Explore
Explore Validate
Validate Learn
Learn14-9999-80
antibody from Invitrogen Antibodies
Targeting: CXCR4
CD184, D2S201E, fusin, HM89, HSY3RR, LESTR, NPY3R, NPYR, NPYY3R
 Flow cytometry
Flow cytometryAntibody data
- Antibody Data
- Antigen structure
- References [26]
- Comments [0]
- Validations
- Flow cytometry [1]
- Other assay [19]
Submit
Validation data
Reference
Comment
Report error
- Product number
- 14-9999-80 - Provider product page

- Provider
- Invitrogen Antibodies
- Product name
- CD184 (CXCR4) Monoclonal Antibody (12G5), eBioscience™
- Antibody type
- Monoclonal
- Antigen
- Other
- Description
- Description: The 12G5 monoclonal antibody reacts with human CD184 (CXCR4), also termed Fusin, LESTR, or HUMSTR. CD184 is a member of the G-protein-coupled chemokine receptor family with seven membrane-spanning domains, and functions as a coreceptor for X4 HIV-1 entry into CD4+ cells. CD184 is expressed predominantly on naive T cell subsets of peripheral blood and is rapidly upregulated by PHA and IL-2 stimulation. The 12G5 antibody shows partial inhibition of chemotaxis and calcium influx induced by SDF-1 (the natural ligand of CD184), blocks CD4-independent HIV-2 infection, and blocks CD4-dependent infection by some T-tropic HIV-1 isolates.
- Antibody clone number
- 12G5
- Concentration
- 0.5 mg/mL
Submitted references Parallel functional assessment of m(6)A sites in human endodermal differentiation with base editor screens.
Cotargeting of miR-126-3p and miR-221-3p inhibits PIK3R2 and PTEN, reducing lung cancer growth and metastasis by blocking AKT and CXCR4 signalling.
Longitudinal single-cell profiling reveals molecular heterogeneity and tumor-immune evolution in refractory mantle cell lymphoma.
Senescent Tumor Cells Build a Cytokine Shield in Colorectal Cancer.
The CXCR4 antagonist plerixafor (AMD3100) promotes proliferation of Ewing sarcoma cell lines in vitro and activates receptor tyrosine kinase signaling.
Optimized RNP transfection for highly efficient CRISPR/Cas9-mediated gene knockout in primary T cells.
Modulatory effects of trophoblast-secreted CXCL12 on the migration and invasion of human first-trimester decidual epithelial cells are mediated by CXCR4 rather than CXCR7.
A phase 1 study of the CXCR4 antagonist plerixafor in combination with high-dose cytarabine and etoposide in children with relapsed or refractory acute leukemias or myelodysplastic syndrome: A Pediatric Oncology Experimental Therapeutics Investigators' Consortium study (POE 10-03).
In Vitro Modeling of Alcohol-Induced Liver Injury Using Human-Induced Pluripotent Stem Cells.
Chemokine axes in breast cancer: factors of the tumor microenvironment reshape the CCR7-driven metastatic spread of luminal-A breast tumors.
ST8SIA4-Dependent Polysialylation is Part of a Developmental Program Required for Germ Layer Formation from Human Pluripotent Stem Cells.
The dual action of poly(ADP-ribose) polymerase -1 (PARP-1) inhibition in HIV-1 infection: HIV-1 LTR inhibition and diminution in Rho GTPase activity.
AEG-1 promotes anoikis resistance and orientation chemotaxis in hepatocellular carcinoma cells.
Antibody epitopes on g protein-coupled receptors mapped with genetically encoded photoactivatable cross-linkers.
Prevailing role of contact guidance in intrastromal T-cell trapping in human pancreatic cancer.
LDL cholesterol modulates human CD34+ HSPCs through effects on proliferation and the IL-17 G-CSF axis.
Dynamic chemotherapy-induced upregulation of CXCR4 expression: a mechanism of therapeutic resistance in pediatric AML.
Flagellin induces myeloid-derived suppressor cells: implications for Pseudomonas aeruginosa infection in cystic fibrosis lung disease.
Human cytomegalovirus infection inhibits CXCL12- mediated migration and invasion of human extravillous cytotrophoblasts.
Osteopontin enhances the expression and activity of MMP-2 via the SDF-1/CXCR4 axis in hepatocellular carcinoma cell lines.
CD137 ligand signaling induces human monocyte to dendritic cell differentiation.
Erythromycin-induced CXCR4 expression on microvascular endothelial cells.
Human mast cell activation with virus-associated stimuli leads to the selective chemotaxis of natural killer cells by a CXCL8-dependent mechanism.
The HIV coreceptors CXCR4 and CCR5 are differentially expressed and regulated on human T lymphocytes.
The HIV coreceptors CXCR4 and CCR5 are differentially expressed and regulated on human T lymphocytes.
Inhibition of human immunodeficiency virus fusion by a monoclonal antibody to a coreceptor (CXCR4) is both cell type and virus strain dependent.
Cheng W, Liu F, Ren Z, Chen W, Chen Y, Liu T, Ma Y, Cao N, Wang J
Nature communications 2022 Jan 25;13(1):478
Nature communications 2022 Jan 25;13(1):478
Cotargeting of miR-126-3p and miR-221-3p inhibits PIK3R2 and PTEN, reducing lung cancer growth and metastasis by blocking AKT and CXCR4 signalling.
Di Paolo D, Pontis F, Moro M, Centonze G, Bertolini G, Milione M, Mensah M, Segale M, Petraroia I, Borzi C, Suatoni P, Brignole C, Perri P, Ponzoni M, Pastorino U, Sozzi G, Fortunato O
Molecular oncology 2021 Nov;15(11):2969-2988
Molecular oncology 2021 Nov;15(11):2969-2988
Longitudinal single-cell profiling reveals molecular heterogeneity and tumor-immune evolution in refractory mantle cell lymphoma.
Zhang S, Jiang VC, Han G, Hao D, Lian J, Liu Y, Zhang R, McIntosh J, Wang R, Dang M, Dai E, Wang Y, Santos D, Badillo M, Leeming A, Chen Z, Hartig K, Bigcal J, Zhou J, Kanagal-Shamanna R, Ok CY, Lee H, Steiner RE, Zhang J, Song X, Nair R, Ahmed S, Rodriquez A, Thirumurthi S, Jain P, Wagner-Bartak N, Hill H, Nomie K, Flowers C, Futreal A, Wang L, Wang M
Nature communications 2021 May 17;12(1):2877
Nature communications 2021 May 17;12(1):2877
Senescent Tumor Cells Build a Cytokine Shield in Colorectal Cancer.
Choi YW, Kim YH, Oh SY, Suh KW, Kim YS, Lee GY, Yoon JE, Park SS, Lee YK, Park YJ, Kim HS, Park SH, Kim JH, Park TJ
Advanced science (Weinheim, Baden-Wurttemberg, Germany) 2021 Feb;8(4):2002497
Advanced science (Weinheim, Baden-Wurttemberg, Germany) 2021 Feb;8(4):2002497
The CXCR4 antagonist plerixafor (AMD3100) promotes proliferation of Ewing sarcoma cell lines in vitro and activates receptor tyrosine kinase signaling.
Berning P, Schaefer C, Clemens D, Korsching E, Dirksen U, Potratz J
Cell communication and signaling : CCS 2018 May 18;16(1):21
Cell communication and signaling : CCS 2018 May 18;16(1):21
Optimized RNP transfection for highly efficient CRISPR/Cas9-mediated gene knockout in primary T cells.
Seki A, Rutz S
The Journal of experimental medicine 2018 Mar 5;215(3):985-997
The Journal of experimental medicine 2018 Mar 5;215(3):985-997
Modulatory effects of trophoblast-secreted CXCL12 on the migration and invasion of human first-trimester decidual epithelial cells are mediated by CXCR4 rather than CXCR7.
Zheng J, Wang H, Zhou W
Reproductive biology and endocrinology : RB&E 2018 Mar 2;16(1):17
Reproductive biology and endocrinology : RB&E 2018 Mar 2;16(1):17
A phase 1 study of the CXCR4 antagonist plerixafor in combination with high-dose cytarabine and etoposide in children with relapsed or refractory acute leukemias or myelodysplastic syndrome: A Pediatric Oncology Experimental Therapeutics Investigators' Consortium study (POE 10-03).
Cooper TM, Sison EAR, Baker SD, Li L, Ahmed A, Trippett T, Gore L, Macy ME, Narendran A, August K, Absalon MJ, Boklan J, Pollard J, Magoon D, Brown PA
Pediatric blood & cancer 2017 Aug;64(8)
Pediatric blood & cancer 2017 Aug;64(8)
In Vitro Modeling of Alcohol-Induced Liver Injury Using Human-Induced Pluripotent Stem Cells.
Tian L, Prasad N, Jang YY
Methods in molecular biology (Clifton, N.J.) 2016;1353:271-83
Methods in molecular biology (Clifton, N.J.) 2016;1353:271-83
Chemokine axes in breast cancer: factors of the tumor microenvironment reshape the CCR7-driven metastatic spread of luminal-A breast tumors.
Weitzenfeld P, Kossover O, Körner C, Meshel T, Wiemann S, Seliktar D, Legler DF, Ben-Baruch A
Journal of leukocyte biology 2016 Jun;99(6):1009-25
Journal of leukocyte biology 2016 Jun;99(6):1009-25
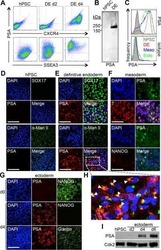
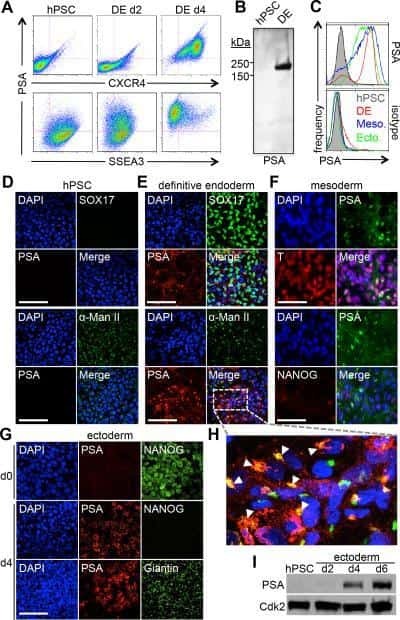
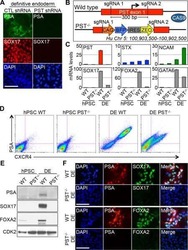
ST8SIA4-Dependent Polysialylation is Part of a Developmental Program Required for Germ Layer Formation from Human Pluripotent Stem Cells.
Berger RP, Sun YH, Kulik M, Lee JK, Nairn AV, Moremen KW, Pierce M, Dalton S
Stem cells (Dayton, Ohio) 2016 Jul;34(7):1742-52
Stem cells (Dayton, Ohio) 2016 Jul;34(7):1742-52
The dual action of poly(ADP-ribose) polymerase -1 (PARP-1) inhibition in HIV-1 infection: HIV-1 LTR inhibition and diminution in Rho GTPase activity.
Rom S, Reichenbach NL, Dykstra H, Persidsky Y
Frontiers in microbiology 2015;6:878
Frontiers in microbiology 2015;6:878
AEG-1 promotes anoikis resistance and orientation chemotaxis in hepatocellular carcinoma cells.
Zhou Z, Deng H, Yan W, Luo M, Tu W, Xia Y, He J, Han P, Fu Y, Tian D
PloS one 2014;9(6):e100372
PloS one 2014;9(6):e100372
Antibody epitopes on g protein-coupled receptors mapped with genetically encoded photoactivatable cross-linkers.
Ray-Saha S, Huber T, Sakmar TP
Biochemistry 2014 Mar 4;53(8):1302-10
Biochemistry 2014 Mar 4;53(8):1302-10
Prevailing role of contact guidance in intrastromal T-cell trapping in human pancreatic cancer.
Hartmann N, Giese NA, Giese T, Poschke I, Offringa R, Werner J, Ryschich E
Clinical cancer research : an official journal of the American Association for Cancer Research 2014 Jul 1;20(13):3422-33
Clinical cancer research : an official journal of the American Association for Cancer Research 2014 Jul 1;20(13):3422-33
LDL cholesterol modulates human CD34+ HSPCs through effects on proliferation and the IL-17 G-CSF axis.
Cimato TR, Palka BA, Lang JK, Young RF
PloS one 2013;8(8):e73861
PloS one 2013;8(8):e73861
Dynamic chemotherapy-induced upregulation of CXCR4 expression: a mechanism of therapeutic resistance in pediatric AML.
Sison EA, McIntyre E, Magoon D, Brown P
Molecular cancer research : MCR 2013 Sep;11(9):1004-16
Molecular cancer research : MCR 2013 Sep;11(9):1004-16
Flagellin induces myeloid-derived suppressor cells: implications for Pseudomonas aeruginosa infection in cystic fibrosis lung disease.
Rieber N, Brand A, Hector A, Graepler-Mainka U, Ost M, Schäfer I, Wecker I, Neri D, Wirth A, Mays L, Zundel S, Fuchs J, Handgretinger R, Stern M, Hogardt M, Döring G, Riethmüller J, Kormann M, Hartl D
Journal of immunology (Baltimore, Md. : 1950) 2013 Feb 1;190(3):1276-84
Journal of immunology (Baltimore, Md. : 1950) 2013 Feb 1;190(3):1276-84
Human cytomegalovirus infection inhibits CXCL12- mediated migration and invasion of human extravillous cytotrophoblasts.
Warner JA, Zwezdaryk KJ, Day B, Sullivan DE, Pridjian G, Morris CA
Virology journal 2012 Nov 1;9:255
Virology journal 2012 Nov 1;9:255
Osteopontin enhances the expression and activity of MMP-2 via the SDF-1/CXCR4 axis in hepatocellular carcinoma cell lines.
Zhang R, Pan X, Huang Z, Weber GF, Zhang G
PloS one 2011;6(8):e23831
PloS one 2011;6(8):e23831
CD137 ligand signaling induces human monocyte to dendritic cell differentiation.
Kwajah M M S, Schwarz H
European journal of immunology 2010 Jul;40(7):1938-49
European journal of immunology 2010 Jul;40(7):1938-49
Erythromycin-induced CXCR4 expression on microvascular endothelial cells.
Takagi Y, Hashimoto N, Phan SH, Imaizumi K, Matsuo M, Nakashima H, Hashimoto I, Hayashi Y, Kawabe T, Shimokata K, Hasegawa Y
American journal of physiology. Lung cellular and molecular physiology 2009 Sep;297(3):L420-31
American journal of physiology. Lung cellular and molecular physiology 2009 Sep;297(3):L420-31
Human mast cell activation with virus-associated stimuli leads to the selective chemotaxis of natural killer cells by a CXCL8-dependent mechanism.
Burke SM, Issekutz TB, Mohan K, Lee PW, Shmulevitz M, Marshall JS
Blood 2008 Jun 15;111(12):5467-76
Blood 2008 Jun 15;111(12):5467-76
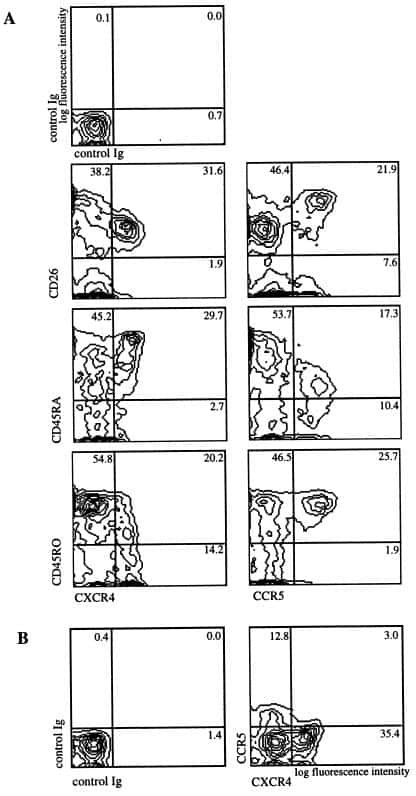
The HIV coreceptors CXCR4 and CCR5 are differentially expressed and regulated on human T lymphocytes.
Bleul CC, Wu L, Hoxie JA, Springer TA, Mackay CR
Proceedings of the National Academy of Sciences of the United States of America 1997 Mar 4;94(5):1925-30
Proceedings of the National Academy of Sciences of the United States of America 1997 Mar 4;94(5):1925-30
The HIV coreceptors CXCR4 and CCR5 are differentially expressed and regulated on human T lymphocytes.
Bleul CC, Wu L, Hoxie JA, Springer TA, Mackay CR
Proceedings of the National Academy of Sciences of the United States of America 1997 Mar 4;94(5):1925-30
Proceedings of the National Academy of Sciences of the United States of America 1997 Mar 4;94(5):1925-30
Inhibition of human immunodeficiency virus fusion by a monoclonal antibody to a coreceptor (CXCR4) is both cell type and virus strain dependent.
McKnight A, Wilkinson D, Simmons G, Talbot S, Picard L, Ahuja M, Marsh M, Hoxie JA, Clapham PR
Journal of virology 1997 Feb;71(2):1692-6
Journal of virology 1997 Feb;71(2):1692-6
No comments: Submit comment
Supportive validation
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
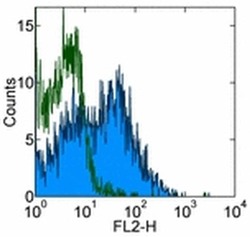
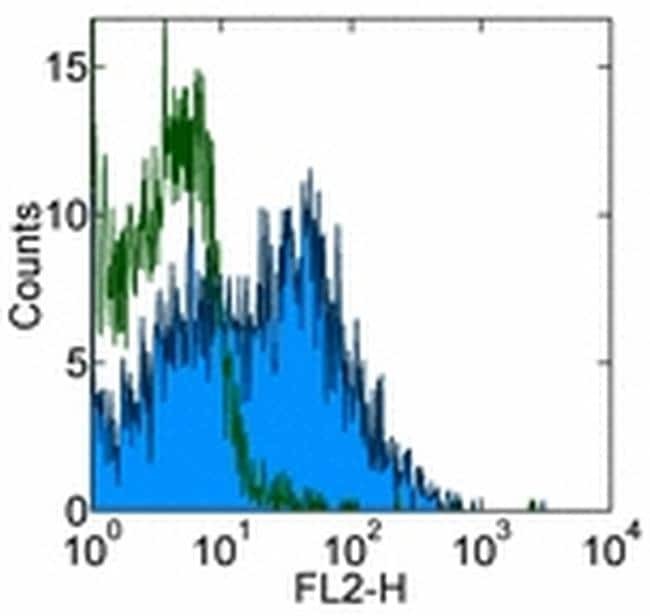
- Experimental details
- Staining of normal human peripheral blood cells with 0.25 µg of Mouse IgG2a K Isotype Control Purified (Product # 14-4724-82) (open histogram) or 0.25 µg of Anti-Human CD184 (CXCR4) Purified (filled histogram) followed by Anti-Mouse IgG Biotin (Product # 13-4013-85) and Streptavidin PE (Product # 12-4317-87).Cells in the lymphocyte gate were used for analysis.
Supportive validation
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
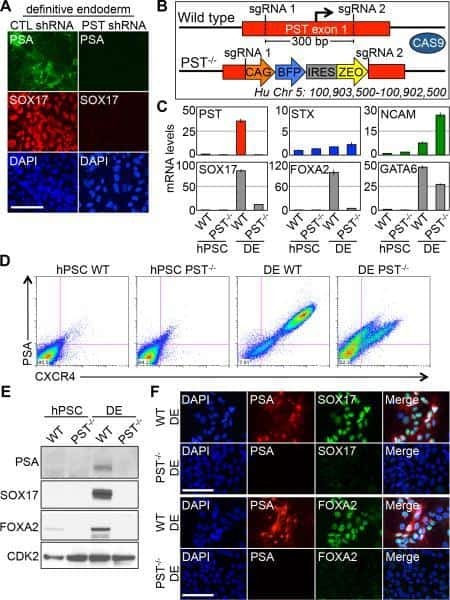
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
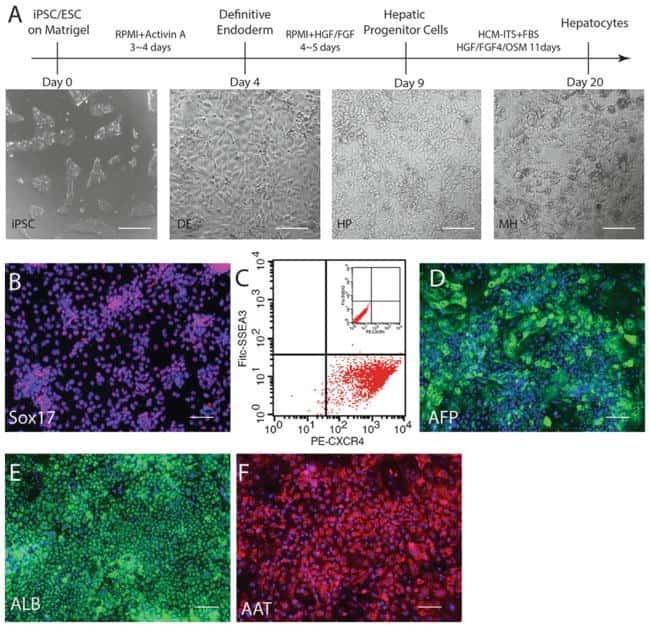
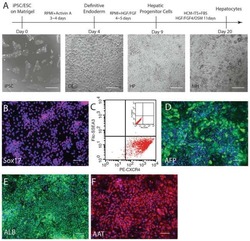
- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
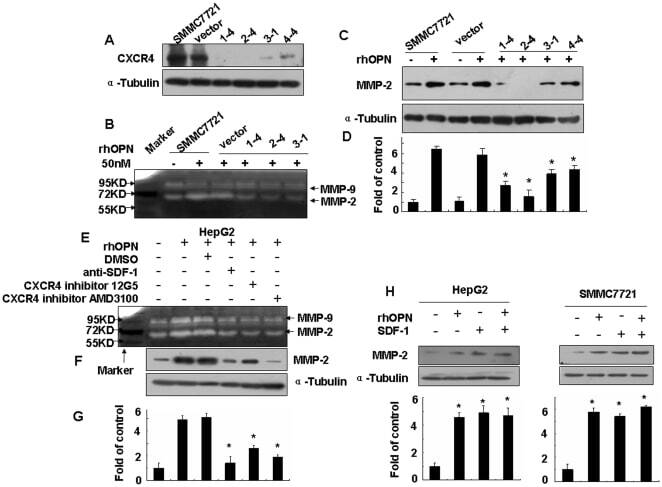
- Figure 2 Effects of the SDF-1alpha/CXCR4 axis on rhOPN-induced MMP-2 expression and activity. ( A ) Verification by Western blotting of the miRNA knockdown of CXCR4 showed a significant reduction of the CXCR4 protein in all clones (1-4, 2-4, 3-1, 4-4). After blocking the SDF-1alpha/CXCR4 axis with miRNA-CXCR4 and inhibitors (SDF-1 neutralizing antibody at 100 ng/ml, CXCR4 inhibitor 12G5 at 50 ug/ml, or CXCR4 inhibitor AMD3100 at 500 ng/ml), the cells were stimulated by rhOPN in serum-free medium for 60 hours, the cells were collected and analyzed by Western blotting in SMMC7721 cells ( C ) and in HepG2 cells ( F ). The supernatants of SMMC7721 cells ( B ) and HepG2 cells ( E ) were analyzed by gelatin zymography. ( D ) and ( G ) show the densitometric ratio of MMP-2 protein/alpha-tubulin. ( H ) Western blotting was used to assay the MMP-2 expression induced by rhOPN (50 nM) or/and SDF-1 (30 nM) for 48 hours. * denotes P
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
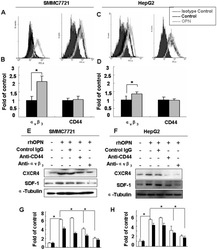
- Figure 3 Integrin alpha v beta 3 and CD44 mediated OPN-induced CXCR4 expression in SMMC7721 and HepG2 cells. FACS analysis using monoclonal antibodies to alpha v beta 3 integrin ( left ) and CD44 ( right ) was done for SMMC7721 cells ( A ) and HepG2 cells ( C ), stimulated by rhOPN for 24 hours. The grey area represents isotype control, while the dark line represents the control and the grey line represents the experimental group. SMMC7721 ( E ) and HepG2 ( F ) cells were treated with rhOPN (50 nM), in the presence of neutralizing antibodies to integrin alpha v beta 3 or CD44v6, or control IgG. After 60 hours, the cells were collected and Western blotting was performed to detect CXCR4. ( B ), ( D ), ( G ) and ( H ) are quantitative evaluations. The results are shown as mean +- standard deviation (n = 3). * denotes P

- Submitted by
- Invitrogen Antibodies (provider)
- Main image
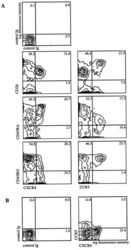
- Experimental details
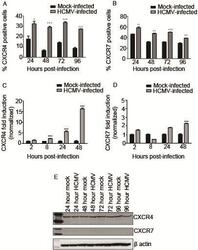
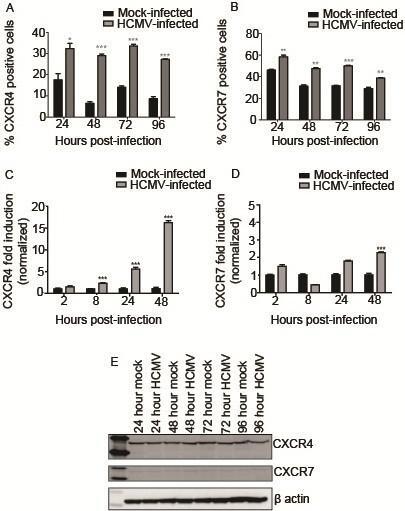
- Figure 3 HCMV infection alters expression of CXCR4 and CXCR7. SGHPL-4 cells were mock-infected or infected with HCMV, and then CXCR4 and CXCR7 receptor expression was determined. HCMV infection induced upregulation of cell surface staining for CXCR4 ( A ) and CXCR7 ( B ) at all time points studied. (*, p
- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
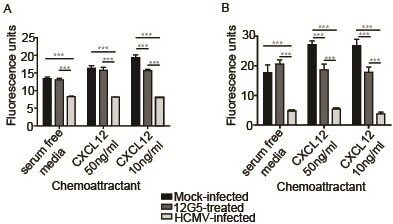
- Figure 4 HCMV infection blocks the ability of SGHPL-4 cells to migrate or invade toward CXCL12. Serum-starved SGHPL-4 cells were mock-infected or infected with Towne HCMV (MOI of 1) for 24 hours. Mock-infected cells were also pretreated with a neutralizing antibody against CXCR4 (12G5) prior to the assay. The cells were loaded into Fluoroblok inserts with 8 mum pores and chemoattractants (CXCL12 or EGF) were loaded into the lower chamber. Migration was allowed to proceed for 6 hours, whereas invasion assays were incubated for 24 hours, at which point they were labeled with Calcein AM and 10X images were acquired. HCMV infection significantly impaired migration ( A ) and invasion ( B ) toward CXCL12 at both 50 ng/ml and 10 ng/ml (p
- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
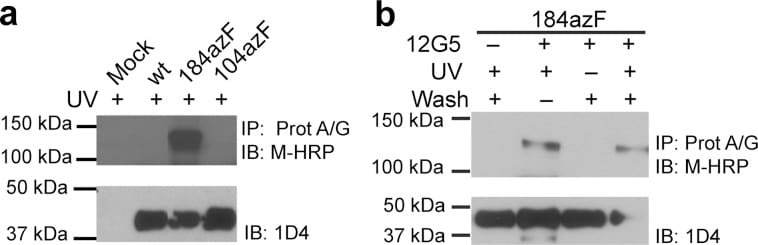
- Figure 2 Western blot analysis of photo-cross-linking of CXCR4variantsto 12G5. Detergent-solubilized cells were incubated with Protein A/GUltraLink resin, and immunoprecipitated samples were probed with theHRP-coupled anti-mouse secondary antibody. Receptor expression incrude lysates was detected with the 1D4 antibody to a C-terminal epitope.(a) UV-exposed samples containing CXCR4 incorporating azF at position104 (EC1) or 184 (EC2) are compared to wt and mock transfections.Specific bands appeared only in the Y184azF-CXCR4 lane. (b) Appearanceof unique bands in the Y184azF-CXCR4 sample tested in the presenceand absence of 12G5, UV exposure, and subsequent stringent washes.The bands are specific to 12G5 incubation and UV exposure. Blots arerepresentative of at least two independent trials.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
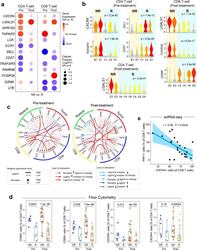
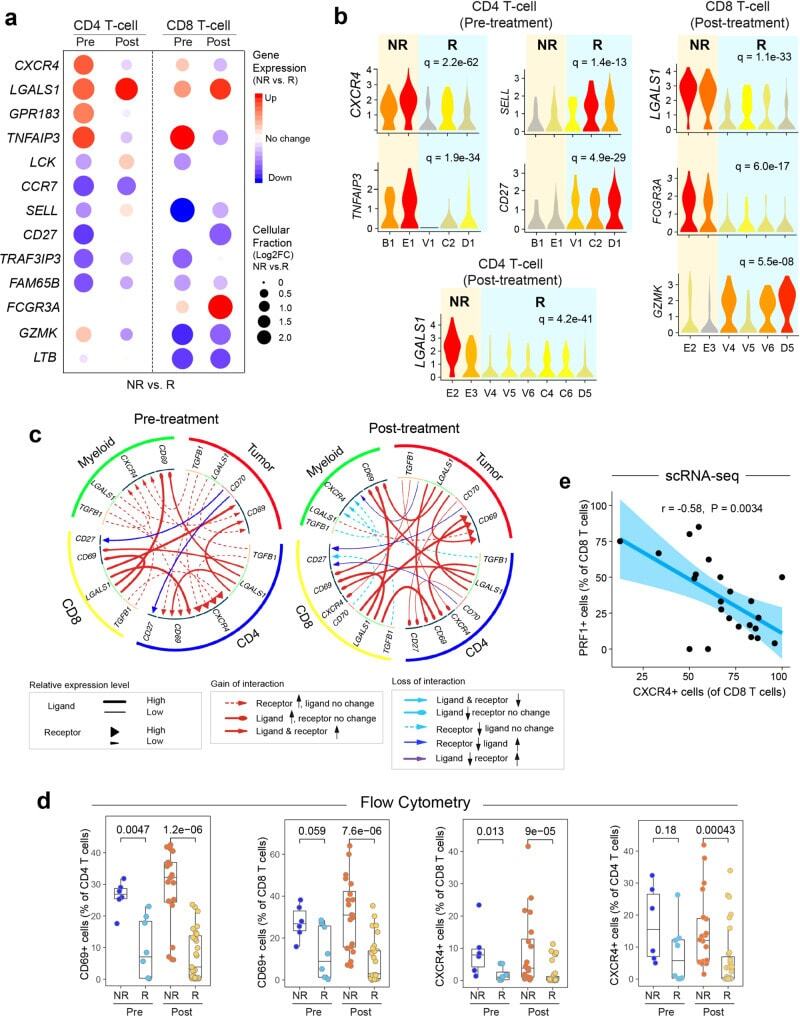
- Fig. 7 Aberrant cell-to-cell communication signaling associated with therapeutic resistance. a Differentially expressed genes (NR vs. R) in CD4+ and CD8+ T-cells pre- and post-ibrutinib treatment, respectively. Filled circle sizes are proportional to the Log2-scaled fold changes of each gene. Upregulated genes are shown in red; downregulated genes are shown in blue. Pre: pre-treatment; Post: post-treatment. b Representative genes are shown in violin plots. c Alterations (NR vs. R) of ligand-receptor-based cell-to-cell communication networks based in pre- and post-treatment samples. d Flow cytometry validation of upregulated CD69 and CXCR4 expression in ibrutinib nonresponders in comparison to the responders in additional patient cohorts ( n = 65 samples collected from 22 patients). The line in the box is the median value. The bottom and top of the box are the 25th and 75th percentiles of the sample. The bottom and top of the whiskers are the minimum and maximum values of the sample. p Values from the two-side Wilcoxon Signed-rank Test are shown. e Reverse correlation between the proportion of PRF1+ CD8 T cells (cytotoxic) and the expression of CXCR4 using scRNA-seq. The bounds of shape correspond to 95% confidence band for the regression line. The Pearson correlation coefficient ( r ) is shown. p Value corresponds to F test of linear regression model.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
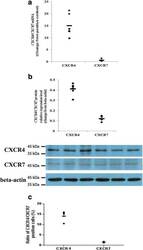
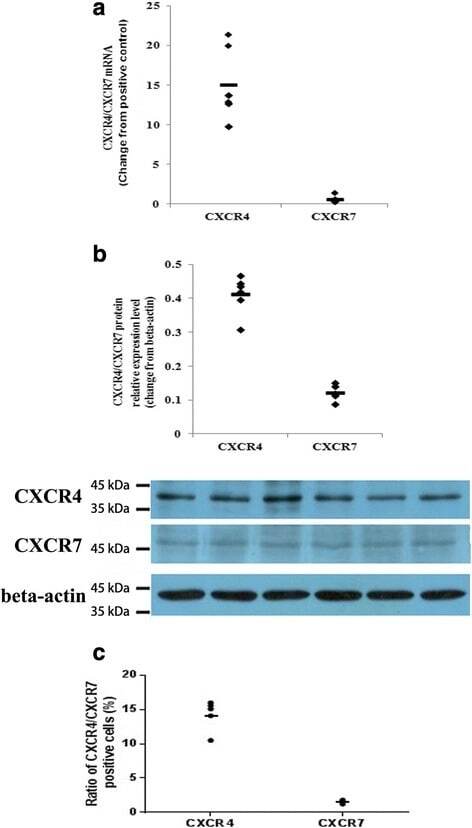
- Fig. 4 Expression levels of CXCR4 and CXCR7 in primary human first-trimester decidual epithelial cell cultures. Expression of the CXCR4 and CXCR7 mRNAs and proteins was detected in primary human first-trimester decidual epithelial cell (DEC) cultures. a The expression of the CXCR4 and CXCR7 genes in DECs was examined by real-time PCR. The level of the CXCR4 or CXCR7 gene was equal to the ratio of the absorbance of the target gene to the control. The transverse line represents the average CXCR4 and CXCR7 levels in DECs. ( n = 6). b The expression of CXCR4 and CXCR7 proteins in DECs was detected by western blotting. ( n = 6). c Flow cytometry results of the expression of CXCR4 and CXCR7 on the membranes human first-trimester DECs ( n = 5). The transverse line represents mean percentage of CXCR4 or CXCR7 positive cells
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
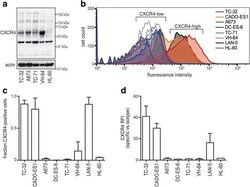
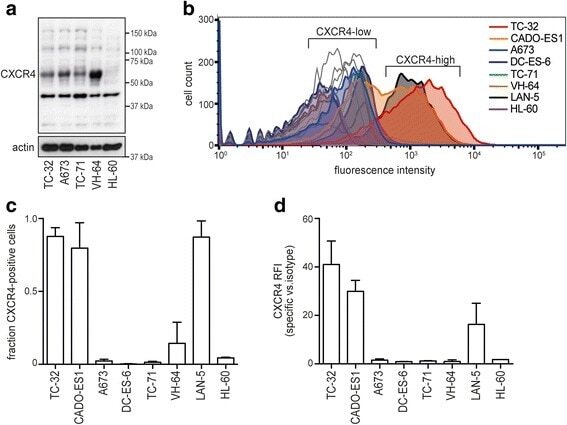
- Fig. 3 Ewing sarcoma cell lines group into CXCR4-high and -low surface expression. a Western blot analysis of total CXCR4 protein reveals multiple isoforms. Cells were grown to 70-80% confluence in standard conditions. Actin served as loading control. ( b )-( d ) CXCR4 surface expression distinguishes CXCR4-high and -low cell lines. Cells were grown as to 70-80% confluence in standard conditions and analyzed for CXCR4 surface expression by flow cytometry. LAN-5 neuroblastoma cells served as CXCR4-high positive control. b Representative flow cytometry plots; open graphs represent isotype-antibody controls. c CXCR4 positive cell populations depicted as mean +- SD of three independent analyses. d Relative fluorescence intensities of CXCR4 positive cell populations in ( c ). Corresponding data in serum-free conditions are provided in Additional file 1
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
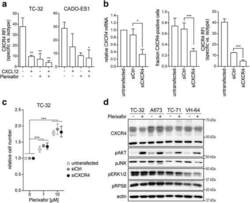
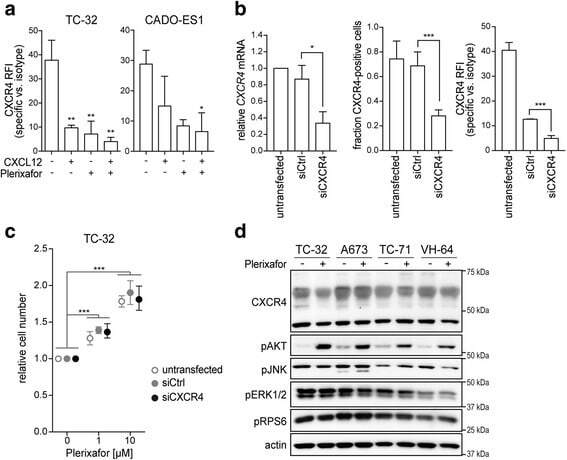
- Fig. 4 Plerixafor-induced proliferation does not require substantial CXCR4 surface expression but is associated with AKT activation. a CXCL12 and plerixafor bind to the CXCR4 receptor. TC-32 and CADO-ES1 cells were grown in standard conditions and treated with CXCL12 (100 ng/ml) and/or plerixafor (1 muM) for 12 h before flow cytometric analysis of CXCR4 signal with a 12G5-CXCR4 antibody. b Significant reduction in CXCR4 mRNA (left panel) and surface protein expression (middle and right panel). Cells were transfected with CXCR4-targeting siRNA (siCXCR4) or non-silencing control (siCtrl) and analyzed by real-time quantitative PCR and flow cytometry after 48 h. c Proliferative plerixafor effects are maintained despite significant reduction in CXCR4 surface expression. 48 h after siRNA transfection, cells from ( b ) were treated with plerixafor for another 72 h. Relative cell number was measured by WST-1 assay. All graphs of this figure represent means +- SD of three independent experiments. d AKT is activated in response to plerixafor in CXCR4-high and -low cell lines. Cells grown in standard conditions were treated with plerixafor (1 muM) for 12 h or remained untreated. Whole cell lysates were analyzed by Western blotting. Actin was loading control
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
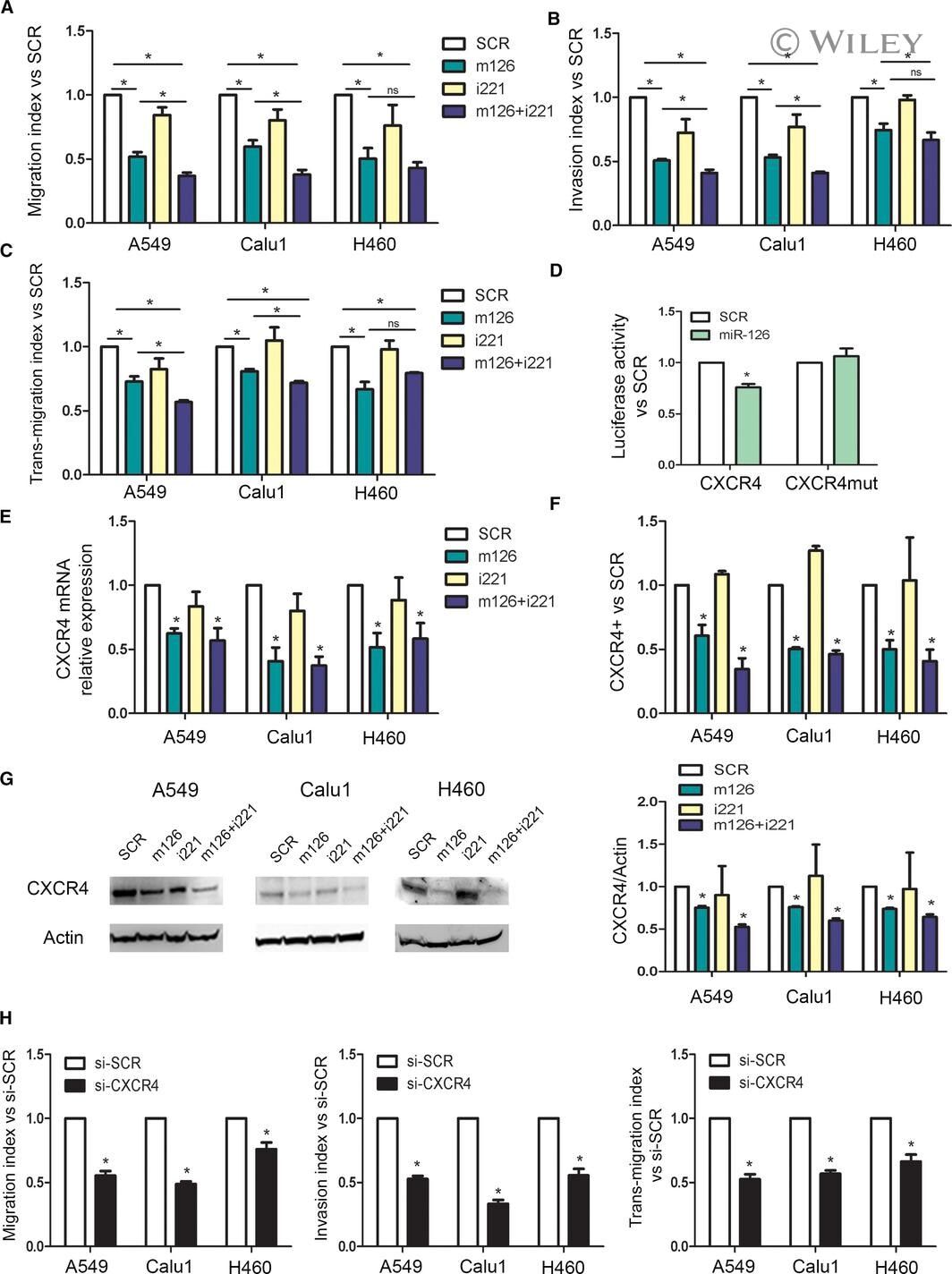
- 4 Fig. miR-126-3p replacement and miR-221 inhibition reduced lung dissemination by inhibiting CXCR4 levels. (A) Migration index after miR-126-3p and miR-221-3p modulation of three lung cancer cells ( n = 5). (B) Invasive capacity of lung cancer cells transfected with miR-126/221 ( n = 4). (C) Number of transmigrated lung cancer cells through endothelial monolayer after miRNA modulation ( n = 3). (D) Luciferase activity for 3'UTR of CXCR4 wild-type and mutated sequence after miR-126 and SCR transfection in A549 cells ( n = 4). CXCR4 levels after miR-126-3p replenishment and miR-221-3p inhibition evaluated by qPCR ( n = 3) (E) and flow cytometry ( n = 3) (F). (G) Representative images and quantification of CXCR4 in all the three cell lines after miRNA modulation ( n = 3). (H) Migration, invasion and transendothelial index of lung cancer cells transient transfected with CXCR4 siRNA and controls ( n = 3 for each cell line). Statistically significant differences were determined with Student's t -test when comparing two groups or ANOVA test for multiple comparisons. * P < 0.05 versus controls. Data are expressed as mean +- S.E.M.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
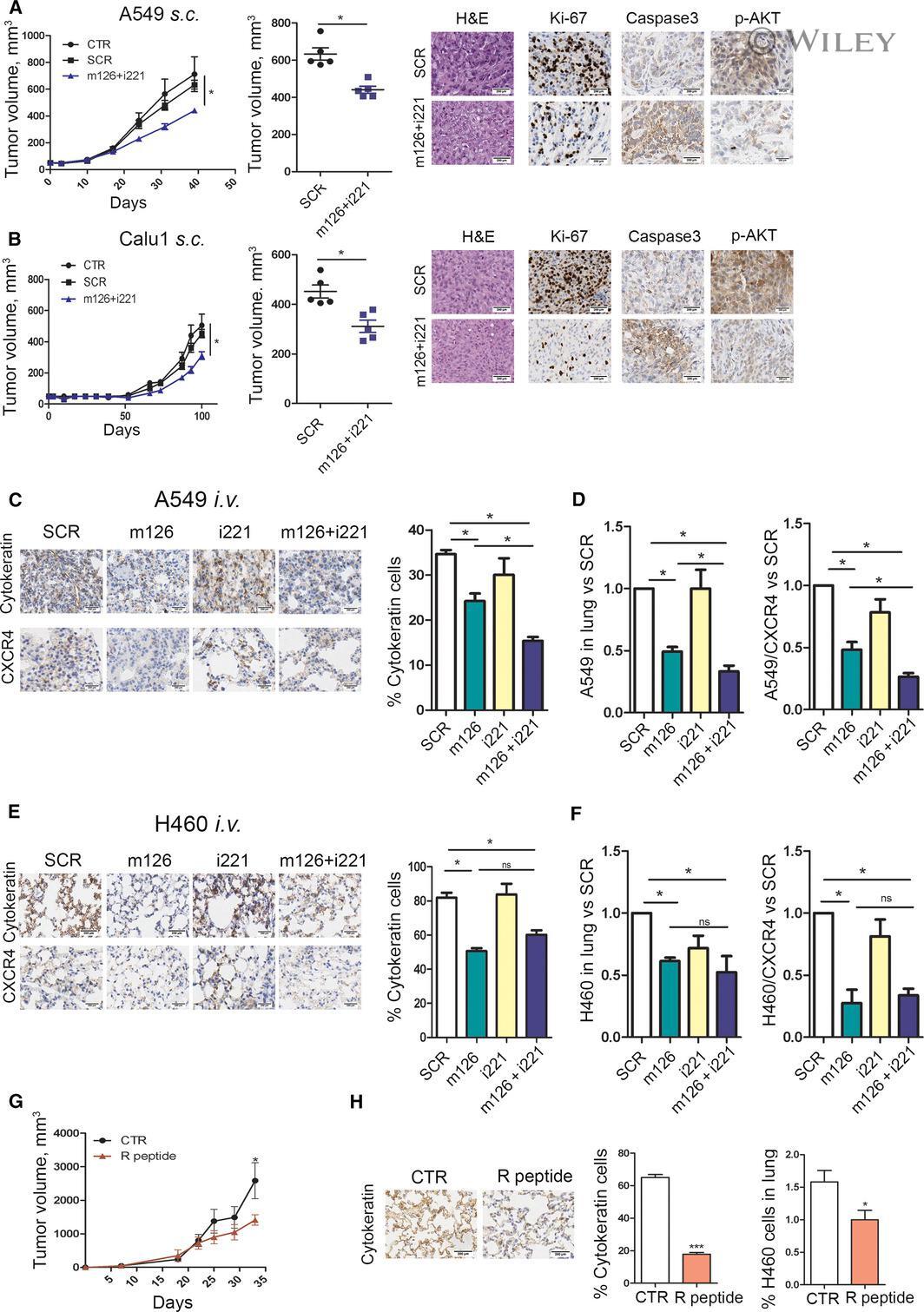
- 5 Fig. Combined replenishment of miR-126-3p and inhibition of miR-221-3p reduced xenograft growth in vivo . (A) A549 transfected with both miR-126-3p mimic (m126) and miR-221-3p inhibitor (i221) showed a reduction in tumour growth in immunodeficient mice compared with SCR and CTR controls. Ki-67, cleaved caspase 3 and phosphor-AKT quantification and representative images in A549 m126+i221 cells compared with controls. Necrosis was evaluated on haematoxylin & eosin (H&E) staining in subcutaneously implanted xenograft after miRNA transfection ( n = 5 for each group) Scale bar: 200 um (B) miR-126-3p replacement and miR-221-3p inhibition reduced tumour growth in subcutaneous Calu1 xenografts in immunodeficient mice. Ki-67, cleaved caspase 3 and phospho-AKT count and staining revealed a reduction in proliferation rate and increase in apoptosis of these cells. Necrosis was evaluated on H&E staining ( n = 5 for each group). Scale bar: 200 um. (C) Pan-cytokeratin and CXCR4 staining and quantification in the lungs of mice treated with A549 transfected miR-126 and/or miR-221 alone or in combination ( n = 4 for each group). Scale bar: 200 um. (D) Quantification of A549 and A549/CXCR4+ cells in the lungs of mice by flow cytometry ( n = 4). (E) Representative images and quantification of pan-cytokeratin and CXCR4 staining of the lungs from H460 transfected with miR-126-3p, miR-221 inhibitors alone or in combination and control ( n = 4 mice for each group). Scale bar: 200 um. (F) Flow cyto
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
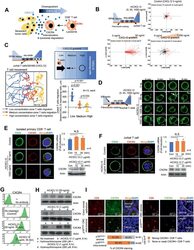
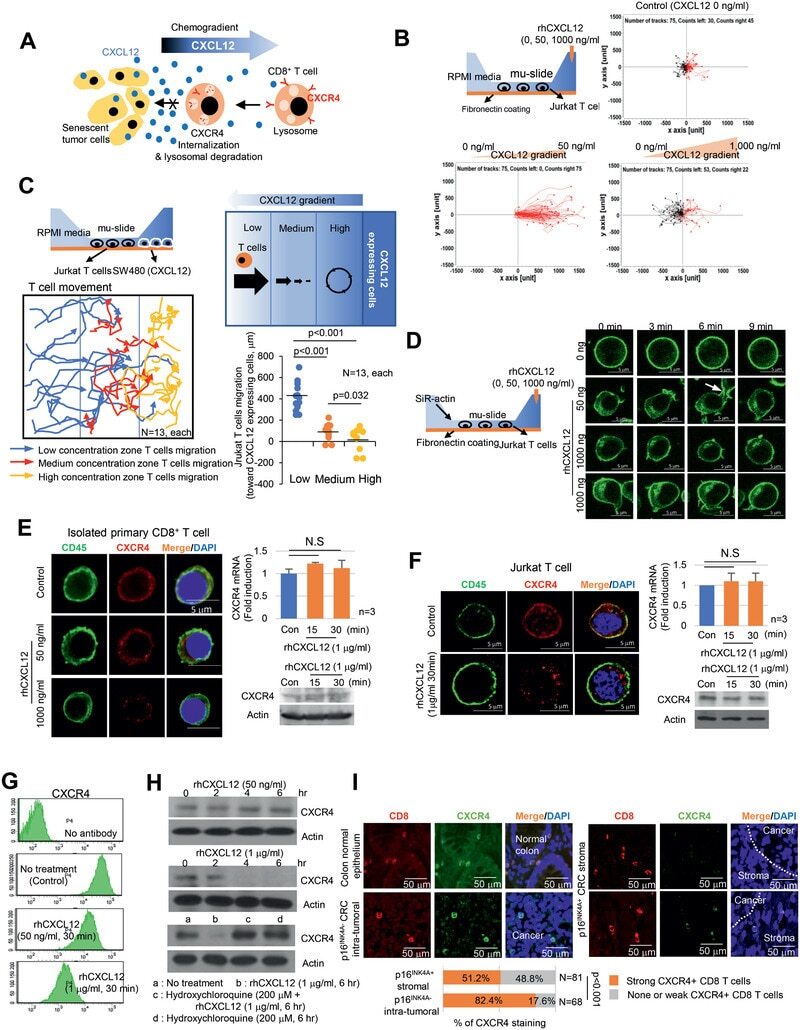
- Figure 4 High concentrations of CXCL12 induce the loss of the plasma membrane CXCR4 in T cells. A) Schematic representation of CD8 + T cell migration in the presence of senescent tumor cells. B) CXCL12 chemogradient was developed using u-slide and then Jurkat T cell migration was analyzed. The migrated T cells were tracked, and the results are presented graphically ( n = 75, each). The red and black lines indicate the tracks of the chemoattracted and chemorepulsed cells, respectively. C) The CXCL12 chemogradient was developed by CXCL12 overexpressing SW480 cells in u-slide, and T cell migration was analyzed. The migrated T cells were tracked, and the result is presented as a line graph (left panel). The migration distances were measured, and the result is presented graphically (lower right panel). The p value was calculated by one-way ANOVA and post hoc analysis. D) Low and high concentrations of rhCXCL12 were treated with u-slide and lamellipodia formation was analyzed with SiR-actin staining. The arrow indicates lamellipodia. E and F) CXCR4 expression in isolated primary CD8 + T cells E) and Jurkat T cells F). Isolated primary CD8 + T or Jurkat cells were treated with 50 or 1000 ng mL -1 of rhCXCL12 for 30 min and analyzed for CD45 by immunocytochemistry and CXCR4 expression by immunocytochemistry, real-time PCR and western blot. G) Jurkat T cells were treated with rhCXCL12 (50 or 1000 ng mL -1 ) for 30 min and analyzed for CXCR4 expression in the plasma membrane by FACS. H
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
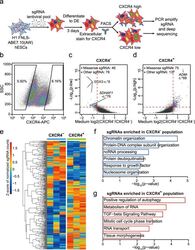
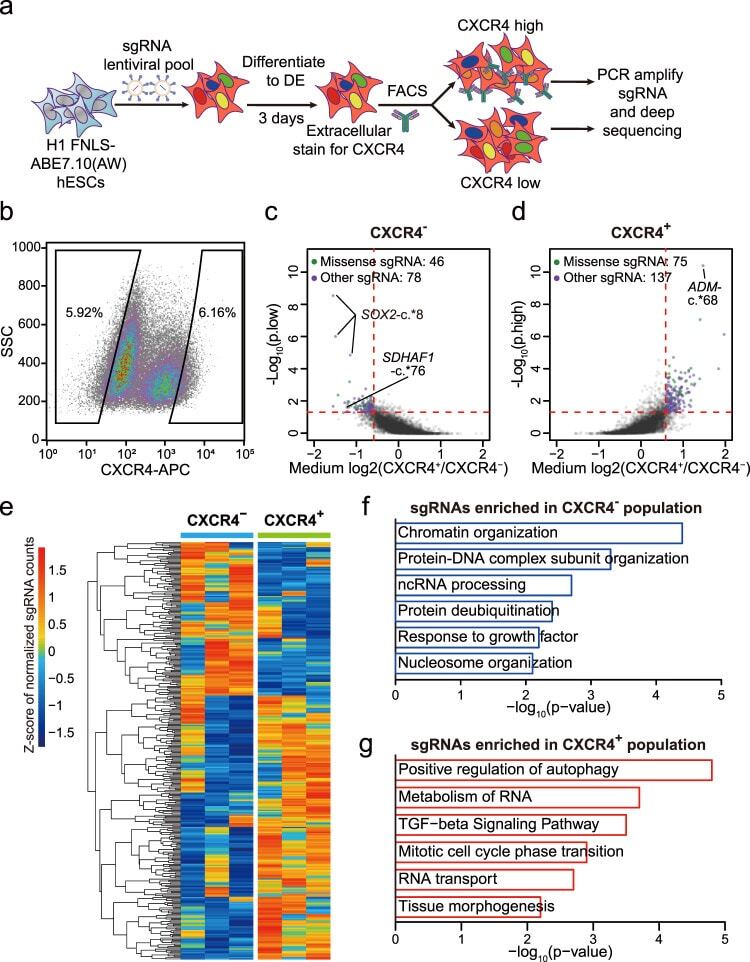
- Fig. 3 FNLS-ABE7.10(AW) base editor-based functional screening of m 6 A sites in H1 hESCs. a Flow chart of ABE-based screening of m 6 A sites is important for endoderm specification. b Representative CXCR4 FACS gating strategy for sorting CXCR4 - and CXCR4 + populations, respectively. SSC side scatter. c , d Scatter plot showing the strategy to determine the sgRNAs significantly enriched in CXCR4 - ( c ) and CXCR4 + ( d ) populations, respectively. e Heatmap showing the normalized counts of sgRNAs enriched in CXCR4 - or CXCR4 + populations for each replicate. f , g GO-enrichment analyses of the genes targeted by the sgRNAs significantly enriched in the CXCR4 - ( f ) or CXCR4 + ( g ) populations, respectively.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
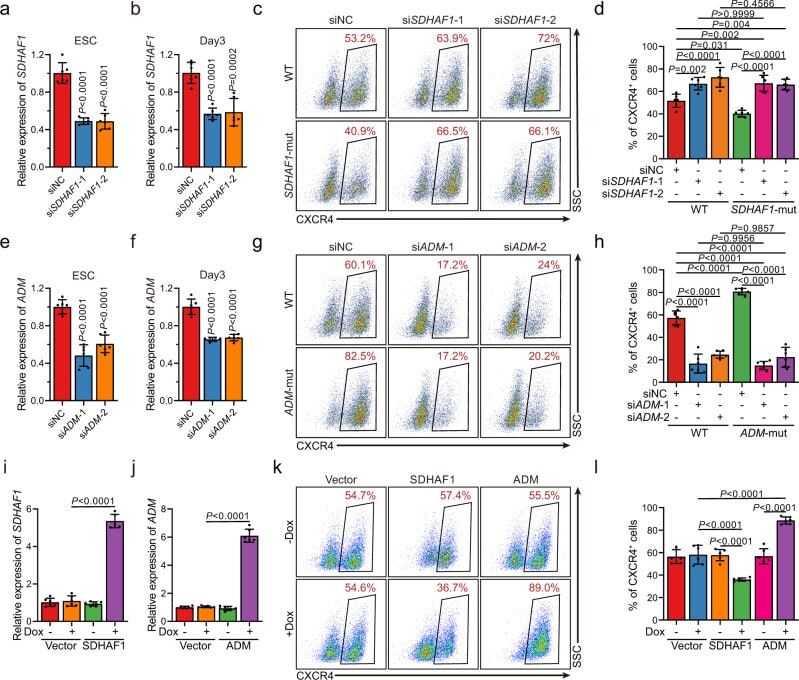
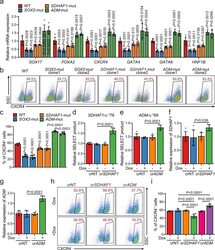
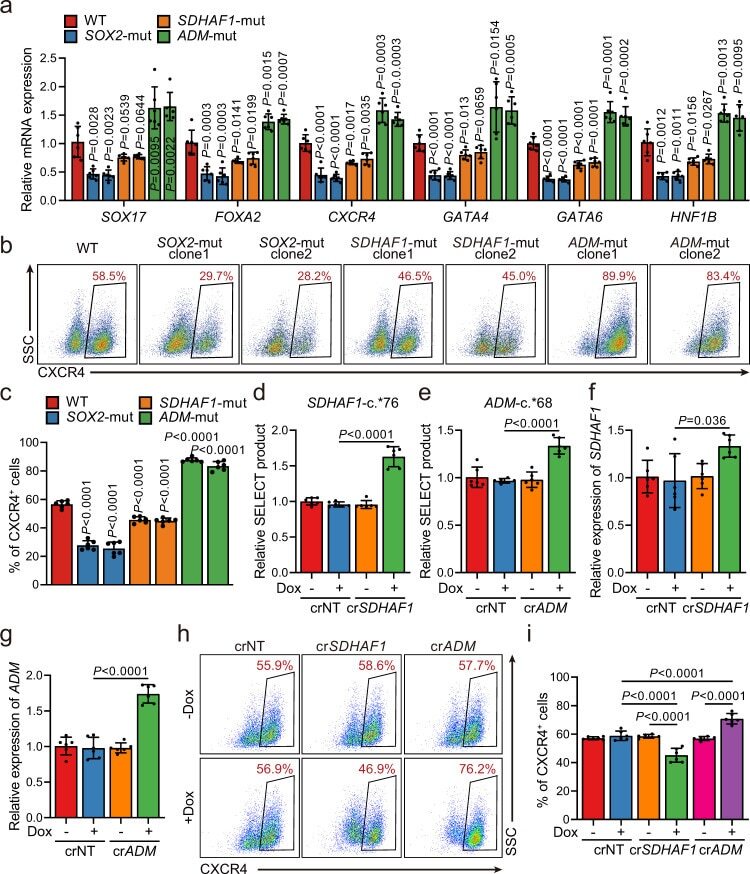
- Fig. 5 Selected m 6 A-disruptive mutations regulate human endodermal specification through m 6 A. a mRNA levels of DE marker genes SOX17 , FOXA2 , CXCR4 , GATA4 , GATA6 , and HNF1B in cultures derived from WT, SOX2 -mut, SDHAF1 -mut, and ADM -mut as well as WT hESCs at day 3 of DE differentiation ( n = 6 biologically independent samples). P -values were calculated vs. WT (two-tailed Student's t -test). b , c Representative ( b ) and quantitative ( c ) flow cytometry analyses of CXCR4 expression in cultures derived from WT, SOX2 -mut, SDHAF1 -mut, and ADM -mut hESCs at day 3 of DE differentiation ( n = 6 biologically independent samples). P -values were calculated vs. WT (one-way ANOVA with Tukey's post hoc test). d , e Measurement of m 6 A levels of SDHAF1 -c. *76 ( d ) and ADM -c. *68 ( e ) after TRME editor induced site-specific m 6 A demethylation by SELECT assay ( n = 6 biologically independent samples). NT non-targeting, Dox doxycycline (one-way ANOVA with Tukey's post hoc test). f , g mRNA expression levels of SDHAF1 ( f ) and ADM ( g ) on day 2 of DE differentiation after TRME editor induced site-specific m 6 A demethylation ( n = 6 biologically independent samples). NT non-targeting, Dox doxycycline (one-way ANOVA with Tukey's post hoc test). h , i Representative ( h ) and quantitative ( i ) flow cytometry analyses of CXCR4 expression in cultures derived from hESCs with or without induced site-specific m 6 A demethylation by TRME editor at day 3 of DE differentiation
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
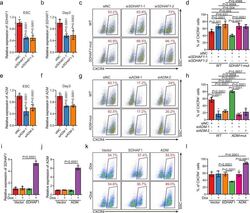
- Fig. 6 The mRNAs of SDHAF1 and ADM mediate the effects of their targeting sgRNAs on DE specification. a , b siRNA knockdown efficiency of SDHAF1 in hESCs ( a ) and at day 3 of DE differentiation ( b ), respectively ( n = 6 biologically independent samples), measured by RT-qPCR. si siRNA, NC non-targeting control. P -values were calculated vs. siNC (one-way ANOVA with Tukey's post hoc test). c , d Representative ( c ) and quantitative ( d ) flow cytometry analyses of CXCR4 expression in cultures derived from WT or SDHAF1- mut hESCs that with NC or SDHAF1 -targeting siRNAs at day 3 of DE differentiation ( n = 6 biologically independent samples). ns non-significant (one-way ANOVA with Tukey's post hoc test). e , f siRNA knockdown efficiency of ADM in hESCs ( e ) and at day 3 of DE differentiation ( f ) respectively ( n = 6 biologically independent samples), measured by RT-qPCR. si siRNA, NC non-targeting control. P -values were calculated vs. siNC (one-way ANOVA with Tukey's post hoc test). g , h Representative ( g ) and quantitative ( h ) flow cytometry analyses of CXCR4 expression in cultures derived from WT or ADM -mut hESCs that with NC or ADM -targeting siRNAs at day 3 of DE differentiation ( n = 6 biologically independent samples). ns non-significant (one-way ANOVA with Tukey's post hoc test). i , j mRNA levels of SDHAF1 ( i ) and ADM ( j ) in overexpressed hESCs ( n = 6 biologically independent samples), measured by RT-qPCR. Dox, doxycycline (one-way ANOVA with Tukey's po