Explore
Explore Validate
Validate Learn
Learn Western blot
Western blotAntibody data
- Antibody Data
- Antigen structure
- References [107]
- Comments [0]
- Validations
- Western blot [3]
- Immunohistochemistry [1]
- Other assay [76]
Submit
Validation data
Reference
Comment
Report error
- Product number
- 51-2700 - Provider product page

- Provider
- Invitrogen Antibodies
- Product name
- beta Amyloid Polyclonal Antibody (CT695)
- Antibody type
- Polyclonal
- Antigen
- Synthetic peptide
- Description
- This antibody can be used to specifically detect the beta-amyloid precursor protein. The antibody reacts with full-length (APP695, 751, 770) and N-terminal truncated forms of beta-APP. The antibody can also be used to detect the C-terminal membrane-anchored fragment of beta-APP that remains after alpha- or beta-secretase cleavage. This antibody does not detect the beta-APP product N-terminal to the gamma-secretase cleavage site.
- Antibody clone number
- CT695
- Concentration
- 0.25 mg/mL
Submitted references Enhancing autophagy maturation with CCZ1-MON1A complex alleviates neuropathology and memory defects in Alzheimer disease models.
Cholesterol biosynthesis defines oligodendrocyte precursor heterogeneity between brain and spinal cord.
Malaria parasite heme biosynthesis promotes and griseofulvin protects against cerebral malaria in mice.
Amelioration of Alzheimer's disease pathology by mitophagy inducers identified via machine learning and a cross-species workflow.
Blast-induced axonal degeneration in the rat cerebellum in the absence of head movement.
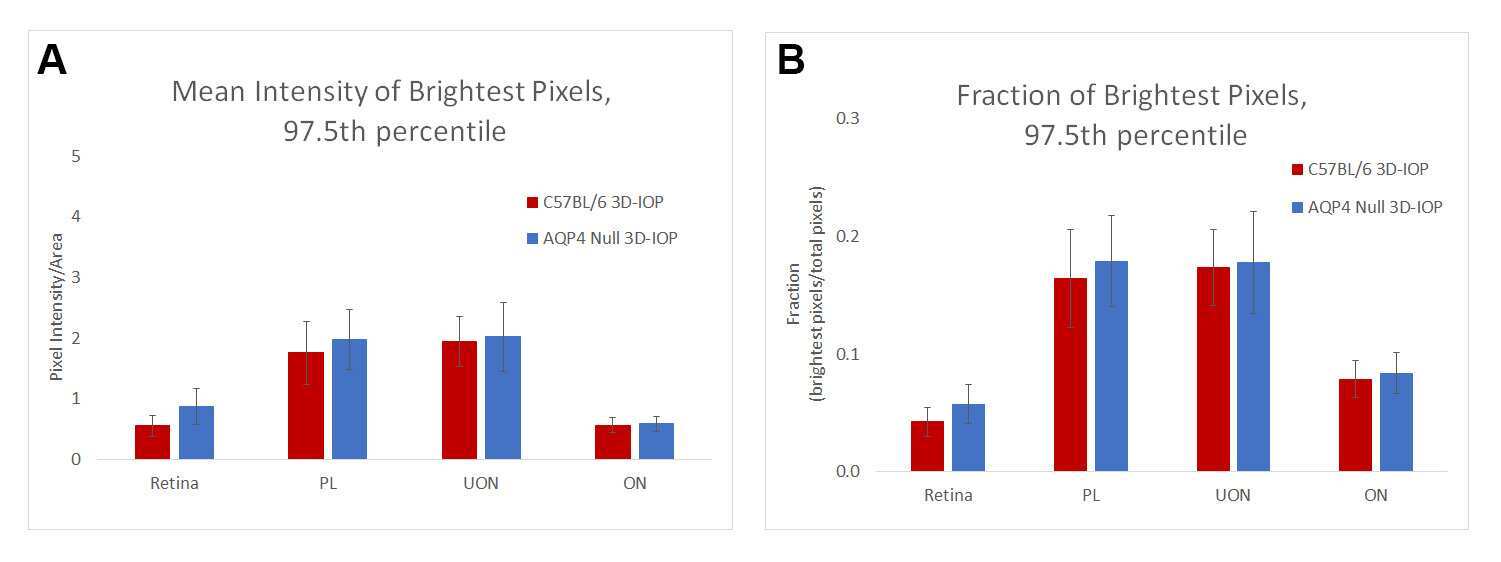
The role of aquaporin-4 in optic nerve head astrocytes in experimental glaucoma.
Electroacupuncture ameliorates beta-amyloid pathology and cognitive impairment in Alzheimer disease via a novel mechanism involving activation of TFEB (transcription factor EB).
Deletion of Abi3 gene locus exacerbates neuropathological features of Alzheimer's disease in a mouse model of Aβ amyloidosis.
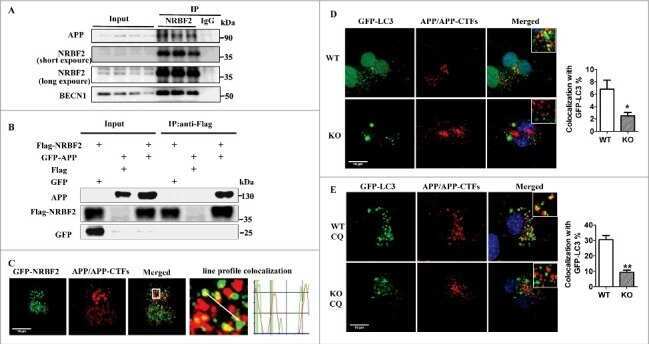
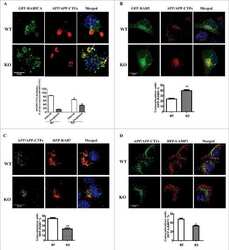
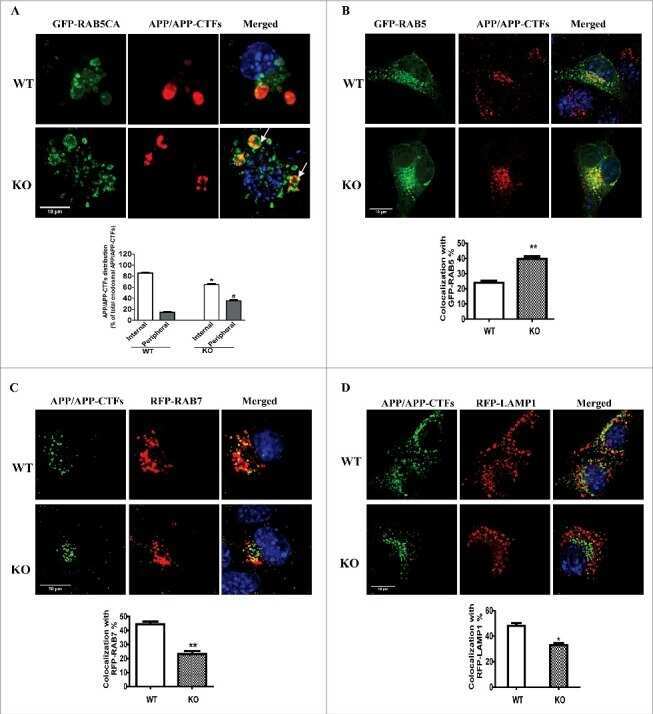
NRBF2 is a RAB7 effector required for autophagosome maturation and mediates the association of APP-CTFs with active form of RAB7 for degradation.
Genetic inactivation of SARM1 axon degeneration pathway improves outcome trajectory after experimental traumatic brain injury based on pathological, radiological, and functional measures.
Mouse closed head traumatic brain injury replicates the histological tau pathology pattern of human disease: characterization of a novel model and systematic review of the literature.
Protein farnesylation is upregulated in Alzheimer's human brains and neuron-specific suppression of farnesyltransferase mitigates pathogenic processes in Alzheimer's model mice.
Hyperoxygenation Treatment Reduces Beta-amyloid Deposition via MeCP2-dependent Upregulation of MMP-2 and MMP-9 in the Hippocampus of Tg-APP/PS1 Mice.
Endothelium-specific deletion of amyloid-β precursor protein exacerbates endothelial dysfunction induced by aging.
Buprenorphine alters microglia and astrocytes acutely following diffuse traumatic brain injury.
Phosphoinositide 3-kinase inhibitor AS605240 ameliorates streptozotocin-induced Alzheimer's disease like sporadic dementia in experimental rats.
Operation Brain Trauma Therapy: An Exploratory Study of Levetiracetam Treatment Following Mild Traumatic Brain Injury in the Micro Pig.
Sex-Specific Differences in Rodents Following a Single Primary Blast Exposure: Focus on the Monoamine and Galanin Systems.
The endogenous neuronal complement inhibitor SRPX2 protects against complement-mediated synapse elimination during development.
Systemic Exposure to Air Pollution Induces Oxidative Stress and Inflammation in Mouse Brain, Contributing to Neurodegeneration Onset.
The Proteasomal Deubiquitinating Enzyme PSMD14 Regulates Macroautophagy by Controlling Golgi-to-ER Retrograde Transport.
Periventricular White Matter Alterations From Explosive Blast in a Large Animal Model: Mild Traumatic Brain Injury or "Subconcussive" Injury?
A method to quantify regional axonal transport blockade at the optic nerve head after short term intraocular pressure elevation in mice.
Long-term cognitive impairment without diffuse axonal injury following repetitive mild traumatic brain injury in rats.
A small molecule transcription factor EB activator ameliorates beta-amyloid precursor protein and Tau pathology in Alzheimer's disease models.
Diffuse Axonal Injury in the Rat Brain: Axonal Injury and Oligodendrocyte Activity Following Rotational Injury.
Repetitive Concussive and Subconcussive Injury in a Human Tau Mouse Model Results in Chronic Cognitive Dysfunction and Disruption of White Matter Tracts, But Not Tau Pathology.
Mechanical Stretch of High Magnitude Provokes Axonal Injury, Elongation of Paranodal Junctions, and Signaling Alterations in Oligodendrocytes.
Vascular phenotype of amyloid precursor protein-deficient mice.
EPPS treatment attenuates traumatic brain injury in mice by reducing Aβ burden and ameliorating neuronal autophagic flux.
The Importance of Inter-Species Variation in Traumatic Brain Injury-Induced Alterations of Microglial-Axonal Interactions.
A Comparative Study of Two Blast-Induced Traumatic Brain Injury Models: Changes in Monoamine and Galanin Systems Following Single and Repeated Exposure.
A Mild Traumatic Brain Injury in Mice Produces Lasting Deficits in Brain Metabolism.
modCHIMERA: a novel murine closed-head model of moderate traumatic brain injury.
Mild Traumatic Brain Injury Induces Structural and Functional Disconnection of Local Neocortical Inhibitory Networks via Parvalbumin Interneuron Diffuse Axonal Injury.
Wnt signaling loss accelerates the appearance of neuropathological hallmarks of Alzheimer's disease in J20-APP transgenic and wild-type mice.
Matrix-Assisted Laser Desorption Ionization Mapping of Lysophosphatidic Acid Changes after Traumatic Brain Injury and the Relationship to Cellular Pathology.
Primary Traumatic Axonopathy in Mice Subjected to Impact Acceleration: A Reappraisal of Pathology and Mechanisms with High-Resolution Anatomical Methods.
NRBF2 is involved in the autophagic degradation process of APP-CTFs in Alzheimer disease models.
Neuronal Injury and Glial Changes Are Hallmarks of Open Field Blast Exposure in Swine Frontal Lobe.
Defining an Analytic Framework to Evaluate Quantitative MRI Markers of Traumatic Axonal Injury: Preliminary Results in a Mouse Closed Head Injury Model.
The connectomics of brain demyelination: Functional and structural patterns in the cuprizone mouse model.
The Amyloid Precursor Protein of Alzheimer's Disease Clusters at the Organelle/Microtubule Interface on Organelles that Bind Microtubules in an ATP Dependent Manner.
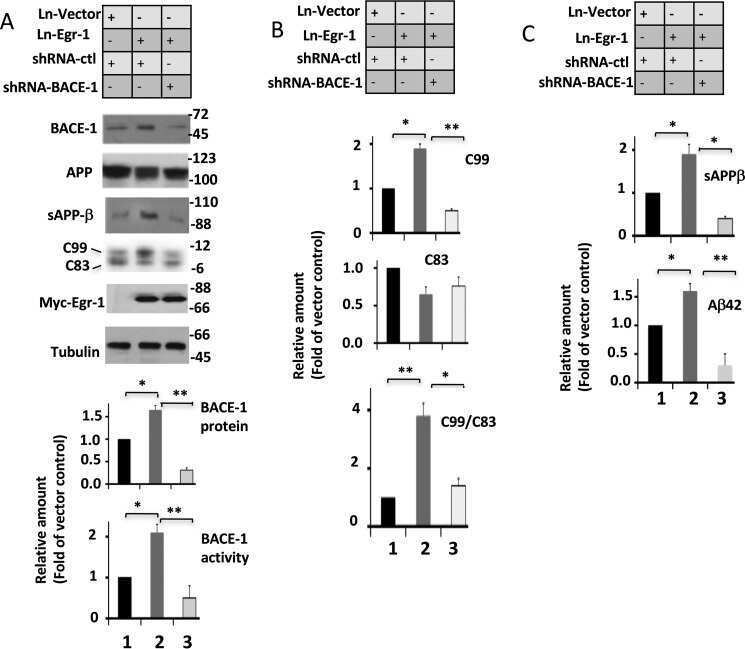
Early Growth Response 1 (Egr-1) Is a Transcriptional Activator of β-Secretase 1 (BACE-1) in the Brain.
Inhibition of Wnt signaling induces amyloidogenic processing of amyloid precursor protein and the production and aggregation of Amyloid-β (Aβ)(42) peptides.
Tetrahydrohyperforin Inhibits the Proteolytic Processing of Amyloid Precursor Protein and Enhances Its Degradation by Atg5-Dependent Autophagy.
Losartan Treatment Protects Retinal Ganglion Cells and Alters Scleral Remodeling in Experimental Glaucoma.
A novel closed-body model of spinal cord injury caused by high-pressure air blasts produces extensive axonal injury and motor impairments.
Kalirin-9 and Kalirin-12 Play Essential Roles in Dendritic Outgrowth and Branching.
Blast overpressure induced axonal injury changes in rat brainstem and spinal cord.
Microglia processes associate with diffusely injured axons following mild traumatic brain injury in the micro pig.
TREM2 regulates microglial cell activation in response to demyelination in vivo.
Traumatic brain injury-induced axonal phenotypes react differently to treatment.
Ccr2 deletion dissociates cavity size and tau pathology after mild traumatic brain injury.
Defects of Lipid Synthesis Are Linked to the Age-Dependent Demyelination Caused by Lamin B1 Overexpression.
Neurotransmitter Systems in a Mild Blast Traumatic Brain Injury Model: Catecholamines and Serotonin.
Array tomography for the detection of non-dilated, injured axons in traumatic brain injury.
Fragile X mental retardation protein expression in Alzheimer's disease.
Identification and Preclinical Pharmacology of the γ-Secretase Modulator BMS-869780.
Moderately elevated intracranial pressure after diffuse traumatic brain injury is associated with exacerbated neuronal pathology and behavioral morbidity in the rat.
Acute reduction of microglia does not alter axonal injury in a mouse model of repetitive concussive traumatic brain injury.
Experimental traumatic brain injury induces rapid aggregation and oligomerization of amyloid-beta in an Alzheimer's disease mouse model.
Cortical hypoexcitation defines neuronal responses in the immediate aftermath of traumatic brain injury.
AAD-2004 Attenuates Progressive Neuronal Loss in the Brain of Tg-betaCTF99/B6 Mouse Model of Alzheimer Disease.
Myelin loss and oligodendrocyte pathology in white matter tracts following traumatic brain injury in the rat.
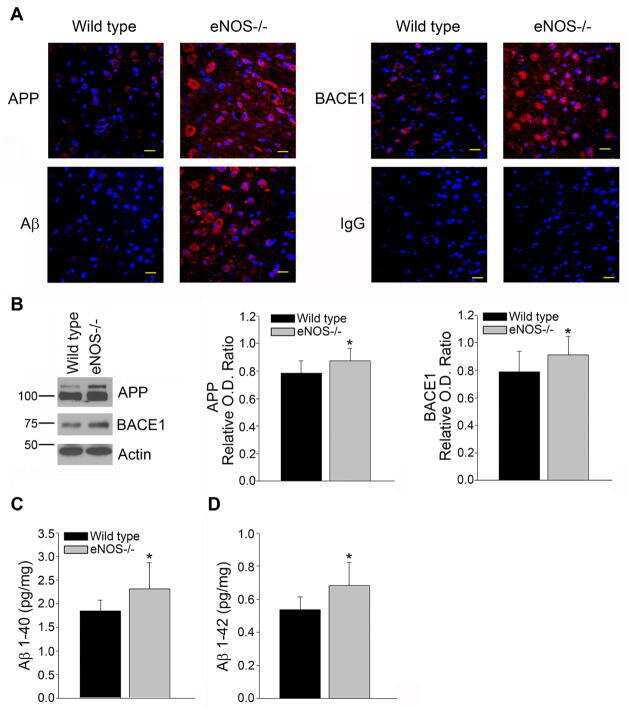
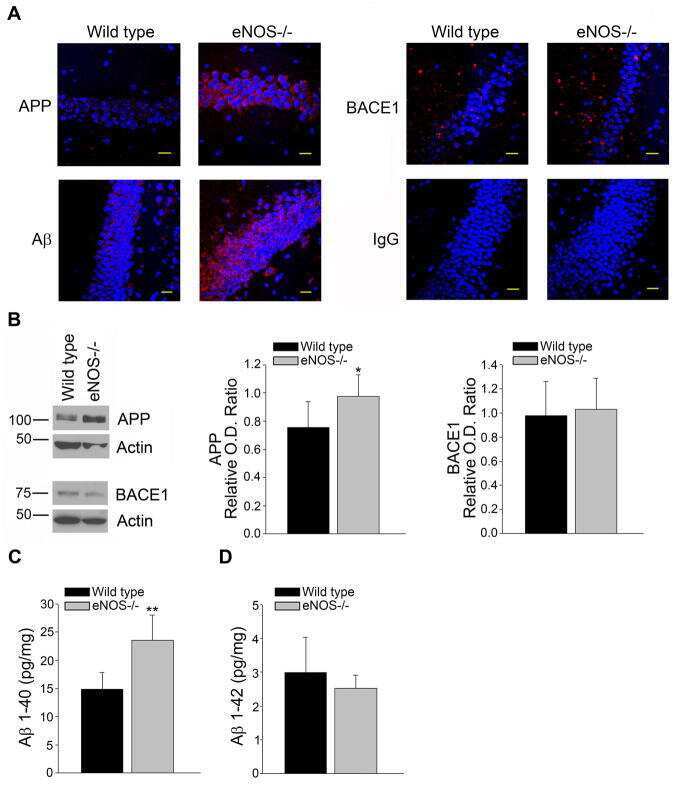
Endothelial nitric oxide deficiency promotes Alzheimer's disease pathology.
Clinically relevant intronic splicing enhancer mutation in myelin proteolipid protein leads to progressive microglia and astrocyte activation in white and gray matter regions of the brain.
Increased intracranial pressure after diffuse traumatic brain injury exacerbates neuronal somatic membrane poration but not axonal injury: evidence for primary intracranial pressure-induced neuronal perturbation.
Secondary damage caused by CD11b+ microglia following diffuse axonal injury in rats.
Internal jugular vein compression mitigates traumatic axonal injury in a rat model by reducing the intracranial slosh effect.
Intraneuronal Aβ detection in 5xFAD mice by a new Aβ-specific antibody.
Temporal assessment of traumatic axonal injury in the rat corpus callosum and optic chiasm.
Inhibition of JNK by a peptide inhibitor reduces traumatic brain injury-induced tauopathy in transgenic mice.
Small-molecule inducers of Aβ-42 peptide production share a common mechanism of action.
Reversal of fragile X phenotypes by manipulation of AβPP/Aβ levels in Fmr1KO mice.
A new model to produce sagittal plane rotational induced diffuse axonal injuries.
Quantitative relationship between axonal injury and mechanical response in a rodent head impact acceleration model.
Rodent model of direct cranial blast injury.
Rate of neurodegeneration in the mouse controlled cortical impact model is influenced by impactor tip shape: implications for mechanistic and therapeutic studies.
A mouse model of blast injury to brain: initial pathological, neuropathological, and behavioral characterization.
Omega-3 fatty acid supplementation and reduction of traumatic axonal injury in a rodent head injury model.
Human neural progenitors from different foetal forebrain regions remyelinate the adult mouse spinal cord.
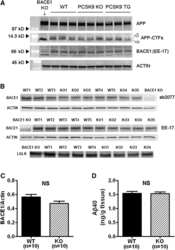
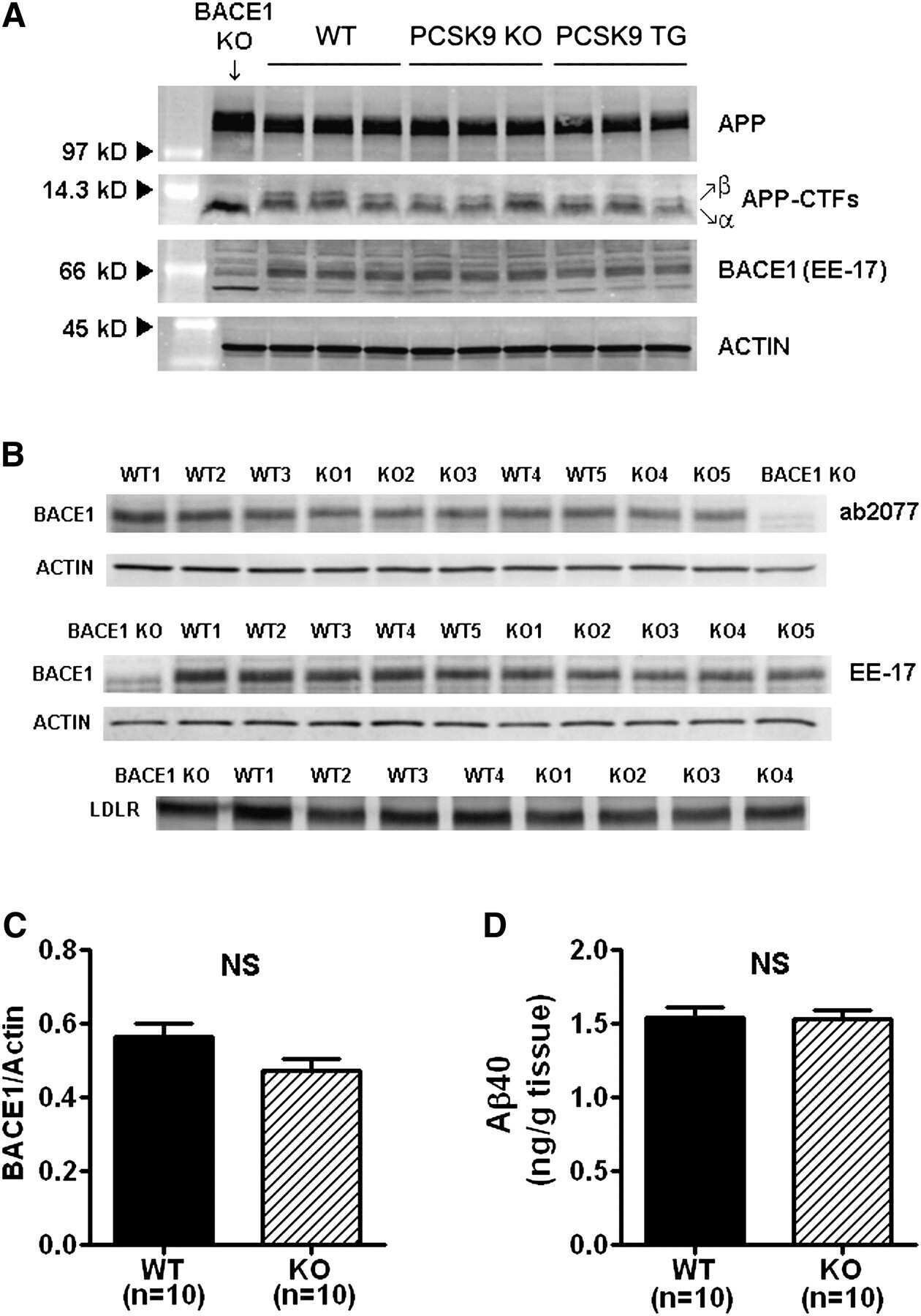
PCSK9 is not involved in the degradation of LDL receptors and BACE1 in the adult mouse brain.
Docosahexaenoic acid reduces traumatic axonal injury in a rodent head injury model.
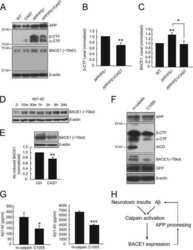
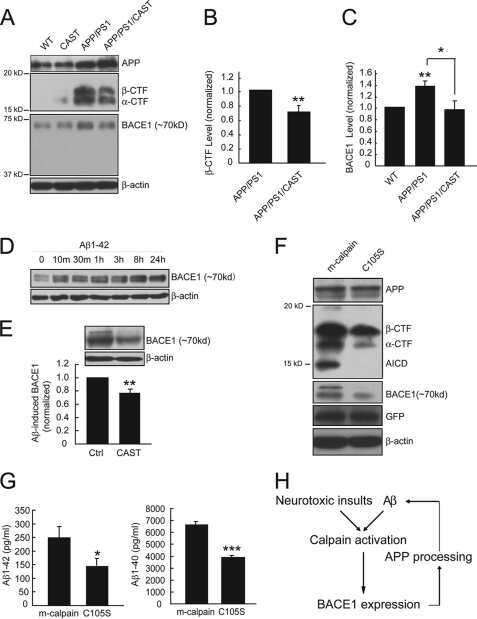
Calpain activation promotes BACE1 expression, amyloid precursor protein processing, and amyloid plaque formation in a transgenic mouse model of Alzheimer disease.
Blockade of acute microglial activation by minocycline promotes neuroprotection and reduces locomotor hyperactivity after closed head injury in mice: a twelve-week follow-up study.
SK-PC-B70M confers anti-oxidant activity and reduces Abeta levels in the brain of Tg2576 mice.
The cleavage products of amyloid-beta precursor protein are sorted to distinct carrier vesicles that are independently transported within neurites.
Behavioral stress accelerates plaque pathogenesis in the brain of Tg2576 mice via generation of metabolic oxidative stress.
Rck/p54 interacts with APP mRNA as part of a multi-protein complex and enhances APP mRNA and protein expression in neuronal cell lines.
Dual-specificity tyrosine(Y)-phosphorylation regulated kinase 1A-mediated phosphorylation of amyloid precursor protein: evidence for a functional link between Down syndrome and Alzheimer's disease.
Overexpression of ABCA1 reduces amyloid deposition in the PDAPP mouse model of Alzheimer disease.
Myopathy with tubulin-reactive inclusions in two cats.
Amyloid precursor protein and Presenilin1 interact with the adaptor GRB2 and modulate ERK 1,2 signaling.
FMRP mediates mGluR5-dependent translation of amyloid precursor protein.
Selective induction of ultrastructural (neurofilament) compaction in axons by means of a new head-injury apparatus.
Evolution of a neuroprotective function of central nervous system myelin.
Alzheimer precursor protein interaction with the Nogo-66 receptor reduces amyloid-beta plaque deposition.
Amyloid precursor protein modulates ERK-1 and -2 signaling.
Amyloid-beta protein precursor (AbetaPP) intracellular domain-associated protein-1 proteins bind to AbetaPP and modulate its processing in an isoform-specific manner.
The low density lipoprotein receptor-related protein 1B retains beta-amyloid precursor protein at the cell surface and reduces amyloid-beta peptide production.
Ginkgo biloba extract (Egb 761) inhibits beta-amyloid production by lowering free cholesterol levels.
A tissue level tolerance criterion for living brain developed with an in vitro model of traumatic mechanical loading.
Fast anterograde transport of herpes simplex virus: role for the amyloid precursor protein of alzheimer's disease.
Sporadic inclusion body myositis correlates with increased expression and cross-linking by transglutaminases 1 and 2.
Sporadic inclusion body myositis correlates with increased expression and cross-linking by transglutaminases 1 and 2.
Antibodies to the C-terminus of the beta-amyloid precursor protein (APP): a site specific marker for the detection of traumatic axonal injury.
Cai CZ, Zhuang XX, Zhu Q, Wu MY, Su H, Wang XJ, Iyaswamy A, Yue Z, Wang Q, Zhang B, Xue Y, Tan J, Li M, He H, Lu JH
Theranostics 2022;12(4):1738-1755
Theranostics 2022;12(4):1738-1755
Cholesterol biosynthesis defines oligodendrocyte precursor heterogeneity between brain and spinal cord.
Khandker L, Jeffries MA, Chang YJ, Mather ML, Evangelou AV, Bourne JN, Tafreshi AK, Ornelas IM, Bozdagi-Gunal O, Macklin WB, Wood TL
Cell reports 2022 Mar 1;38(9):110423
Cell reports 2022 Mar 1;38(9):110423
Malaria parasite heme biosynthesis promotes and griseofulvin protects against cerebral malaria in mice.
Chandana M, Anand A, Ghosh S, Das R, Beura S, Jena S, Suryawanshi AR, Padmanaban G, Nagaraj VA
Nature communications 2022 Jul 12;13(1):4028
Nature communications 2022 Jul 12;13(1):4028
Amelioration of Alzheimer's disease pathology by mitophagy inducers identified via machine learning and a cross-species workflow.
Xie C, Zhuang XX, Niu Z, Ai R, Lautrup S, Zheng S, Jiang Y, Han R, Gupta TS, Cao S, Lagartos-Donate MJ, Cai CZ, Xie LM, Caponio D, Wang WW, Schmauck-Medina T, Zhang J, Wang HL, Lou G, Xiao X, Zheng W, Palikaras K, Yang G, Caldwell KA, Caldwell GA, Shen HM, Nilsen H, Lu JH, Fang EF
Nature biomedical engineering 2022 Jan;6(1):76-93
Nature biomedical engineering 2022 Jan;6(1):76-93
Blast-induced axonal degeneration in the rat cerebellum in the absence of head movement.
Bishop R, Won SJ, Irvine KA, Basu J, Rome ES, Swanson RA
Scientific reports 2022 Jan 7;12(1):143
Scientific reports 2022 Jan 7;12(1):143
The role of aquaporin-4 in optic nerve head astrocytes in experimental glaucoma.
Kimball E, Schaub J, Quillen S, Keuthan C, Pease ME, Korneva A, Quigley H
PloS one 2021;16(2):e0244123
PloS one 2021;16(2):e0244123
Electroacupuncture ameliorates beta-amyloid pathology and cognitive impairment in Alzheimer disease via a novel mechanism involving activation of TFEB (transcription factor EB).
Zheng X, Lin W, Jiang Y, Lu K, Wei W, Huo Q, Cui S, Yang X, Li M, Xu N, Tang C, Song JX
Autophagy 2021 Nov;17(11):3833-3847
Autophagy 2021 Nov;17(11):3833-3847
Deletion of Abi3 gene locus exacerbates neuropathological features of Alzheimer's disease in a mouse model of Aβ amyloidosis.
Karahan H, Smith DC, Kim B, Dabin LC, Al-Amin MM, Wijeratne HRS, Pennington T, Viana di Prisco G, McCord B, Lin PB, Li Y, Peng J, Oblak AL, Chu S, Atwood BK, Kim J
Science advances 2021 Nov 5;7(45):eabe3954
Science advances 2021 Nov 5;7(45):eabe3954
NRBF2 is a RAB7 effector required for autophagosome maturation and mediates the association of APP-CTFs with active form of RAB7 for degradation.
Cai CZ, Yang C, Zhuang XX, Yuan NN, Wu MY, Tan JQ, Song JX, Cheung KH, Su H, Wang YT, Tang BS, Behrends C, Durairajan SSK, Yue Z, Li M, Lu JH
Autophagy 2021 May;17(5):1112-1130
Autophagy 2021 May;17(5):1112-1130
Genetic inactivation of SARM1 axon degeneration pathway improves outcome trajectory after experimental traumatic brain injury based on pathological, radiological, and functional measures.
Bradshaw DV Jr, Knutsen AK, Korotcov A, Sullivan GM, Radomski KL, Dardzinski BJ, Zi X, McDaniel DP, Armstrong RC
Acta neuropathologica communications 2021 May 17;9(1):89
Acta neuropathologica communications 2021 May 17;9(1):89
Mouse closed head traumatic brain injury replicates the histological tau pathology pattern of human disease: characterization of a novel model and systematic review of the literature.
Kahriman A, Bouley J, Smith TW, Bosco DA, Woerman AL, Henninger N
Acta neuropathologica communications 2021 Jun 29;9(1):118
Acta neuropathologica communications 2021 Jun 29;9(1):118
Protein farnesylation is upregulated in Alzheimer's human brains and neuron-specific suppression of farnesyltransferase mitigates pathogenic processes in Alzheimer's model mice.
Jeong A, Cheng S, Zhong R, Bennett DA, Bergö MO, Li L
Acta neuropathologica communications 2021 Jul 27;9(1):129
Acta neuropathologica communications 2021 Jul 27;9(1):129
Hyperoxygenation Treatment Reduces Beta-amyloid Deposition via MeCP2-dependent Upregulation of MMP-2 and MMP-9 in the Hippocampus of Tg-APP/PS1 Mice.
Choi J, Kwon H, Han PL
Experimental neurobiology 2021 Aug 31;30(4):294-307
Experimental neurobiology 2021 Aug 31;30(4):294-307
Endothelium-specific deletion of amyloid-β precursor protein exacerbates endothelial dysfunction induced by aging.
d'Uscio LV, Katusic ZS
Aging 2021 Aug 12;13(15):19165-19185
Aging 2021 Aug 12;13(15):19165-19185
Buprenorphine alters microglia and astrocytes acutely following diffuse traumatic brain injury.
Ryu J, Stone P, Lee S, Payne B, Gorse K, Lafrenaye A
Scientific reports 2021 Apr 21;11(1):8620
Scientific reports 2021 Apr 21;11(1):8620
Phosphoinositide 3-kinase inhibitor AS605240 ameliorates streptozotocin-induced Alzheimer's disease like sporadic dementia in experimental rats.
Alluri R, Ambati SR, Routhu K, Kopalli SR, Koppula S
EXCLI journal 2020;19:71-85
EXCLI journal 2020;19:71-85
Operation Brain Trauma Therapy: An Exploratory Study of Levetiracetam Treatment Following Mild Traumatic Brain Injury in the Micro Pig.
Lafrenaye A, Mondello S, Povlishock J, Gorse K, Walker S, Hayes R, Wang K, Kochanek PM
Frontiers in neurology 2020;11:586958
Frontiers in neurology 2020;11:586958
Sex-Specific Differences in Rodents Following a Single Primary Blast Exposure: Focus on the Monoamine and Galanin Systems.
Kawa L, Arborelius UP, Hökfelt T, Risling M
Frontiers in neurology 2020;11:540144
Frontiers in neurology 2020;11:540144
The endogenous neuronal complement inhibitor SRPX2 protects against complement-mediated synapse elimination during development.
Cong Q, Soteros BM, Wollet M, Kim JH, Sia GM
Nature neuroscience 2020 Sep;23(9):1067-1078
Nature neuroscience 2020 Sep;23(9):1067-1078
Systemic Exposure to Air Pollution Induces Oxidative Stress and Inflammation in Mouse Brain, Contributing to Neurodegeneration Onset.
Milani C, Farina F, Botto L, Massimino L, Lonati E, Donzelli E, Ballarini E, Crippa L, Marmiroli P, Bulbarelli A, Palestini P
International journal of molecular sciences 2020 May 24;21(10)
International journal of molecular sciences 2020 May 24;21(10)
The Proteasomal Deubiquitinating Enzyme PSMD14 Regulates Macroautophagy by Controlling Golgi-to-ER Retrograde Transport.
Bustamante HA, Cereceda K, González AE, Valenzuela GE, Cheuquemilla Y, Hernández S, Arias-Muñoz E, Cerda-Troncoso C, Bandau S, Soza A, Kausel G, Kerr B, Mardones GA, Cancino J, Hay RT, Rojas-Fernandez A, Burgos PV
Cells 2020 Mar 23;9(3)
Cells 2020 Mar 23;9(3)
Periventricular White Matter Alterations From Explosive Blast in a Large Animal Model: Mild Traumatic Brain Injury or "Subconcussive" Injury?
Kim JH, Goodrich JA, Situ R, Rapuano A, Hetherington H, Du F, Parks S, Taylor W, Westmoreland T, Ling G, Bandak FA, de Lanerolle NC
Journal of neuropathology and experimental neurology 2020 Jun 1;79(6):605-617
Journal of neuropathology and experimental neurology 2020 Jun 1;79(6):605-617
A method to quantify regional axonal transport blockade at the optic nerve head after short term intraocular pressure elevation in mice.
Korneva A, Schaub J, Jefferys J, Kimball E, Pease ME, Nawathe M, Johnson TV, Pitha I, Quigley H
Experimental eye research 2020 Jul;196:108035
Experimental eye research 2020 Jul;196:108035
Long-term cognitive impairment without diffuse axonal injury following repetitive mild traumatic brain injury in rats.
Tadepalli SA, Bali ZK, Bruszt N, Nagy LV, Amrein K, Fazekas B, Büki A, Czeiter E, Hernádi I
Behavioural brain research 2020 Jan 27;378:112268
Behavioural brain research 2020 Jan 27;378:112268
A small molecule transcription factor EB activator ameliorates beta-amyloid precursor protein and Tau pathology in Alzheimer's disease models.
Song JX, Malampati S, Zeng Y, Durairajan SSK, Yang CB, Tong BC, Iyaswamy A, Shang WB, Sreenivasmurthy SG, Zhu Z, Cheung KH, Lu JH, Tang C, Xu N, Li M
Aging cell 2020 Feb;19(2):e13069
Aging cell 2020 Feb;19(2):e13069
Diffuse Axonal Injury in the Rat Brain: Axonal Injury and Oligodendrocyte Activity Following Rotational Injury.
Losurdo M, Davidsson J, Sköld MK
Brain sciences 2020 Apr 10;10(4)
Brain sciences 2020 Apr 10;10(4)
Repetitive Concussive and Subconcussive Injury in a Human Tau Mouse Model Results in Chronic Cognitive Dysfunction and Disruption of White Matter Tracts, But Not Tau Pathology.
Gangolli M, Benetatos J, Esparza TJ, Fountain EM, Seneviratne S, Brody DL
Journal of neurotrauma 2019 Mar 1;36(5):735-755
Journal of neurotrauma 2019 Mar 1;36(5):735-755
Mechanical Stretch of High Magnitude Provokes Axonal Injury, Elongation of Paranodal Junctions, and Signaling Alterations in Oligodendrocytes.
Chierto E, Simon A, Castoldi F, Meffre D, Cristinziano G, Sapone F, Carrete A, Borderie D, Etienne F, Rannou F, Morrison B 3rd, Massaad C, Jafarian-Tehrani M
Molecular neurobiology 2019 Jun;56(6):4231-4248
Molecular neurobiology 2019 Jun;56(6):4231-4248
Vascular phenotype of amyloid precursor protein-deficient mice.
d'Uscio LV, Katusic ZS
American journal of physiology. Heart and circulatory physiology 2019 Jun 1;316(6):H1297-H1308
American journal of physiology. Heart and circulatory physiology 2019 Jun 1;316(6):H1297-H1308
EPPS treatment attenuates traumatic brain injury in mice by reducing Aβ burden and ameliorating neuronal autophagic flux.
Anthony Jalin AMA, Jin R, Wang M, Li G
Experimental neurology 2019 Apr;314:20-33
Experimental neurology 2019 Apr;314:20-33
The Importance of Inter-Species Variation in Traumatic Brain Injury-Induced Alterations of Microglial-Axonal Interactions.
Gorse KM, Lafrenaye AD
Frontiers in neurology 2018;9:778
Frontiers in neurology 2018;9:778
A Comparative Study of Two Blast-Induced Traumatic Brain Injury Models: Changes in Monoamine and Galanin Systems Following Single and Repeated Exposure.
Kawa L, Kamnaksh A, Long JB, Arborelius UP, Hökfelt T, Agoston DV, Risling M
Frontiers in neurology 2018;9:479
Frontiers in neurology 2018;9:479
A Mild Traumatic Brain Injury in Mice Produces Lasting Deficits in Brain Metabolism.
Lyons DN, Vekaria H, Macheda T, Bakshi V, Powell DK, Gold BT, Lin AL, Sullivan PG, Bachstetter AD
Journal of neurotrauma 2018 Oct 15;35(20):2435-2447
Journal of neurotrauma 2018 Oct 15;35(20):2435-2447
modCHIMERA: a novel murine closed-head model of moderate traumatic brain injury.
Sauerbeck AD, Fanizzi C, Kim JH, Gangolli M, Bayly PV, Wellington CL, Brody DL, Kummer TT
Scientific reports 2018 May 16;8(1):7677
Scientific reports 2018 May 16;8(1):7677
Mild Traumatic Brain Injury Induces Structural and Functional Disconnection of Local Neocortical Inhibitory Networks via Parvalbumin Interneuron Diffuse Axonal Injury.
Vascak M, Jin X, Jacobs KM, Povlishock JT
Cerebral cortex (New York, N.Y. : 1991) 2018 May 1;28(5):1625-1644
Cerebral cortex (New York, N.Y. : 1991) 2018 May 1;28(5):1625-1644
Wnt signaling loss accelerates the appearance of neuropathological hallmarks of Alzheimer's disease in J20-APP transgenic and wild-type mice.
Tapia-Rojas C, Inestrosa NC
Journal of neurochemistry 2018 Feb;144(4):443-465
Journal of neurochemistry 2018 Feb;144(4):443-465
Matrix-Assisted Laser Desorption Ionization Mapping of Lysophosphatidic Acid Changes after Traumatic Brain Injury and the Relationship to Cellular Pathology.
McDonald WS, Jones EE, Wojciak JM, Drake RR, Sabbadini RA, Harris NG
The American journal of pathology 2018 Aug;188(8):1779-1793
The American journal of pathology 2018 Aug;188(8):1779-1793
Primary Traumatic Axonopathy in Mice Subjected to Impact Acceleration: A Reappraisal of Pathology and Mechanisms with High-Resolution Anatomical Methods.
Ziogas NK, Koliatsos VE
The Journal of neuroscience : the official journal of the Society for Neuroscience 2018 Apr 18;38(16):4031-4047
The Journal of neuroscience : the official journal of the Society for Neuroscience 2018 Apr 18;38(16):4031-4047
NRBF2 is involved in the autophagic degradation process of APP-CTFs in Alzheimer disease models.
Yang C, Cai CZ, Song JX, Tan JQ, Durairajan SSK, Iyaswamy A, Wu MY, Chen LL, Yue Z, Li M, Lu JH
Autophagy 2017;13(12):2028-2040
Autophagy 2017;13(12):2028-2040
Neuronal Injury and Glial Changes Are Hallmarks of Open Field Blast Exposure in Swine Frontal Lobe.
Kallakuri S, Desai A, Feng K, Tummala S, Saif T, Chen C, Zhang L, Cavanaugh JM, King AI
PloS one 2017;12(1):e0169239
PloS one 2017;12(1):e0169239
Defining an Analytic Framework to Evaluate Quantitative MRI Markers of Traumatic Axonal Injury: Preliminary Results in a Mouse Closed Head Injury Model.
Haber M, Hutchinson EB, Sadeghi N, Cheng WH, Namjoshi D, Cripton P, Irfanoglu MO, Wellington C, Diaz-Arrastia R, Pierpaoli C
eNeuro 2017 Sep-Oct;4(5)
eNeuro 2017 Sep-Oct;4(5)
The connectomics of brain demyelination: Functional and structural patterns in the cuprizone mouse model.
Hübner NS, Mechling AE, Lee HL, Reisert M, Bienert T, Hennig J, von Elverfeldt D, Harsan LA
NeuroImage 2017 Feb 1;146:1-18
NeuroImage 2017 Feb 1;146:1-18
The Amyloid Precursor Protein of Alzheimer's Disease Clusters at the Organelle/Microtubule Interface on Organelles that Bind Microtubules in an ATP Dependent Manner.
Stevenson JW, Conaty EA, Walsh RB, Poidomani PJ, Samoriski CM, Scollins BJ, DeGiorgis JA
PloS one 2016;11(1):e0147808
PloS one 2016;11(1):e0147808
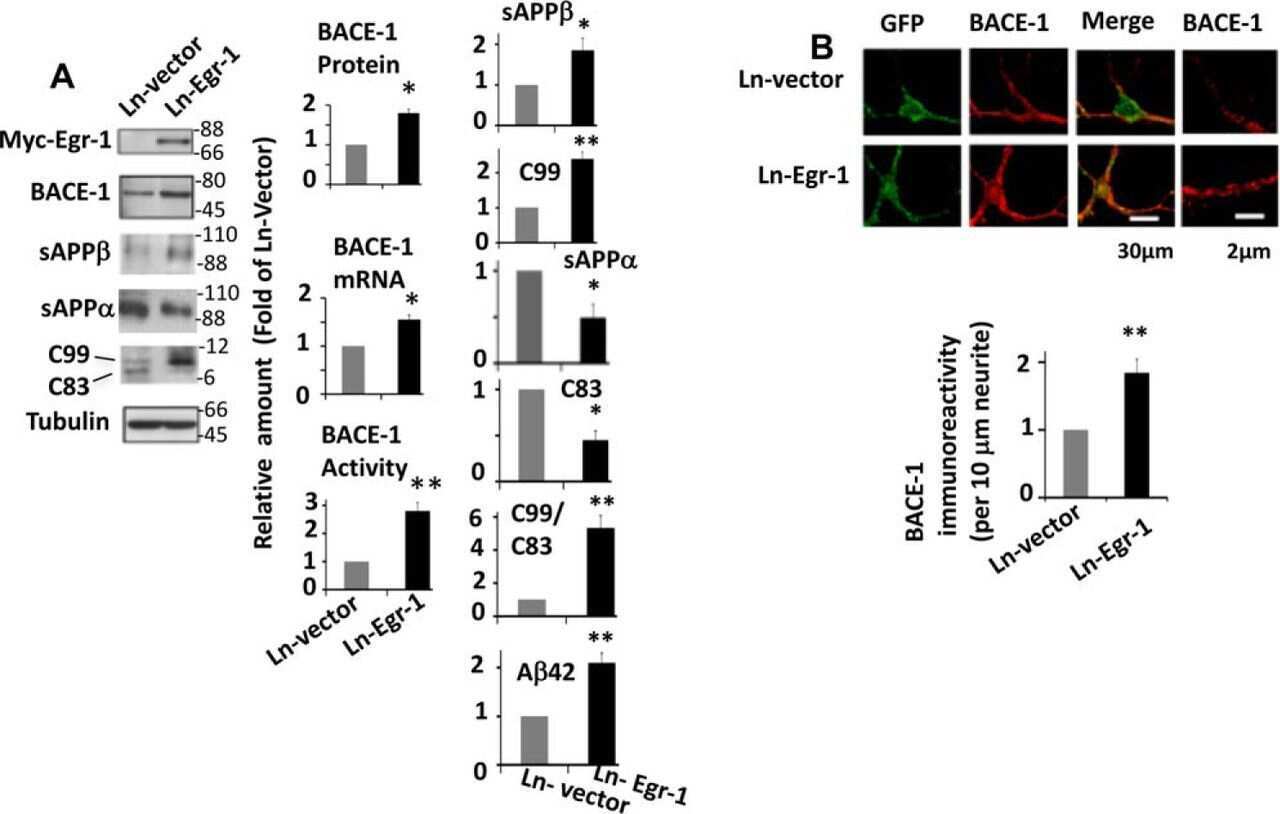
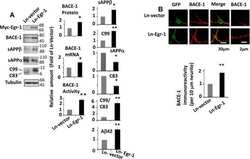
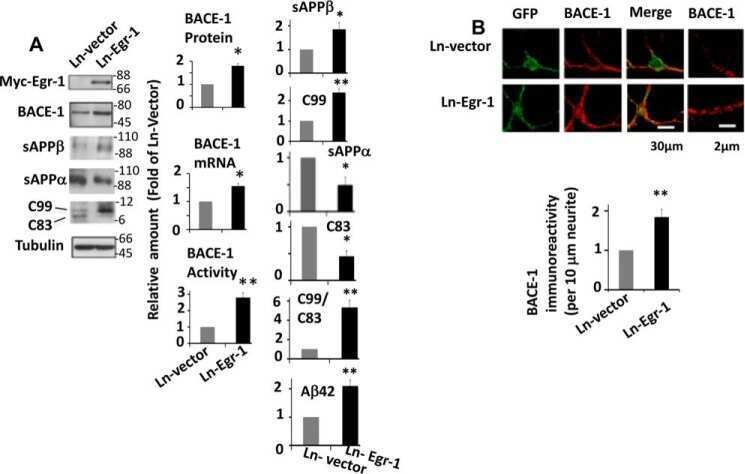
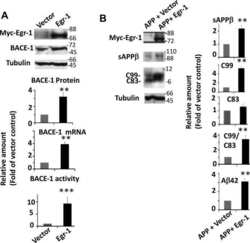
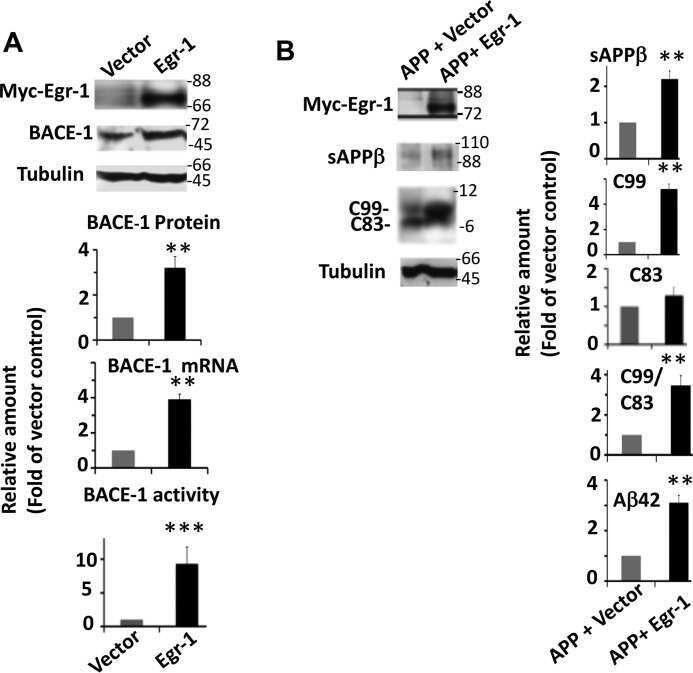
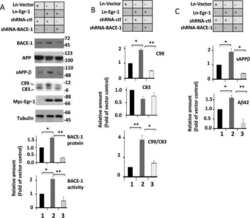
Early Growth Response 1 (Egr-1) Is a Transcriptional Activator of β-Secretase 1 (BACE-1) in the Brain.
Qin X, Wang Y, Paudel HK
The Journal of biological chemistry 2016 Oct 14;291(42):22276-22287
The Journal of biological chemistry 2016 Oct 14;291(42):22276-22287
Inhibition of Wnt signaling induces amyloidogenic processing of amyloid precursor protein and the production and aggregation of Amyloid-β (Aβ)(42) peptides.
Tapia-Rojas C, Burgos PV, Inestrosa NC
Journal of neurochemistry 2016 Dec;139(6):1175-1191
Journal of neurochemistry 2016 Dec;139(6):1175-1191
Tetrahydrohyperforin Inhibits the Proteolytic Processing of Amyloid Precursor Protein and Enhances Its Degradation by Atg5-Dependent Autophagy.
Cavieres VA, González A, Muñoz VC, Yefi CP, Bustamante HA, Barraza RR, Tapia-Rojas C, Otth C, Barrera MJ, González C, Mardones GA, Inestrosa NC, Burgos PV
PloS one 2015;10(8):e0136313
PloS one 2015;10(8):e0136313
Losartan Treatment Protects Retinal Ganglion Cells and Alters Scleral Remodeling in Experimental Glaucoma.
Quigley HA, Pitha IF, Welsbie DS, Nguyen C, Steinhart MR, Nguyen TD, Pease ME, Oglesby EN, Berlinicke CA, Mitchell KL, Kim J, Jefferys JJ, Kimball EC
PloS one 2015;10(10):e0141137
PloS one 2015;10(10):e0141137
A novel closed-body model of spinal cord injury caused by high-pressure air blasts produces extensive axonal injury and motor impairments.
del Mar N, von Buttlar X, Yu AS, Guley NH, Reiner A, Honig MG
Experimental neurology 2015 Sep;271:53-71
Experimental neurology 2015 Sep;271:53-71
Kalirin-9 and Kalirin-12 Play Essential Roles in Dendritic Outgrowth and Branching.
Yan Y, Eipper BA, Mains RE
Cerebral cortex (New York, N.Y. : 1991) 2015 Oct;25(10):3487-501
Cerebral cortex (New York, N.Y. : 1991) 2015 Oct;25(10):3487-501
Blast overpressure induced axonal injury changes in rat brainstem and spinal cord.
Kallakuri S, Purkait HS, Dalavayi S, VandeVord P, Cavanaugh JM
Journal of neurosciences in rural practice 2015 Oct-Dec;6(4):481-7
Journal of neurosciences in rural practice 2015 Oct-Dec;6(4):481-7
Microglia processes associate with diffusely injured axons following mild traumatic brain injury in the micro pig.
Lafrenaye AD, Todani M, Walker SA, Povlishock JT
Journal of neuroinflammation 2015 Oct 6;12:186
Journal of neuroinflammation 2015 Oct 6;12:186
TREM2 regulates microglial cell activation in response to demyelination in vivo.
Cantoni C, Bollman B, Licastro D, Xie M, Mikesell R, Schmidt R, Yuede CM, Galimberti D, Olivecrona G, Klein RS, Cross AH, Otero K, Piccio L
Acta neuropathologica 2015 Mar;129(3):429-47
Acta neuropathologica 2015 Mar;129(3):429-47
Traumatic brain injury-induced axonal phenotypes react differently to treatment.
Hånell A, Greer JE, McGinn MJ, Povlishock JT
Acta neuropathologica 2015 Feb;129(2):317-32
Acta neuropathologica 2015 Feb;129(2):317-32
Ccr2 deletion dissociates cavity size and tau pathology after mild traumatic brain injury.
Gyoneva S, Kim D, Katsumoto A, Kokiko-Cochran ON, Lamb BT, Ransohoff RM
Journal of neuroinflammation 2015 Dec 3;12:228
Journal of neuroinflammation 2015 Dec 3;12:228
Defects of Lipid Synthesis Are Linked to the Age-Dependent Demyelination Caused by Lamin B1 Overexpression.
Rolyan H, Tyurina YY, Hernandez M, Amoscato AA, Sparvero LJ, Nmezi BC, Lu Y, Estécio MR, Lin K, Chen J, He RR, Gong P, Rigatti LH, Dupree J, Bayır H, Kagan VE, Casaccia P, Padiath QS
The Journal of neuroscience : the official journal of the Society for Neuroscience 2015 Aug 26;35(34):12002-17
The Journal of neuroscience : the official journal of the Society for Neuroscience 2015 Aug 26;35(34):12002-17
Neurotransmitter Systems in a Mild Blast Traumatic Brain Injury Model: Catecholamines and Serotonin.
Kawa L, Arborelius UP, Yoshitake T, Kehr J, Hökfelt T, Risling M, Agoston D
Journal of neurotrauma 2015 Aug 15;32(16):1190-9
Journal of neurotrauma 2015 Aug 15;32(16):1190-9
Array tomography for the detection of non-dilated, injured axons in traumatic brain injury.
Bennett RE, Brody DL
Journal of neuroscience methods 2015 Apr 30;245:25-36
Journal of neuroscience methods 2015 Apr 30;245:25-36
Fragile X mental retardation protein expression in Alzheimer's disease.
Renoux AJ, Carducci NM, Ahmady AA, Todd PK
Frontiers in genetics 2014;5:360
Frontiers in genetics 2014;5:360
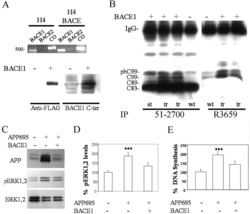
Identification and Preclinical Pharmacology of the γ-Secretase Modulator BMS-869780.
Toyn JH, Thompson LA, Lentz KA, Meredith JE Jr, Burton CR, Sankaranararyanan S, Guss V, Hall T, Iben LG, Krause CM, Krause R, Lin XA, Pierdomenico M, Polson C, Robertson AS, Denton RR, Grace JE, Morrison J, Raybon J, Zhuo X, Snow K, Padmanabha R, Agler M, Esposito K, Harden D, Prack M, Varma S, Wong V, Zhu Y, Zvyaga T, Gerritz S, Marcin LR, Higgins MA, Shi J, Wei C, Cantone JL, Drexler DM, Macor JE, Olson RE, Ahlijanian MK, Albright CF
International journal of Alzheimer's disease 2014;2014:431858
International journal of Alzheimer's disease 2014;2014:431858
Moderately elevated intracranial pressure after diffuse traumatic brain injury is associated with exacerbated neuronal pathology and behavioral morbidity in the rat.
Lafrenaye AD, Krahe TE, Povlishock JT
Journal of cerebral blood flow and metabolism : official journal of the International Society of Cerebral Blood Flow and Metabolism 2014 Oct;34(10):1628-36
Journal of cerebral blood flow and metabolism : official journal of the International Society of Cerebral Blood Flow and Metabolism 2014 Oct;34(10):1628-36
Acute reduction of microglia does not alter axonal injury in a mouse model of repetitive concussive traumatic brain injury.
Bennett RE, Brody DL
Journal of neurotrauma 2014 Oct 1;31(19):1647-63
Journal of neurotrauma 2014 Oct 1;31(19):1647-63
Experimental traumatic brain injury induces rapid aggregation and oligomerization of amyloid-beta in an Alzheimer's disease mouse model.
Washington PM, Morffy N, Parsadanian M, Zapple DN, Burns MP
Journal of neurotrauma 2014 Jan 1;31(1):125-34
Journal of neurotrauma 2014 Jan 1;31(1):125-34
Cortical hypoexcitation defines neuronal responses in the immediate aftermath of traumatic brain injury.
Johnstone VP, Yan EB, Alwis DS, Rajan R
PloS one 2013;8(5):e63454
PloS one 2013;8(5):e63454
AAD-2004 Attenuates Progressive Neuronal Loss in the Brain of Tg-betaCTF99/B6 Mouse Model of Alzheimer Disease.
Baek IS, Kim TK, Seo JS, Lee KW, Lee YA, Cho J, Gwag BJ, Han PL
Experimental neurobiology 2013 Mar;22(1):31-7
Experimental neurobiology 2013 Mar;22(1):31-7
Myelin loss and oligodendrocyte pathology in white matter tracts following traumatic brain injury in the rat.
Flygt J, Djupsjö A, Lenne F, Marklund N
The European journal of neuroscience 2013 Jul;38(1):2153-65
The European journal of neuroscience 2013 Jul;38(1):2153-65
Endothelial nitric oxide deficiency promotes Alzheimer's disease pathology.
Austin SA, Santhanam AV, Hinton DJ, Choi DS, Katusic ZS
Journal of neurochemistry 2013 Dec;127(5):691-700
Journal of neurochemistry 2013 Dec;127(5):691-700
Clinically relevant intronic splicing enhancer mutation in myelin proteolipid protein leads to progressive microglia and astrocyte activation in white and gray matter regions of the brain.
Bachstetter AD, Webster SJ, Van Eldik LJ, Cambi F
Journal of neuroinflammation 2013 Dec 5;10:146
Journal of neuroinflammation 2013 Dec 5;10:146
Increased intracranial pressure after diffuse traumatic brain injury exacerbates neuronal somatic membrane poration but not axonal injury: evidence for primary intracranial pressure-induced neuronal perturbation.
Lafrenaye AD, McGinn MJ, Povlishock JT
Journal of cerebral blood flow and metabolism : official journal of the International Society of Cerebral Blood Flow and Metabolism 2012 Oct;32(10):1919-32
Journal of cerebral blood flow and metabolism : official journal of the International Society of Cerebral Blood Flow and Metabolism 2012 Oct;32(10):1919-32
Secondary damage caused by CD11b+ microglia following diffuse axonal injury in rats.
Jia X, Cong B, Wang S, Dong L, Ma C, Li Y
The journal of trauma and acute care surgery 2012 Nov;73(5):1168-74
The journal of trauma and acute care surgery 2012 Nov;73(5):1168-74
Internal jugular vein compression mitigates traumatic axonal injury in a rat model by reducing the intracranial slosh effect.
Smith DW, Bailes JE, Fisher JA, Robles J, Turner RC, Mills JD
Neurosurgery 2012 Mar;70(3):740-6
Neurosurgery 2012 Mar;70(3):740-6
Intraneuronal Aβ detection in 5xFAD mice by a new Aβ-specific antibody.
Youmans KL, Tai LM, Kanekiyo T, Stine WB Jr, Michon SC, Nwabuisi-Heath E, Manelli AM, Fu Y, Riordan S, Eimer WA, Binder L, Bu G, Yu C, Hartley DM, LaDu MJ
Molecular neurodegeneration 2012 Mar 16;7:8
Molecular neurodegeneration 2012 Mar 16;7:8
Temporal assessment of traumatic axonal injury in the rat corpus callosum and optic chiasm.
Zakaria N, Kallakuri S, Bandaru S, Cavanaugh JM
Brain research 2012 Jul 27;1467:81-90
Brain research 2012 Jul 27;1467:81-90
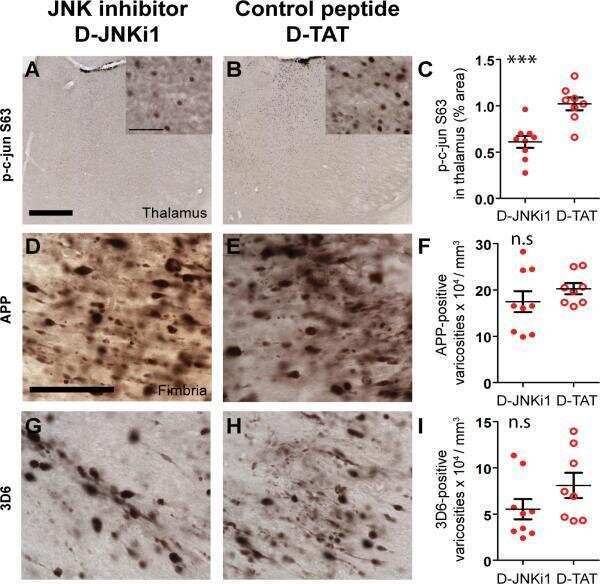
Inhibition of JNK by a peptide inhibitor reduces traumatic brain injury-induced tauopathy in transgenic mice.
Tran HT, Sanchez L, Brody DL
Journal of neuropathology and experimental neurology 2012 Feb;71(2):116-29
Journal of neuropathology and experimental neurology 2012 Feb;71(2):116-29
Small-molecule inducers of Aβ-42 peptide production share a common mechanism of action.
Bettayeb K, Oumata N, Zhang Y, Luo W, Bustos V, Galons H, Greengard P, Meijer L, Flajolet M
FASEB journal : official publication of the Federation of American Societies for Experimental Biology 2012 Dec;26(12):5115-23
FASEB journal : official publication of the Federation of American Societies for Experimental Biology 2012 Dec;26(12):5115-23
Reversal of fragile X phenotypes by manipulation of AβPP/Aβ levels in Fmr1KO mice.
Westmark CJ, Westmark PR, O'Riordan KJ, Ray BC, Hervey CM, Salamat MS, Abozeid SH, Stein KM, Stodola LA, Tranfaglia M, Burger C, Berry-Kravis EM, Malter JS
PloS one 2011;6(10):e26549
PloS one 2011;6(10):e26549
A new model to produce sagittal plane rotational induced diffuse axonal injuries.
Davidsson J, Risling M
Frontiers in neurology 2011;2:41
Frontiers in neurology 2011;2:41
Quantitative relationship between axonal injury and mechanical response in a rodent head impact acceleration model.
Li Y, Zhang L, Kallakuri S, Zhou R, Cavanaugh JM
Journal of neurotrauma 2011 Sep;28(9):1767-82
Journal of neurotrauma 2011 Sep;28(9):1767-82
Rodent model of direct cranial blast injury.
Kuehn R, Simard PF, Driscoll I, Keledjian K, Ivanova S, Tosun C, Williams A, Bochicchio G, Gerzanich V, Simard JM
Journal of neurotrauma 2011 Oct;28(10):2155-69
Journal of neurotrauma 2011 Oct;28(10):2155-69
Rate of neurodegeneration in the mouse controlled cortical impact model is influenced by impactor tip shape: implications for mechanistic and therapeutic studies.
Pleasant JM, Carlson SW, Mao H, Scheff SW, Yang KH, Saatman KE
Journal of neurotrauma 2011 Nov;28(11):2245-62
Journal of neurotrauma 2011 Nov;28(11):2245-62
A mouse model of blast injury to brain: initial pathological, neuropathological, and behavioral characterization.
Koliatsos VE, Cernak I, Xu L, Song Y, Savonenko A, Crain BJ, Eberhart CG, Frangakis CE, Melnikova T, Kim H, Lee D
Journal of neuropathology and experimental neurology 2011 May;70(5):399-416
Journal of neuropathology and experimental neurology 2011 May;70(5):399-416
Omega-3 fatty acid supplementation and reduction of traumatic axonal injury in a rodent head injury model.
Mills JD, Bailes JE, Sedney CL, Hutchins H, Sears B
Journal of neurosurgery 2011 Jan;114(1):77-84
Journal of neurosurgery 2011 Jan;114(1):77-84
Human neural progenitors from different foetal forebrain regions remyelinate the adult mouse spinal cord.
Buchet D, Garcia C, Deboux C, Nait-Oumesmar B, Baron-Van Evercooren A
Brain : a journal of neurology 2011 Apr;134(Pt 4):1168-83
Brain : a journal of neurology 2011 Apr;134(Pt 4):1168-83
PCSK9 is not involved in the degradation of LDL receptors and BACE1 in the adult mouse brain.
Liu M, Wu G, Baysarowich J, Kavana M, Addona GH, Bierilo KK, Mudgett JS, Pavlovic G, Sitlani A, Renger JJ, Hubbard BK, Fisher TS, Zerbinatti CV
Journal of lipid research 2010 Sep;51(9):2611-8
Journal of lipid research 2010 Sep;51(9):2611-8
Docosahexaenoic acid reduces traumatic axonal injury in a rodent head injury model.
Bailes JE, Mills JD
Journal of neurotrauma 2010 Sep;27(9):1617-24
Journal of neurotrauma 2010 Sep;27(9):1617-24
Calpain activation promotes BACE1 expression, amyloid precursor protein processing, and amyloid plaque formation in a transgenic mouse model of Alzheimer disease.
Liang B, Duan BY, Zhou XP, Gong JX, Luo ZG
The Journal of biological chemistry 2010 Sep 3;285(36):27737-44
The Journal of biological chemistry 2010 Sep 3;285(36):27737-44
Blockade of acute microglial activation by minocycline promotes neuroprotection and reduces locomotor hyperactivity after closed head injury in mice: a twelve-week follow-up study.
Homsi S, Piaggio T, Croci N, Noble F, Plotkine M, Marchand-Leroux C, Jafarian-Tehrani M
Journal of neurotrauma 2010 May;27(5):911-21
Journal of neurotrauma 2010 May;27(5):911-21
SK-PC-B70M confers anti-oxidant activity and reduces Abeta levels in the brain of Tg2576 mice.
Seo JS, Kim TK, Leem YH, Lee KW, Park SK, Baek IS, Kim KS, Im GJ, Lee SM, Park YH, Han PL
Brain research 2009 Mar 19;1261:100-8
Brain research 2009 Mar 19;1261:100-8
The cleavage products of amyloid-beta precursor protein are sorted to distinct carrier vesicles that are independently transported within neurites.
Muresan V, Varvel NH, Lamb BT, Muresan Z
The Journal of neuroscience : the official journal of the Society for Neuroscience 2009 Mar 18;29(11):3565-78
The Journal of neuroscience : the official journal of the Society for Neuroscience 2009 Mar 18;29(11):3565-78
Behavioral stress accelerates plaque pathogenesis in the brain of Tg2576 mice via generation of metabolic oxidative stress.
Lee KW, Kim JB, Seo JS, Kim TK, Im JY, Baek IS, Kim KS, Lee JK, Han PL
Journal of neurochemistry 2009 Jan;108(1):165-75
Journal of neurochemistry 2009 Jan;108(1):165-75
Rck/p54 interacts with APP mRNA as part of a multi-protein complex and enhances APP mRNA and protein expression in neuronal cell lines.
Broytman O, Westmark PR, Gurel Z, Malter JS
Neurobiology of aging 2009 Dec;30(12):1962-74
Neurobiology of aging 2009 Dec;30(12):1962-74
Dual-specificity tyrosine(Y)-phosphorylation regulated kinase 1A-mediated phosphorylation of amyloid precursor protein: evidence for a functional link between Down syndrome and Alzheimer's disease.
Ryoo SR, Cho HJ, Lee HW, Jeong HK, Radnaabazar C, Kim YS, Kim MJ, Son MY, Seo H, Chung SH, Song WJ
Journal of neurochemistry 2008 Mar;104(5):1333-44
Journal of neurochemistry 2008 Mar;104(5):1333-44
Overexpression of ABCA1 reduces amyloid deposition in the PDAPP mouse model of Alzheimer disease.
Wahrle SE, Jiang H, Parsadanian M, Kim J, Li A, Knoten A, Jain S, Hirsch-Reinshagen V, Wellington CL, Bales KR, Paul SM, Holtzman DM
The Journal of clinical investigation 2008 Feb;118(2):671-82
The Journal of clinical investigation 2008 Feb;118(2):671-82
Myopathy with tubulin-reactive inclusions in two cats.
Shelton GD, Sturges BK, Lyons LA, Williams DC, Aleman M, Jiang Y, Mizisin AP
Acta neuropathologica 2007 Nov;114(5):537-42
Acta neuropathologica 2007 Nov;114(5):537-42
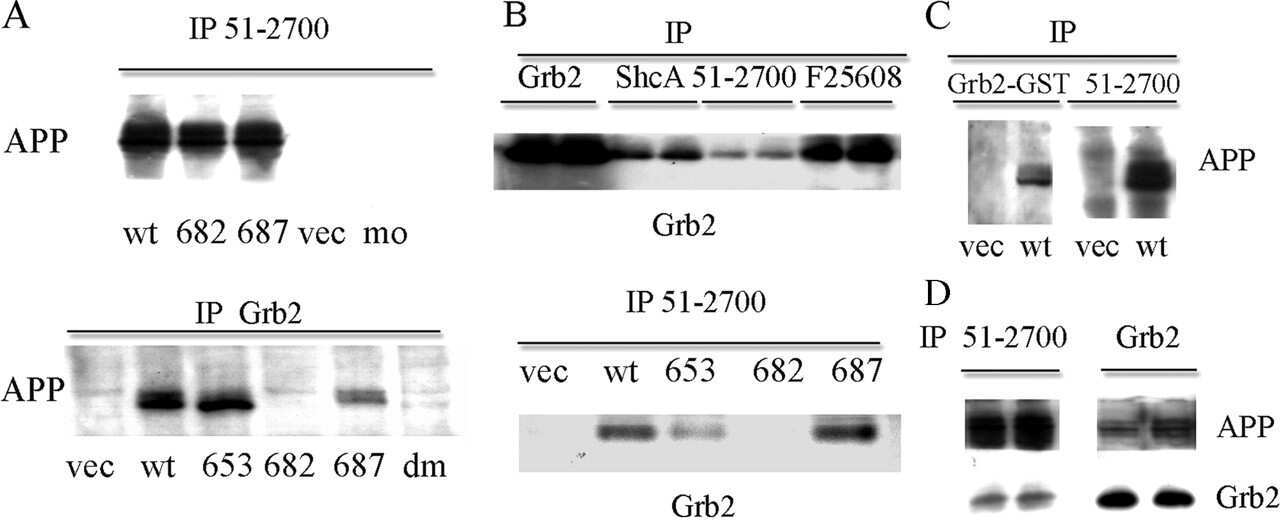
Amyloid precursor protein and Presenilin1 interact with the adaptor GRB2 and modulate ERK 1,2 signaling.
Nizzari M, Venezia V, Repetto E, Caorsi V, Magrassi R, Gagliani MC, Carlo P, Florio T, Schettini G, Tacchetti C, Russo T, Diaspro A, Russo C
The Journal of biological chemistry 2007 May 4;282(18):13833-44
The Journal of biological chemistry 2007 May 4;282(18):13833-44
FMRP mediates mGluR5-dependent translation of amyloid precursor protein.
Westmark CJ, Malter JS
PLoS biology 2007 Mar;5(3):e52
PLoS biology 2007 Mar;5(3):e52
Selective induction of ultrastructural (neurofilament) compaction in axons by means of a new head-injury apparatus.
Pál J, Tóth Z, Farkas O, Kellényi L, Dóczi T, Gallyas F
Journal of neuroscience methods 2006 Jun 15;153(2):283-9
Journal of neuroscience methods 2006 Jun 15;153(2):283-9
Evolution of a neuroprotective function of central nervous system myelin.
Yin X, Baek RC, Kirschner DA, Peterson A, Fujii Y, Nave KA, Macklin WB, Trapp BD
The Journal of cell biology 2006 Jan 30;172(3):469-78
The Journal of cell biology 2006 Jan 30;172(3):469-78
Alzheimer precursor protein interaction with the Nogo-66 receptor reduces amyloid-beta plaque deposition.
Park JH, Gimbel DA, GrandPre T, Lee JK, Kim JE, Li W, Lee DH, Strittmatter SM
The Journal of neuroscience : the official journal of the Society for Neuroscience 2006 Feb 1;26(5):1386-95
The Journal of neuroscience : the official journal of the Society for Neuroscience 2006 Feb 1;26(5):1386-95
Amyloid precursor protein modulates ERK-1 and -2 signaling.
Venezia V, Nizzari M, Repetto E, Violani E, Corsaro A, Thellung S, Villa V, Carlo P, Schettini G, Florio T, Russo C
Annals of the New York Academy of Sciences 2006 Dec;1090:455-65
Annals of the New York Academy of Sciences 2006 Dec;1090:455-65
Amyloid-beta protein precursor (AbetaPP) intracellular domain-associated protein-1 proteins bind to AbetaPP and modulate its processing in an isoform-specific manner.
Ghersi E, Noviello C, D'Adamio L
The Journal of biological chemistry 2004 Nov 19;279(47):49105-12
The Journal of biological chemistry 2004 Nov 19;279(47):49105-12
The low density lipoprotein receptor-related protein 1B retains beta-amyloid precursor protein at the cell surface and reduces amyloid-beta peptide production.
Cam JA, Zerbinatti CV, Knisely JM, Hecimovic S, Li Y, Bu G
The Journal of biological chemistry 2004 Jul 9;279(28):29639-46
The Journal of biological chemistry 2004 Jul 9;279(28):29639-46
Ginkgo biloba extract (Egb 761) inhibits beta-amyloid production by lowering free cholesterol levels.
Yao ZX, Han Z, Drieu K, Papadopoulos V
The Journal of nutritional biochemistry 2004 Dec;15(12):749-56
The Journal of nutritional biochemistry 2004 Dec;15(12):749-56
A tissue level tolerance criterion for living brain developed with an in vitro model of traumatic mechanical loading.
Morrison B 3rd, Cater HL, Wang CC, Thomas FC, Hung CT, Ateshian GA, Sundstrom LE
Stapp car crash journal 2003 Oct;47:93-105
Stapp car crash journal 2003 Oct;47:93-105
Fast anterograde transport of herpes simplex virus: role for the amyloid precursor protein of alzheimer's disease.
Satpute-Krishnan P, DeGiorgis JA, Bearer EL
Aging cell 2003 Dec;2(6):305-18
Aging cell 2003 Dec;2(6):305-18
Sporadic inclusion body myositis correlates with increased expression and cross-linking by transglutaminases 1 and 2.
Choi YC, Park GT, Kim TS, Sunwoo IN, Steinert PM, Kim SY
The Journal of biological chemistry 2000 Mar 24;275(12):8703-10
The Journal of biological chemistry 2000 Mar 24;275(12):8703-10
Sporadic inclusion body myositis correlates with increased expression and cross-linking by transglutaminases 1 and 2.
Choi YC, Park GT, Kim TS, Sunwoo IN, Steinert PM, Kim SY
The Journal of biological chemistry 2000 Mar 24;275(12):8703-10
The Journal of biological chemistry 2000 Mar 24;275(12):8703-10
Antibodies to the C-terminus of the beta-amyloid precursor protein (APP): a site specific marker for the detection of traumatic axonal injury.
Stone JR, Singleton RH, Povlishock JT
Brain research 2000 Jul 21;871(2):288-302
Brain research 2000 Jul 21;871(2):288-302
No comments: Submit comment
Supportive validation
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
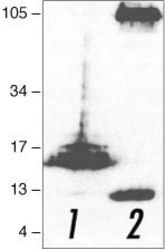
- Experimental details
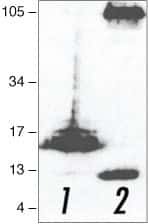
- Western blot analysis of (lane 1) APP C-terminal fragment (C100)- and (lane 2) full-length APP (751)-transfected HEK293 cells using Rb anti-Amyloid-b Precursor Protein (APP) (Product # 51-2700).
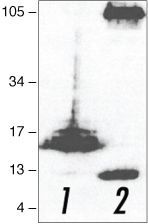
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- Western blot analysis of (lane 1) APP C-terminal fragment (C100)- and (lane 2) full-length APP (751)-transfected HEK293 cells using Rb anti-Amyloid-b Precursor Protein (APP) (Product # 51-2700).
- Submitted by
- Invitrogen Antibodies (provider)
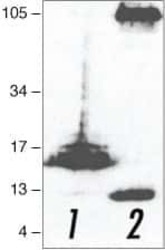
- Main image
- Experimental details
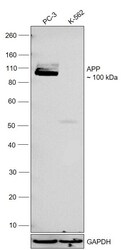
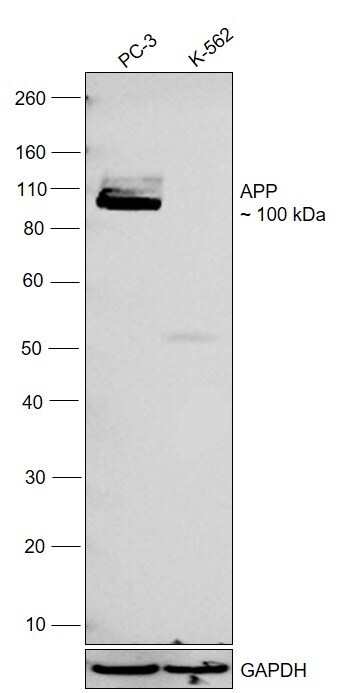
- Western blot was performed using Anti-beta Amyloid Polyclonal Antibody (CT695) (Product # 51-2700) and a 100 kDa band corresponding to APP was observed across in PC-3 and no band was observed in K-562 which is a low expression model. Whole cell extracts (30 µg lysate) of PC-3 (Lane 1), K-562 (Lane 2) were electrophoresed using NuPAGE™ 4-12% Bis-Tris Protein Gel (Product # NP0321BOX). Resolved proteins were then transferred onto a nitrocellulose membrane (Product # IB23001) by iBlot® 2 Dry Blotting System (Product # IB21001). The blot was probed with the primary antibody (1:500) and detected by chemiluminescence with Goat anti-Rabbit IgG (H+L) Superclonal™ Recombinant Secondary Antibody, HRP (Product # A27036,1:10000) using the iBright™ FL1500 Imaging System (Product # A44115). Chemiluminescent detection was performed using SuperSignal™ West Pico PLUS Chemiluminescent Substrate (Product # 34580).
Supportive validation
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
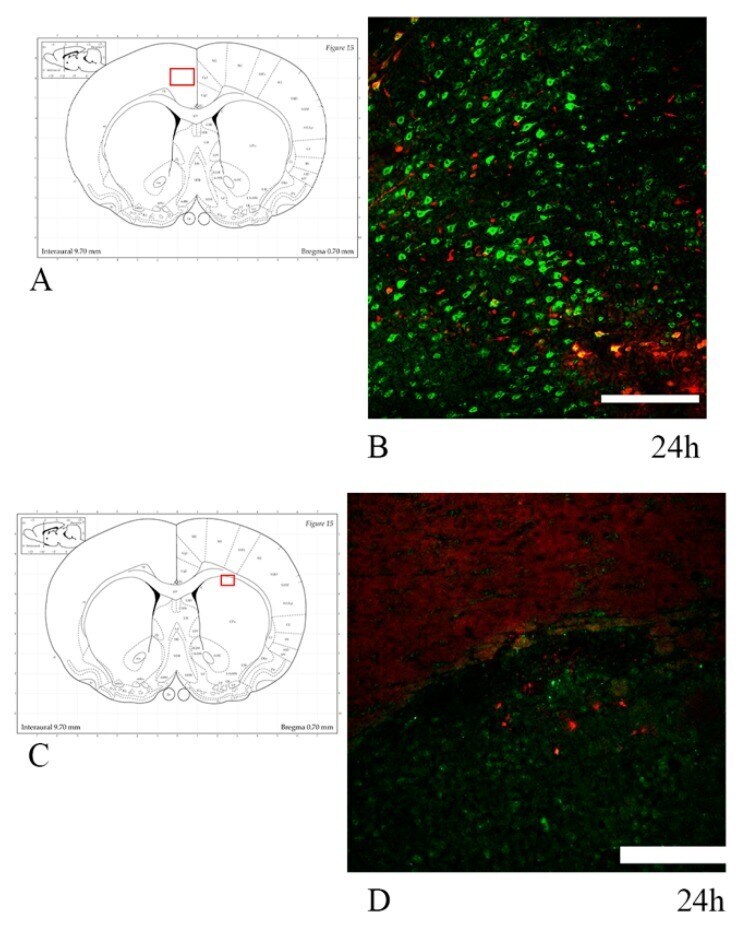
- Experimental details
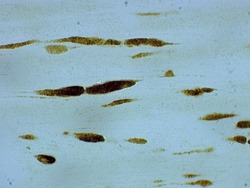
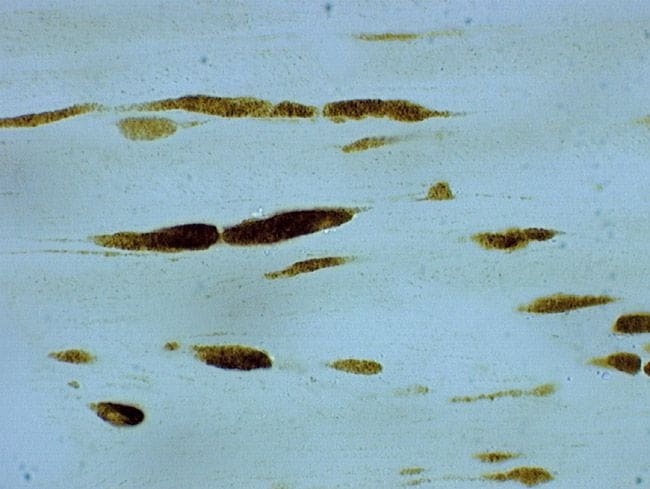
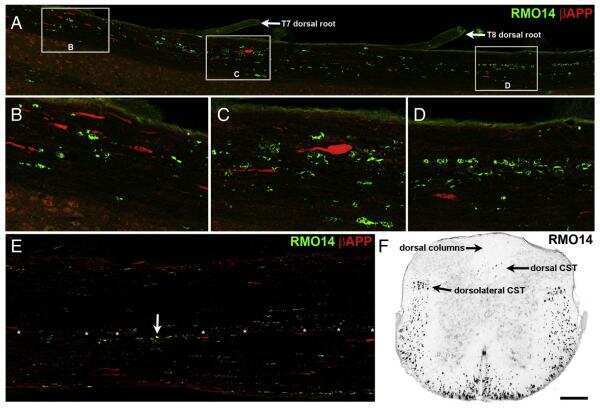
- Rabbit anti-Beta-Amyloid Precursor Protein (CT695) stained rat corticospinal tract after 24 hrs Traumatic Axonal Injury. Image: Courtesy of Dr. James R. Stone, University of Virginia, Dept. of Neurosurgery.
Supportive validation
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL

- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL

- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
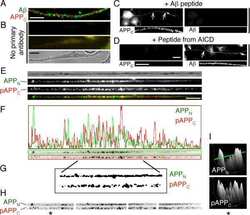
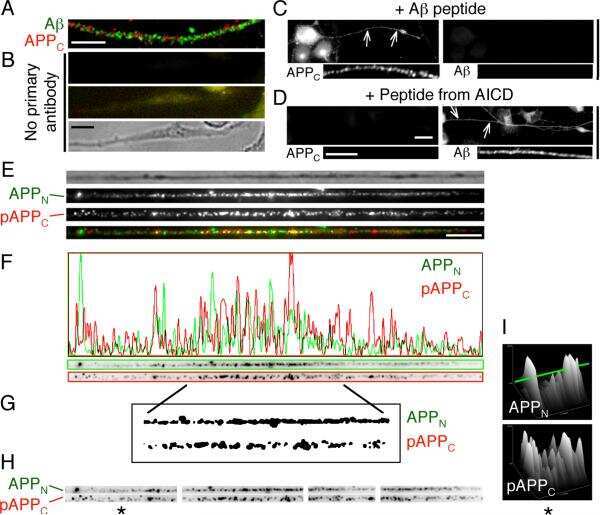
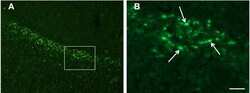
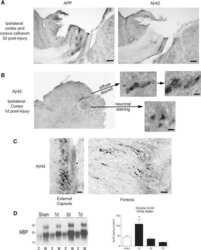
- Figure 3 Confocal images of beta-APP stained tissue, coronal plane, frontal sections . In (A) a low magnification image shows the border between the corpus callosum (lower part) and subcortical white matter. A larger number of beta-APP-positive profiles are visible at the border between the corpus callosum and the subcortical white matter, 24 h after high acceleration trauma. The box in (A) indicates the area that is shown in higher magnification in (B) , in which beta-APP-positive profiles have been indicated by arrows (scale bar = 25 mum).
- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
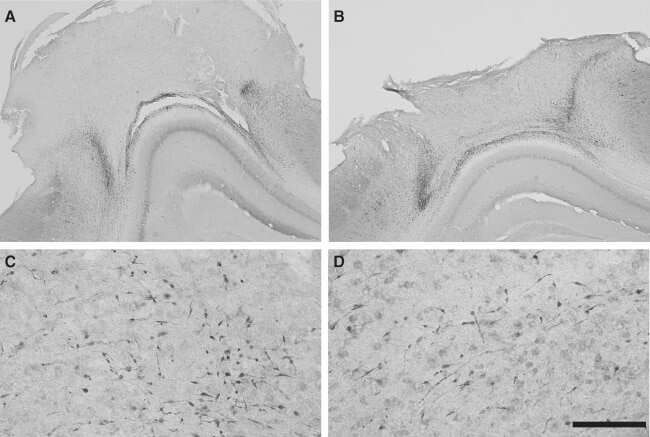
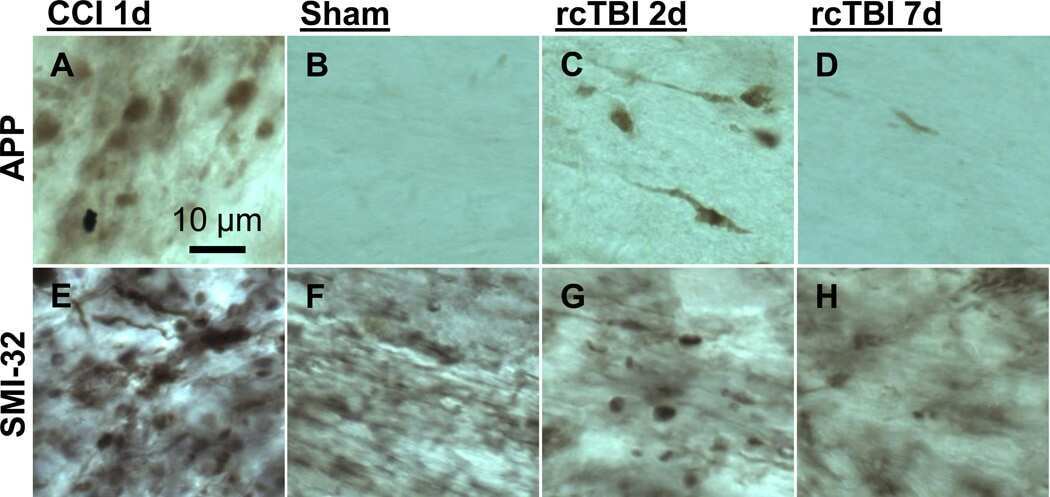
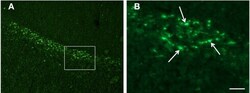
- Fig. 3 Abundant DAI is readily apparent 6 h following cFPI in the micro pig thalamus. Representative photomicrographs of APP immunofluorescence in the thalamus of animals sustaining sham ( a ) or cFPI ( b ). While sham-injured animals had little to no APP labeling, prevalent APP+ axonal swellings, indicative of DAI, were apparent following injury. c Bar graph depicting the average number of APP labeled axonal swellings/ 0.72 mm 2 of thalamic tissue. Graph depicts mean +- standard error of the mean. * p < 0.05. Scale bar: 50 mum
- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
- NULL

- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
- NULL

- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL

- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL

- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
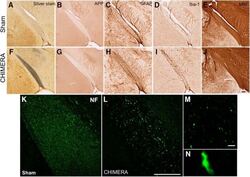
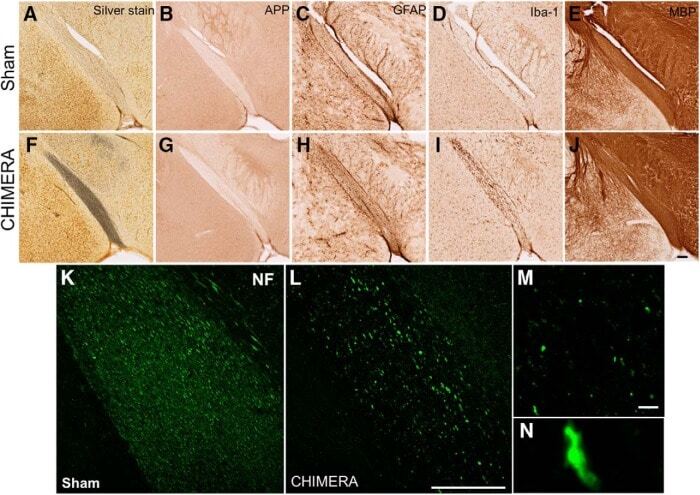
- Figure 4. Regions with altered DTI metrics exhibit histologic abnormalities consistent with injury. Representative photomicrographs of sham ( A-E ) and 0.65J CHIMERA-injured ( F-J ) optic tract. Sections were stained with silver stain ( A , F ), APP ( B , G ), GFAP ( C , H ), IBA-1 ( D , I ), MBP ( E , J ), and NF ( K , L ). Scale bar = 100 mum. Retraction bulbs and axonal varicosities are prominent in magnified photomicrographs of the 0.65J tissue ( M , N ). Scale bar = 10 mum.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
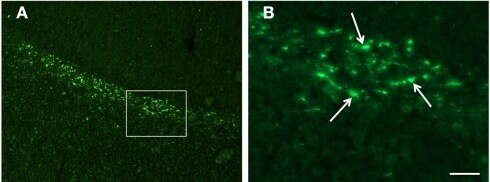
- Figure 3 Confocal images of beta-APP stained tissue, coronal plane, frontal sections . In (A) a low magnification image shows the border between the corpus callosum (lower part) and subcortical white matter. A larger number of beta-APP-positive profiles are visible at the border between the corpus callosum and the subcortical white matter, 24 h after high acceleration trauma. The box in (A) indicates the area that is shown in higher magnification in (B) , in which beta-APP-positive profiles have been indicated by arrows (scale bar = 25 mum).
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
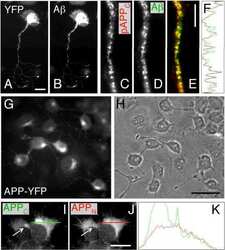
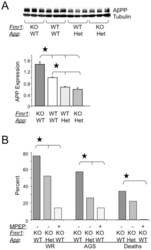
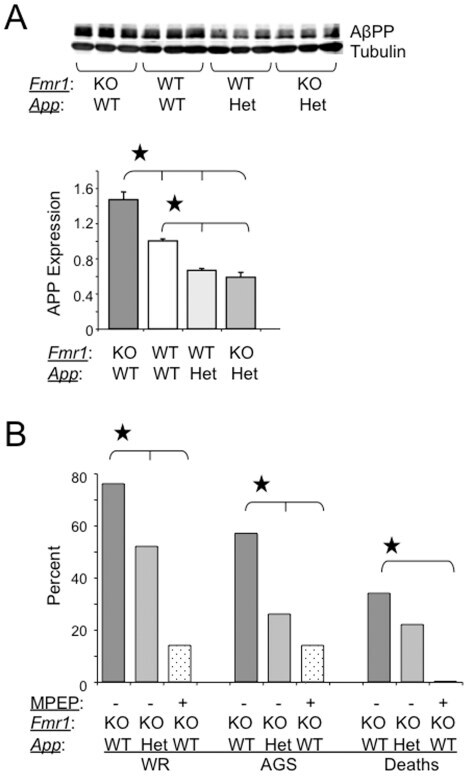
- Figure 1 AGS are rescued by genetic manipulation of App or mGluR 5 blockade. (A) western blot analyses of AbetaPP levels in Fmr1 KO , WT, App HE T and Fmr1 KO /App HET mice (n = 3 male mice per strain, 1 month old). Statistics: one-way ANOVA p
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
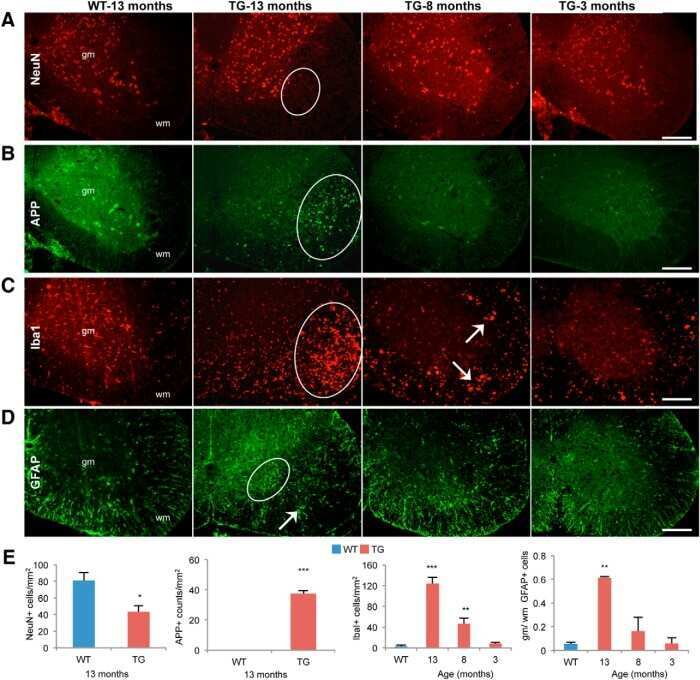
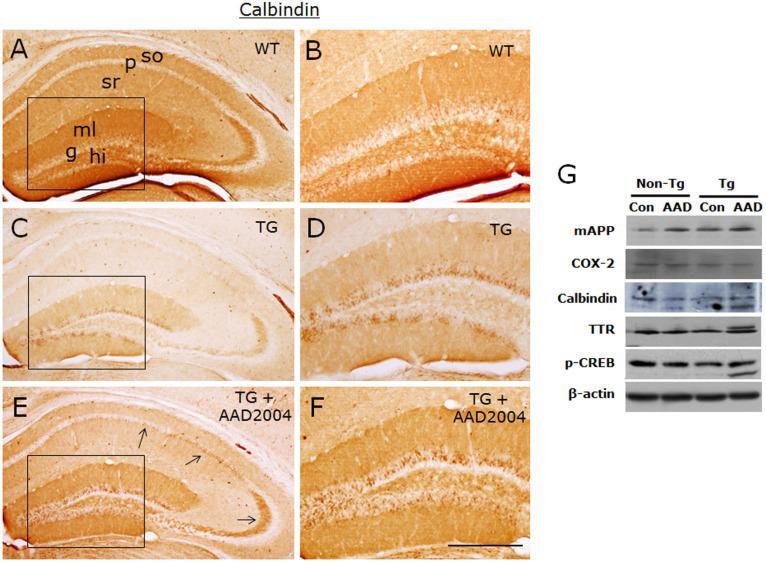
- Fig. 4 AAD-2004 partially reversed the down-regulated expression of calbindin in the brain of Tg-betaCTF99/B6 mice. (A-F) Photomicrographs showing anti-calbindin-stained hippocampus of non-transgenic control mice (WT; A, B), control Tg-betaCTF99/B6 mice (TG; C, D), and Tg-betaCTF99/B6 mice fed with AAD-2004 (TG+AAD-2004; E, F). Photomicrographs with high magnification of rectangles on the left panels (A, C, E) were shown (B, D, F). Note the anti-calbindin immunoreactivity not only in the granular layer (g) of the dentate gyrus, but also in the pyramidal cells (arrows), in the molecular layer (ml) of the dentate gyrus, and in the stratum oriens (so) and stratum radiatum (sr) of the hippocampus. p, pyramidal layer; hi, hilus. Scale bar, 200 um. (G) Representative Western blot images showing the expression of mouse amyloid precursor protein (mAPP), COX-2, calbindin, transthyretin (TTR), phospho-CREB, and beta-actin in the hippocampus. Total 5-8 animals were analyzed for each group.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
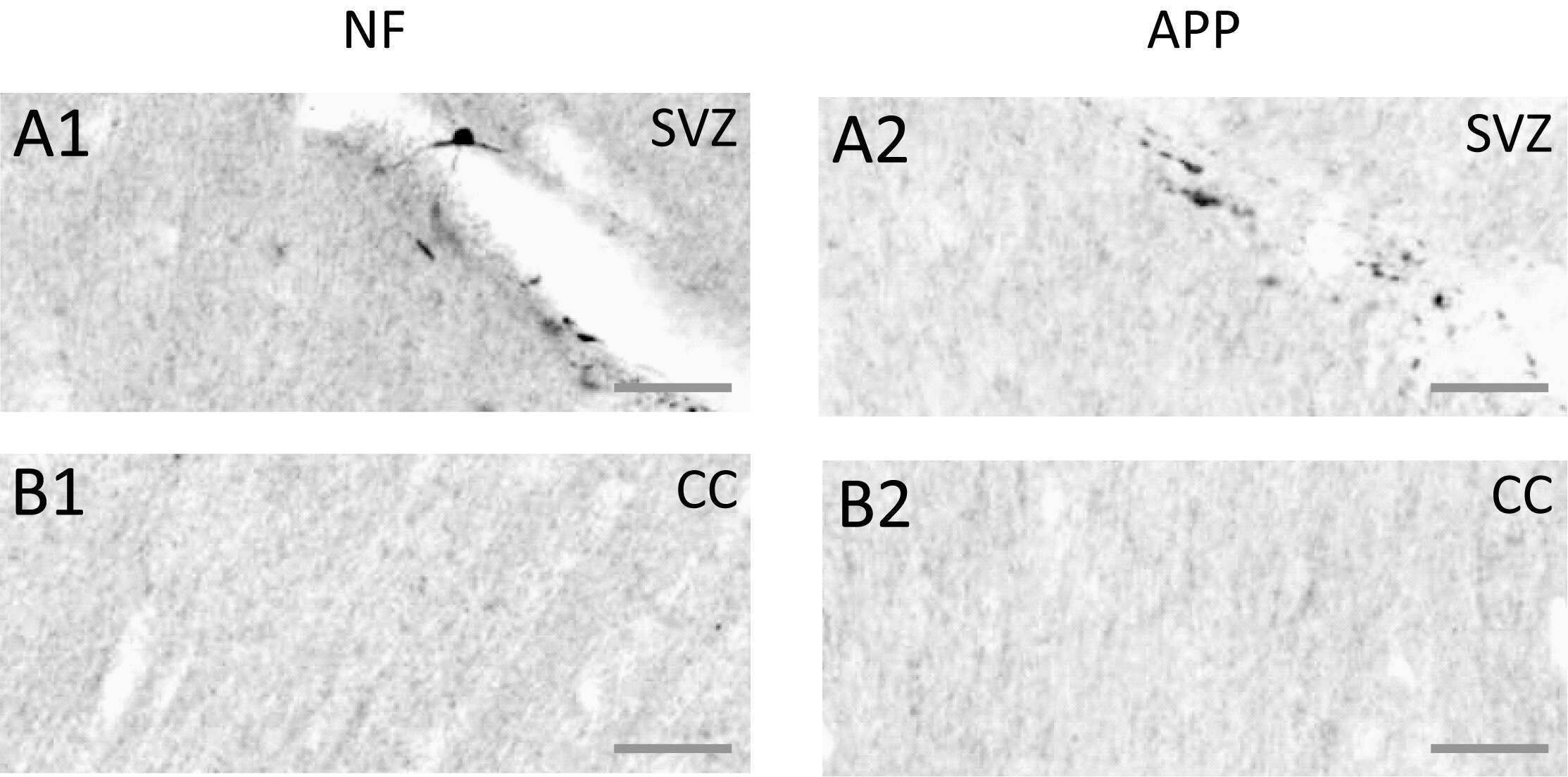
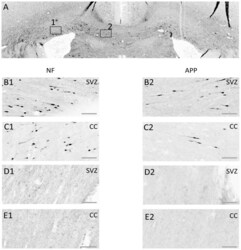
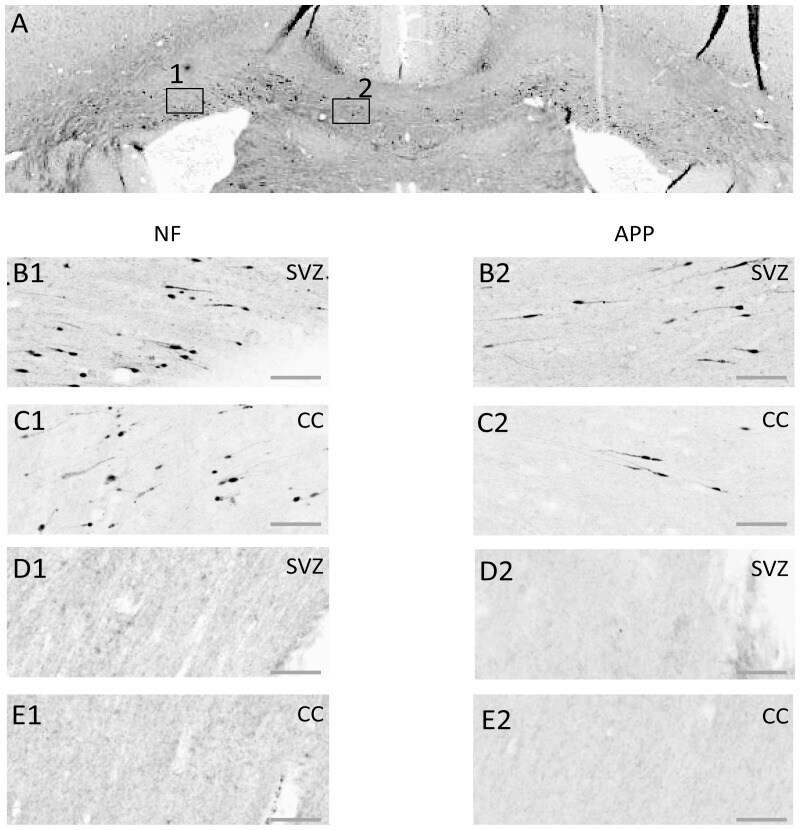
- Figure 8 Axonal injury as indicated by immunohistochemical staining for neurofilament heavy-chain (""NF-H"") OR beta-amyloid precursor protein (APP). Exemplar coronal section (x6 magnification) 1.3 mm caudal of bregma analysed for staining for NF-H and for beta-APP is shown in ( A ). The boxes indicate the sub-ventricular zone (Box 1; SVZ) or the corpus callosum (Box 2; CC) shown in greater detail (x20 magnification) in the next four rows of panels. ( B, C ) Staining in a TBI animal 24 hrs post-TBI for NF-H (B1, C1) or beta-APP (B2, C2) in the SVZ (row B) or the CC (row C). ( D, E ) As for ( B, C ) for equivalent brain regions, from a Sham control animal. Scale bar indicates 50 um.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
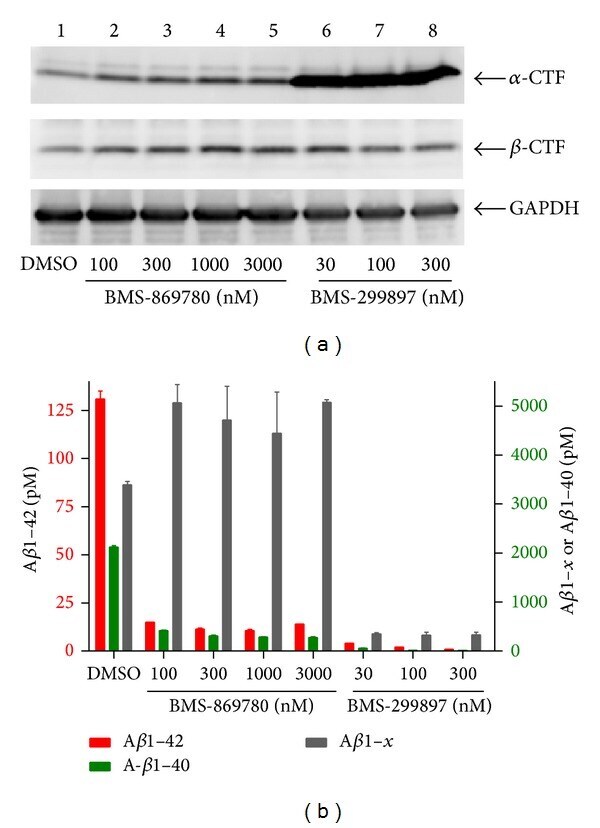
- Figure 4 BMS-869780 had minimal effect on APP-CTF accumulation in vitro . H4-APPsw cell cultures were treated overnight with the indicated concentrations of BMS-869780, BMS-299897 or vehicle (0.1% DMSO). (a) Cells were harvested and analyzed by western blotting for APP-CTF alpha , APP-CTF beta , and GAPDH. Lane 1; culture treated with vehicle 0.1% DMSO. Lanes 2-5; cultures treated with BMS-869780 at 100 nM, 300 nM, 1000 nM, or 3000 nM, respectively. Lanes 6-8; cultures treated with BMS-299897 at 30 nM, 100 nM, or 300 nM, respectively. (b) Levels of A beta 1-42 (red; left Y -axis), A beta 1-40 (green; right Y -axis), and A beta 1- x (grey; right Y -axis) were quantified.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
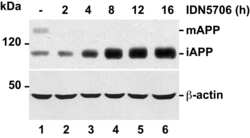
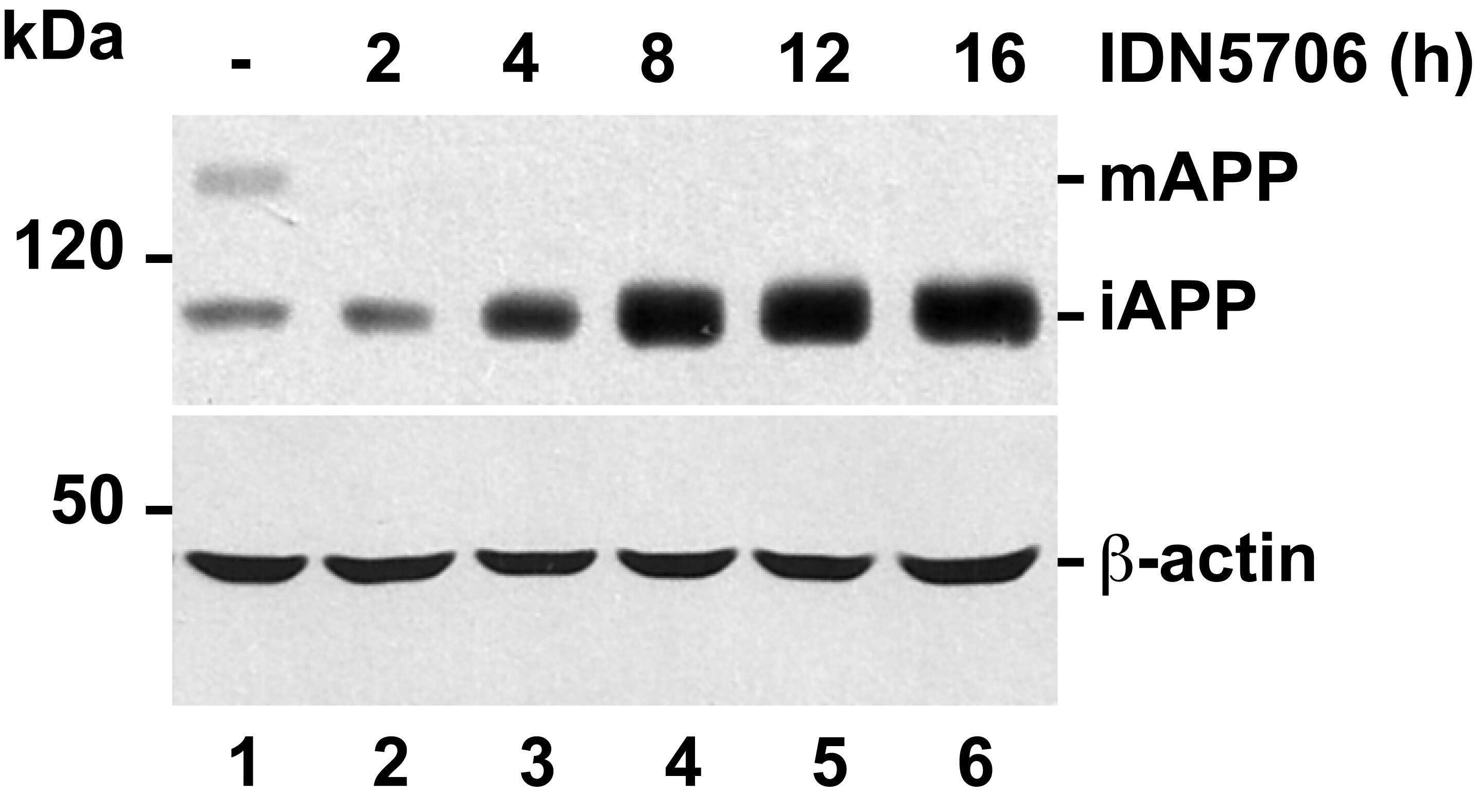
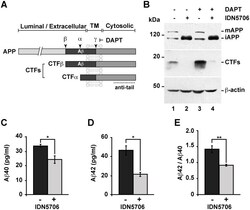
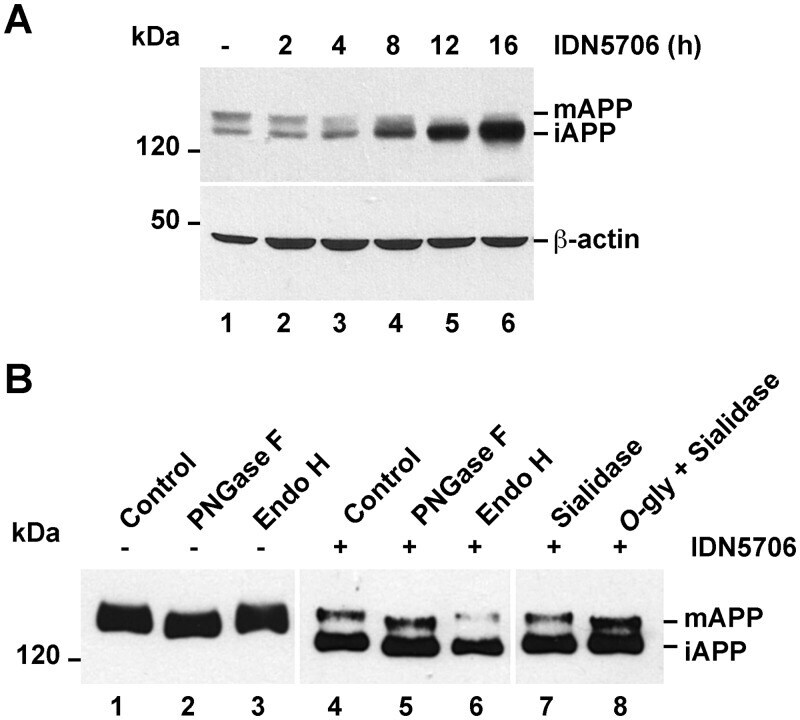
- Fig 6 IDN5706 inhibits proteolytic processing of APP to CTFs, and production of Abeta species. (A) Schematic representation of APP and carboxy-terminal fragments (CTFs) indicating their topological domains and the position of the proteolytic cleavage sites by alpha, beta and gamma secretases. The cytosolic region, recognized by the anti-tail antibody, and the gamma-secretase inhibitor, DAPT, are indicated. (B) H4 cells were left untreated (lane 1) or treated for 16 h either with 250 muM IDN5706 (lane 2), 1 muM DAPT (lane 3), or with a combination of 250 muM IDN5706 and 1 muM DAPT (lane 4). Cell extracts were subjected to Western blot analysis using the anti-tail antibody to the cytosolic C-terminal region of APP. Western blotting with antibody to beta-actin was used as loading control. mAPP, mature APP; iAPP, immature APP. The position of molecular mass markers is indicated on the left. (C-D) CHO 7AP2 cells were cultured in DMEM containing low glucose and without fetal bovine serum, in the absence or presence of 250 muM IDN5706 for 16 h. The amount of Abeta40 and Abeta42 peptides in the culture medium was analyzed by ELISA. (E) Ratio of the amount of Abeta42 and Abeta40 peptides as an indicator of toxicity. (C-E) Values are presented as the mean +- SD of three independent experiments. *, P < 0,05 and **, P < 0,005.

- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
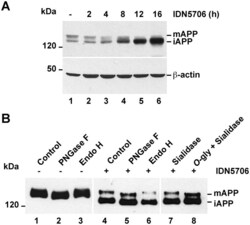
- Fig 7 IDN5706 disrupts glycosylation of APP. (A) H4 cells stably expressing an amyloidogenic version of APP tagged to GFP (APP-GFP) were treated with 250 muM IDN5706 for the indicated periods of time. Cell extracts were subjected to Western blot analysis with an antibody to GFP. Western blotting with antibody to beta-actin was used as loading control. (B) H4 cells stably expressing APP-GFP, were left untreated (lanes 1-3) or treated for 16 h with 250 muM IDN5706 (lanes 4-8). Cell extracts were subjected to immunoprecipitation with a an antibody to GFP, followed by denaturation and digestion with the indicated glycosidases for 1 h at 37degC. Immunoprecipitated proteins were subjected to Western blot analysis with anti-GFP-HRP. (A-B) The position of molecular mass markers is indicated on the left. mAPP, mature APP; iAPP, immature APP.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
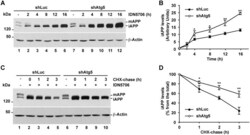
- Fig 9 Accumulated immature APP in response to IDN5706 is degraded by Atg5-dependent autophagy. (A) H4 cells stably expressing an amyloidogenic version of APP tagged to GFP, and stably expressing either luciferase shRNA (control; shLuc) or Atg5 shRNA (shAtg5), were treated with 250 muM IDN5706 for the indicated periods of time, or (C) were left untreated (lanes 1 and 6) or treated with 250 muM IDN5706 (lanes 2-5 and lanes 7-10) for 12 h, followed by cycloheximide-chase with 150 mug/ml cycloheximide and 40 mug/ml chloramphenicol for 1-3 h in the presence of 250 muM IDN5706 (lanes 2-5 and lanes 7-10). Equivalent amounts of cell extracts were subjected to SDS-PAGE in 7.5% acrylamide gels, followed by Western blotting with an antibody to GFP. Western blotting with antibody to beta-actin was used as loading control. mAPP, mature APP; iAPP, immature APP. The position of molecular mass markers is indicated on the left. (B and D) Densitometric quantification of the levels of iAPP shown either in A or C. The values depicted in the graphs represent the mean +- SD of three independent experiments. *, P < 0.05; **, P < 0.01.

- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
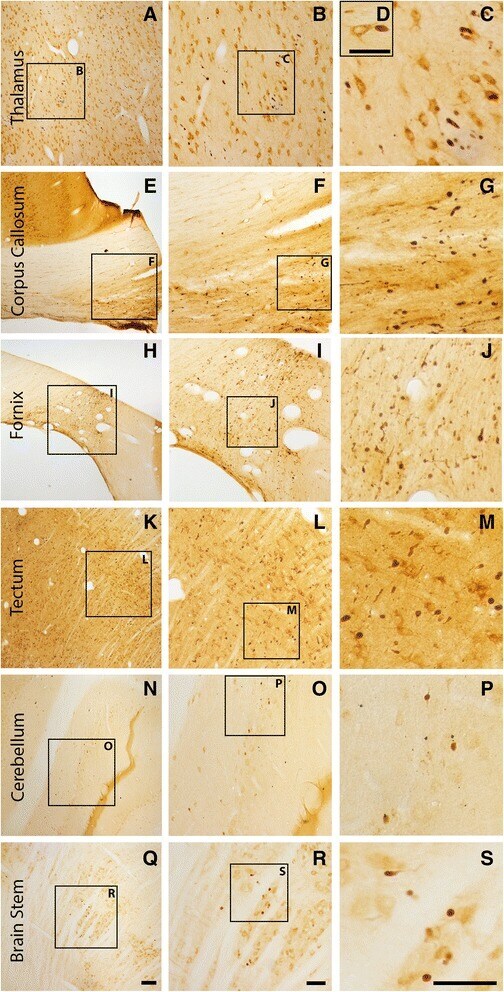
- Fig. 2 Axonal injury is observed in various regions throughout the micro pig brain following cFPI. Representative photomicrographs of APP immunohistochemistry in regions of the micro pig brain that demonstrated DAI in animals sustaining cFPI. Images in the middle panel ( b , f , i , l , o , r ) are magnified regions indicated in the images of the left panel ( a , e , h , k , n , q ) and images in the right panel ( c , g , j , m , p , s ) are magnified regions indicated in the middle panel ( b , f , i , l , o , r ), respectively. Note that DAI within the thalamus and tectum was diffusely distributed throughout the domain, while DAI within the other regions was more localized. Also note that while not common, APP+ proximal axonal swellings in continuity with the neuronal soma ( d ) were observed in the thalamus. Scale bar in q : 200 mum; r and s : 100 mum; d : 50 mum
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
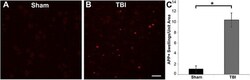
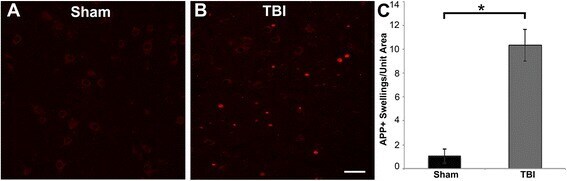
- Fig. 3 Abundant DAI is readily apparent 6 h following cFPI in the micro pig thalamus. Representative photomicrographs of APP immunofluorescence in the thalamus of animals sustaining sham ( a ) or cFPI ( b ). While sham-injured animals had little to no APP labeling, prevalent APP+ axonal swellings, indicative of DAI, were apparent following injury. c Bar graph depicting the average number of APP labeled axonal swellings/ 0.72 mm 2 of thalamic tissue. Graph depicts mean +- standard error of the mean. * p < 0.05. Scale bar: 50 mum
- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
- Fig. 6 Microglia activation occurs in thalamic sectors sustaining acute DAI 6 h following mTBI. Representative confocal micrographs of APP ( red ; a and d ) and Iba-1 ( green ; b and e ), with overlays in c and f , in the thalamus of the same injured animal. Interestingly, areas lacking axonal injury ( a - c ) appear to contain inactive ramified microglia ( arrows ), whereas thalamic sites exhibiting DAI, indicated by accumulation of APP in axonal swellings ( d - f ), also appear to contain the majority of morphologically activated Iba-1+ microglia ( arrow heads ). Scale bar: 20 mum

- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
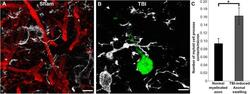
- Fig. 7 Microglia processes appear to preferentially contact TBI-induced proximal axonal swellings. Representative 3D reconstructions of MBP+ myelinated axons ( red ) or APP+ axonal swellings ( green ) and Iba-1+ microglia ( white ) in sham-injured ( a ) or central fluid percussion injured ( b ) thalami. c Bar graph depicting the average number of Iba-1+ microglial processes contacting either MBP+ myelinated fibers in the sham animals or APP+ axonal swellings in injured animals. Graph depicts the mean +- standard error of the mean. * p < 0.05. Scale bar: 5 mum

- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
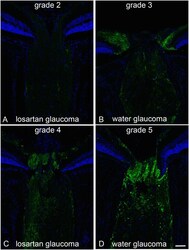
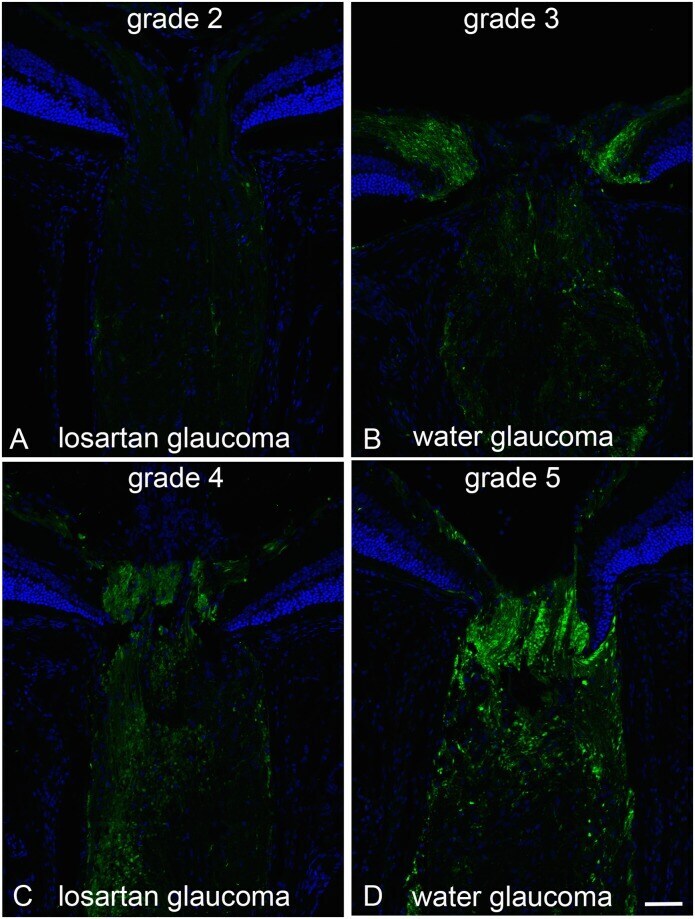
- Fig 2 Axonal transport effects, APP labeling with losartan-treated vs water control. Immunolabeling for APP in the optic nerve head of mice treated with oral losartan (A and C) and water alone (B and D) shows axonal transport obstruction 3 days after IOP elevation. The examples show 4 of the 5 grade levels for APP accumulation (level 1, not illustrated, had no label, indicating that there was no accumulation and no background or non-specific labeling). The grades of losartan-treated eyes ranged from 2 to 4 (A and C; mean = 2.6 (sd 0.8), while water-treated eyes ranged from 3 to 5 (B and D; mean = 4.0 (sd 0.9); difference from losartan, p = 0.007, multivariable regression adjusted for IOP exposure). Bar equals 50 um.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
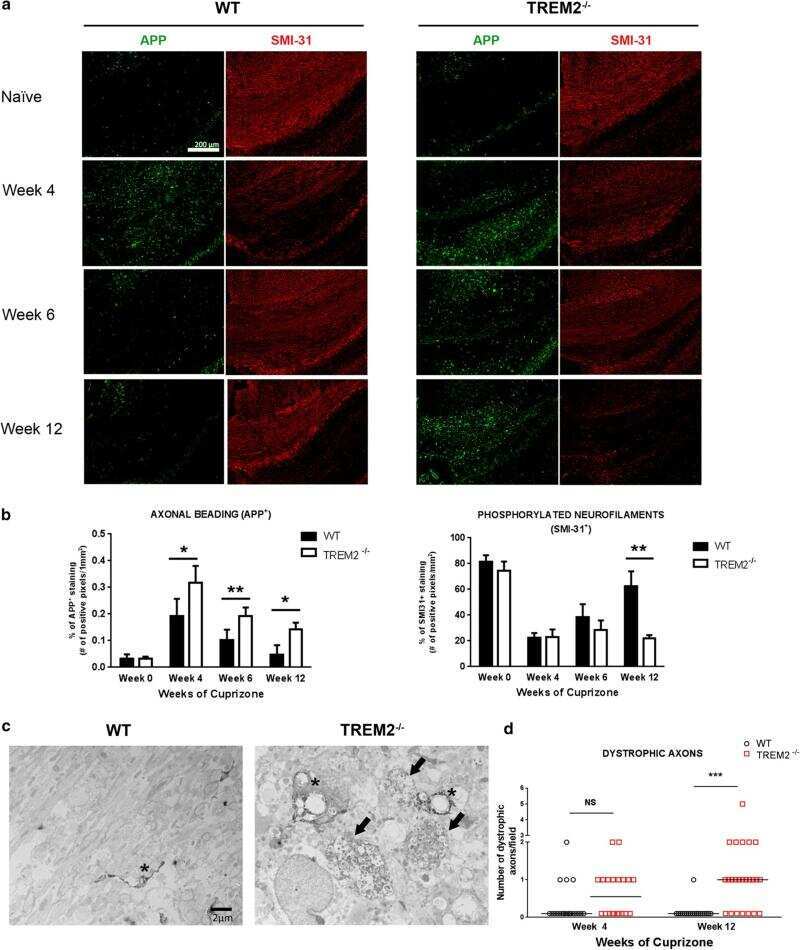
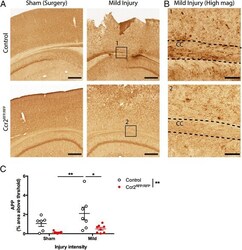
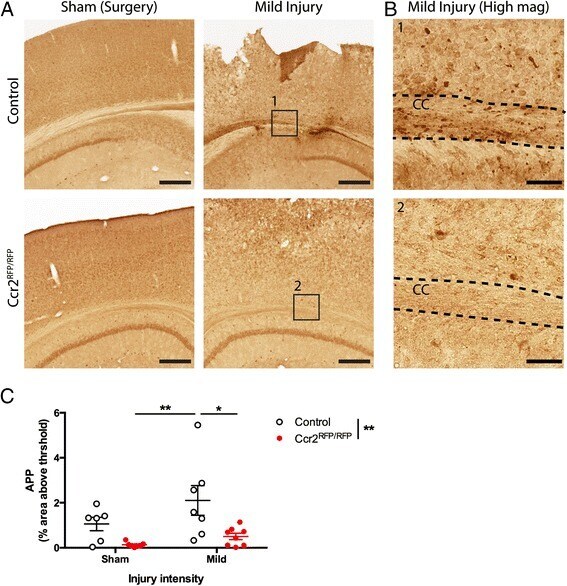
- Fig. 3 Effects of CCR2 signaling on axonal damage after TBI. Axonal pathology was evaluated at 3 dpi in control heterozygous and Ccr2 RFP / RFP mice by APP staining. a The injury induces APP accumulation in axons of the ipsilateral side to the injury, particularly in the corpus callosum. Ccr2 deletion decreases APP accumulation in axons. Scale bar , 500 mum. b Higher magnification images of the indicated regions ( boxes ) show that APP immunoreactivity aligns with axonal tracks in the corpus callosum (CC). Scale bar , 100 mum. c Quantification of axonal pathology as percent area of the corpus callosum with APP immunoreactivity above threshold. Statistics: two-way ANOVA and Tukey's post hoc test. Comparisons between groups are shown with horizontal lines ; vertical line in figure legend indicates main effect of genotype. * p < 0.05; ** p < 0.01
- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
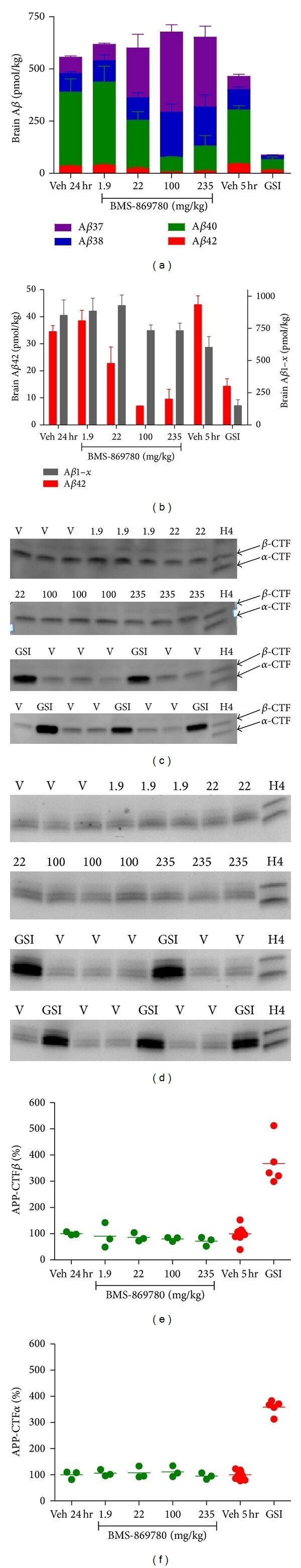
- Figure 5 BMS-869780 modulated A beta but did not cause accumulation of beta CTF or alpha CTF in rat brain. Rats were given oral doses of BMS-869780, and levels of brain A beta , beta CTF, and alpha CTF were determined 24 hours later. For comparison, BMS-698861 was dosed in a separate experiment and samples were taken 5 hours later. (a) Brain levels of A beta 1-42 (red), A beta 1-40 (green), A beta 1-38 (blue), and A beta 1-37 (purple) are shown as bars stacked upon one another. The total height of each bar therefore represents the sum of the four peptides. (b) A beta 1-42 (red--left Y axis) and A beta 1- x (grey--right Y axis). The same results for A beta 1-42 are plotted in both (a) and (b). (c) Rat brain beta CTF was detected by western blotting of immunoprecipitates from samples of the same rat brains used for A beta determinations. V, vehicle groups; results from rats dosed with 1.9, 22, 100, and 235 mg/kg of BMS-869780 and 10 mg/kg BMS-698861 (GSI) are indicated. (d) Western blots of immunoprecipitated alpha CTF from the same rat brain samples. (e) and (f) quantification of western blots shown in (c) and (d), respectively, expressed relative to percent of average level of CTF in vehicle-treated rats. Actual doses of BMS-869780 were determined by analysis of concentrations in left-over dosing solutions.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
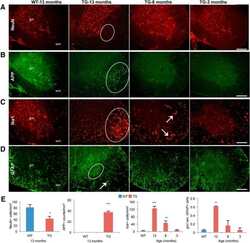
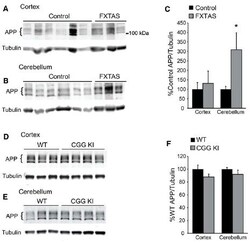
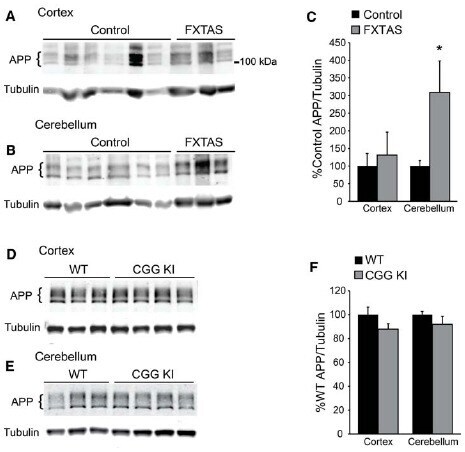
- FIGURE 5 Amyloid precursor protein expression in FXTAS patients and mouse models. (A) Frontal cortex lysates from control ( n = 10) and FXTAS patients ( n = 3) probed with a C-terminal alphaAPP antibody which detects the three primary isoforms of full-length APP (100-125 kDa). (B) Cerebellar lysates from the same individuals as in (A) . (C) Normalized APP expression relative to tubulin expressed as a percent of controls, performed in technical triplicate. (D) Twelve-month-old CGG KI ( n = 4) and WT littermate control ( n = 3) cortex and subcortical lysates probed with alphaAPP. (E) Cerebellar lysates from the same animals as in (D) . (F) Normalized APP expression in CGG KI mice expressed as a percent of WT controls, performed in technical triplicate. * P < 0.05 Student's t-test.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
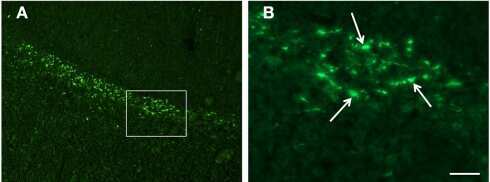
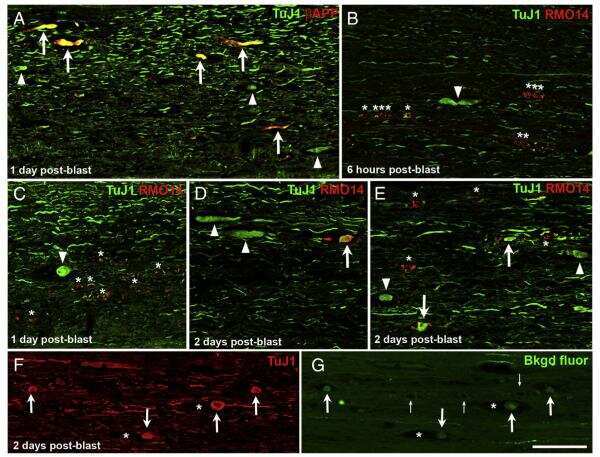
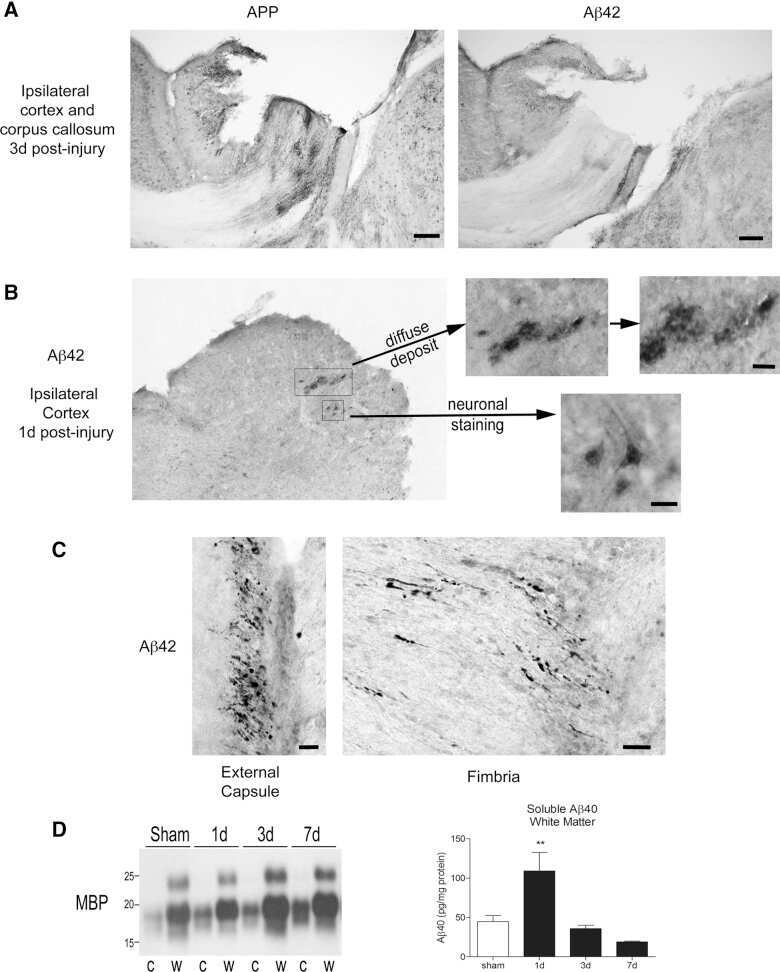
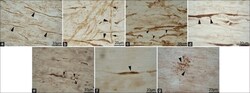
- Figure 3 (a) Normal looking axons in the white matter tracts of cervical spinal cord of a 24 h survival sham animal (b) axons with prominent swellings (arrowheads) and swollen axons with vacuolations (arrows) in the white matter tracts of cervical spinal cord at 6 h (c and d) prominent disruptions of axonal membranes (arrowheads) at 6 h and 24 h respectively. These disruptions in the form of wide spaces on the margins or projections with ragged edges could be seen in various large caliber axons (e) axonal injury in the form of vacuolations in the axonal core at 24 h (f) a beta amyloid precursor protein reactive swollen axon in the cervical spinal cord white matter tract at 6 h post blast period (g) beta amyloid precursor protein reactive punctate axons 24 h post blast

- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
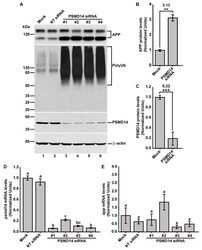
- Figure 2 PSMD14 is validated as a regulator of the endogenous APP levels. ( A ) Protein extracts of parental H4 cells either untransfected (Mock), transfected with NT siRNA, or transfected with four different PSMD14 siRNA sequences for 72 h were analyzed by western blot. Polyclonal antibodies to endogenous APP (CT695) and to Ub (that recognizes all types of Ub conjugates), and monoclonal antibodies to PSMD14 (clone D18C7) and to beta-actin (clone BA3R), were tested. The position of molecular mass markers are indicated on the left. Densitometric quantification of the levels of endogenous APP ( B ) and PSMD14 ( C ) in H4 cells transfected with PSMD14 siRNA#1, compared to untransfected cells (Mock). Statistical significance was determined by Student's t-test. Bars represent the mean +- SD of biological replicates (APP n =5; PSMD14 n = 4). ** p < 0.01 and *** p < 0.001. ( D ) mRNA levels of psmd14 and ( E ) mRNA levels of app were measured using RT-qPCR from parental H4 cells transfected for 72 h. All data were normalized for TATA binding protein expression in either untransfected cells (Mock), cells transfected with NT siRNA or cells transfected with four different PSMD14 siRNAs duplexes. Statistical significance was determined by One-Way ANOVA, followed by Tukey's test. Bars represent the mean +- SD of biological replicates ( psmd14 n = 3; app n = 3). Different letters above the mean bars apply to significant differences between groups p < 0.01.

- Submitted by
- Invitrogen Antibodies (provider)
- Main image
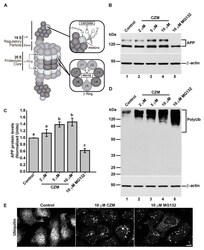
- Experimental details
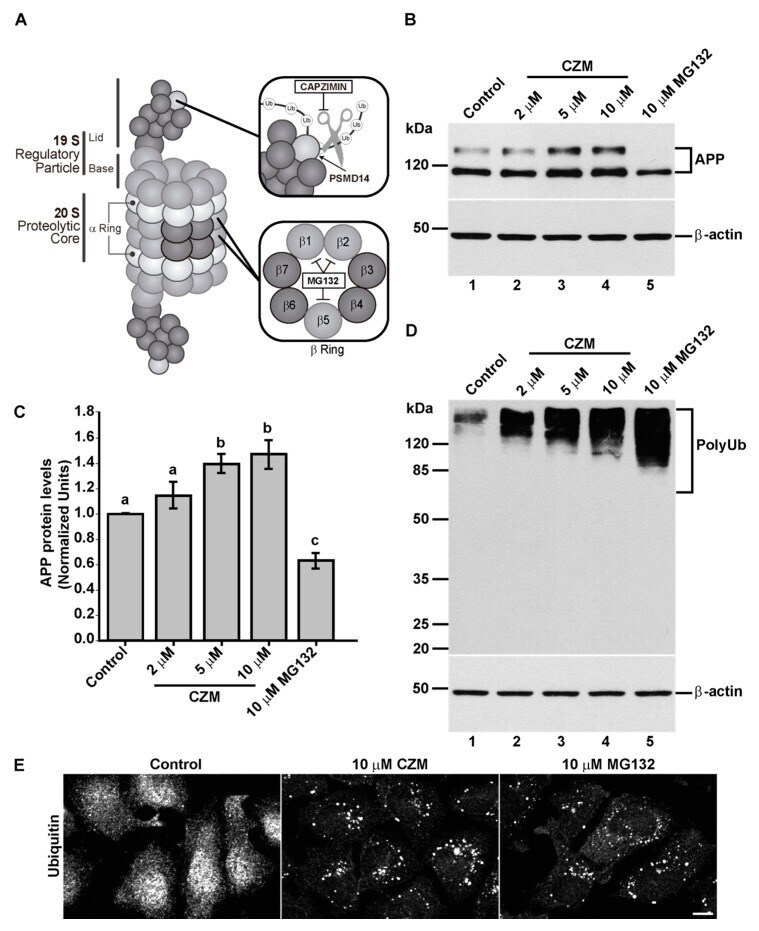
- Figure 3 Acute inhibition of PSMD14 by Capzimin CZM shows a similar phenotype as that of PSMD14 KD on the levels of APP and high molecular weight Ub conjugates. ( A ) Schematic diagram of the molecular targets of Capzimin and MG132 in the 19S RP and 20S catalytic core of the proteasome, respectively. ( B ) Parental H4 cells were treated either with vehicle (DMSO; Control), or increasing doses of CZM for 4 h, or MG132 for 6 h. Protein extracts were analyzed by western blot with a polyclonal antibody to endogenous APP. Monoclonal antibody to beta-actin (clone BA3R) was used as a loading control. The position of molecular mass markers is indicated on the left. ( C ) Densitometric quantification of APP protein levels as shown in ( D ). Statistical significance was determined by one-way ANOVA, followed by Tukey's test. Bars represent the mean +- SD of biological replicates ( n = 4). Different letters above the mean bars apply to significant differences between groups p < 0.05. ( D ) Parental H4 cells were treated as in ( B ), and the protein extracts were analyzed by western blot with a polyclonal antibody to Ub that recognizes all types of Ub conjugate. Monoclonal antibody to beta-actin (clone BA3R) was used as a loading control. The position of molecular mass markers is indicated on the left. ( E ) Immunofluorescence microscopy images of the cellular localization of Ub in parental H4 cells treated with either the vehicle (DMSO; Control), CZM for 4 h or MG132 for 6 h. Cells were
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
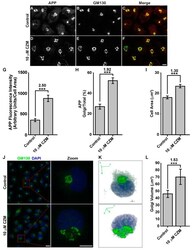
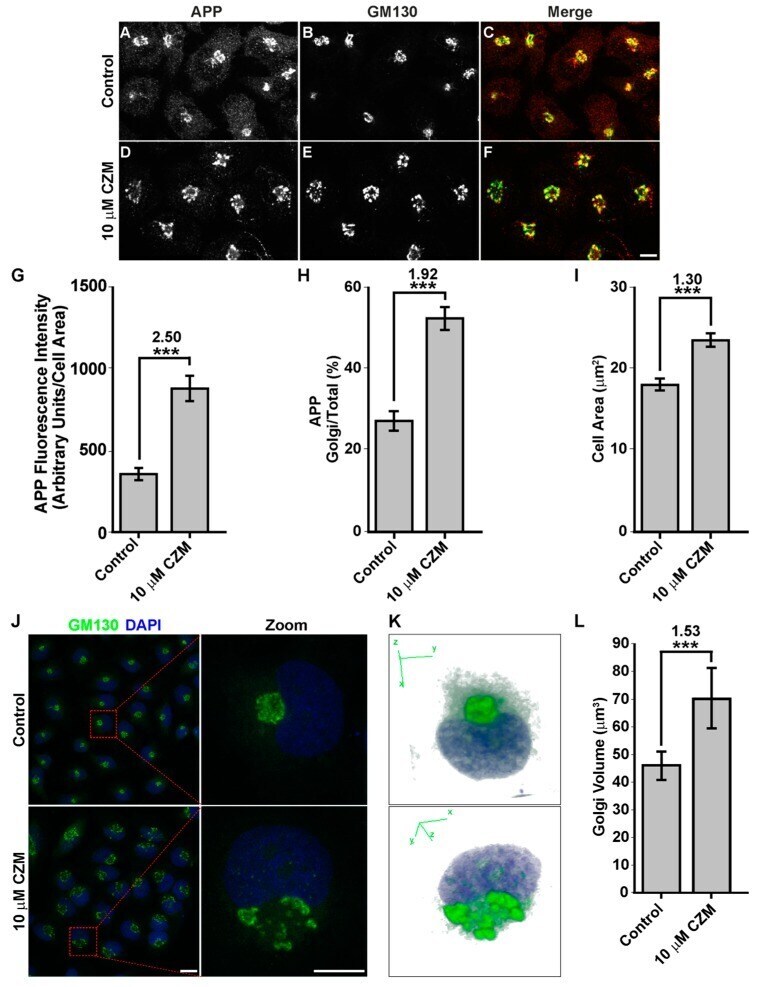
- Figure 4 Acute inhibition of PSMD14 by CZM triggers the accumulation of APP in a swollen Golgi apparatus. Immunofluorescence analysis of endogenous APP in H4 parental cells treated either with the vehicle (DMSO; Control) ( A - C ) or CZM ( D - F ) for 4 h. Cells were fixed, permeabilized, and double stained with a rabbit polyclonal antibody to APP (CT695) ( A , D ) and a mouse monoclonal antibody to GM130 (clone35/GM130) ( B , E ), followed by Alexa-594-conjugated donkey anti-Rabbit IgG and Alexa-488-conjugated donkey anti-Mouse IgG. Merging of the images generated the third picture ( C , F ). Scale bar, 10 mm. ( G ) Quantitative analysis of the mean of total fluorescence intensity of APP upon treatment with CZM, in comparison to control cells. The statistical significance was determined by Student's t-test. Bars represent the mean +- SD of the fluorescent signal per cell area ( n = 43 cells). *** p < 0.001. ( H ) Quantitative analysis of the fraction of APP colocalizing with GM130 under CZM treatment and compared to control cells. Statistical significance was determined by Student's t-test. Bars represent the mean +- SD of the fluorescent signal per cell area ( n = 43 cells). *** p < 0.001. ( I ) Quantitative analysis of the cell area. Statistical significance was determined by Student's t-test. Bars represent the mean +- SD of the cell area ( n = 43 cells) ** p < 0.001. ( J ) Immunofluorescence microscopy analysis of GM130 in parental H4 cells treated either with the vehicl
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
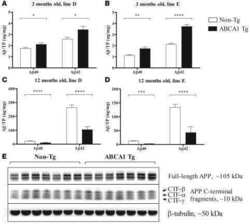
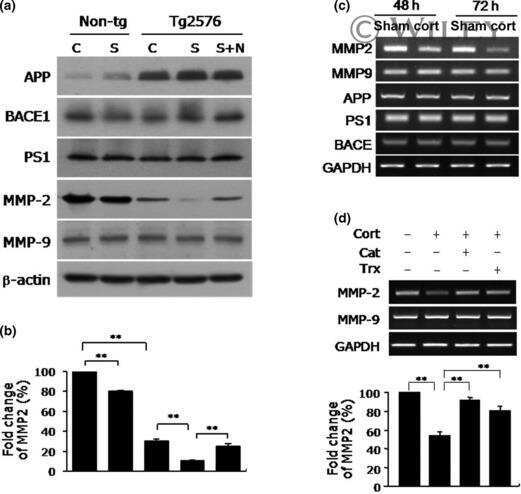
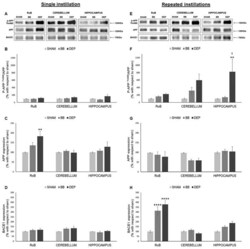
- Figure 3 Amyloidogenic precursor protein (APP) processing analysis after single and repeated instillations of BB and DEP. Representative immunoblotting images of amyloid precursor protein (APP), phosphorylated APP on threonine 668 (p-APP Thr668 ), and beta-secretase 1 (BACE1) analysis in mice after single ( A ) and repeated ( E ) instillations with 50 µg of BB or DEP/100 uL 0.9% NaCl. Histograms display p-APP Thr668 /APP, APP, and BACE1 protein levels in mice after single ( B - D ) and repeated ( F - H ) instillations with BB and DEP, with respect to sham. Proteins are normalized to corresponding total proteins revealed by Ponceau in each lane ( Figure S3, Supplementary Materials ), and the data are expressed as means +- SEM ( n = 6). Statistical differences were tested accordingly by one-way ANOVA followed by Tukey post hoc comparison. ** p < 0.01 vs. sham mice; **** p < 0.0001 vs. sham mice; SS p < 0.05 vs. BB-treated mice.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
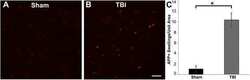
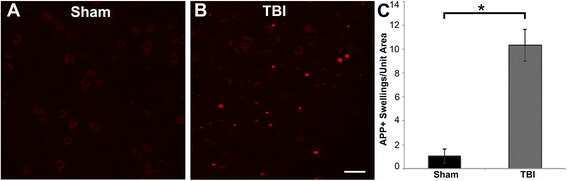
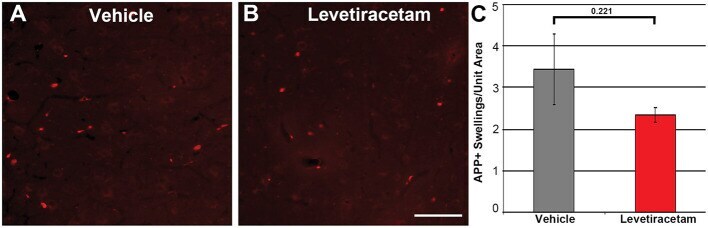
- Figure 3 Representative micrographs of amyloid precursor protein (APP) immunofluorescence in the thalamus of pigs sustaining central fluid percussion injury (cFPI) followed by (A) vehicle or (B) levetiracetam (LEV) treatment. (C) Bar graph depicting the average number of APP-labeled axonal swelling/unit area of thalamic tissue at 6 h post-cFPI (vehicle, n = 7 pigs; LEV, n = 7 pigs). The number of axonal swellings at 6 h was not significantly different with LEV treatment vs. vehicle. Scale = 100 um.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
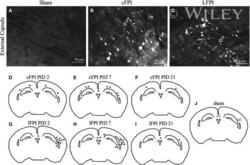
- Experimental details
- Axonal injury indicated by post-injury accumulation of APP . (A-C) Example of APP immunostaining from the external capsule. (A and J) No APP accumulation was observed in sham-injured controls. (B and C) Following TBI , APP accumulation was observed, showing both the classical varicosities (open arrows) and axonal bulbs (white arrows) in both TBI models. (D-I) Schematic overview of post-injury accumulation of APP in the two TBI models from 2 to 21 days post-injury. Each asterisk indicates a region of APP -positive axonal profile in the evaluated white matter regions that was not observed in sham-injured controls. Note the decreased APP immunostaining at the late time points in both TBI models. CC , corpus callosum; EC , external capsule; F, fimbriae; PID , post-injury day.
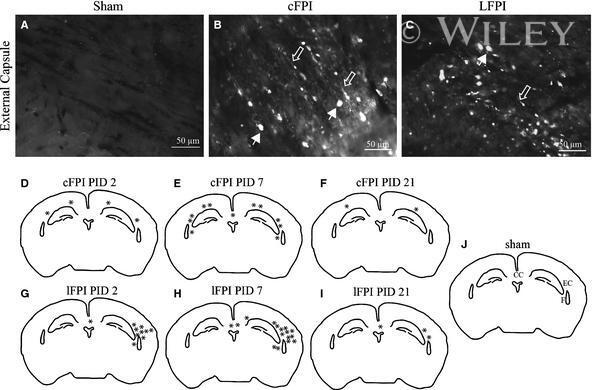
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
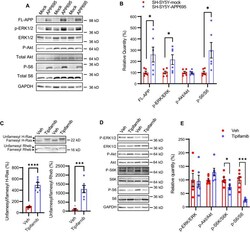
- Experimental details
- Fig. 10 Pharmacological inhibition of FT reverses the hyperactivation of Ras-Rheb-mTORC1 signaling in SH-SY5Y-APP695 cells. A Immunoblot analysis of FL-APP and downstream targets of Ras-Rheb-mTORC1 signaling, including ERK and ribosomal protein S6, in the lysates of SH-SY5Y-APP695 and mock-transfected control cells. B Densitometric analysis of immunoblots normalized by the amount of GAPDH or total proteins for phosphorylated proteins (P-ERK, P-Akt, P-S6) with the levels in the control group set as 100%. n = 6 replicates from 2 independent experiments. C Immunoblot analysis of farnesylation status of H-Ras and Rheb, and D phosphorylation status of downstream targets of Ras and Rheb in the lysates of SH-SY5Y-APP695 cells after treated with vehicle (DMSO) or 1 muM tipifarnib for 48 h. E Densitometric analysis of immunoblots of P-ERK, P-S6K, and P-S6 normalized by the amount of total ERK, S6K, and S6 with the levels in the control group set as 100%. n = 6 replicates from 2 independent experiments. Student's t-test, 2-tailed; * p < 0.05; *** p < 0.001; **** p < 0.0001
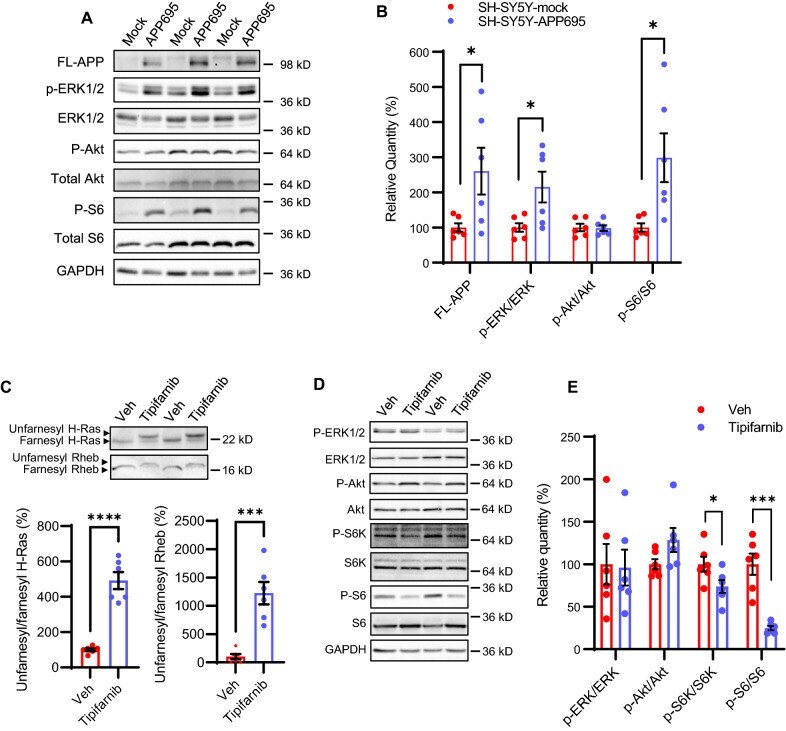
- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
- Fig. 7 Reduced APP proteolytic processing in APP/PS1/FTnKO mice. A , B Immunoblots and densitometric analysis of full-length APP (FL-APP), and APP cleavage products, carboxyl-terminal fragments (CTFalpha and CTFbeta), amino-terminal fragments of APP (sAPPalpha and sAPPbeta), and beta-secretase 1 (BACE1). Protein levels were normalized by the amount of tubulin with the levels in the APP/PS1 control group set as 100%. C , D Immunoblots and densitometric analysis of proteins involved in Abeta clearance pathways including APOE, LRP1, IDE, and neprilysin. Protein levels were normalized by the amount of tubulin or GAPDH, and the levels in the control group set as 100%. n = 8/genotype; Student's t-test, 2-tailed; # p = 0.08; * p < 0.05
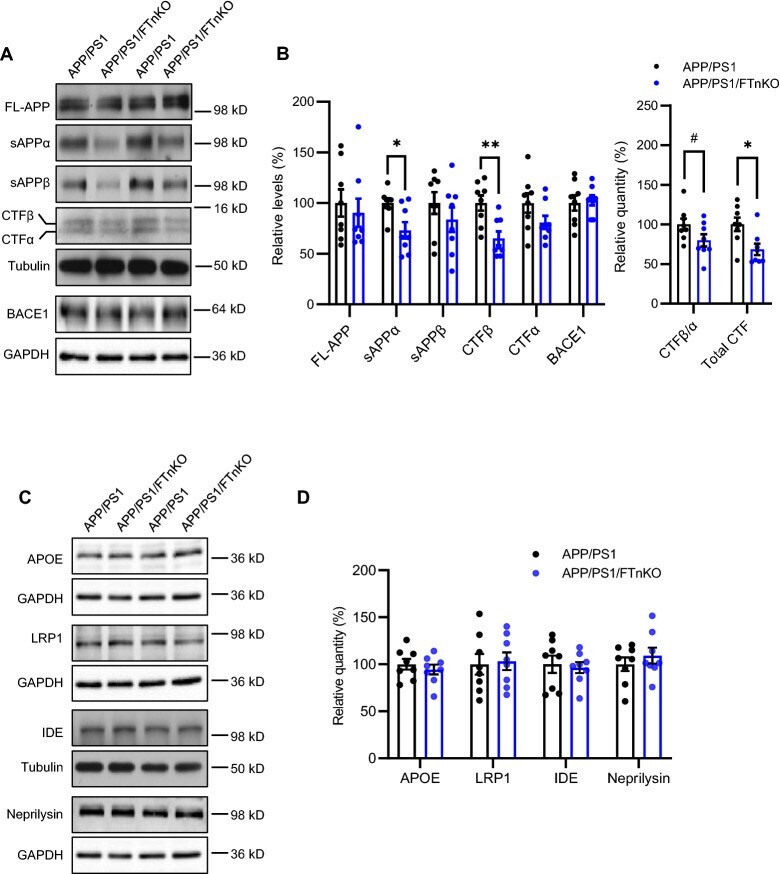
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
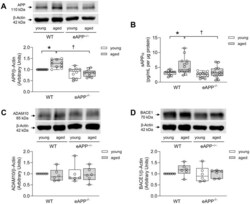
- Experimental details
- Figure 1 ( A ) Effects of aging on protein expression of APP in the aortas of wild-type (WT) littermates and eAPP -/- mice (n=10 per group). ( B ) Effects of aging on ex-vivo sAPPalpha secretion from wild-type (WT) littermates and eAPP -/- mice aortas. The supernatants were collected and analyzed for sAPPalpha levels. Results were normalized against tissue protein levels (n=12 per group for young WT littermates and eAPP -/- mice and n=15 per group for aged WT littermates and eAPP -/- mice). ( C ) Effects of aging on protein expression of ADAM10 in the aortas of wild-type (WT) littermates and eAPP -/- mice (n=6 per group). ( D ) Effects of aging on protein expression of BACE1 in the aortas of wild-type (WT) littermates and eAPP -/- mice (n=6 per group). Western blot results are the relative densitometry compared with beta-actin protein. All results are representing box plots with whiskers showing the median, 25 th to 75 th percentiles, and min-max range. * P
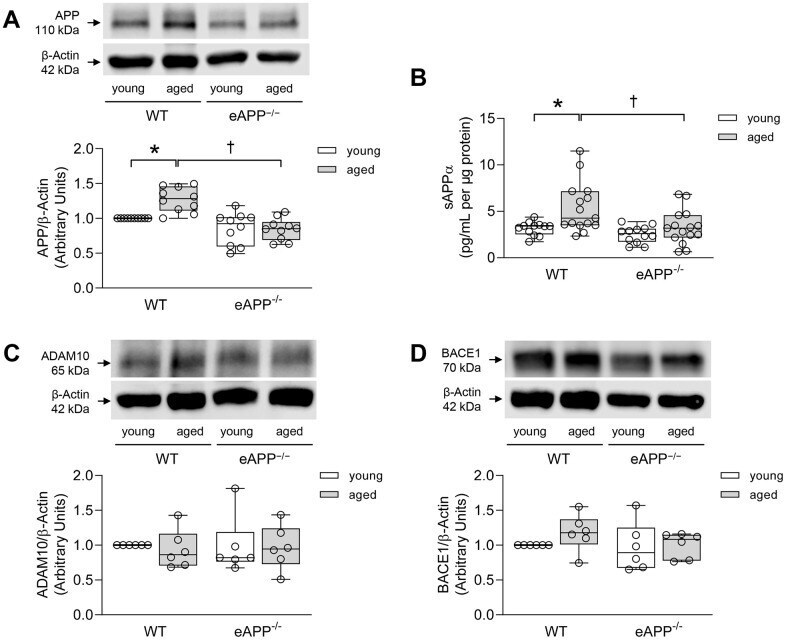
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
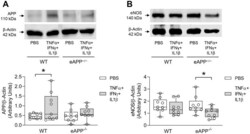
- Experimental details
- Figure 9 Effects of ex-vivo treatment for 24 hours with cytokines cocktail on protein expressions of APP ( A ) and eNOS ( B ) in the aortas of young wild-type (WT) littermates and eAPP -/- mice. Western blot results are the relative densitometry compared with beta-actin protein (n=9 per group). All results are representing box plots with whiskers showing the median, 25 th to 75 th percentiles, and min-max range. * P
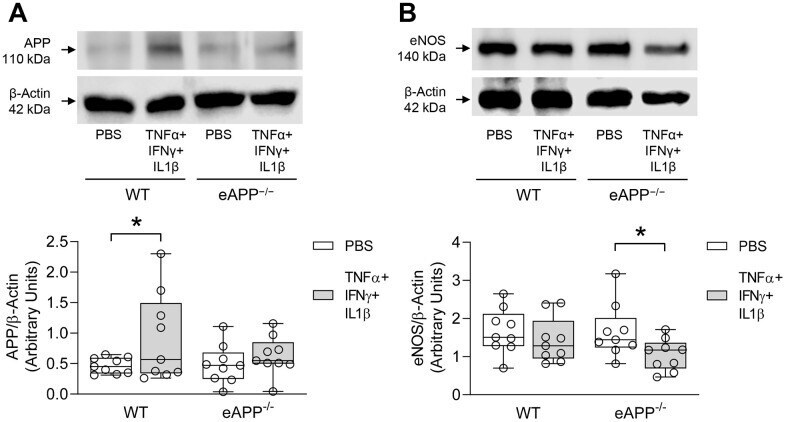
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
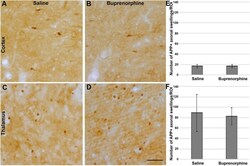
- Experimental details
- Figure 3 Buprenorphine did not alter acute diffuse axonal injury in either the thalamus or cortex. Representative photomicrographs of ( A , C ) saline or ( B , D ) buprenorphine treated Sprague Dawley rats 1d following cFPI. Tissue samples of ( A , B ) somatosensory neocortex and ( C , D ) hemithalamus were labeled immunohistochemically with a primary antibody against amyloid precursor protein (APP) and a secondary biotinylated antibody followed by DAB reaction. Swellings were visualized and quantified to assess axonal injury in each brain region at 1d following cFPI and either saline or buprenorphine treatment. ( E , F ) Corresponding bar graphs depicting the number of swellings per region of interest in the ( E ) cortex and ( F ) thalamus. The number of swellings was consistent between saline and buprenorphine treated animals, indicating that buprenorphine does not affect axonal injury in observed regions following TBI. Figure was compiled using Adobe Photoshop CS., version 22.0 (2020), San Diego, CA. n = 6 rats/group; Mean +- SEM. scale = 50 mum.

- Submitted by
- Invitrogen Antibodies (provider)
- Main image
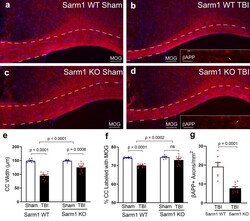
- Experimental details
- Fig. 3 Sarm1 knockout reduces corpus callosum atrophy, myelin loss, and axon damage at 10 weeks post-TBI. a - d Representative images from CC coronal sections of Sarm1 WT and Sarm1 KO mice after sham or TBI procedures. Myelin is detected with immunolabeling for MOG (red). DAPI nuclear stain is shown in blue. Insets show axonal swellings with beta-APP immunoreactivity (examples indicated by arrows). CC borders are indicated by dashed lines. e Sarm1 knockout attenuates CC atrophy, which is quantified based on the CC width. f TBI results in significant myelin loss as detected by reduced MOG immunoreactivity. Myelin loss after TBI is significantly reduced in Sarm1 KO mice compared to Sarm1 WT mice. g Axon damage in TBI mice was significantly reduced in Sarm1 KO mice compared to Sarm1 WT mice. MOG quantification included Sarm1 WT: n = 7 sham, n = 11 TBI; Sarm1 KO: n = 6 sham, n = 11 TBI. beta-APP quantification included Sarm1 WT: n = 6 TBI; Sarm1 KO: n = 9 TBI. ns = not significant. Further statistical details are provided in Additional File 1 : Table S3. Scale bars a - d = 100 um, insets b , d = 25 um
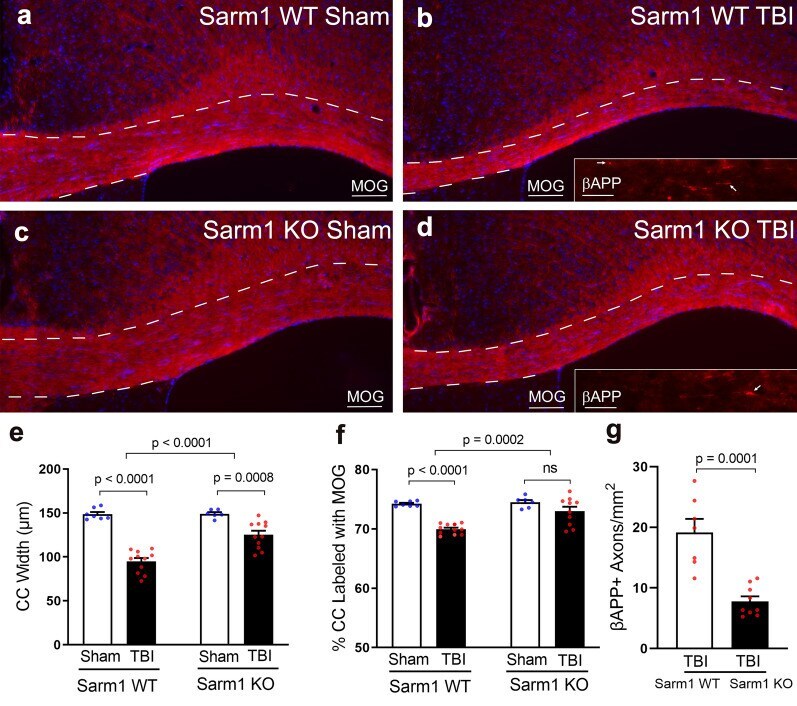
- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
- Fig. 2 Hyperoxygenation treatment increased the expression of MMP-2, MMP-9, and tPA in the hippocampus of Tg-APP/PS1 mice. (A) Real-time PCR data showing transcript levels of Bace1 , Bace2 , Psen1 , Psen2 , Adam10 , and Apoe genes in the hippocampus of wildtype control (WT-CON), wildtype mice exposed to hyperoxygenation (WT-HO 2 ), Tg-APP/PS1 control mice (Tg-CON), and Tg-APP/PS1 mice exposed to hyperoxygenation (Tg-HO 2 ). n=6-7 animals/group, 4 PCR repeats. (C) Western blot data showing expression levels of APP, BACE1, PSEN1, and ApoE in the hippocampus of WT-CON, WT-HO 2 , Tg-CON, and Tg-HO 2 mice. n=6~8 animals/group, 4~11 repeats. (D) Real-time PCR data showing transcript levels of Mmp-2 , Mmp-9 , tPA , Ide , Nep , and Lrp1 genes in the hippocampus of WT-CON, WT-HO 2 , Tg-CON, and Tg-HO 2 mice. n=6~7 animals, 4 data points. (E) Western blot data showing expression levels of MMP-2, MMP-9, tPA, uPA, and Ide genes in the hippocampus of WT-CON, WT-HO 2 , Tg-CON, and Tg-HO 2 mice. Tissue samples were prepared at the time point of Figure 1A . n=6~8 animals/group, 4~10 data points. Data are presented as mean+-SEM. * , **difference between indicated groups. *p

- Submitted by
- Invitrogen Antibodies (provider)
- Main image
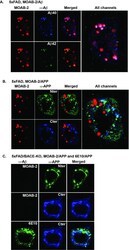
- Experimental details
- Figure 3 MOAB-2 detection of intraneuronal Abeta but not intraneuronal APP in 5xFAD brain tissue . Immunofluorescent detection of Abeta and APP in the cortex of: (A) 1-month old 5xFAD mice using: MOAB-2, Abeta42- or Abeta40-specific antibodies; (B) 1-month old 5xFAD mice with MOAB-2 and antibodies against the C-terminus (Cter) (CT695) or N-terminus (Nter) (2211) of APP; and (C) 4-month old 5xFAD/BACE -/- with 6E10 or MOAB-2, plus antibodies CT695 and 22C11. (Coronal sections, representative confocal images at 100x). Antibody concentrations: MOAB-2 (1:1000), anti-Abeta40, (1:1700), anti-Abeta42 (1:200), 22C11 (1:40), CT695 (1:500), 6E10 (1:1000).
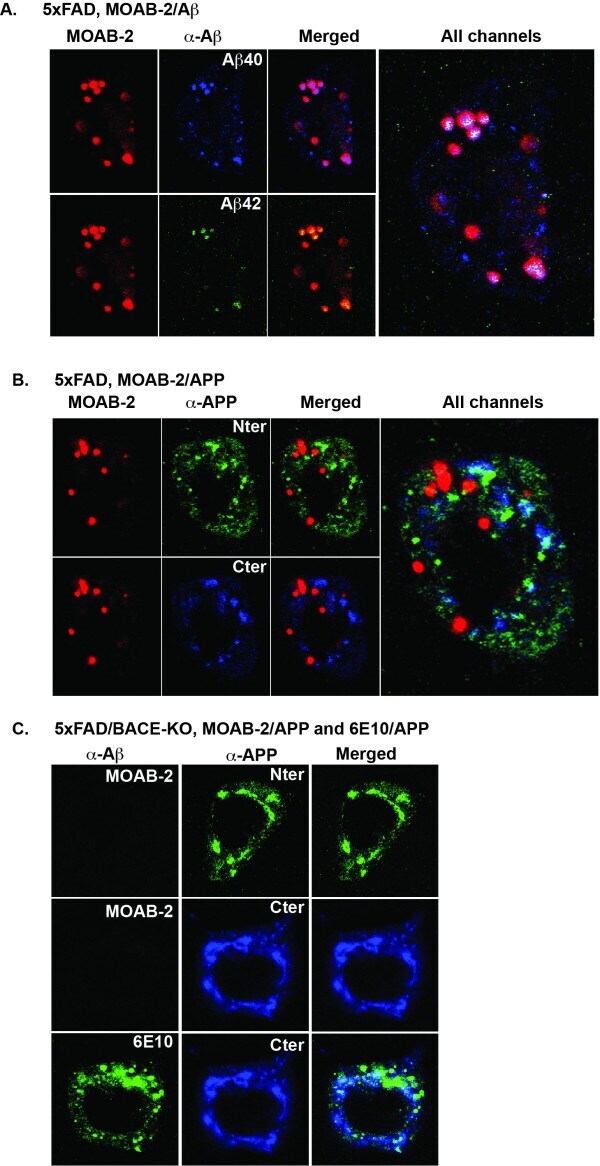
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- Figure 2 Curcumin analog C1 activates transcription factor EB (TFEB), degrades beta-amyloid precursor protein (APP) and beta-amyloid peptides (Abeta), and prevents synaptic and cognitive failures in 5xFAD mice. (a, b) Western blots showed the levels of full-length APP (Fl-APP), CTF-beta/alpha, Abeta, LAMP1, CTSD, LC3B, cytosolic and nuclear TFEB (cTFEB and nTFEB) with their respective loading controls in the brain lysates of male mice (Another batch of blots for male mice and all blots for female mice were shown in Figure S2 B). (c, d) All the values from male ( n = 6) and female ( n = 6) mice were quantified as average +- SEM . * p < .05, ** p < .01, and *** p < .001 vs. Veh. group, respectively, analyzed by one-way ANOVA. The interaction between sex and drug effects was analyzed by two-way ANOVA, and the P values were summarized in Table S1 . (e, f) Immunohistochemistry analysis of Abeta load. (e) Representative images show Abeta labeled with biotinylated 4G8 antibody. (f) Data are quantified as mean +- SEM in male ( n = 6) and female ( n = 6) mice. ** p < .01 and *** p < .001 vs. Veh. group, respectively. (g) ELISA assay of Abeta42 and Abeta40 in male ( n = 6) and female ( n = 6) mice. Whole-brain samples were separated into Triton soluble and formic acid (FA) fractions. Data were presented as mean +- SEM . # p < .05, ## p < .01, and ### p < .001 vs. Veh. group (Abeta42); * p < .05, ** p < .01, and *** p < .001 vs. Veh. group (Abeta40). The interaction between sex and drug

- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
- Figure 4 Curcumin analog C1 activates transcription factor EB (TFEB) and attenuates both beta-amyloid precursor protein (APP) and Tau pathology in 3xTg mice. (a, b) Representative blots showed the levels of Fl-APP, CTF-beta/alpha, phosphorylated Tau (PHF-1, AT-8), total Tau, LAMP1, CTSD, cytosolic and nuclear TFEB (cTFEB and nTFEB), and LC3B with their respective loading controls in the hippocampus of female mice (Another batch of blots were shown in Figure S3 A). (c, d) Data are quantified as mean +- SEM ( n = 6). * p < .05, ** p < .01, *** p < .001 vs. Veh. treatment analyzed by one-way ANOVA. (e, f) Immunohistochemistry analysis of beta-amyloid peptides (Abeta) load. (e) Representative images show the Abeta labeled with biotinylated 4G8 antibody. (f) Data are quantified as mean +- SEM ( n = 10). * p < .05 vs. Veh. treatment analyzed by one-way ANOVA. (g, h) Immunohistochemistry analysis of p-Tau (AT8). (g) Representative images show AT8 staining in the posterior hippocampus. (h) Data are quantified as mean +- SEM ( n = 7-12). * p < .05, ** p < .01 vs. Veh. treatment analyzed by one-way ANOVA. CTSD, cathepsin D; LAMP, lysosomal-associated membrane protein 1

- Submitted by
- Invitrogen Antibodies (provider)
- Main image
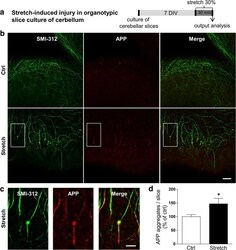
- Experimental details
- Fig. 1 Mechanical tensile strain of 30% induces axonal injury in organotypic cerebellar slice culture. a Ex vivo culture and 30% strain model using organotypic slice cultures of cerebellum. b Double immunostaining of organotypic cerebellar slices for APP in red and SMI-312 (pan-axonal neurofilament marker) in green. Images represent control (Ctrl) and stretched (Stretch) cultures. The response of slices was evaluated at time 0 h post-stretch. Scale bar, 50 mum. c High magnification of the white square in the stretch panel from b . Scale bar, 20 mum. d Quantitative analysis of the number of APP aggregates per slice. Results represent the mean +- SEM ( n = 3). * p < 0.05. APP amyloid precursor protein, DIV days in vitro
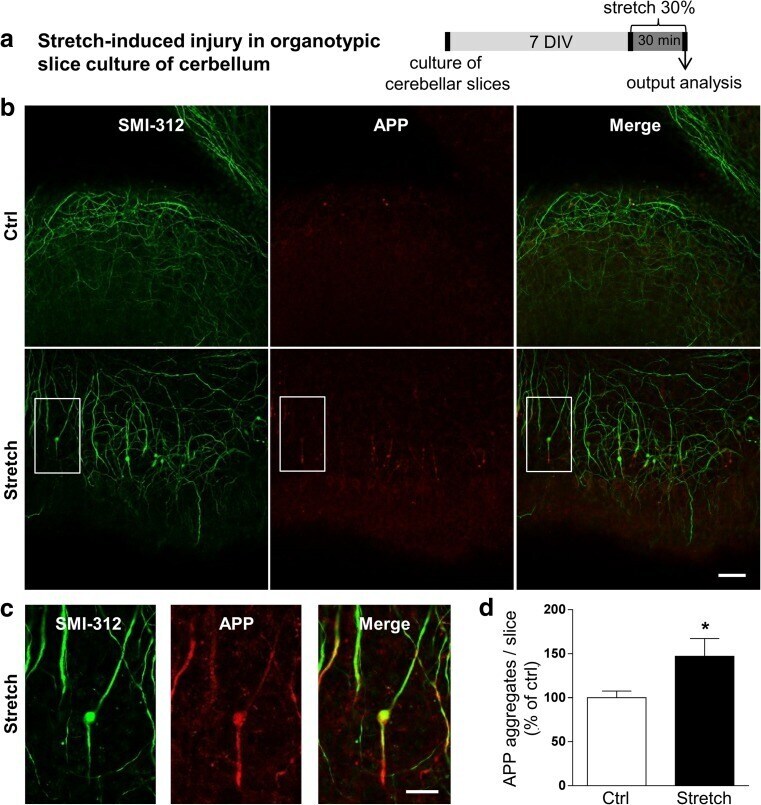
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
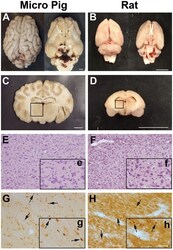
- Experimental details
- Figure 1 Central fluid percussion injury does not result in focal brain damage, but does precipitate diffuse axonal injury in the thalamus of both rats and micro pigs 1 day following injury. Representative photographs of the gross (A,C) micro pig and (B,D) rat brain 1 day following cFPI. The top panels are dorsal and ventral views of the entire brain 1 day post-injury while panels (C) and (D) represent 5 mm thick coronal sections at the level of the rostral thalamus in the (C) micro pig and (D) rat. The boxes indicate the regions of analysis of microglia process convergence in the thalamus of both species. Diffuse cFPI also did not result in cell damage/death. Hematoxylin and eosin staining reveled no square wave cellular damage in the thalamus of either the (E) micro pig or (F) rat thalamus at 1 day following cFPI. Amyloid precursor protein (APP) immunohistochemistry in the (G) micro pig and (H) rat thalamus, however, demonstrated diffuse axonal injury (arrows) 1 day following cFPI. Insets (e-h) depict higher magnification images of panels (E-H) , respectively. Note that the cFPI model employed did not result in contusion, hematoma formation or square wave cell damage/death, however did demonstrate diffusely distributed axonal injury, consistent with diffuse injury. Scale bar A-D : 1 mm; E-H : 100 um; e-h: 50 um.
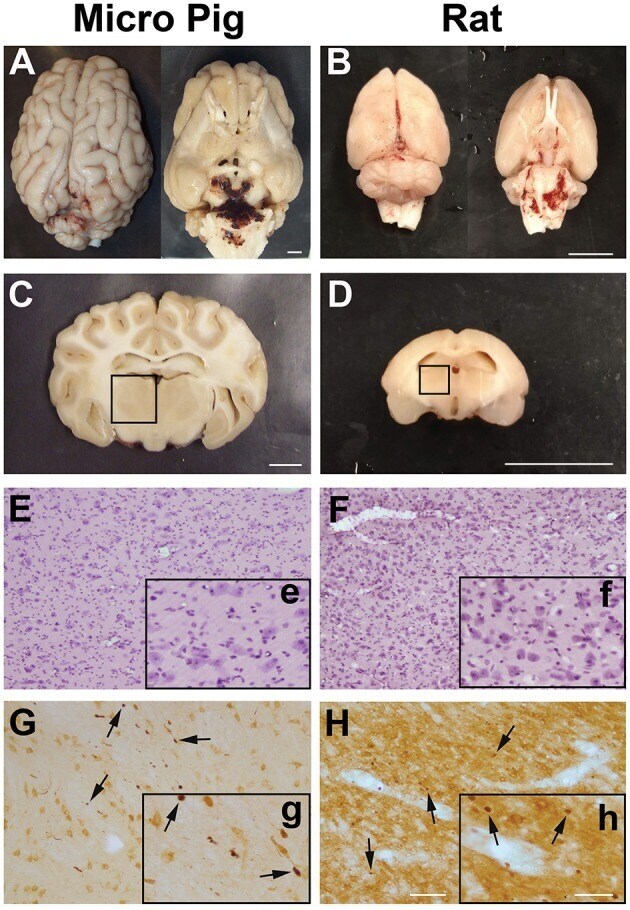
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
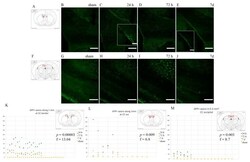
- Experimental details
- Figure 3 APP expression in the corpus callosum. Fluorescence microscopy photographs of the corpus callosum (coronal sections) at around 1mm from bregma (top row) and around -3 mm from bregma (bottom row). ( A , F ) The highlighted area refers to the area represented in the photographs on the same row. ( C - E ) Following injury, axonal bulbs localize mostly at the borders between the corpus callosum and grey matter. Insert show details of the CC-lateral ventricle border area. ( H - J , M ) Moving occipitally, APP expression is comparatively less. ( B , G ) Sham animals do not show APP accumulation in the axonal terminals at the regions of interest. ( K - M ) Scatter plots showing the amount of APP+ profiles in each ROI (y-axis represents the number of APP+ profiles; x-axis represents each observation). APP expression is maximal at 24 h and decreases thereafter, with p
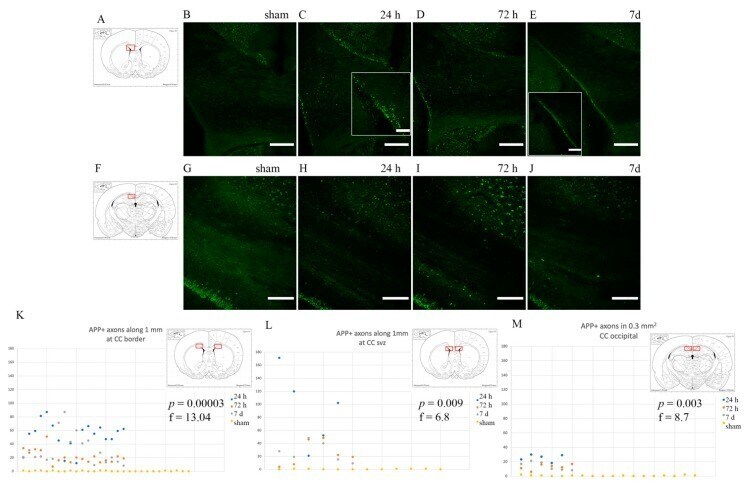
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- Figure 4 Olig2 expression in the corpus callosum. ( A , F ) The highlighted area refers to the area represented in the photographs on the same row. ( B - E ) Olig2 expression in the corpus callosum, at its interface with the lateral ventricle at around 1 mm from bregma. ( G - J ) Olig2 expression in the corpus callosum at around -3 mm from bregma. Scale bar = 200 um.
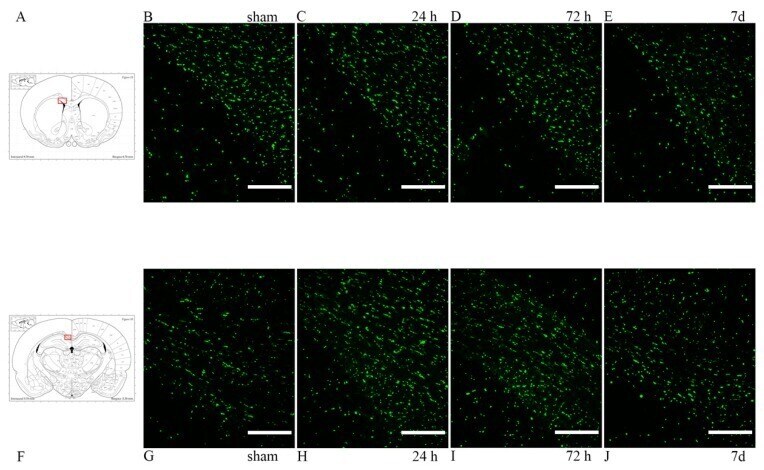
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
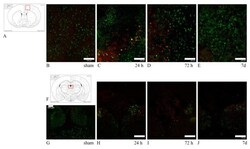
- Experimental details
- Figure 12 HSP70/ATF3/APP expression reflecting neuronal stress. ( A , F ) The highlighted area refers to the area represented in each row. ( B - E ) HSP70 (red) and APP (green) double staining in the dorsal cortex. While APP expression remains elevated through 7 d, HSP70 expression is comparatively decreased at 7 d. Few neurons are HSP70+/APP+. ( G - J ) ATF3 (red) and APP (green) double staining in the habenular nuclei. ATF3 follows the same expression pattern as HSP70, with decreased marker uptake at 7 d. In all cases, sham animals do not express any of the tested markers, with only the baseline marker uptake visible. Scale bar = 150 um (Scale bar = 100 um in image ( F )).
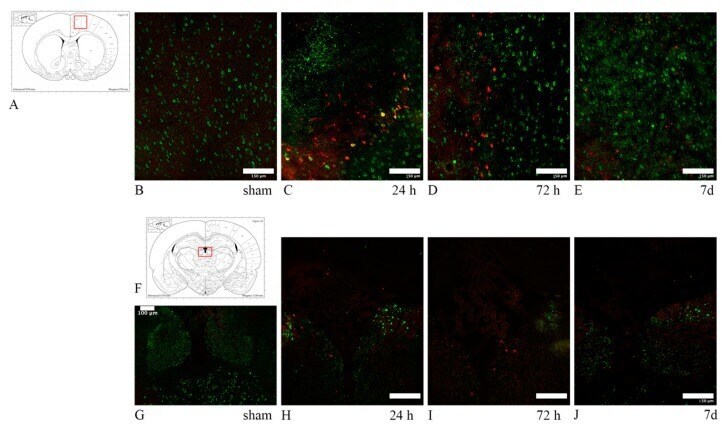
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
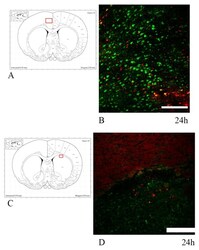
- Experimental details
- Figure 13 HSP70/APP double-staining in 24 h trauma-exposed animals. ( A , C ) The highlighted area refers to the area represented in the adjacent microphotograph. ( B ) HSP70 and APP expression in the cingulate cortex. Few neurons are positive for both markers. ( D ) HSP70 and APP expression in the caudoputamen, close to its interface with the corpus callosum. Scale bar = 200 um.