Explore
Explore Validate
Validate Learn
Learn Western blot
Western blotAntibody data
- Antibody Data
- Antigen structure
- References [14]
- Comments [0]
- Validations
- Western blot [2]
- Immunohistochemistry [1]
- Other assay [10]
Submit
Validation data
Reference
Comment
Report error
- Product number
- 14-9109-80 - Provider product page

- Provider
- Invitrogen Antibodies
- Product name
- Phospho-ERK1/2 (Thr202, Tyr204) Monoclonal Antibody (MILAN8R), eBioscience™
- Antibody type
- Monoclonal
- Antigen
- Other
- Description
- Description: This MILAN8R monoclonal antibody recognizes human and mouse extracellular signal-regulated kinases 1 and 2 (also known as ERK1/2, p44/p42, or MAPK3/1) when phosphorylated on T202/Y204. ERK1/2 belong to a family of conserved serine/threonine protein kinases known as mitogen-activated protein kinases (MAPKs) that are involved in many cellular programs such as proliferation, differentiation, motility, and survival. ERK1/2 signaling is activated in response to numerous extracellular stimuli including mitogens, growth factors, and cytokines. The primary activators of ERK1/2 are MEK1 and MEK2 which act by phosphorylating the activation loop residues T202/Y204 and T185/Y187 in ERK1 and ERK2, respectively. Several downstream targets of ERK1/2 have been identified, including p90RSK and the transcription factor Elk-1. ERK1/2 are negatively regulated by MAPK phosphatases, known as DUSPs or MKPs, as well as by chemical inhibitors of MEK including U0126 and PD98059. Disruption of the ERK pathway is common in many types of cancer.
- Antibody clone number
- MILAN8R
- Concentration
- 0.5 mg/mL
Submitted references Suppression of 4.1R enhances the potency of NKG2D-CAR T cells against pancreatic carcinoma via activating ERK signaling pathway.
MM-129 as a Novel Inhibitor Targeting PI3K/AKT/mTOR and PD-L1 in Colorectal Cancer.
Antitumor and Radiosensitizing Effects of Zinc Oxide-Caffeic Acid Nanoparticles against Solid Ehrlich Carcinoma in Female Mice.
Fibroblast subtypes define a metastatic matrisome in breast cancer.
Cancer cell-intrinsic expression of MHC II in lung cancer cell lines is actively restricted by MEK/ERK signaling and epigenetic mechanisms.
Immune myopathy with large histiocyte-related myofiber necrosis.
Cas9-mediated excision of proximal DNaseI/H3K4me3 signatures confers robust silencing of microRNA and long non-coding RNA genes.
Butyrate upregulates the TLR4 expression and the phosphorylation of MAPKs and NK-κB in colon cancer cell in vitro.
MicroRNA-126 deficiency enhanced the activation and function of CD4(+) T cells by elevating IRS-1 pathway.
IL-4/IL-13 Signaling Inhibits the Potential of Early Thymic Progenitors To Commit to the T Cell Lineage.
Heterogeneity of leukemia-initiating capacity of chronic myelogenous leukemia stem cells.
Germline and somatic FGFR1 abnormalities in dysembryoplastic neuroepithelial tumors.
The Bacterial Enzyme IdeS Cleaves the IgG-Type of B Cell Receptor (BCR), Abolishes BCR-Mediated Cell Signaling, and Inhibits Memory B Cell Activation.
Fish Oil-Rich Diet Promotes Hematopoiesis and Alters Hematopoietic Niche.
Gao Y, Lin H, Guo D, Cheng S, Zhou Y, Zhang L, Yao J, Farooq MA, Ajmal I, Duan Y, He C, Tao L, Wu S, Liu M, Jiang W
Oncogenesis 2021 Sep 21;10(9):62
Oncogenesis 2021 Sep 21;10(9):62
MM-129 as a Novel Inhibitor Targeting PI3K/AKT/mTOR and PD-L1 in Colorectal Cancer.
Hermanowicz JM, Pawlak K, Sieklucka B, Czarnomysy R, Kwiatkowska I, Kazberuk A, Surazynski A, Mojzych M, Pawlak D
Cancers 2021 Jun 26;13(13)
Cancers 2021 Jun 26;13(13)
Antitumor and Radiosensitizing Effects of Zinc Oxide-Caffeic Acid Nanoparticles against Solid Ehrlich Carcinoma in Female Mice.
Sayed HM, Said MM, Morcos NYS, El Gawish MA, Ismail AFM
Integrative cancer therapies 2021 Jan-Dec;20:15347354211021920
Integrative cancer therapies 2021 Jan-Dec;20:15347354211021920
Fibroblast subtypes define a metastatic matrisome in breast cancer.
Brechbuhl HM, Barrett AS, Kopin E, Hagen JC, Han AL, Gillen AE, Finlay-Schultz J, Cittelly DM, Owens P, Horwitz KB, Sartorius CA, Hansen K, Kabos P
JCI insight 2020 Feb 27;5(4)
JCI insight 2020 Feb 27;5(4)
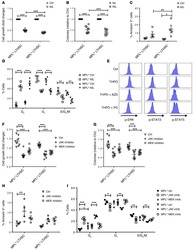
Cancer cell-intrinsic expression of MHC II in lung cancer cell lines is actively restricted by MEK/ERK signaling and epigenetic mechanisms.
Neuwelt AJ, Kimball AK, Johnson AM, Arnold BW, Bullock BL, Kaspar RE, Kleczko EK, Kwak JW, Wu MH, Heasley LE, Doebele RC, Li HY, Nemenoff RA, Clambey ET
Journal for immunotherapy of cancer 2020 Apr;8(1)
Journal for immunotherapy of cancer 2020 Apr;8(1)
Immune myopathy with large histiocyte-related myofiber necrosis.
Pestronk A, Sinha N, Alhumayyd Z, Ly C, Schmidt R, Bucelli R
Neurology 2019 Apr 9;92(15):e1763-e1772
Neurology 2019 Apr 9;92(15):e1763-e1772
Cas9-mediated excision of proximal DNaseI/H3K4me3 signatures confers robust silencing of microRNA and long non-coding RNA genes.
Janga H, Aznaourova M, Boldt F, Damm K, Grünweller A, Schulte LN
PloS one 2018;13(2):e0193066
PloS one 2018;13(2):e0193066
Butyrate upregulates the TLR4 expression and the phosphorylation of MAPKs and NK-κB in colon cancer cell in vitro.
Xiao T, Wu S, Yan C, Zhao C, Jin H, Yan N, Xu J, Wu Y, Li C, Shao Q, Xia S
Oncology letters 2018 Oct;16(4):4439-4447
Oncology letters 2018 Oct;16(4):4439-4447
MicroRNA-126 deficiency enhanced the activation and function of CD4(+) T cells by elevating IRS-1 pathway.
Chu F, Hu Y, Zhou Y, Guo M, Lu J, Zheng W, Xu H, Zhao J, Xu L
Clinical and experimental immunology 2018 Feb;191(2):166-179
Clinical and experimental immunology 2018 Feb;191(2):166-179
IL-4/IL-13 Signaling Inhibits the Potential of Early Thymic Progenitors To Commit to the T Cell Lineage.
Barik S, Miller MM, Cattin-Roy AN, Ukah TK, Chen W, Zaghouani H
Journal of immunology (Baltimore, Md. : 1950) 2017 Oct 15;199(8):2767-2776
Journal of immunology (Baltimore, Md. : 1950) 2017 Oct 15;199(8):2767-2776
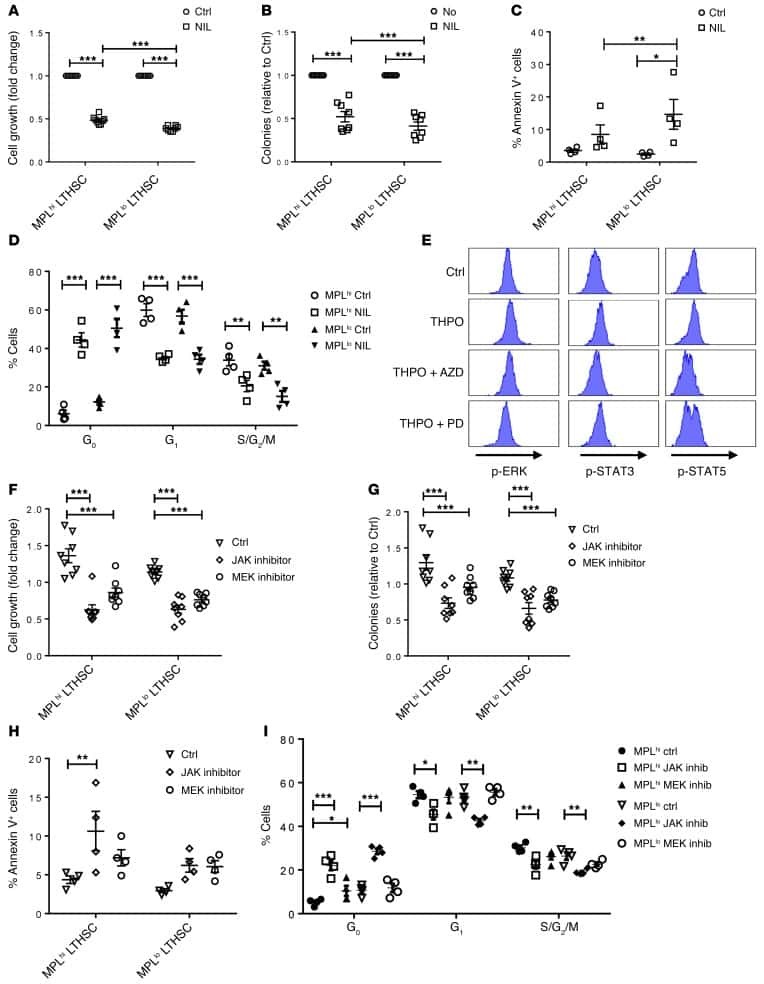
Heterogeneity of leukemia-initiating capacity of chronic myelogenous leukemia stem cells.
Zhang B, Li L, Ho Y, Li M, Marcucci G, Tong W, Bhatia R
The Journal of clinical investigation 2016 Mar 1;126(3):975-91
The Journal of clinical investigation 2016 Mar 1;126(3):975-91
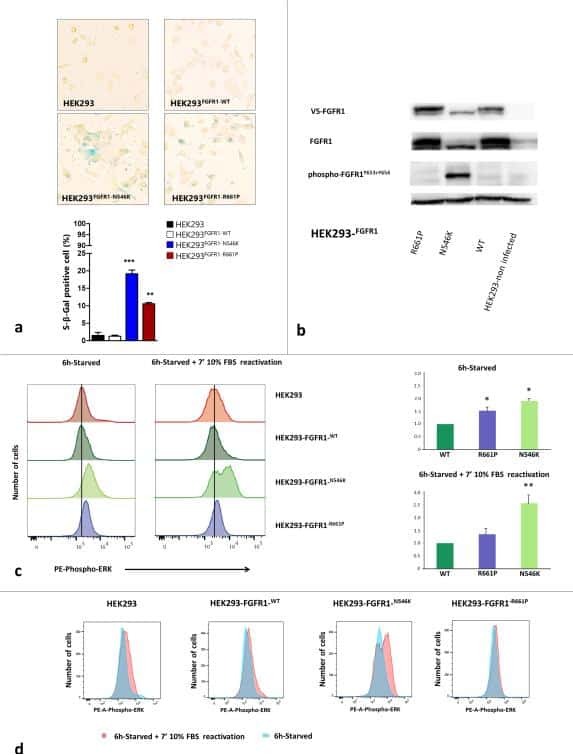
Germline and somatic FGFR1 abnormalities in dysembryoplastic neuroepithelial tumors.
Rivera B, Gayden T, Carrot-Zhang J, Nadaf J, Boshari T, Faury D, Zeinieh M, Blanc R, Burk DL, Fahiminiya S, Bareke E, Schüller U, Monoranu CM, Sträter R, Kerl K, Niederstadt T, Kurlemann G, Ellezam B, Michalak Z, Thom M, Lockhart PJ, Leventer RJ, Ohm M, MacGregor D, Jones D, Karamchandani J, Greenwood CM, Berghuis AM, Bens S, Siebert R, Zakrzewska M, Liberski PP, Zakrzewski K, Sisodiya SM, Paulus W, Albrecht S, Hasselblatt M, Jabado N, Foulkes WD, Majewski J
Acta neuropathologica 2016 Jun;131(6):847-63
Acta neuropathologica 2016 Jun;131(6):847-63
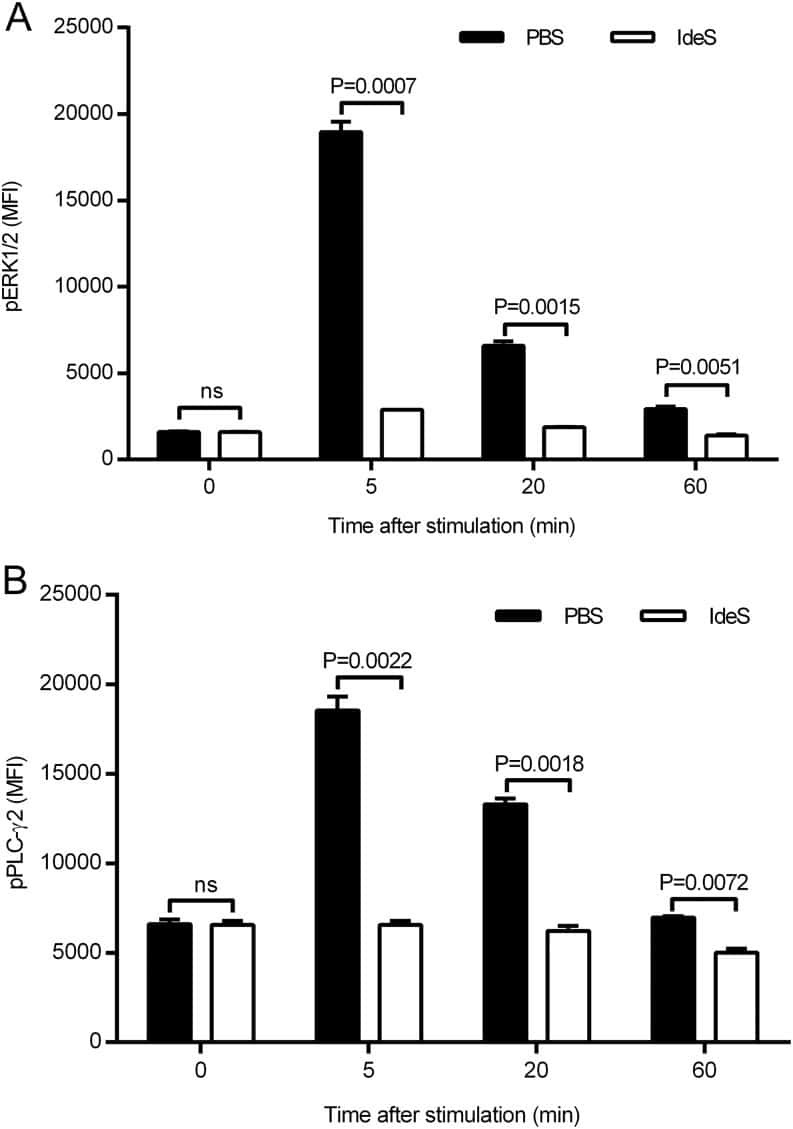
The Bacterial Enzyme IdeS Cleaves the IgG-Type of B Cell Receptor (BCR), Abolishes BCR-Mediated Cell Signaling, and Inhibits Memory B Cell Activation.
Järnum S, Bockermann R, Runström A, Winstedt L, Kjellman C
Journal of immunology (Baltimore, Md. : 1950) 2015 Dec 15;195(12):5592-601
Journal of immunology (Baltimore, Md. : 1950) 2015 Dec 15;195(12):5592-601
Fish Oil-Rich Diet Promotes Hematopoiesis and Alters Hematopoietic Niche.
Xia S, Li XP, Cheng L, Han MT, Zhang MM, Shao QX, Xu HX, Qi L
Endocrinology 2015 Aug;156(8):2821-30
Endocrinology 2015 Aug;156(8):2821-30
No comments: Submit comment
Supportive validation
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
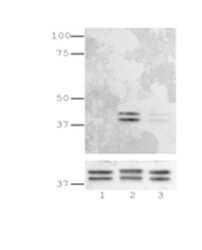
- Immunoblotting of reduced lysates from Jurkat T cells left unstimulated (lane 1), stimulated with PMA+ionomycin (lane 2), or stimulated with PMA+ionomycin in the presence of the MEK1/2 inhibitor, PD98059 (lane 3) using 5 µg/mL of Anti-Human/Mouse phospho-ERK1/2 (T202/Y204) Purified.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
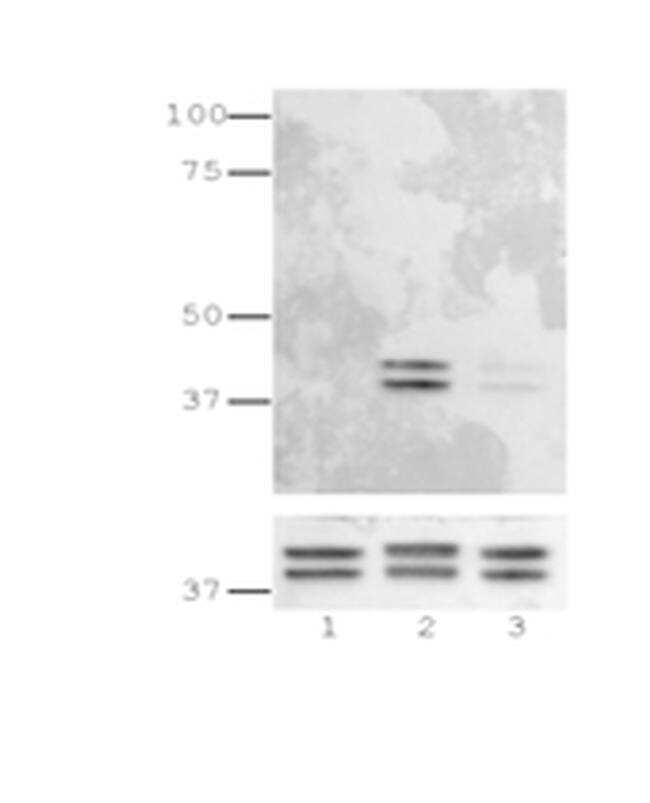
- Western blot analysis was performed on whole cell extracts (30 µg lysate) of A-431 (Lane 1), A-431 serum starved for overnight followed by EGF (200 ng/mL for 10 min) (Lane 2), A-431 treated with Afatinib followed by EGF (0.5 µM for 6hr, 200 ng/mL for 10 min) (Lane 3), Jurkat (Lane 4), Jurkat treated with TPA (200 nM for 10 min) (Lane 5), NIH/3T3 (Lane 6), NIH/3T3 treated with TPA (200 nM for 10 min) (Lane 7) was probed with Anti-Phospho-ERK1/2 (Thr202, Tyr204) Monoclonal Antibody (Product # 14-9109-82, 1 µg/mL) and detected by chemiluminescence using Goat anti-Mouse IgG (H+L) Superclonal™ Secondary Antibody, HRP conjugate (Product # A28177, 0.25 µg/mL, 1:4000 dilution). A 44, 42 kDa band corresponding to Phospho-ERK1/2 (Thr202, Tyr204) was observed upon treatment across the cell lines tested and was inhibited upon Afatinib treatment in A431 cell line.

Supportive validation
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
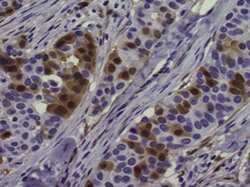
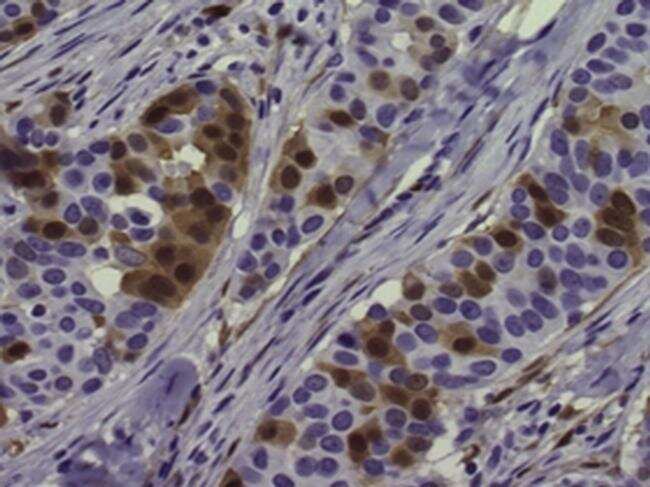
- Immunohistochemistry of formalin-fixed paraffin embedded human breast cancer tissue stained with 5 µg/mL of Anti-Human/Mouse phospho-ERK1/2 (T202/Y204) Purified followed by Anti-Mouse IgG Biotin, Avidin HRP, and DAB visualization.Nuclei are counterstained with hematoxylin.
Supportive validation
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
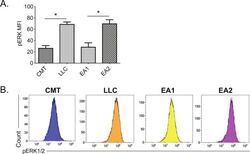
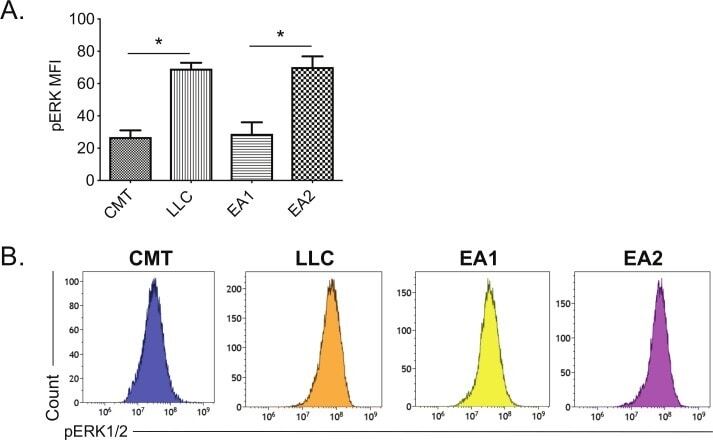
- Figure 3 Correlation between MHC II induction and basal phosphorylated ERK1/2 levels in murine NSCLC cells. Mouse NSCLC lines were cultured for 48 hours, and analyzed for basal levels of intracellular pERK1/2 by flow cytometric analysis. (A) Relative pERK1/2 levels in four murine NSCLC cell lines, including (B) representative histograms. Data are from two independent experiments, with n=3 replicates total. Statistical comparisons of pERK levels were focused on comparison of oncogene-matched cancer cell lines; pERK levels were not significantly different between CMT167 and EA1 cells. MFI is x10 6 . Graphs show mean+-SEM. Flow cytometry data analyzed singlets with MFI data visualized x10 6 values. LLC, Lewis lung carcinoma; MFI, median fluorescent intensity; NSCLC, non-small cell lung cancer.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
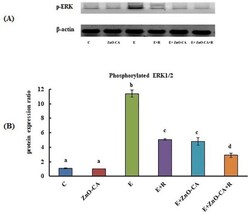
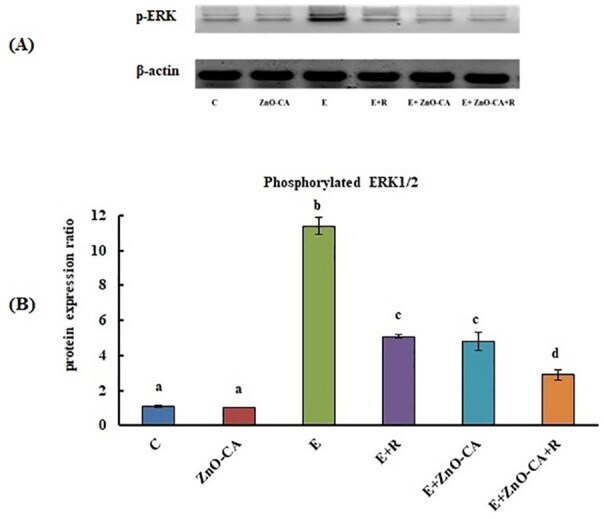
- Figure 4. (A): Western blot analysis of phosphorylated extracellular signal-regulated kinase 1 and 2 (p-ERK1/2) protein expression relative to beta-actin. (B): Effect of gamma-Radiation Exposure and/or Zinc Oxide-Caffeic Acid Nanoparticles treatment on the phosphorylated extracellular signal-regulated kinase 1 and 2 (p-ERK 1/2) protein expression relative to beta-actin in the thigh muscle (control) or tumor tissues. Results are mean +- SE of 3 mice. Values with dissimilar superscripts are significantly different. C: control, E: Solid Ehrlich (EC) bearing mice, R: mice exposed to 2 Gy of whole-body gamma (gamma)-irradiation/week for 3 consecutive weeks, and ZnO-CA: Mice injected intraperitoneally with 5 mg of Zinc Oxide-Caffeic acid nanoparticles (ZnO-CA NPs)/100 g bodyweight, every other day for 3 consecutive weeks. Relative gene expression was measured in thigh muscle of non EC bearing mice (C and ZnO-CA groups) or tumor tissues of solid EC bearing mice of different treated groups. P -value < 0.001.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
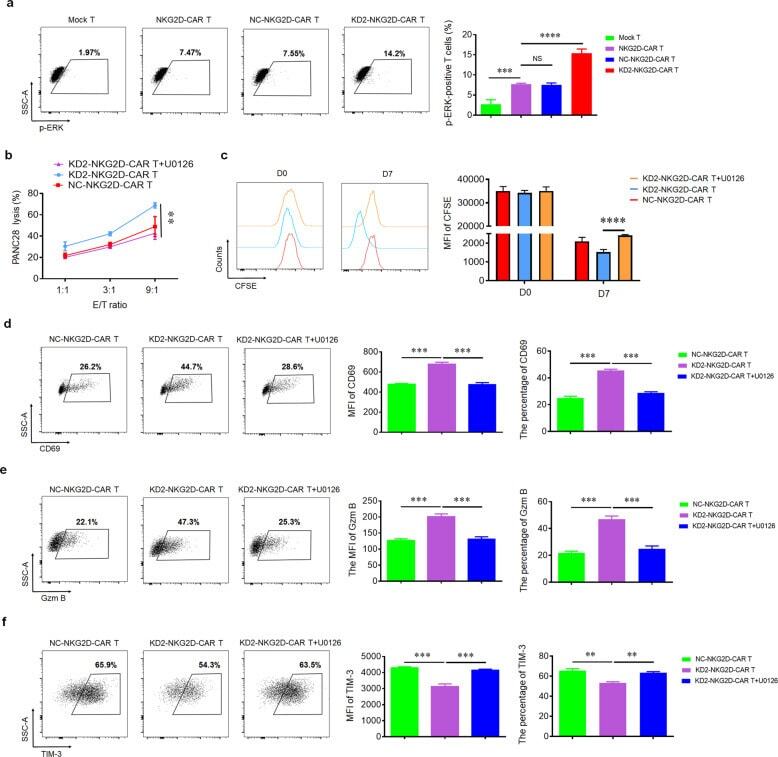
- Fig. 4 4.1R deficiency regulated the function of CAR T cells via ERK signaling pathway. a Mock T, NKG2D-CAR T, NC-NKG2D-CAR T, and KD2-NKG2D-CAR T were co-incubated with PANC28 at a 9:1 ratio for 16 h. The expression of p-ERK was detected by flow cytometry (left), and the percentage of p-ERK-positive T cells was statistically analyzed (right) ( n = 3). b Line plots displayed the cytotoxicity of NC-NKG2D-CAR T and KD2-NKG2D-CAR T against PANC28 at a different effector to target ( E : T ) ratios for 16 h in the absence and presence of 10 muM U0126. c NC-NKG2D-CAR T and KD2-NKG2D-CAR T were co-incubated with PANC28 at a different effector to target ( E : T ) ratios for 7 days in the absence and presence of 10 muM U0126. CFSE dilution was used as a measure of cell proliferation (left), and MFI was calculated (right) ( n = 3). NC-NKG2D-CAR T and KD2-NKG2D-CAR T were co-incubated with PANC28 at a different effector to target ( E : T ) ratios for 16 h in the absence and presence of 10 muM U0126. The expression of CD69 ( d ), Gzm B ( e ), and TIM-3 ( f ) was detected by flow cytometry (left). MFI and percentage were statistically analyzed and shown in column chart (middle and right) ( n = 3). Data were representative of three independent experiments. ** P < 0.01, *** P < 0.001, NS not significant.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
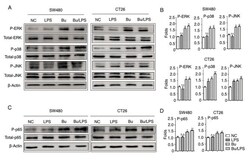
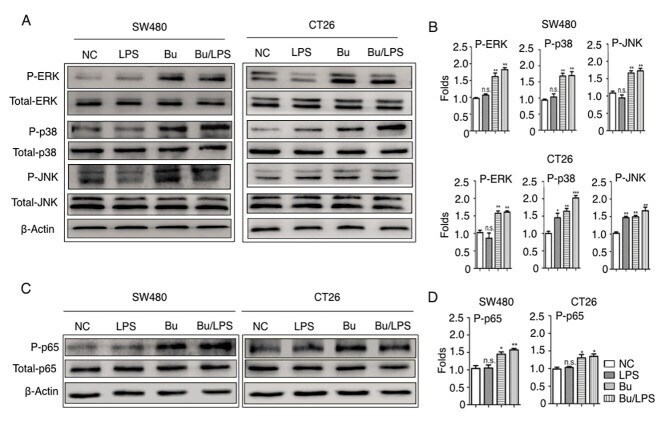
- Figure 5. Butyrate modifies the phosphorylation of ERK, p38, JNK and NF-kappaB p65 in colon cancer cells. The phosphorylation of ERK, p38, JNK and NF-kappaB p65 proteins in SW480 and CT26 cells treated with butyrate and/or LPS were detected using a western blot analysis assay. (A) Representative western blot analysis images of P-ERK/ERK, P-p38/p38 and P-JNK/JNK are presented. (B) The ratios of P-ERK/ERK, P-p38/p38 and P-JNK/JNK protein intensities were calculated. (C) Representative western blot analysis images of P-NF-kappaB p65/NF-kappaB p65 protein expression in cells are presented. (D) P-NF-kappaB p65/p65 protein intensities were calculated. beta-actin was used as the control. Experiments were performed in triplicate and data are expressed as the mean +- standard deviation. Compared with the NC group, *P
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
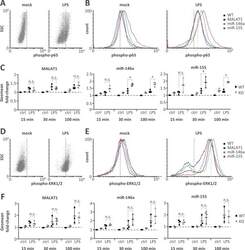
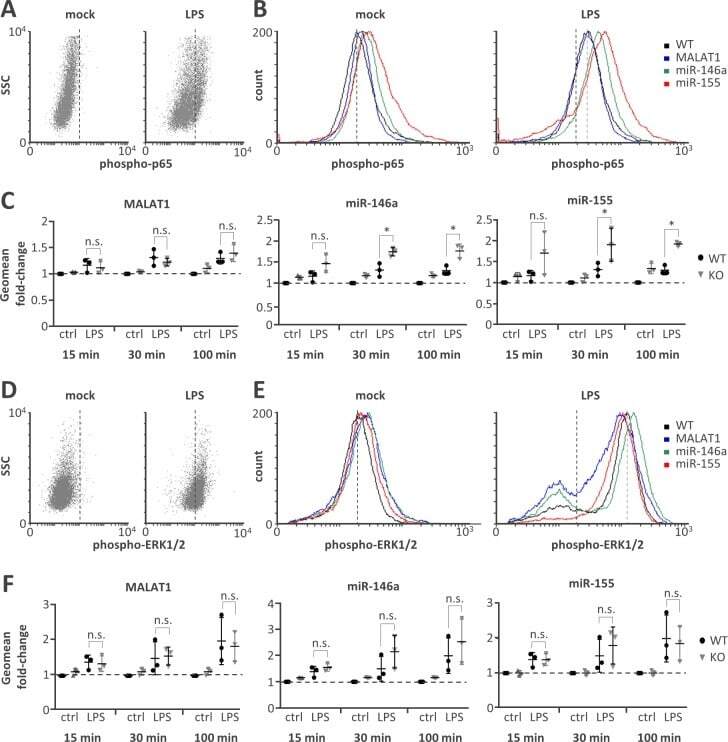
- Fig 4 Elevated NFkappaB p65 but not ERK1/2 activity on miR-146a and miR-155 knockout. A) Representative FACS scatter plots showing a right-shift of 30 min LPS-stimulated (1 mug / ml) compared to mock-treated monocytes stained with phospho-p65 antibody (PE-channel). B) Representative histogram plots showing an increased right-shift of miR-146a and miR-155 deficient compared to control or MALAT1 deficient monocytes after 30 min LPS-stimulation (1 mug / ml) and staining with a phospho-p65 antibody (PE-channel). C) Fold change in phospho-p65 signal in monocytes stimulated with LPS (1 mug / ml) for 15, 30 or 100 min compared to mock-treatment (ctrl) in wild-type (WT) or the indicated ncRNA knockout (KO) cells. All fold-changes are relative to the respective WT mock control. D-F) Same as A-C) but with phospho-ERK1/2 staining (APC-channel). Statistical significance was determined by a one-way ANOVA test with multiple comparisons (* p
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
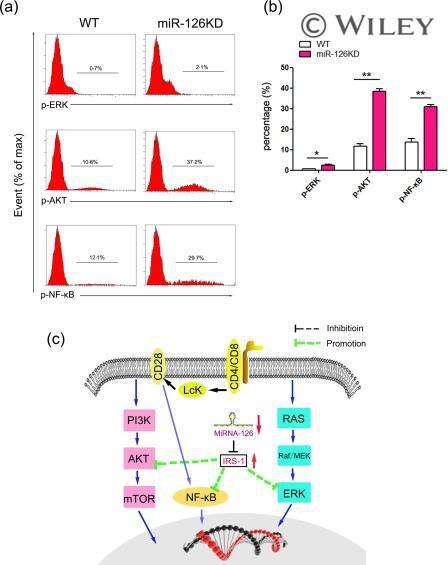
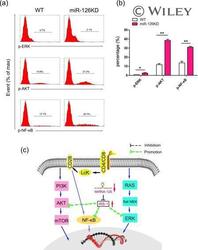
- The effect of microRNA-126 (Mir-126) deficiency on the transduction of related signalling pathway. CD4 + CD62L + T cells purified by magnetic-activated cell sorting (MACS) from miR-126 knock-down (KD) mice and wild-type (WT) mice (8-10 weeks old, n = 6), respectively, Then, cells were cultured in the presence of anti-CD3 (20 mug/ml)/anti-CD28 (4 ng/ml) antibody plus interleukin (IL)-2 (10 ng/ml) for 48 h. (a) The expression levels of phospho-extracellular regulated kinase (p-ERK), phospho-protein kinase B (p-AKT) and phospho-nuclear factor kinase kappa B (p-NF-kappaB) were determined by fluorescence activated cell sorting (FACS) and calculated (b). (c) Schematic representation of the underlying mechanism of miR-126 deficiency on the activation and function of CD4 + T cells. miR-126 deficiency leads to up-regulation of insulin receptor substrate 1 (IRS-1), which could successively alter the transduction of the AKT, ERK and NF-kappaB signalling pathway. [Colour figure can be viewed at wileyonlinelibrary.com ]