Explore
Explore Validate
Validate Learn
Learn Western blot
Western blotAntibody data
- Antibody Data
- Antigen structure
- References [60]
- Comments [0]
- Validations
- Western blot [3]
- Immunohistochemistry [3]
- Other assay [35]
Submit
Validation data
Reference
Comment
Report error
- Product number
- 44-680G - Provider product page

- Provider
- Invitrogen Antibodies
- Product name
- Phospho-ERK1/ERK2 (Thr185, Tyr187) Polyclonal Antibody
- Antibody type
- Polyclonal
- Antigen
- Synthetic peptide
- Description
- Purified from rabbit serum by sequential epitope-specific chromatography, this product contains enough material for 10 mini-blots. The antibody has been negatively preadsorbed using a non-phosphopeptide corresponding to the sites of phosphorylation to remove antibody that is reactive with non-phosphorylated ERK1&2. The final product is generated by affinity chromatography using an ERK1&2-derived peptide that is phosphorylated at threonine 202/185 and tyrosine 204/187, respectively, within the activation loop.
Submitted references Sialylation of TLR2 initiates osteoclast fusion.
Hematopoietic Cell Kinase (HCK) Is a Player of the Crosstalk Between Hematopoietic Cells and Bone Marrow Niche Through CXCL12/CXCR4 Axis.
Response of human cancer cells to simultaneous treatment with sorafenib and radiofrequency current.
Leptin reduces in vitro cementoblast mineralization and survival as well as induces PGE2 release by ERK1/2 commitment.
Adiponectin Interacts In-Vitro With Cementoblasts Influencing Cell Migration, Proliferation and Cementogenesis Partly Through the MAPK Signaling Pathway.
Role of thiocyanate in the modulation of myeloperoxidase-derived oxidant induced damage to macrophages.
A novel protective role of sacubitril/valsartan in cyclophosphamide induced lung injury in rats: impact of miRNA-150-3p on NF-κB/MAPK signaling trajectories.
Involvement of the EGF Receptor in MAPK Signaling Activation by a 50 Hz Magnetic Field in Human Neuroblastoma Cells.
Simvastatin Mitigates Apoptosis and Transforming Growth Factor-Beta Upregulation in Stretch-Induced Endothelial Cells.
ANO6 promotes cell proliferation and invasion in glioma through regulating the ERK signaling pathway.
Repeated Sigma-1 Receptor Antagonist MR309 Administration Modulates Central Neuropathic Pain Development After Spinal Cord Injury in Mice.
Response of neuroblastoma cells to RF currents as a function of the signal frequency.
KLF9-dependent ROS regulate melanoma progression in stage-specific manner.
The role of fibroblast growth factor signalling in Echinococcus multilocularis development and host-parasite interaction.
Intermittent hypoxia alleviates increased VEGF and pro-angiogenic potential in liver cancer cells.
CRB3 and the FERM protein EPB41L4B regulate proliferation of mammary epithelial cells through the release of amphiregulin.
ZO-1 protein is required for hydrogen peroxide to increase MDCK cell paracellular permeability in an ERK 1/2-dependent manner.
FGF21 acts as a negative regulator of bile acid synthesis.
Critical role of sigma-1 receptors in central neuropathic pain-related behaviours after mild spinal cord injury in mice.
Development of MK-8353, an orally administered ERK1/2 inhibitor, in patients with advanced solid tumors.
Hematopoietic cell kinase (HCK) is a potential therapeutic target for dysplastic and leukemic cells due to integration of erythropoietin/PI3K pathway and regulation of erythropoiesis: HCK in erythropoietin/PI3K pathway.
Targeting the PI3K/AKT/mTOR pathway overcomes the stimulating effect of dabrafenib on the invasive behavior of melanoma cells with acquired resistance to the BRAF inhibitor.
WNK1 kinase balances T cell adhesion versus migration in vivo.
Haem-dependent dimerization of PGRMC1/Sigma-2 receptor facilitates cancer proliferation and chemoresistance.
Oxidative pentose phosphate pathway inhibition is a key determinant of antimalarial induced cancer cell death.
Anti-c-Met monoclonal antibody ABT-700 breaks oncogene addiction in tumors with MET amplification.
Endosomal Na+/H+ exchanger NHE5 influences MET recycling and cell migration.
ERK and p38 MAPK Activities Determine Sensitivity to PI3K/mTOR Inhibition via Regulation of MYC and YAP.
Power Frequency Magnetic Fields Affect the p38 MAPK-Mediated Regulation of NB69 Cell Proliferation Implication of Free Radicals.
All-Trans Retinoic Acid Induces Proliferation, Survival, and Migration in A549 Lung Cancer Cells by Activating the ERK Signaling Pathway through a Transcription-Independent Mechanism.
A rapid and convenient method for fluorescence analysis of in vitro cultivated metacestode vesicles from Echinococcus multilocularis.
A new water soluble MAPK activator exerts antitumor activity in melanoma cells resistant to the BRAF inhibitor vemurafenib.
ATG12-ATG3 interacts with Alix to promote basal autophagic flux and late endosome function.
Candidate tumor suppressor B-cell translocation gene 3 impedes neoplastic progression by suppression of AKT.
Possible Mechanisms of Di(2-ethylhexyl) Phthalate-Induced MMP-2 and MMP-9 Expression in A7r5 Rat Vascular Smooth Muscle Cells.
The MEK1/2-ERK1/2 pathway is activated in chronic rhinosinusitis with nasal polyps.
Increased ERK and JNK activation and decreased ERK/JNK ratio are associated with long-term organ damage in patients with systemic lupus erythematosus.
Silver and fullerene nanoparticles' effect on interleukin-2-dependent proliferation of CD4 (+) T cells.
The C-terminus of human nucleotide receptor P2X7 is critical for receptor oligomerization and N-linked glycosylation.
Molecular pathway profiling of T lymphocyte signal transduction pathways; Th1 and Th2 genomic fingerprints are defined by TCR and CD28-mediated signaling.
TFG-1 function in protein secretion and oncogenesis.
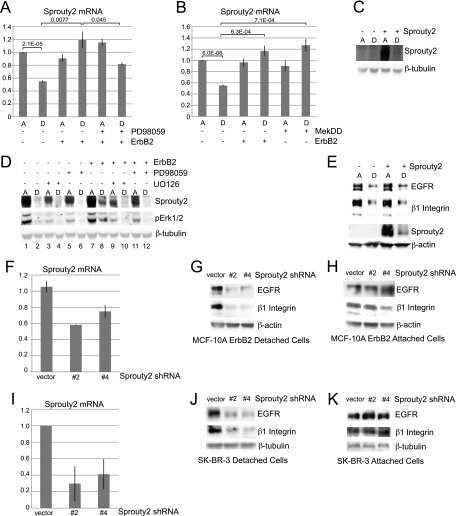
ErbB2 stabilizes epidermal growth factor receptor (EGFR) expression via Erk and Sprouty2 in extracellular matrix-detached cells.
Extranuclear estrogen receptors mediate the neuroprotective effects of estrogen in the rat hippocampus.
3-phosphoinositide-dependent protein kinase-1 regulates proliferation and survival of cancer cells with an activated mitogen-activated protein kinase pathway.
Mutation of putative N-linked glycosylation sites on the human nucleotide receptor P2X7 reveals a key residue important for receptor function.
Induction of invasion in an organotypic oral cancer model by CoCl2, a hypoxia mimetic.
The absence of caveolin-1 increases proliferation and anchorage- independent growth by a Rac-dependent, Erk-independent mechanism.
Antioxidant and oncogene rescue of metabolic defects caused by loss of matrix attachment.
A study of phosphorylated ERK1/2 and COX-2 in early stage (T1-T2) oral squamous cell carcinomas.
Dissection of the osteogenic effects of laminin-332 utilizing specific LG domains: LG3 induces osteogenic differentiation, but not mineralization.
Quantitative measurement of epidermal growth factor receptor-mitogen-activated protein kinase signal transduction using a nine-plex, peptide-based immunoassay.
Molecular mechanisms involved in interleukin-4-induced human neutrophils: expression and regulation of suppressor of cytokine signaling.
Activation of FAK is necessary for the osteogenic differentiation of human mesenchymal stem cells on laminin-5.
The mitogen-activated protein kinases (MAPK) p38 and JNK are markers of tumor progression in breast carcinoma.
Epidermal growth factor receptor induced Erk phosphorylation in the suprachiasmatic nucleus.
Phospho-caveolin-1 mediates integrin-regulated membrane domain internalization.
Heat shock protein 60 inhibits Th1-mediated hepatitis model via innate regulation of Th1/Th2 transcription factors and cytokines.
Mitogen-activated protein kinase expression and activation does not differentiate benign from malignant mesothelial cells.
CHIR-258: a potent inhibitor of FLT3 kinase in experimental tumor xenograft models of human acute myelogenous leukemia.
Chemokine receptor CCR2 expression by systemic sclerosis fibroblasts: evidence for autocrine regulation of myofibroblast differentiation.
Dou C, Zhen G, Dan Y, Wan M, Limjunyawong N, Cao X
Bone research 2022 Mar 2;10(1):24
Bone research 2022 Mar 2;10(1):24
Hematopoietic Cell Kinase (HCK) Is a Player of the Crosstalk Between Hematopoietic Cells and Bone Marrow Niche Through CXCL12/CXCR4 Axis.
Roversi FM, Bueno MLP, Pericole FV, Saad STO
Frontiers in cell and developmental biology 2021;9:634044
Frontiers in cell and developmental biology 2021;9:634044
Response of human cancer cells to simultaneous treatment with sorafenib and radiofrequency current.
Hernández-Bule ML, Martínez MA, Trillo MÁ, Martínez L, Toledano-Macías E, Úbeda A
Oncology letters 2021 Nov;22(5):807
Oncology letters 2021 Nov;22(5):807
Leptin reduces in vitro cementoblast mineralization and survival as well as induces PGE2 release by ERK1/2 commitment.
Ruiz-Heiland G, Yong JW, von Bremen J, Ruf S
Clinical oral investigations 2021 Apr;25(4):1933-1944
Clinical oral investigations 2021 Apr;25(4):1933-1944
Adiponectin Interacts In-Vitro With Cementoblasts Influencing Cell Migration, Proliferation and Cementogenesis Partly Through the MAPK Signaling Pathway.
Yong J, von Bremen J, Ruiz-Heiland G, Ruf S
Frontiers in pharmacology 2020;11:585346
Frontiers in pharmacology 2020;11:585346
Role of thiocyanate in the modulation of myeloperoxidase-derived oxidant induced damage to macrophages.
Guo C, Davies MJ, Hawkins CL
Redox biology 2020 Sep;36:101666
Redox biology 2020 Sep;36:101666
A novel protective role of sacubitril/valsartan in cyclophosphamide induced lung injury in rats: impact of miRNA-150-3p on NF-κB/MAPK signaling trajectories.
Abdel-Latif GA, Elwahab AHA, Hasan RA, ElMongy NF, Ramzy MM, Louka ML, Schaalan MF
Scientific reports 2020 Aug 3;10(1):13045
Scientific reports 2020 Aug 3;10(1):13045
Involvement of the EGF Receptor in MAPK Signaling Activation by a 50 Hz Magnetic Field in Human Neuroblastoma Cells.
Martínez MA, Úbeda A, Trillo MÀ
Cellular physiology and biochemistry : international journal of experimental cellular physiology, biochemistry, and pharmacology 2019;52(4):893-907
Cellular physiology and biochemistry : international journal of experimental cellular physiology, biochemistry, and pharmacology 2019;52(4):893-907
Simvastatin Mitigates Apoptosis and Transforming Growth Factor-Beta Upregulation in Stretch-Induced Endothelial Cells.
Dong G, Huang X, Jiang S, Ni L, Chen S
Oxidative medicine and cellular longevity 2019;2019:6026051
Oxidative medicine and cellular longevity 2019;2019:6026051
ANO6 promotes cell proliferation and invasion in glioma through regulating the ERK signaling pathway.
Xuan ZB, Wang YJ, Xie J
OncoTargets and therapy 2019;12:6721-6731
OncoTargets and therapy 2019;12:6721-6731
Repeated Sigma-1 Receptor Antagonist MR309 Administration Modulates Central Neuropathic Pain Development After Spinal Cord Injury in Mice.
Castany S, Codony X, Zamanillo D, Merlos M, Verdú E, Boadas-Vaello P
Frontiers in pharmacology 2019;10:222
Frontiers in pharmacology 2019;10:222
Response of neuroblastoma cells to RF currents as a function of the signal frequency.
Hernández-Bule ML, Medel E, Colastra C, Roldán R, Úbeda A
BMC cancer 2019 Sep 5;19(1):889
BMC cancer 2019 Sep 5;19(1):889
KLF9-dependent ROS regulate melanoma progression in stage-specific manner.
Bagati A, Moparthy S, Fink EE, Bianchi-Smiraglia A, Yun DH, Kolesnikova M, Udartseva OO, Wolff DW, Roll MV, Lipchick BC, Han Z, Kozlova NI, Jowdy P, Berman AE, Box NF, Rodriguez C, Bshara W, Kandel ES, Soengas MS, Paragh G, Nikiforov MA
Oncogene 2019 May;38(19):3585-3597
Oncogene 2019 May;38(19):3585-3597
The role of fibroblast growth factor signalling in Echinococcus multilocularis development and host-parasite interaction.
Förster S, Koziol U, Schäfer T, Duvoisin R, Cailliau K, Vanderstraete M, Dissous C, Brehm K
PLoS neglected tropical diseases 2019 Mar;13(3):e0006959
PLoS neglected tropical diseases 2019 Mar;13(3):e0006959
Intermittent hypoxia alleviates increased VEGF and pro-angiogenic potential in liver cancer cells.
Dong G, Lin XH, Liu HH, Gao DM, Cui JF, Ren ZG, Chen RX
Oncology letters 2019 Aug;18(2):1831-1839
Oncology letters 2019 Aug;18(2):1831-1839
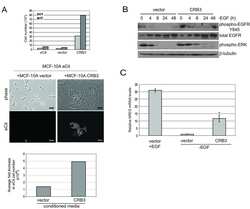
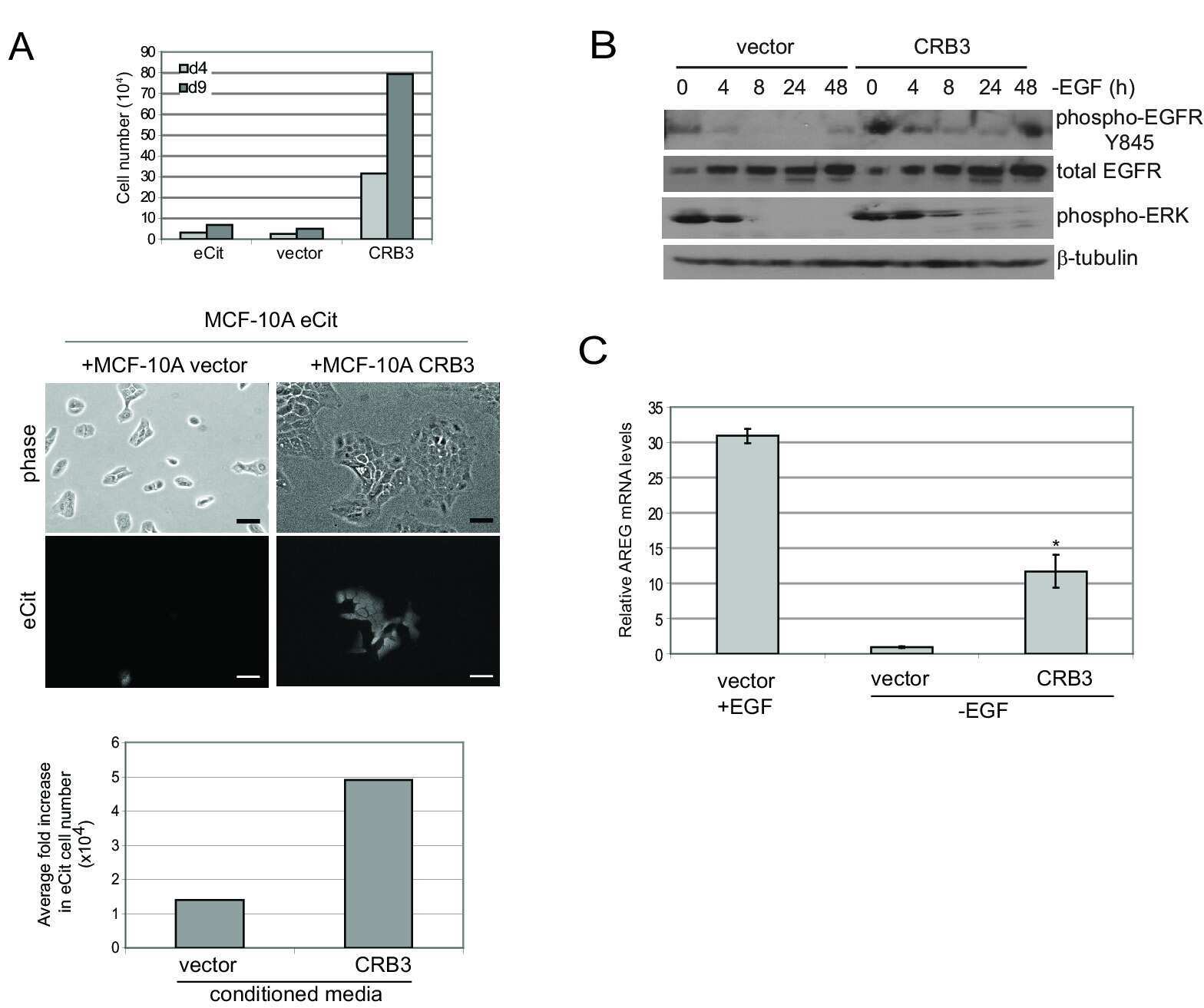
CRB3 and the FERM protein EPB41L4B regulate proliferation of mammary epithelial cells through the release of amphiregulin.
Walker SJ, Selfors LM, Margolis BL, Brugge JS
PloS one 2018;13(11):e0207470
PloS one 2018;13(11):e0207470
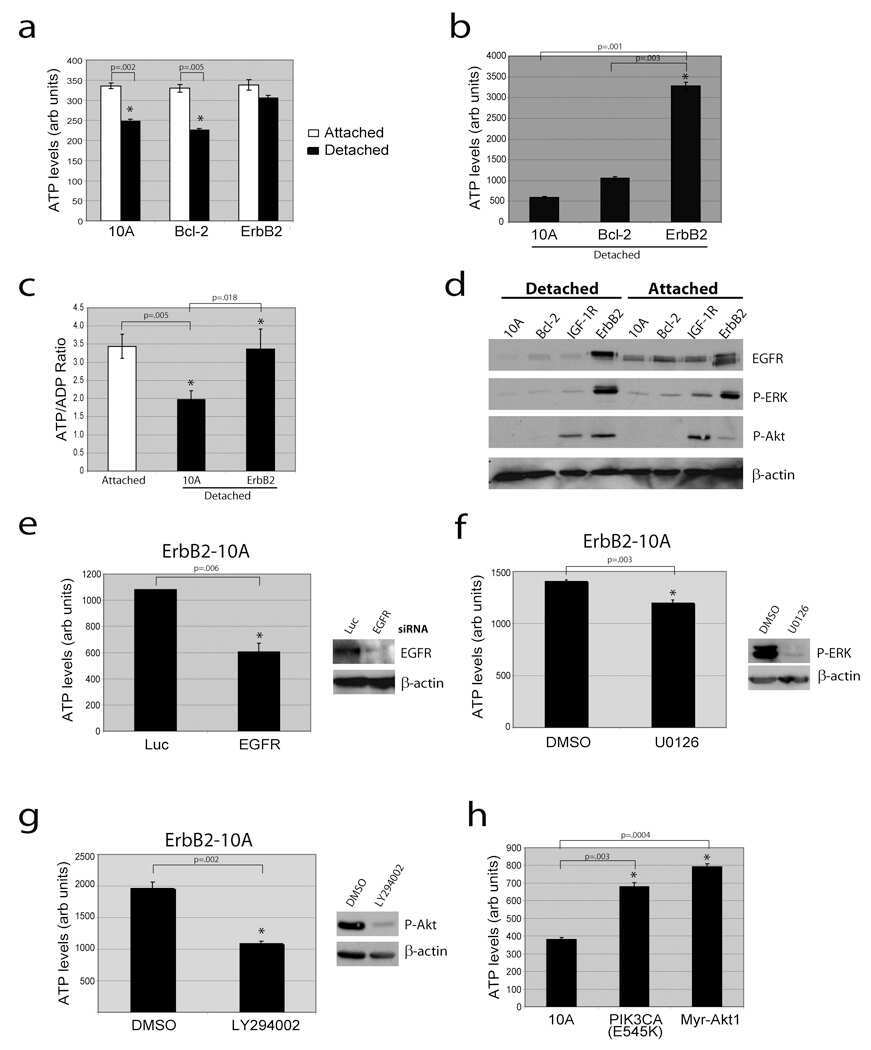
ZO-1 protein is required for hydrogen peroxide to increase MDCK cell paracellular permeability in an ERK 1/2-dependent manner.
Bilal S, Jaggi S, Janosevic D, Shah N, Teymour S, Voronina A, Watari J, Axis J, Amsler K
American journal of physiology. Cell physiology 2018 Sep 1;315(3):C422-C431
American journal of physiology. Cell physiology 2018 Sep 1;315(3):C422-C431
FGF21 acts as a negative regulator of bile acid synthesis.
Chen MM, Hale C, Stanislaus S, Xu J, Véniant MM
The Journal of endocrinology 2018 May;237(2):139-152
The Journal of endocrinology 2018 May;237(2):139-152
Critical role of sigma-1 receptors in central neuropathic pain-related behaviours after mild spinal cord injury in mice.
Castany S, Gris G, Vela JM, Verdú E, Boadas-Vaello P
Scientific reports 2018 Mar 1;8(1):3873
Scientific reports 2018 Mar 1;8(1):3873
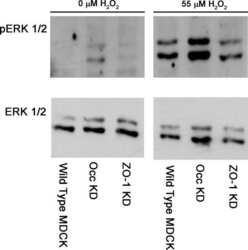
Development of MK-8353, an orally administered ERK1/2 inhibitor, in patients with advanced solid tumors.
Moschos SJ, Sullivan RJ, Hwu WJ, Ramanathan RK, Adjei AA, Fong PC, Shapira-Frommer R, Tawbi HA, Rubino J, Rush TS 3rd, Zhang D, Miselis NR, Samatar AA, Chun P, Rubin EH, Schiller J, Long BJ, Dayananth P, Carr D, Kirschmeier P, Bishop WR, Deng Y, Cooper A, Shipps GW, Moreno BH, Robert L, Ribas A, Flaherty KT
JCI insight 2018 Feb 22;3(4)
JCI insight 2018 Feb 22;3(4)
Hematopoietic cell kinase (HCK) is a potential therapeutic target for dysplastic and leukemic cells due to integration of erythropoietin/PI3K pathway and regulation of erythropoiesis: HCK in erythropoietin/PI3K pathway.
Roversi FM, Pericole FV, Machado-Neto JA, da Silva Santos Duarte A, Longhini AL, Corrocher FA, Palodetto B, Ferro KP, Rosa RG, Baratti MO, Verjovski-Almeida S, Traina F, Molinari A, Botta M, Saad ST
Biochimica et biophysica acta. Molecular basis of disease 2017 Feb;1863(2):450-461
Biochimica et biophysica acta. Molecular basis of disease 2017 Feb;1863(2):450-461
Targeting the PI3K/AKT/mTOR pathway overcomes the stimulating effect of dabrafenib on the invasive behavior of melanoma cells with acquired resistance to the BRAF inhibitor.
Caporali S, Alvino E, Lacal PM, Levati L, Giurato G, Memoli D, Caprini E, Antonini Cappellini GC, D'Atri S
International journal of oncology 2016 Sep;49(3):1164-74
International journal of oncology 2016 Sep;49(3):1164-74
WNK1 kinase balances T cell adhesion versus migration in vivo.
Köchl R, Thelen F, Vanes L, Brazão TF, Fountain K, Xie J, Huang CL, Lyck R, Stein JV, Tybulewicz VL
Nature immunology 2016 Sep;17(9):1075-83
Nature immunology 2016 Sep;17(9):1075-83
Haem-dependent dimerization of PGRMC1/Sigma-2 receptor facilitates cancer proliferation and chemoresistance.
Kabe Y, Nakane T, Koike I, Yamamoto T, Sugiura Y, Harada E, Sugase K, Shimamura T, Ohmura M, Muraoka K, Yamamoto A, Uchida T, Iwata S, Yamaguchi Y, Krayukhina E, Noda M, Handa H, Ishimori K, Uchiyama S, Kobayashi T, Suematsu M
Nature communications 2016 Mar 18;7:11030
Nature communications 2016 Mar 18;7:11030
Oxidative pentose phosphate pathway inhibition is a key determinant of antimalarial induced cancer cell death.
Salas E, Roy S, Marsh T, Rubin B, Debnath J
Oncogene 2016 Jun 2;35(22):2913-22
Oncogene 2016 Jun 2;35(22):2913-22
Anti-c-Met monoclonal antibody ABT-700 breaks oncogene addiction in tumors with MET amplification.
Wang J, Goetsch L, Tucker L, Zhang Q, Gonzalez A, Vaidya KS, Oleksijew A, Boghaert E, Song M, Sokolova I, Pestova E, Anderson M, Pappano WN, Ansell P, Bhathena A, Naumovski L, Corvaia N, Reilly EB
BMC cancer 2016 Feb 16;16:105
BMC cancer 2016 Feb 16;16:105
Endosomal Na+/H+ exchanger NHE5 influences MET recycling and cell migration.
Fan SH, Numata Y, Numata M
Molecular biology of the cell 2016 Feb 15;27(4):702-15
Molecular biology of the cell 2016 Feb 15;27(4):702-15
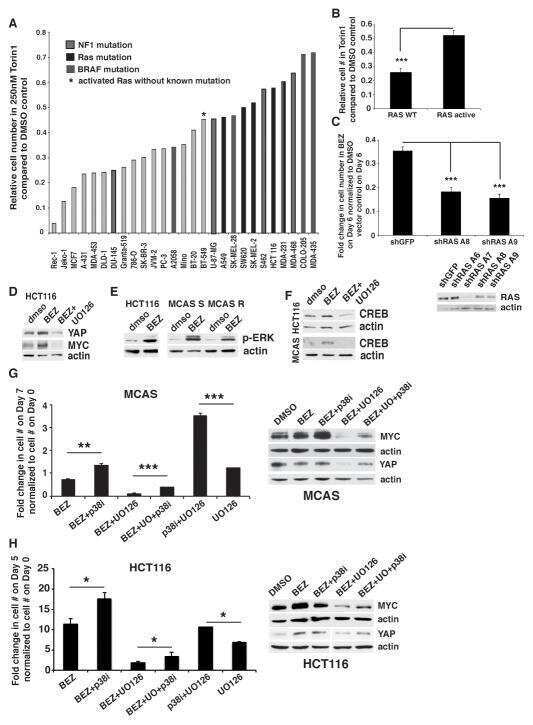
ERK and p38 MAPK Activities Determine Sensitivity to PI3K/mTOR Inhibition via Regulation of MYC and YAP.
Muranen T, Selfors LM, Hwang J, Gallegos LL, Coloff JL, Thoreen CC, Kang SA, Sabatini DM, Mills GB, Brugge JS
Cancer research 2016 Dec 15;76(24):7168-7180
Cancer research 2016 Dec 15;76(24):7168-7180
Power Frequency Magnetic Fields Affect the p38 MAPK-Mediated Regulation of NB69 Cell Proliferation Implication of Free Radicals.
Martínez MA, Úbeda A, Moreno J, Trillo MÁ
International journal of molecular sciences 2016 Apr 6;17(4):510
International journal of molecular sciences 2016 Apr 6;17(4):510
All-Trans Retinoic Acid Induces Proliferation, Survival, and Migration in A549 Lung Cancer Cells by Activating the ERK Signaling Pathway through a Transcription-Independent Mechanism.
Quintero Barceinas RS, García-Regalado A, Aréchaga-Ocampo E, Villegas-Sepúlveda N, González-De la Rosa CH
BioMed research international 2015;2015:404368
BioMed research international 2015;2015:404368
A rapid and convenient method for fluorescence analysis of in vitro cultivated metacestode vesicles from Echinococcus multilocularis.
Cheng Z, Liu F, Zhu S, Tian H, Wang L, Wang Y
PloS one 2015;10(2):e0118215
PloS one 2015;10(2):e0118215
A new water soluble MAPK activator exerts antitumor activity in melanoma cells resistant to the BRAF inhibitor vemurafenib.
Graziani G, Artuso S, De Luca A, Muzi A, Rotili D, Scimeca M, Atzori MG, Ceci C, Mai A, Leonetti C, Levati L, Bonanno E, Tentori L, Caccuri AM
Biochemical pharmacology 2015 May 1;95(1):16-27
Biochemical pharmacology 2015 May 1;95(1):16-27
ATG12-ATG3 interacts with Alix to promote basal autophagic flux and late endosome function.
Murrow L, Malhotra R, Debnath J
Nature cell biology 2015 Mar;17(3):300-10
Nature cell biology 2015 Mar;17(3):300-10
Candidate tumor suppressor B-cell translocation gene 3 impedes neoplastic progression by suppression of AKT.
Cheng YC, Chen PH, Chiang HY, Suen CS, Hwang MJ, Lin TY, Yang HC, Lin WC, Lai PL, Shieh SY
Cell death & disease 2015 Jan 8;6(1):e1584
Cell death & disease 2015 Jan 8;6(1):e1584
Possible Mechanisms of Di(2-ethylhexyl) Phthalate-Induced MMP-2 and MMP-9 Expression in A7r5 Rat Vascular Smooth Muscle Cells.
Shih MF, Pan KH, Cherng JY
International journal of molecular sciences 2015 Dec 4;16(12):28800-11
International journal of molecular sciences 2015 Dec 4;16(12):28800-11
The MEK1/2-ERK1/2 pathway is activated in chronic rhinosinusitis with nasal polyps.
Linke R, Pries R, Könnecke M, Bruchhage KL, Böscke R, Gebhard M, Wollenberg B
Archivum immunologiae et therapiae experimentalis 2014 Jun;62(3):217-29
Archivum immunologiae et therapiae experimentalis 2014 Jun;62(3):217-29
Increased ERK and JNK activation and decreased ERK/JNK ratio are associated with long-term organ damage in patients with systemic lupus erythematosus.
Bloch O, Amit-Vazina M, Yona E, Molad Y, Rapoport MJ
Rheumatology (Oxford, England) 2014 Jun;53(6):1034-42
Rheumatology (Oxford, England) 2014 Jun;53(6):1034-42
Silver and fullerene nanoparticles' effect on interleukin-2-dependent proliferation of CD4 (+) T cells.
Côté-Maurais G, Bernier J
Toxicology in vitro : an international journal published in association with BIBRA 2014 Dec;28(8):1474-81
Toxicology in vitro : an international journal published in association with BIBRA 2014 Dec;28(8):1474-81
The C-terminus of human nucleotide receptor P2X7 is critical for receptor oligomerization and N-linked glycosylation.
Wickert LE, Blanchette JB, Waldschmidt NV, Bertics PJ, Denu JM, Denlinger LC, Lenertz LY
PloS one 2013;8(5):e63789
PloS one 2013;8(5):e63789
Molecular pathway profiling of T lymphocyte signal transduction pathways; Th1 and Th2 genomic fingerprints are defined by TCR and CD28-mediated signaling.
Smeets RL, Fleuren WW, He X, Vink PM, Wijnands F, Gorecka M, Klop H, Bauerschmidt S, Garritsen A, Koenen HJ, Joosten I, Boots AM, Alkema W
BMC immunology 2012 Mar 14;13:12
BMC immunology 2012 Mar 14;13:12
TFG-1 function in protein secretion and oncogenesis.
Witte K, Schuh AL, Hegermann J, Sarkeshik A, Mayers JR, Schwarze K, Yates JR 3rd, Eimer S, Audhya A
Nature cell biology 2011 May;13(5):550-8
Nature cell biology 2011 May;13(5):550-8
ErbB2 stabilizes epidermal growth factor receptor (EGFR) expression via Erk and Sprouty2 in extracellular matrix-detached cells.
Grassian AR, Schafer ZT, Brugge JS
The Journal of biological chemistry 2011 Jan 7;286(1):79-90
The Journal of biological chemistry 2011 Jan 7;286(1):79-90
Extranuclear estrogen receptors mediate the neuroprotective effects of estrogen in the rat hippocampus.
Yang LC, Zhang QG, Zhou CF, Yang F, Zhang YD, Wang RM, Brann DW
PloS one 2010 May 7;5(5):e9851
PloS one 2010 May 7;5(5):e9851
3-phosphoinositide-dependent protein kinase-1 regulates proliferation and survival of cancer cells with an activated mitogen-activated protein kinase pathway.
Lu Z, Cox-Hipkin MA, Windsor WT, Boyapati A
Molecular cancer research : MCR 2010 Mar;8(3):421-32
Molecular cancer research : MCR 2010 Mar;8(3):421-32
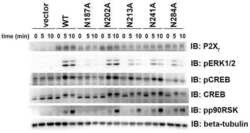
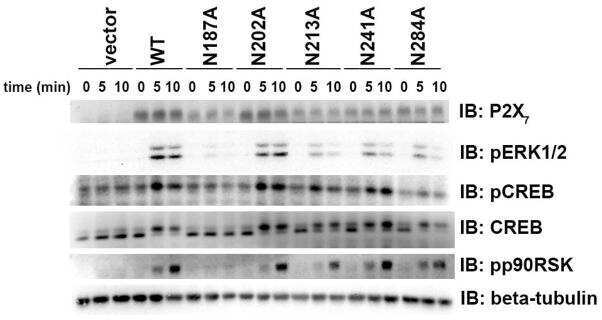
Mutation of putative N-linked glycosylation sites on the human nucleotide receptor P2X7 reveals a key residue important for receptor function.
Lenertz LY, Wang Z, Guadarrama A, Hill LM, Gavala ML, Bertics PJ
Biochemistry 2010 Jun 8;49(22):4611-9
Biochemistry 2010 Jun 8;49(22):4611-9
Induction of invasion in an organotypic oral cancer model by CoCl2, a hypoxia mimetic.
Brusevold IJ, Husvik C, Schreurs O, Schenck K, Bryne M, Søland TM
European journal of oral sciences 2010 Apr;118(2):168-76
European journal of oral sciences 2010 Apr;118(2):168-76
The absence of caveolin-1 increases proliferation and anchorage- independent growth by a Rac-dependent, Erk-independent mechanism.
Cerezo A, Guadamillas MC, Goetz JG, Sánchez-Perales S, Klein E, Assoian RK, del Pozo MA
Molecular and cellular biology 2009 Sep;29(18):5046-59
Molecular and cellular biology 2009 Sep;29(18):5046-59
Antioxidant and oncogene rescue of metabolic defects caused by loss of matrix attachment.
Schafer ZT, Grassian AR, Song L, Jiang Z, Gerhart-Hines Z, Irie HY, Gao S, Puigserver P, Brugge JS
Nature 2009 Sep 3;461(7260):109-13
Nature 2009 Sep 3;461(7260):109-13
A study of phosphorylated ERK1/2 and COX-2 in early stage (T1-T2) oral squamous cell carcinomas.
Søland TM, Husvik C, Koppang HS, Boysen M, Sandvik L, Clausen OP, Christoffersen T, Bryne M
Journal of oral pathology & medicine : official publication of the International Association of Oral Pathologists and the American Academy of Oral Pathology 2008 Oct;37(9):535-42
Journal of oral pathology & medicine : official publication of the International Association of Oral Pathologists and the American Academy of Oral Pathology 2008 Oct;37(9):535-42
Dissection of the osteogenic effects of laminin-332 utilizing specific LG domains: LG3 induces osteogenic differentiation, but not mineralization.
Klees RF, Salasznyk RM, Ward DF, Crone DE, Williams WA, Harris MP, Boskey A, Quaranta V, Plopper GE
Experimental cell research 2008 Feb 15;314(4):763-73
Experimental cell research 2008 Feb 15;314(4):763-73
Quantitative measurement of epidermal growth factor receptor-mitogen-activated protein kinase signal transduction using a nine-plex, peptide-based immunoassay.
Rauh-Adelmann C, Moskow JM, Graham JR, Yen LG, Boucher JI, Murphy CE, Nadler TK, Gordon NF, Radding JA
Analytical biochemistry 2008 Apr 15;375(2):255-64
Analytical biochemistry 2008 Apr 15;375(2):255-64
Molecular mechanisms involved in interleukin-4-induced human neutrophils: expression and regulation of suppressor of cytokine signaling.
Ratthé C, Pelletier M, Chiasson S, Girard D
Journal of leukocyte biology 2007 May;81(5):1287-96
Journal of leukocyte biology 2007 May;81(5):1287-96
Activation of FAK is necessary for the osteogenic differentiation of human mesenchymal stem cells on laminin-5.
Salasznyk RM, Klees RF, Boskey A, Plopper GE
Journal of cellular biochemistry 2007 Feb 1;100(2):499-514
Journal of cellular biochemistry 2007 Feb 1;100(2):499-514
The mitogen-activated protein kinases (MAPK) p38 and JNK are markers of tumor progression in breast carcinoma.
Davidson B, Konstantinovsky S, Kleinberg L, Nguyen MT, Bassarova A, Kvalheim G, Nesland JM, Reich R
Gynecologic oncology 2006 Sep;102(3):453-61
Gynecologic oncology 2006 Sep;102(3):453-61
Epidermal growth factor receptor induced Erk phosphorylation in the suprachiasmatic nucleus.
Hao H, Schwaber J
Brain research 2006 May 9;1088(1):45-8
Brain research 2006 May 9;1088(1):45-8
Phospho-caveolin-1 mediates integrin-regulated membrane domain internalization.
del Pozo MA, Balasubramanian N, Alderson NB, Kiosses WB, Grande-García A, Anderson RG, Schwartz MA
Nature cell biology 2005 Sep;7(9):901-8
Nature cell biology 2005 Sep;7(9):901-8
Heat shock protein 60 inhibits Th1-mediated hepatitis model via innate regulation of Th1/Th2 transcription factors and cytokines.
Zanin-Zhorov A, Bruck R, Tal G, Oren S, Aeed H, Hershkoviz R, Cohen IR, Lider O
Journal of immunology (Baltimore, Md. : 1950) 2005 Mar 15;174(6):3227-36
Journal of immunology (Baltimore, Md. : 1950) 2005 Mar 15;174(6):3227-36
Mitogen-activated protein kinase expression and activation does not differentiate benign from malignant mesothelial cells.
Vintman L, Nielsen S, Berner A, Reich R, Davidson B
Cancer 2005 Jun 1;103(11):2427-33
Cancer 2005 Jun 1;103(11):2427-33
CHIR-258: a potent inhibitor of FLT3 kinase in experimental tumor xenograft models of human acute myelogenous leukemia.
Lopes de Menezes DE, Peng J, Garrett EN, Louie SG, Lee SH, Wiesmann M, Tang Y, Shephard L, Goldbeck C, Oei Y, Ye H, Aukerman SL, Heise C
Clinical cancer research : an official journal of the American Association for Cancer Research 2005 Jul 15;11(14):5281-91
Clinical cancer research : an official journal of the American Association for Cancer Research 2005 Jul 15;11(14):5281-91
Chemokine receptor CCR2 expression by systemic sclerosis fibroblasts: evidence for autocrine regulation of myofibroblast differentiation.
Carulli MT, Ong VH, Ponticos M, Shiwen X, Abraham DJ, Black CM, Denton CP
Arthritis and rheumatism 2005 Dec;52(12):3772-82
Arthritis and rheumatism 2005 Dec;52(12):3772-82
No comments: Submit comment
Supportive validation
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
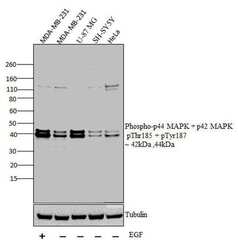
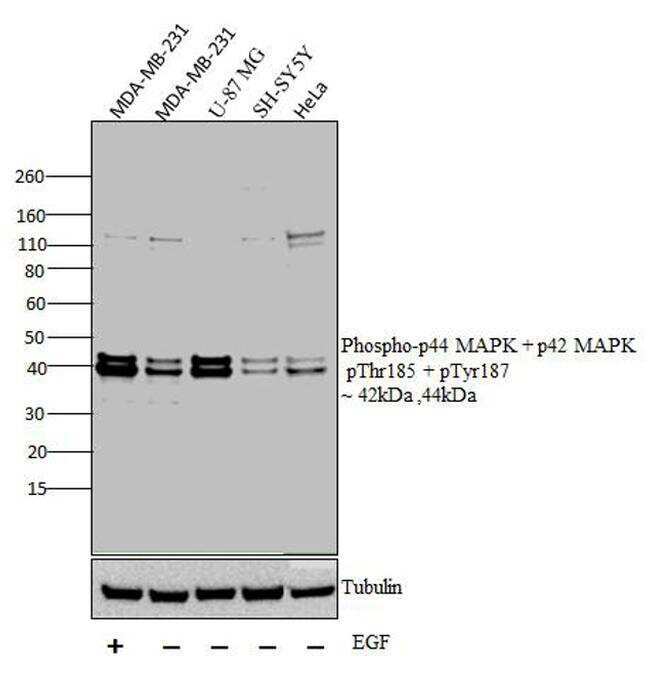
- Western blot analysis was performed on whole cell extracts (30 µg lysate) of MDA-MB-231 with treatment of EGF(100 ng/mL for 15 mins) (Lane 1), MDA-MB-231 (Lane 2), U-87 MG (Lane 3),, SH-SY5Y (Lane 4) and HeLa (Lane 5). The blots were probed with Anti-Phospho-p44 MAPK + p42 MAPK pThr185 + pTyr187 Rabbit polyclonal Antibody (Product# 44680G, 1 in 1000 dilution) and detected by chemiluminescence Goat anti-Rabbit IgG (H+L) Secondary Antibody, HRP conjugate (Product # G-21234, 1:5000 dilution). Bands of 42 kDa and 44 kDa corresponding to Phospho-p44 MAPK + p42 MAPK pThr185 + pTyr187 was observed across cell lines tested. Known quantity of protein samples were electrophoresed using Novex® NuPAGE® 12 % Bis-Tris gel (Product # NP0341BOX), XCell SureLock™ Electrophoresis System (Product # EI0002) and Novex® Sharp Pre-Stained Protein Standard (Product # LC5800). Resolved proteins were then transferred onto a nitrocellulose membrane with iBlot® 2 Dry Blotting System (Product # IB21001). The membrane was probed with the relevant primary and secondary Antibody following blocking with 5 % skimmed milk. Chemiluminescent detection was performed using Pierce™ ECL Western Blotting Substrate (Product # 32106).
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
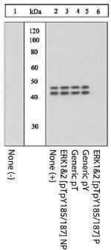
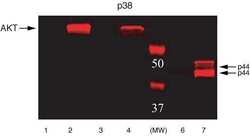
- Extracts of PC12 cells unstimulated (1) or stimulated with 0.5 M sorbitol for 5 minutes (2-6) were resolved by SDS-PAGE on a 10% Tris-glycine gel and transferred to PVDF. The membrane was blocked with a 5% BSA-TBST buffer overnight at 4°C, and then incubated with the ERK1&2 (pTpY 185/187) antibody for two hours at room temperature in a 3% BSA-TBST buffer, following its prior incubation with: no peptide (1, 2), the non-phosphopeptide corresponding to the phosphopeptide immunogen (3), a generic phosphothreonine-containing peptide (4), a generic phosphotyrosine-containing peptide (5), or the phosphopeptide immunogen (6). After washing, the membrane was incubated with goat F (ab')2 anti-rabbit IgG HRP conjugate (Product # ALI4404) and signals were detected using the Pierce SuperSignal™ method.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
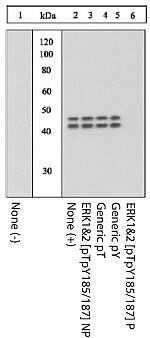
- ERK1&2 (pTpY185/187) phosphospecific antibody. NIH3T3 or NIH3T3-L1 cells were treated with either PDGF or Insulin. Western Blots were performed using either Akt (pS473) (Product # 44-622G) or Erk (pTpY185/187) (Product # 44-680G) primary antibodies. Anti-rabbit secondary antibody conjugated to Alexa fluor 680 was used for detection. Data was analyzed on the LI-COR Odyssey® Infrared Imaging System. Akt (pS473): Lane 1 - NIH3T3 control, 20 µg; Lane 2 - NIH3T3 + PDGF, 20 µg; Lane 3 - NIH3T3-L1 control, 20 µg; Lane 4 - NIH3T3-L1 + Insulin, 20 µg. Erk (pTpY185/187): Lane 6 - NIH3T3 control, 5 µg; Lane 7 - NIH3T3 + PDGF, 5 µg.

Supportive validation
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
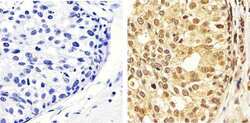
- Immunohistochemistry analysis of ERK1/2 (pTpY185/187) showing staining in the cytoplasm and nucleus of paraffin-embedded human breast carcinoma tissue (right) compared to a negative control without primary antibody (left). To expose target proteins, antigen retrieval was performed using 10mM sodium citrate (pH 6.0), microwaved for 8-15 min. Following antigen retrieval, tissues were blocked in 3% H2O2-methanol for 15 min at room temperature, washed with ddH2O and PBS, and then probed with a ERK1/2 (pTpY185/187) polyclonal antibody (Product # 44-680G) diluted in 3% BSA-PBS at a dilution of 1:50 overnight at 4ºC in a humidified chamber. Tissues were washed extensively in PBST and detection was performed using an HRP-conjugated secondary antibody followed by colorimetric detection using a DAB kit. Tissues were counterstained with hematoxylin and dehydrated with ethanol and xylene to prep for mounting.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
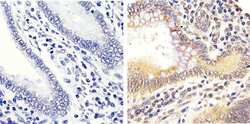
- Experimental details
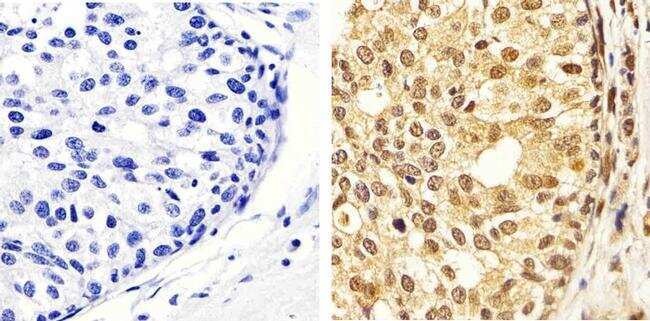
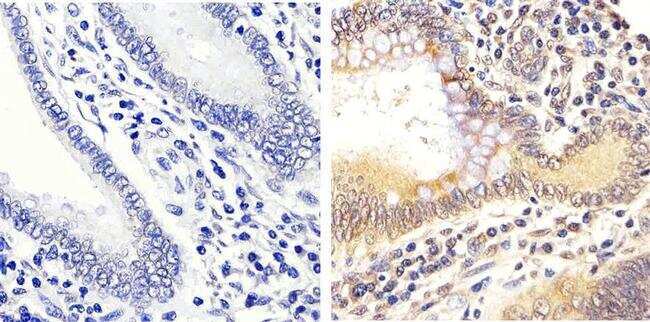
- Immunohistochemistry analysis of ERK1/2 (pTpY185/187) showing staining in the cytoplasm and nucleus of paraffin-embedded human colon carcinoma tissue (right) compared to a negative control without primary antibody (left). To expose target proteins, antigen retrieval was performed using 10mM sodium citrate (pH 6.0), microwaved for 8-15 min. Following antigen retrieval, tissues were blocked in 3% H2O2-methanol for 15 min at room temperature, washed with ddH2O and PBS, and then probed with a ERK1/2 (pTpY185/187) polyclonal antibody (Product # 44-680G) diluted in 3% BSA-PBS at a dilution of 1:20 overnight at 4ºC in a humidified chamber. Tissues were washed extensively in PBST and detection was performed using an HRP-conjugated secondary antibody followed by colorimetric detection using a DAB kit. Tissues were counterstained with hematoxylin and dehydrated with ethanol and xylene to prep for mounting.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
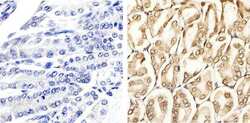
- Experimental details
- Immunohistochemistry analysis of ERK1/2 (pTpY185/187) showing staining in the cytoplasm and nucleus of paraffin-embedded mouse stomach tissue (right) compared to a negative control without primary antibody (left). To expose target proteins, antigen retrieval was performed using 10mM sodium citrate (pH 6.0), microwaved for 8-15 min. Following antigen retrieval, tissues were blocked in 3% H2O2-methanol for 15 min at room temperature, washed with ddH2O and PBS, and then probed with a ERK1/2 (pTpY185/187) polyclonal antibody (Product # 44-680G) diluted in 3% BSA-PBS at a dilution of 1:20 overnight at 4ºC in a humidified chamber. Tissues were washed extensively in PBST and detection was performed using an HRP-conjugated secondary antibody followed by colorimetric detection using a DAB kit. Tissues were counterstained with hematoxylin and dehydrated with ethanol and xylene to prep for mounting.
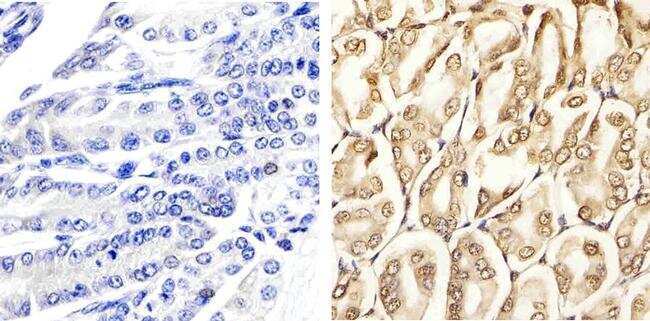
Supportive validation
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
- NULL

- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL

- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
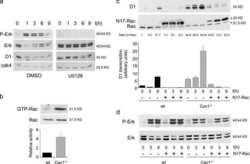
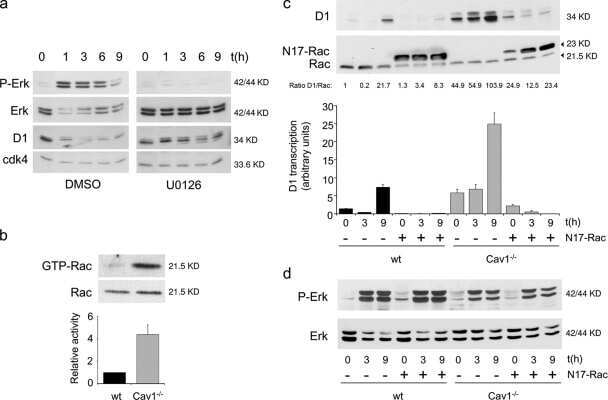
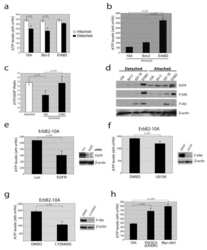
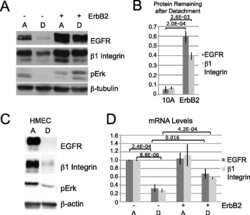
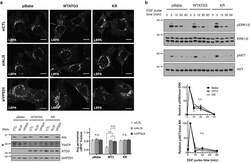
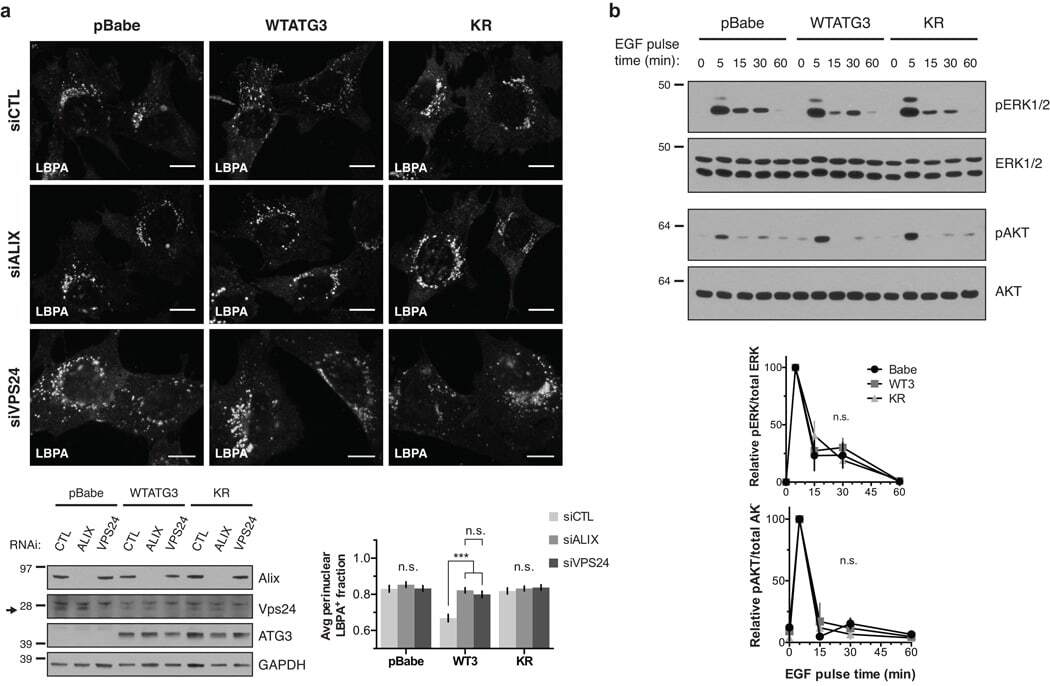
- Figure 5 Loss of ATG12-ATG3 conjugation phenocopies loss of Alix (a) Indicated cell types were transfected with siRNA against ALIX (siALIX), VPS24 (siVPS24), or a non-targeting control (siCTL) and immunostained with anti-LBPA to mark late endosomes. Scale bar, 10 um. Right: Quantification of the perinuclear LBPA + fraction as described in Fig. 2d (mean +- SEM; n = 60 cells pooled from three independent experiments). Statistical significance calculated using ANOVA, followed by Tukey's HSD test (***P < 0.001). Matched lysates were immunoblotted for anti-Alix and anti-Vps24. (b) Indicated cell types were serum-starved overnight at 37 degC followed by treatment with 100 ng/mL EGF for the indicated times. Lysates were immunoblotted for anti-phosphorylated-ERK1/2 (pERK1/2), anti-ERK1/2, anti-phosphorylated-AKT (pAKT), and anti-AKT. Right: Quantification of phosphorylated ERK and AKT levels (mean SEM; n = 3 independent experiments). Statistical significance calculated using ANOVA, followed by Tukey's HSD test. Uncropped images of blots are shown in Supplementary Figure 6 .
- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
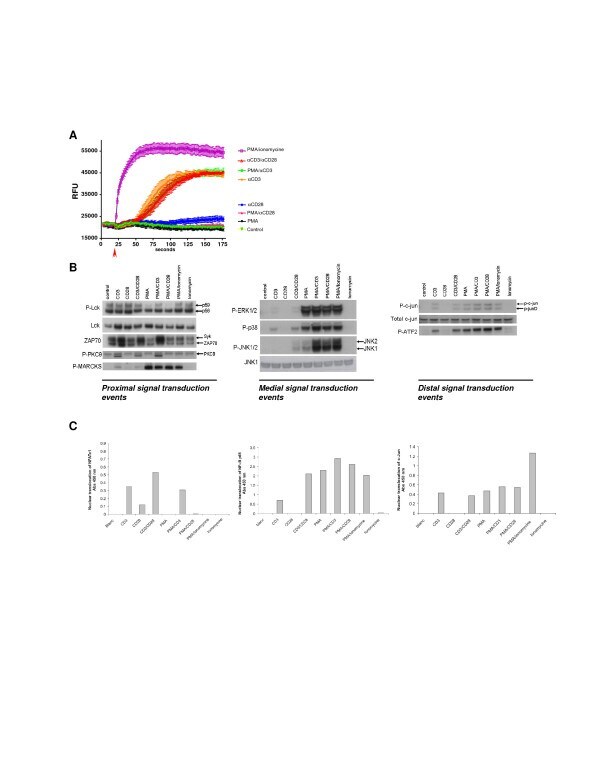
- Figure 1 Signal transduction in Jurkat T cells . Jurkat T cells were stimulated with different combinations of stimuli in order to elucidate the different signal transduction pathways. A ; Jurkat T cells were stimulated as indicated and intracellular Ca 2+ release was monitored over/in time. B ; Intracellular signal transduction routes were charted via phosphoanalysis using western blot. Jurkat T cells were stimulated for 15 min using different stimulations. Proximal (Lck, ZAP70, PKCtheta and the PKC substrate MARCKs), medial (MAPK phosphorylation) and distal (c-Jun and ATF2) signaling was monitored based on the phosphorylation status of the described proteins. C ; Nuclear translocation of the transcription factors NFAT, NFkB and c-Jun was evaluated 15 minutes after stimulation.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
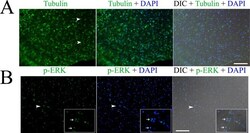
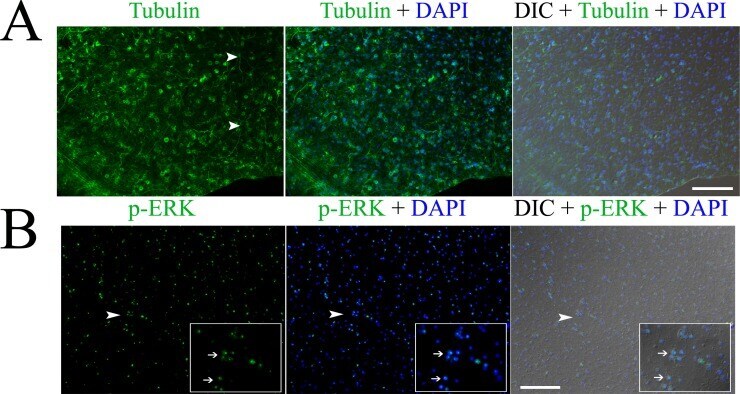
- Fig 1 Immunofluorescence detection of beta tubulin (A) and phosphorylated ERK (p-ERK) (B) in the germinal layer of the whole-mount preparations of in vitro cultured E. multilocularis vesicles. Arrow heads in (A) indicate the neuron cells with long neurites. Arrow heads in (B) indicate the areas which are magnified in the inserts. Note the higher expression of p-ERK in the nucleolus of some cells indicated by the arrows in the inserts. Merges of the differential interference contrast (DIC) bright field microscopy with the fluorescence channels are shown. Bar = 125 mum.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
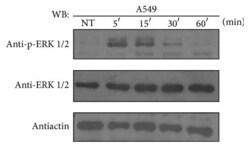
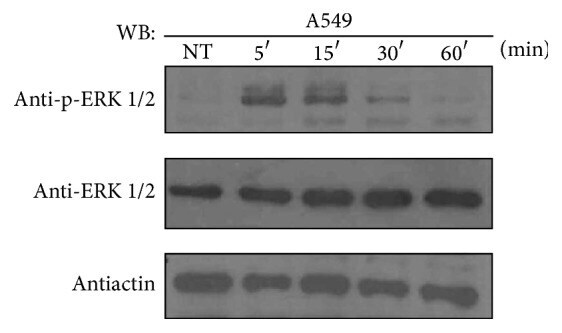
- Figure 1 ATRA activates the ERK pathway through nongenomic mechanisms in A549 cells. A549 cells were serum-starved for 18 h, treated or nontreated (NT) with 5 mu M of ATRA for the times indicated. The phosphorylated form of ERK and total proteins were detected by western blot using specific antibodies.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
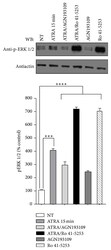
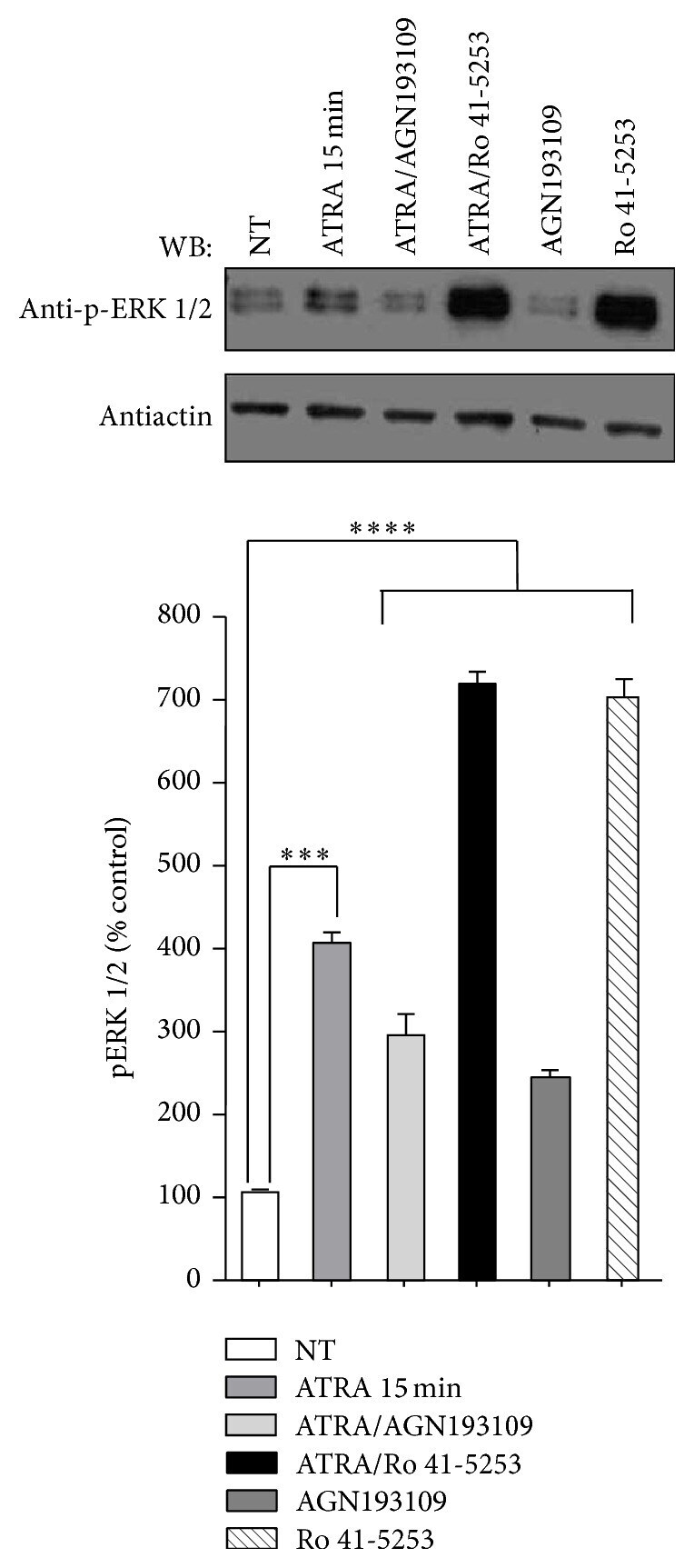
- Figure 2 Effect of AGN193109 and Ro 41-5253 inhibitors on ATRA-induced ERK activation. A549 cells were serum-starved for 18 h, treated or nontreated (NT) with 5 mu M of ATRA for 15 minutes. Cells were preincubated for 1 h with 10 mu M of AGN193109 or 20 mu M of Ro 41-5253 alone or in combination with ATRA. The phosphorylated form of ERK was detected by western blot using specific antibodies. beta -Actin was used as the loading control. The graph represents the densitometric values of ERK phosphorylation in three independent experiments (means +- SEM, *** P < 0.0001; **** P < 0.00001 compared with NT cells, analysis of variance and Newman-Keuls test).
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
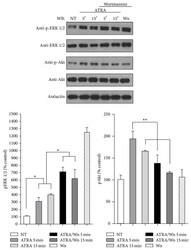
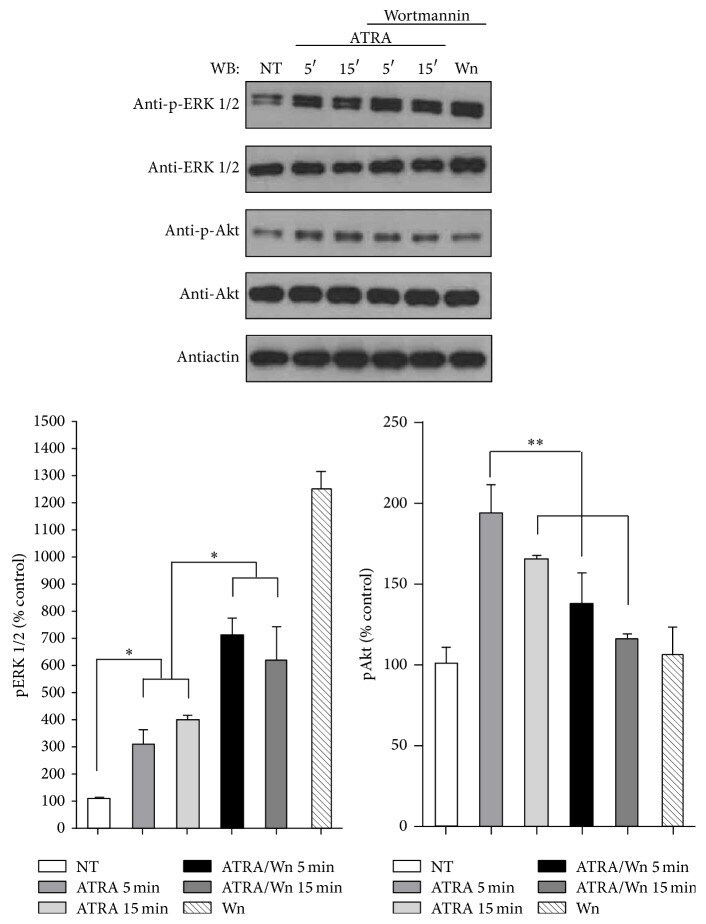
- Figure 3 Effect of PI3K inhibitor wortmannin on ATRA-induced ERK phosphorylation. A549 cells were serum-starved for 18 h and treated or nontreated (NT) with 5 mu M of ATRA at different times. Cells were preincubated for 1 h with 10 mu M of wortmannin (Wm) alone or in combination with ATRA. The phosphorylated form of ERK and Akt and total proteins were detected by western blot using specific antibodies. The graph represents the densitometric analysis of ERK and Akt phosphorylation in three independent experiments (means +- SEM, * P < 0.05; ** P < 0.001; analysis of variance and Newman-Keuls test).
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
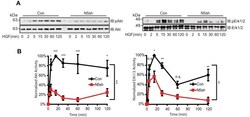
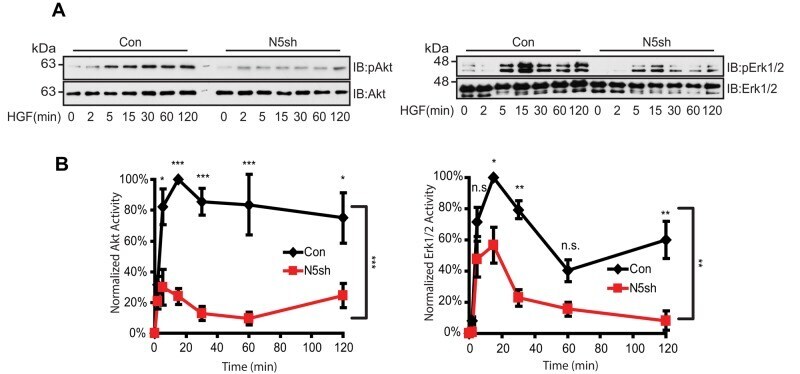
- FIGURE 5: HGF-induced Akt/Erk phosphorylation is attenuated by NHE5 knockdown. Cells serum-starved overnight were stimulated with 50 ng/ml of HGF. The amount of phosphorylated and total Akt and Erk1/2 was detected by immunoblotting. (A) Representative immunoblots are shown. (B) The relative level of phospho-Akt and phospho-Erk are expressed as mean +- SEM of four experiments. p Values for pairwise comparisons at each time point were obtained following repeated-measures ANOVA with time as the within-subject factor. *, p < 0.05, **, p < 0.01, ***, p < 0.001, n.s., not significant.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
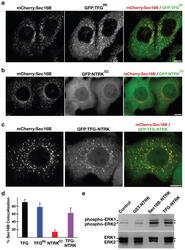
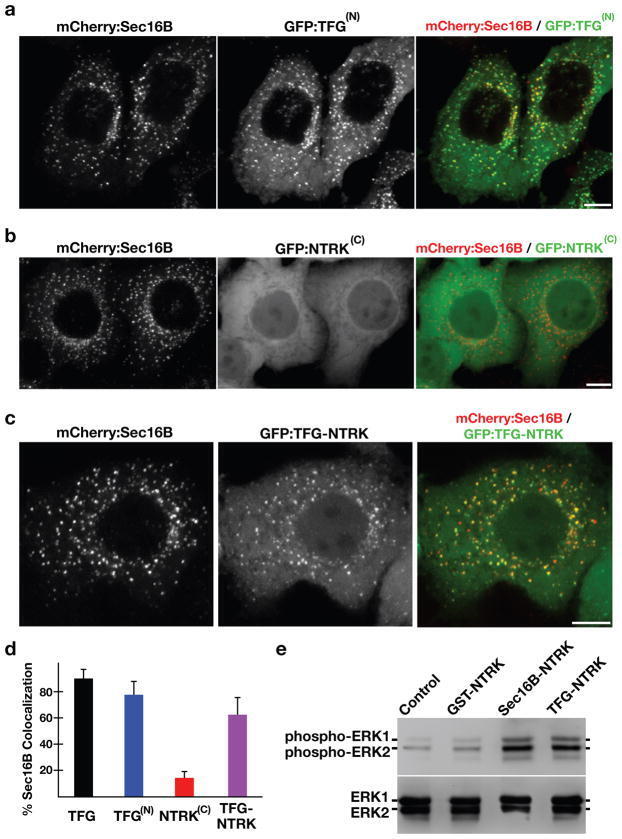
- Figure 7 Targeting of the NTRK1 kinase domain to ER exit sites is sufficient to activate NTRK1-mediated downstream signaling (a-c) Swept field confocal optics were used to image HeLa cells that had been transiently transfected with mCherry:Sec16B and GFP fusions to either the amino terminus of TFG referred to as GFP:TFG (N) (n=18), the transmembrane and kinase domains of NTRK1 referred to as GFP:NTRK (C) (n=15), or a TFG (N) -NTRK1 (C) fusion (n=28), which is equivalent to the oncogene characterized previously ( 21 ). Representative color overlays of mCherry:Sec16B (red) and GFP fusions (green) are shown. Scale bar, 10 mum. (d) Bar graph showing the percent co-localization between the GFP fusions described above and mCherry:Sec16B (error bars represent means +/- SEM for each condition; n=15 different cells for each condition and at least 800 unique ER exit sites were examined). (e) Extracts from hTERT-RPE1 cells stably transfected with GFP alone (Control) or various GFP fusions to the NTRK1 transmembrane and kinase domains (as indicated) were separated by SDS-PAGE and blotted using a phospho-specific ERK1-ERK2 antibody (top) and a pan-ERK1-ERK2 antibody (bottom).
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
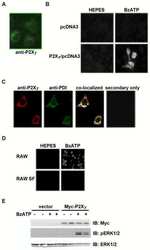
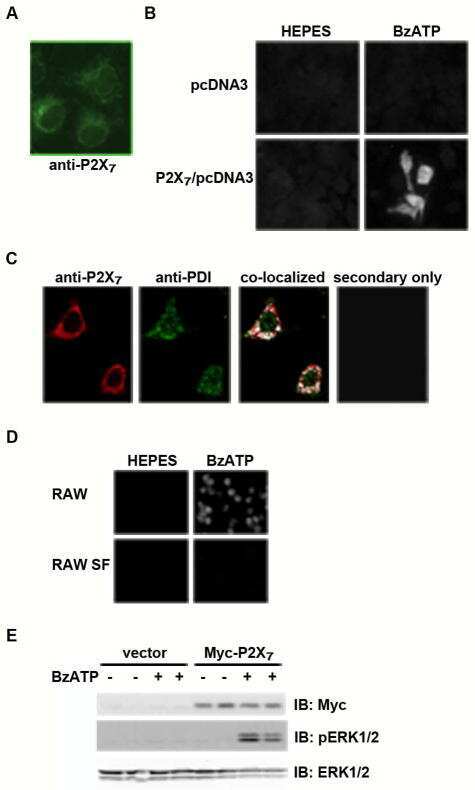
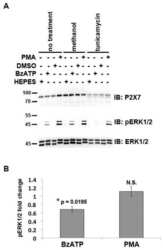
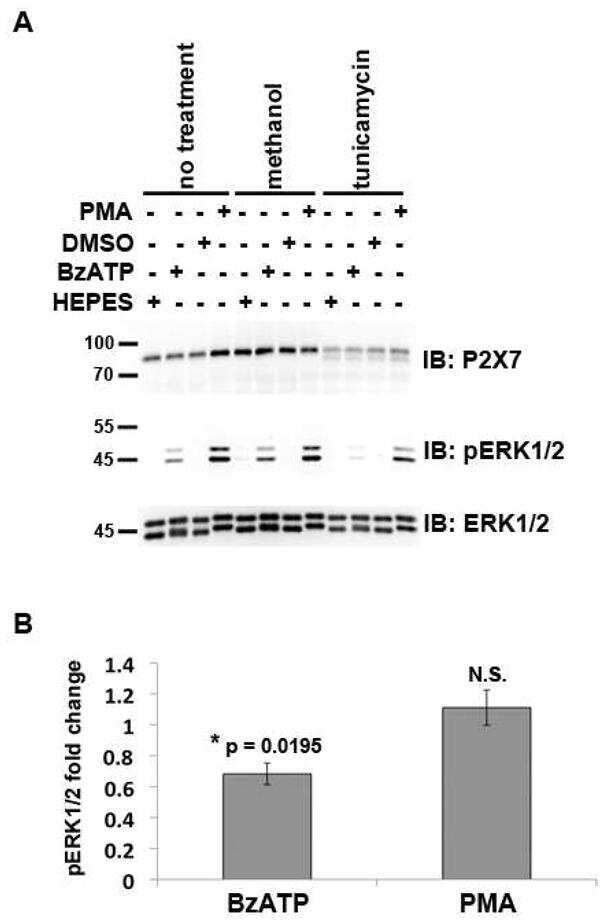
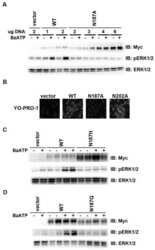
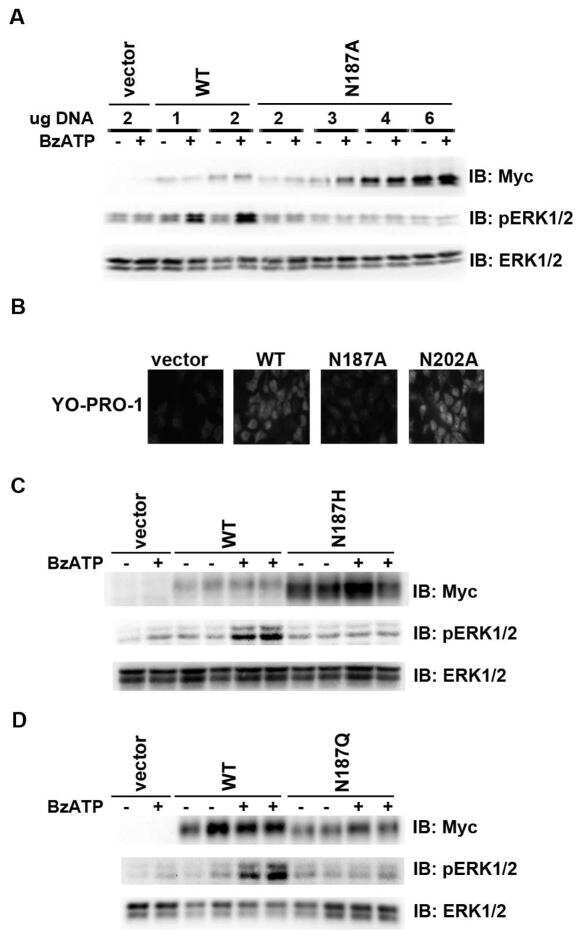
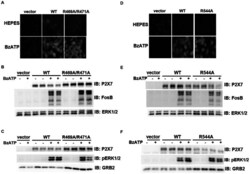
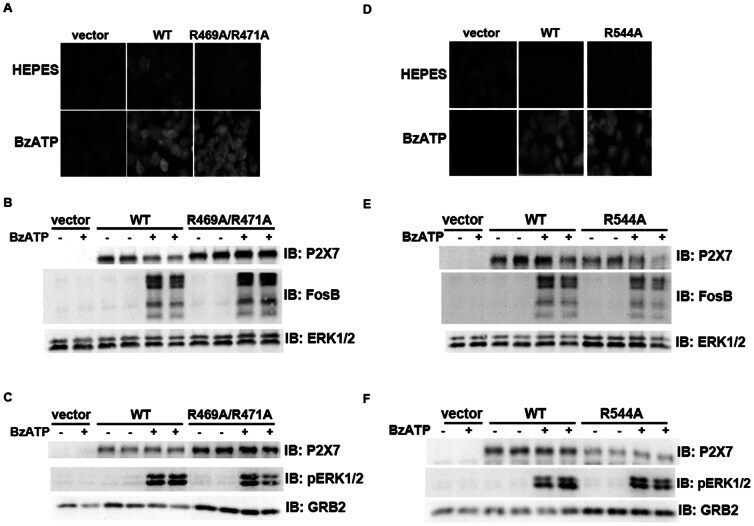
- Figure 7 Mutations within the other Arg-X-Arg sequences do not result in altered P2X 7 activity. A and D, The P2X 7 R469A/R471A double mutant and the P2X 7 R554A single mutant do not display attenuated nucleotide-stimulated pore activity. These data are representative of at least three experiments. B and E, The P2X 7 R469A/R471A and R554A mutants do not display attenuated BzATP-stimulated FosB/DeltaFosB induction. The anti-FosB antibody recognizes both full-length FosB (indicated by the top three bands) and its truncated splice variant DeltaFosB (indicated by the bottom two bands). These data are representative of at least two experiments. C and F, The P2X 7 R469A/R471A and R554A mutants exhibit normal nucleotide-stimulated ERK1/2 activation. These data are representative of two experiments.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
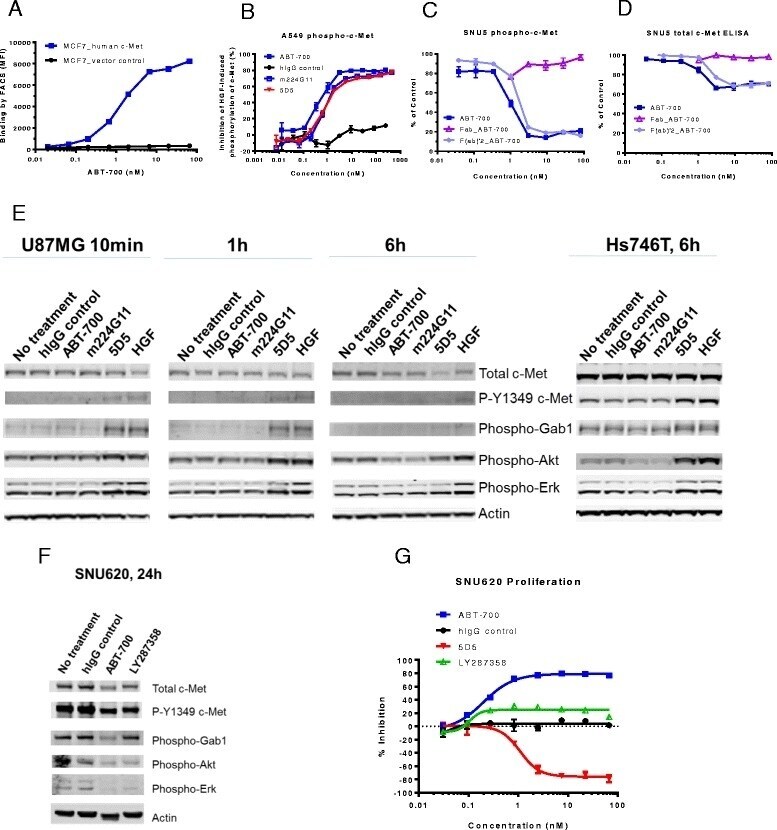
- Fig. 1 ABT-700 specifically binds cellular c-Met and antagonizes c-Met signaling in both HGF-dependent and -independent settings. a FACS analysis of ABT-700 binding to MCF7 transfectants. Stable human c-Met or vector control transfectants of human MCF7 breast cancer cells were incubated with increasing amounts of ABT-700 and bound ABT-700 was detected by FACS with secondary anti-human IgG conjugated with Alexa 488. b ELISA quantification of phospho-c-Met in A549 cells. A549 cells grown in a 96-well plate were pre-incubated for one hour with antibodies in a dose-range as shown, followed by stimulation with 1 nM HGF for 10 min. Total cell lysates were made and phospho-c-Met was detected by ELISA. c ELISA quantification of phospho-c-Met in SNU5 cells. SNU5 cells grown in a 96-well plate were incubated with antibodies in a dose-range as shown for 6 h. Total cell lysates were made and subjected to ELISA for phospho-c-Met. The value of cells in media alone was used as 100 % of control. d ELISA quantification of total c-Met in SNU5 cells. SNU5 cells grown in a 96-well plate were incubated with antibodies in a dose-range as shown for 6 h. Total cell lysates were made and c-Met level was determined by ELISA. The value of cells in media alone was used as 100 % of control. e Western blot analysis of U87MG cell lysates. U87MG cells grown in a 12-well plate were treated with antibodies as shown at 10 mug/mL for 10 min, 1 h or 6 h. Total cell lysates were analyzed for c-Met and other phosp
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
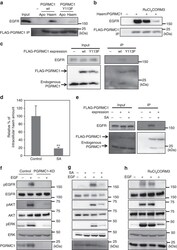
- Figure 4 Haem-dependent dimerization of PGRMC1 is necessary for tumour proliferation mediated by EGFR signalling. ( a ) FLAG-PGRMC1 wild-type (wt) and Y113F mutant proteins (a.a.44-195), in either apo- or haem-bound form, were incubated with purified EGFR and co-immunoprecipitated with anti-FLAG antibody-conjugated beads. Input and bound proteins were detected by Western blotting. ( b ) In vitro binding assay was performed as in ( a ) using haem-bound FLAG-PGRMC1 wt (a.a.44-195) and purified EGFR with or without treatment of RuCl 3 and CORM3. ( c ) FLAG-PGRMC1 wt or Y113F (full length) was over-expressed in HCT116 cells and immunoprecipitated with anti-FLAG antibody-conjugated beads. Co-immunoprecipitated proteins (FLAG-PGRMC1, endogenous PGRMC1 and EGFR) were detected with Western blotting by using anti-PGRMC1 or anti-EGFR antibody. ( d ) HCT116 cells were treated with or without 250 mumol l -1 of succinylacetone (SA) for 48 h. The intracellular haem was extracted and quantified by reverse-phase HPLC. The data represent mean+-s.d. of four separate experiments. ** P
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
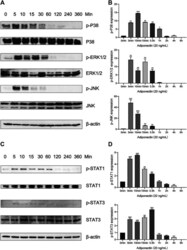
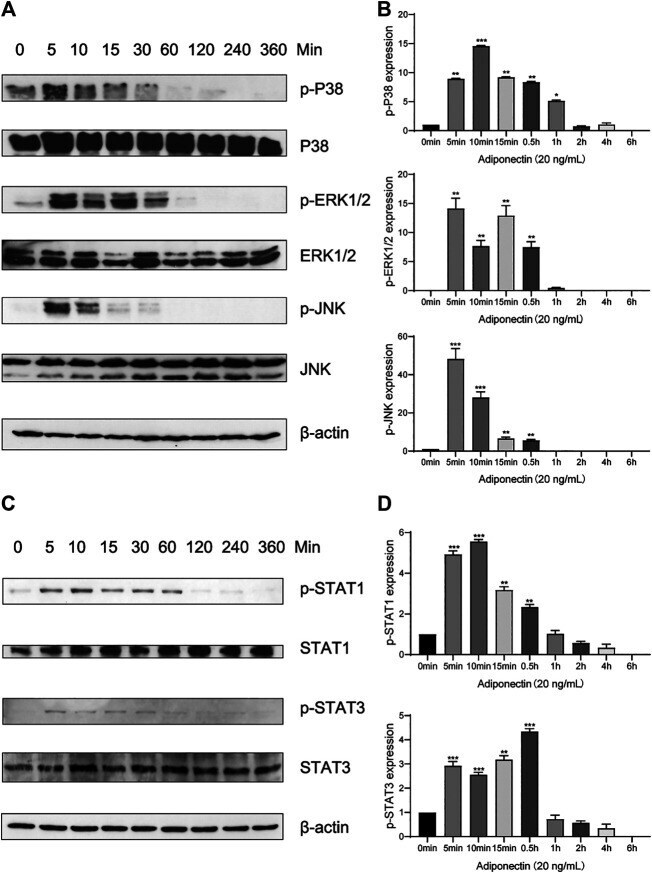
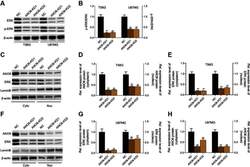
- FIGURE 4 Adiponectin promotes JNK, ERK1/2 and P38 phosphorylation on OCCM-30 cells. (A,B) The expression of JNK (46 and 54 kDa), ERK1/2 (42 and 44 kDa) and P38 (42 kDa) expression as well as their phosphorylated forms after Adiponectin (20 ng/mL) stimulation were analyzed by Western Blots. beta-actin served as a loading control. Graphics show the relative expression of p-JNK, p-ERK1/2 and p-P38 compared to cells at time point 0 min. (C,D) Adiponectin (20 ng/mL) promotes STAT1 (98 KDa) and STAT3 (85 KDa) phosphorylation during a period of 30 min. Graphics show the relative expression of p-STAT1 and p-STAT3 compared to cells at time point 0 min. Values are expressed as means +- SD: Ns (not significant), * p < 0.05; ** p < 0.01; *** p < 0.001.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
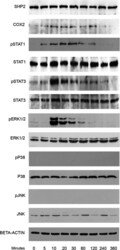
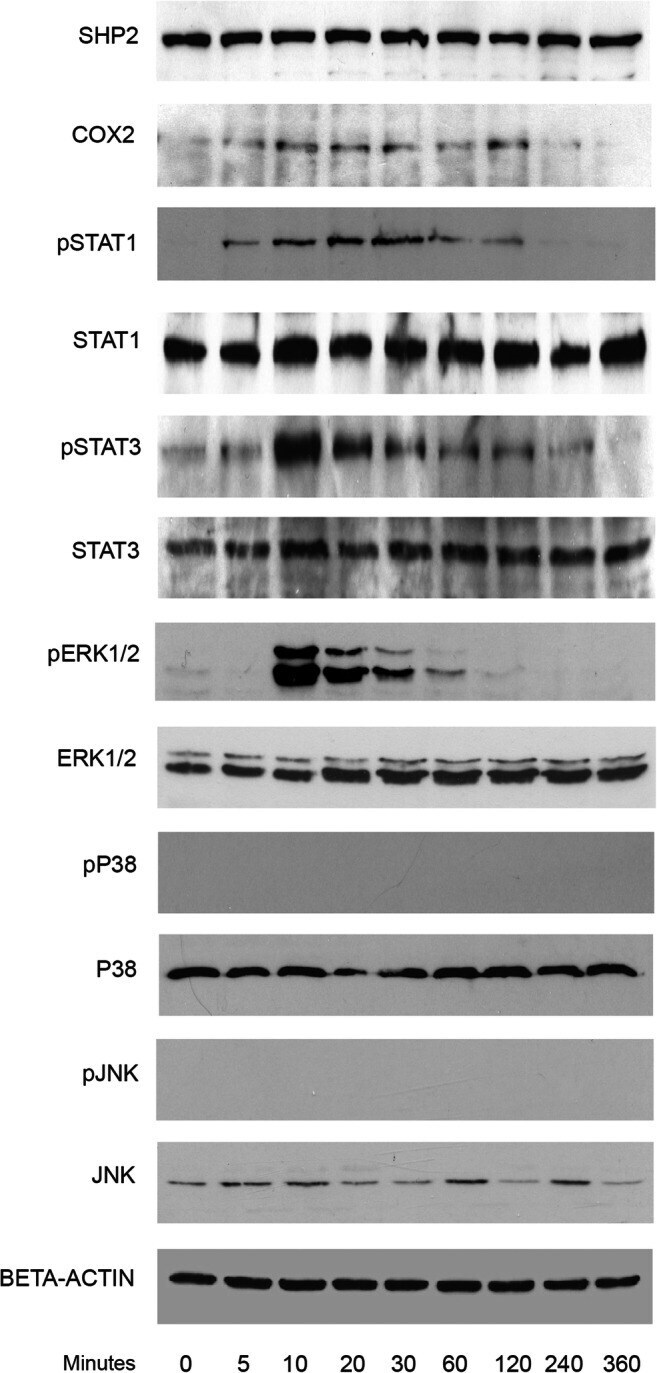
- Fig. 2 Kinetic analysis performed on OCCM-30 cells after stimulation with leptin. The expression of SHP2; COX2; p-STAT1; STAT1; p-STAT3; STAT3; p-ERK1/2; ERK1/2; p-P38; P38; p-JNK; and JNK were visualized by western blot: Leptin upregulates p-STAT1; p-STAT3; and p-ERK1/2 as well as COX2 on OCCM cells whereas no activation of P38 or JNK was observed. Each experiment was repeated three times
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
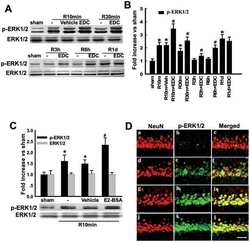
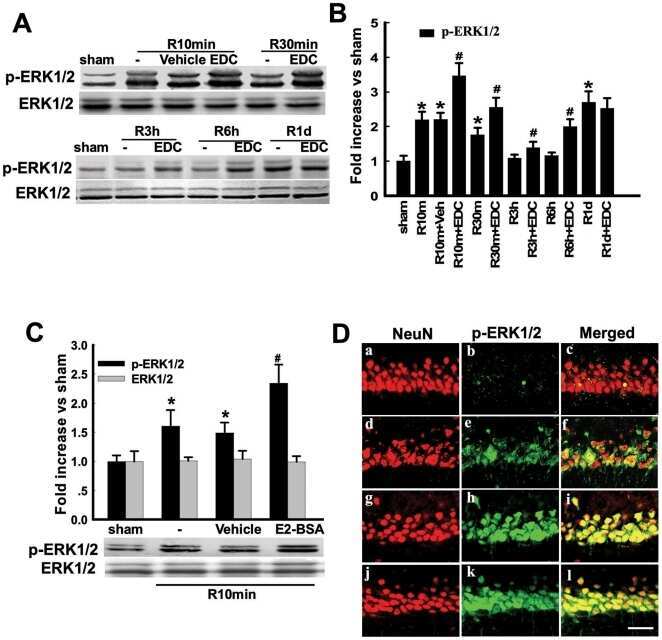
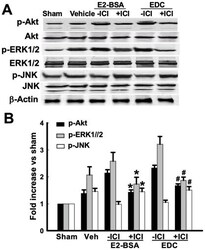
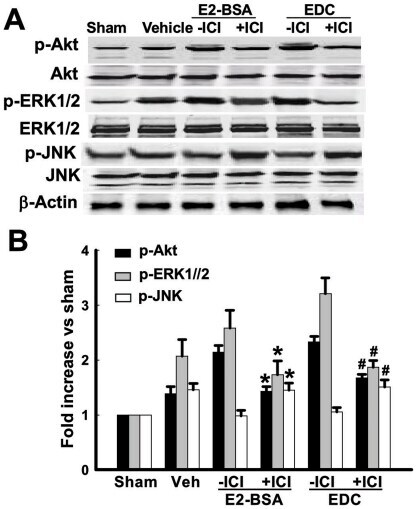
- Figure 6 Effects of ICI 182,780 on phosphorylation of Akt, ERK1/2 and JNK induced by E2-BSA or EDC at 10min of reperfusion in hippocampal CA1 region. ( A ) Western blot analysis showed phosphorylation levels and protein expressions of Akt, ERK1/2 and JNK. Samples were obtained from sham, vehicle, E2-BSA, E2-BSA+ICI, EDC and EDC+ICI treated animals. Bands corresponding to p-Akt, p-ERK1/2 and p-JNK were scanned and the intensities represented as fold increase vs. sham as shown in ( B ). F(5,18) = 106, p
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
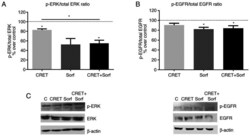
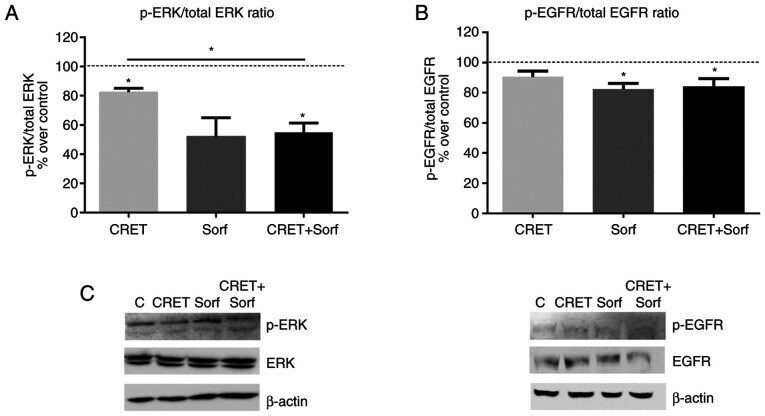
- Experimental details
- Figure 4. Effect of CRET and/or sorafenib on EGFR and ERK expression and activation. Western blotting results for p-ERK, ERK, p-EGFR and EGFR expression are presented. The samples were treated with CRET alone (4 h intermittent exposure), sorafenib alone (4 h) or in combination. Data are presented as the ratio of (A) p-ERK1/2 over total ERK1/2 protein and (B) p-EGFR over total EGFR protein. Data are presented as the mean +- SEM of >=3 experimental replicates. *P
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
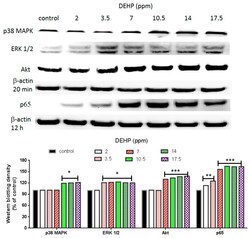
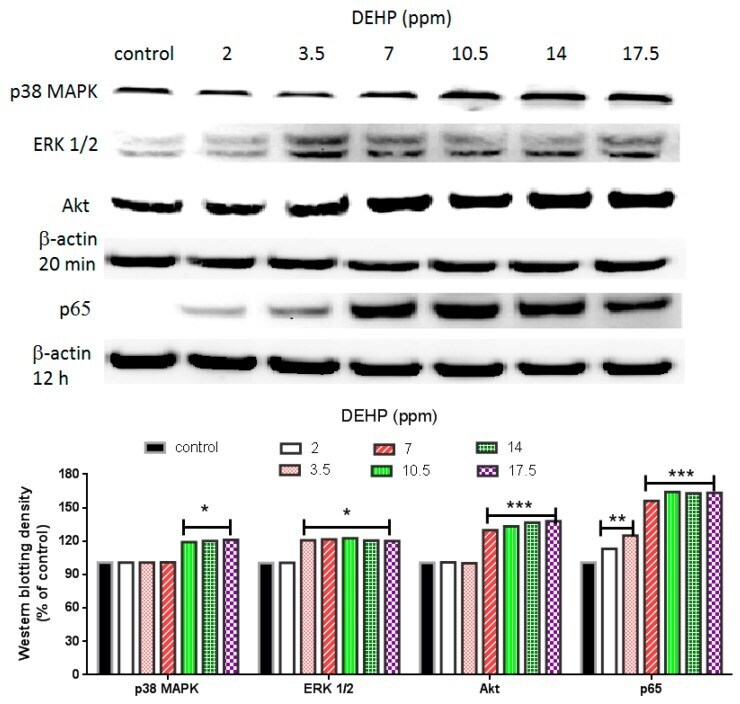
- Experimental details
- Figure 3 Effects of DEHP on phosphorylated p38 MAPK, phosphorylated ERK1/2, phosphorylated Akt, and NF-kappaB (p65) protein expression. VSMC ( n >= 3) were treated with DEHP (concentrations between 2 and 17.5 ppm) for 20 min (p38 MAPK, ERK1/2, and Akt) or 12 h (NF-kappaB) prior to protein extraction. Phosphorylated p38 MAPK, phosphorylated ERK1/2, phosphorylated Akt, and NF-kappaB (p65) were expressed by Western blotting. Statistics are shown for DEHP-treated cells * p < 0.05, ** p < 0.01, and *** p < 0.005, compared to the respective control groups.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
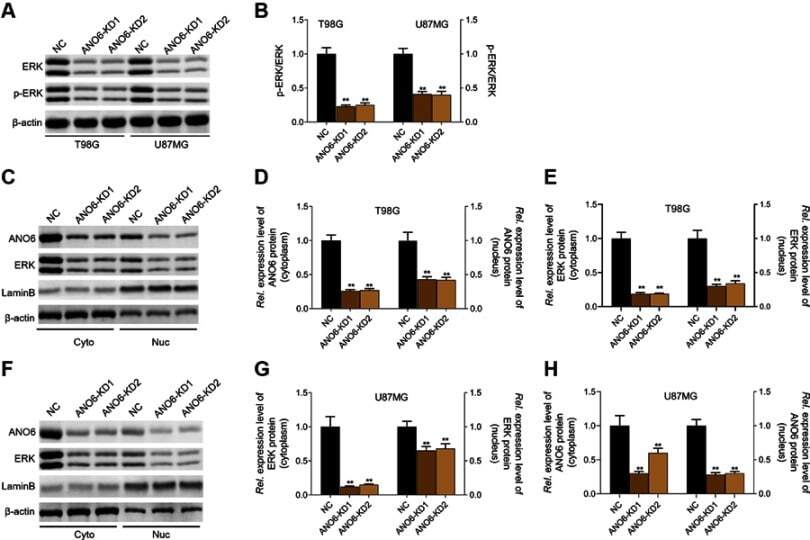
- Knockdown of ANO6 inhibited the activation of ERK signaling. After T98G and U87MG cells transfection with ANO6-KD 1 or ANO6-KD2, (A and B) the protein expression level of ERK and phospho-ERK was detected by Western blot assay; (C) the protein expression levels of ANO6 and ERK were detected by Western blot assay in cytoplasm and cell nucleus of T98G cells; (D and E) Histograms were used to represent statistical results of the protein expression levels of ANO6 and ERK in cytoplasm and cell nucleus of T98G cells; (F) the protein expression levels of ANO6 and ERK were detected by Western blot assay in cytoplasm and cell nucleus of U87MG cells. (G and H) Histograms were used to represent statistical results of the protein expression levels of ANO6 and ERK in cytoplasm and cell nucleus of U87MG cells; β-actin was used as a load control for cytoplasm. LaminB was used as a load control for cell nucleus. Data are presented as the mean standard deviation. **p
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
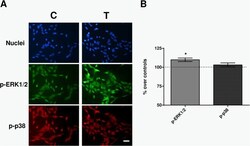
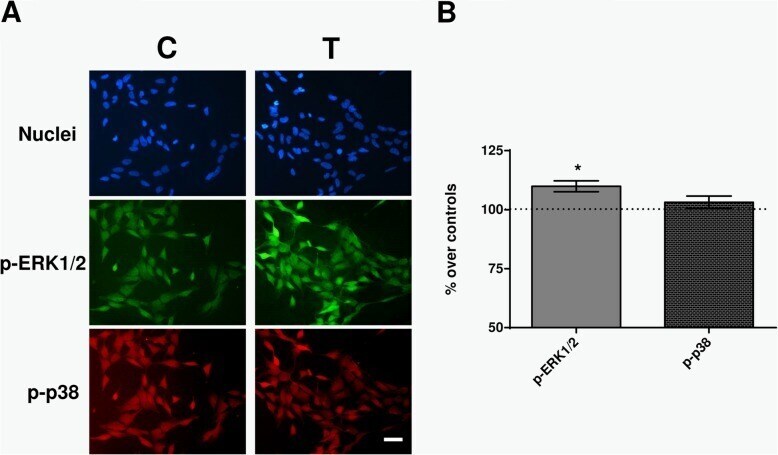
- Fig. 6 Immunofluorescence. a Immunofluorescence for p-ERK and p-p38 at 30 min of sham- or CRET-treatment: representative micrographs. Alexa Green for p-ERK, Alexa Red for p-p38 and Hoechst 33258 for DNA. C = Control, T = Treated with 448 kHz at 50 muA/mm 2 . Bar = 100 mum. b Quantification of p-ERK and p-p38 positive cells. Values are means +- SEM of 3 experimental replicates, normalized over sham-exposed controls. *: 0.05 > p >= 0.01; Student's t-test
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
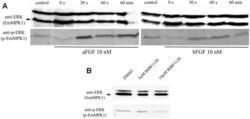
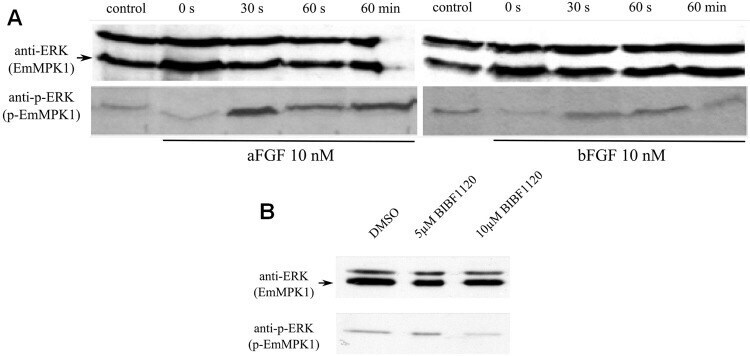
- Experimental details
- Fig 7 Effects of host FGFs and BIBF 1120 on EmMPK1 phosphorylation in metacestode vesicles. A , axenically cultivated metacestode vesicles were incubated in cMEM (control) or in medium without FCS (0 s), upon which FGF1 (aFGF) or FGF2 (bFGF) were added at a concentration of 10 nM for 30 sec (30s), 60 sec (60s) or 60 min (60min). Protein lysates were subsequently separated by 12% SDS-PAGE and Western blots were analysed using polyclonal antibodies against Erk-like MAP kinases (anti-ERK) or double phosphorylated Erk-like MAP kinases (anti-p-ERK). B , axenically cultivated metacestode vesicles were incubated with DMSO (negative control), 5 mM or 10 mM BIBF1120 (30 min each) and cell lysates were subsequently analysed as described above. Both experiments were performed in triplicate with similar results.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
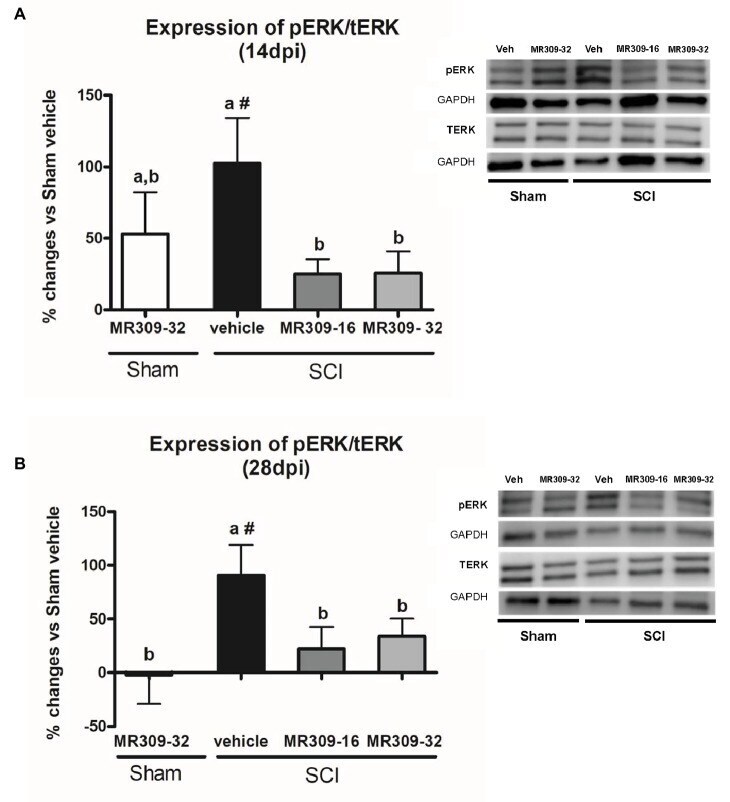
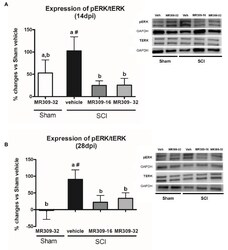
- Figure 2 Spinal ERK1/2 phosphorylation (pERK) expression after preventive sigma-1 receptor antagonist MR309 treatment. Quantification and representative immunoblots of total ERK (tERK), pERK, and glyceraldehyde 3-phosphate dehydrogenase (GAPDH). Experimental groups: Sham-Veh ( n = 5), Sham-MR309-32 ( n = 5), SCI-Veh ( n = 4), SCI-MR309-16 ( n = 5), and SCI-MR309-32 ( n = 5). Protein expressions were normalized to GAPDH, and data are presented as a percentage respect to SCI-Veh mice. ANOVA analysis revealed significant differences at both 14 ( F (4,23) = 3.54, p = 0.025) and 28 ( F (4,24) = 3.65, p = 0.022) days post-injury. a, b: groups not sharing a letter are significantly different, p < 0.05, by Duncan's test; #: significant differences vs. SCI-Veh ( p < 0.05, Duncan's test). MR309 treatments prevent pERK upregulation observed in mild spinal cord injured mice at both 14 (A) and 28 (B) days post-injury. Control images were reused either for illustrative purposes or methodological purposes when several protein levels were assessed in one blot. Full-length blots are presented in Supplementary Figure S1 .