis of renal tissues isolated 5 days after intravenous injection of 10 7 CFU of wild-type S. aureus Newman or its adsA mutant into C57BL/6 CASP3 fl/fl or CASP3 fl/fl Tie2-Cre + mice. Thin sections were stained with hematoxylin and eosin (H&E) (A to D) or examined by immunohistochemistry with anti-Ly-6G antibodies (neutrophils) (E to H) or anti-F4/80 antibodies (macrophages) (I to L). (M to P) Magnifications of boxed area from panels I to L. Macrophages and neutrophils stain brown. Green arrows point to replicating S. aureus cells surrounded by a fibrin capsule. Black bars depict a length of 100 mum. Representative images are shown. (Q) Determination of macrophage-infiltrated areas of renal abscesses of infected C57BL/6 CASP3 fl/fl (black circles) or CASP3 fl/fl Tie2-Cre + mice (open circles) by immunohistochemistry with anti-F4/80 antibodies. Macrophage-infiltrated areas were determined by calculating the total and macrophage-free (anti-F4/80-negative) abscess areas. Multiple abscesses ( n = 25 to 35) from a cohort of 3 to 4 animals per group were analyzed. Horizontal blue bars represent mean values in each cohort. (R) Immunoblotting of lysates from bone marrow-derived macrophages (BMDM). Lysates of BMDM derived from female ( symbol) or male ( symbol) C57BL/6 CASP3 fl/fl or CASP3 fl/fl Tie2-Cre + mice were probed with caspase-3 and GAPDH-specific antibo

- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- FIG 4 Single nucleotide polymorphisms (SNPs) in CASP3 protect human macrophages from S. aureus -derived deoxyadenosine. (A) Caspase-3 protein lollipop plot highlighting amino acid substitutions investigated in this study. Associated SNP identifiers (IDs) are provided in Table S1 . (B) Immunoblotting of lysates from wild-type (WT) U937-derived macrophages (MPhi) and their SLC29A1 -/- , CASP3 -/- , and complemented CASP3 -/- variants (WT and 12 different alleles indicated according to their amino acid substitution in caspase-3) using caspase-3 and GAPDH-specific antibodies (alpha-CASP3 and alpha-GAPDH, respectively). GAPDH was used as a loading control. Numbers to the left of blots indicate the migration of molecular weight markers in kilodaltons. (C) Caspase-3 activity in cell lysates of dAdo-exposed WT MPhi (black bars) and their SLC29A1 -/- (blue bars), CASP3 -/- (red bars), and CASP3 -/- variants complemented with WT (open bars) and various alleles of CASP3 (gray or pink bars). Caspase-3 activity was measured using a colorimetric assay. (D and E) Survival of WT MPhi (black bars) and their SLC29A1 -/- (blue bars), CASP3 -/- (red bars), and CASP3 -/- variants complemented with WT (open bars) and various alleles of CASP3 (gray or pink bars) after treatment with dAdo (D) or after treatment with culture medium (RPMI) that had been conditioned by incubation with either wild-type S. aureus Newman or its adsA mutant in the presence of host DNA, as indicated with + and - signs (E).

- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
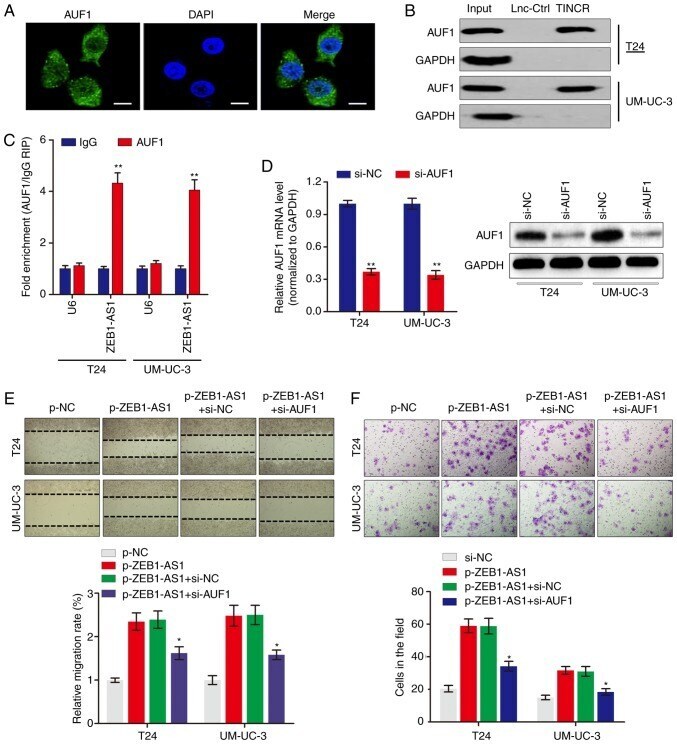
- Figure 3 ZEB1-AS1 interacts with AUF1 to serve key roles in BCa. (A) Immunofluorescence analysis of AUF1 protein in UM-UC-3 cells. Scale bars, 10 u m. (B) The interaction between ZEB1-AS1 and AUF1 was confirmed by RNA pulldown assays and western blot analysis. GAPDH served as negative control. (C) RNA immunoprecipitation assays were performed using anti-AUF1 and control IgG antibodies, followed by RT-qPCR to examine the enrichment of ZEB1-AS1 and U6. U6 served as negative control. ** P