Explore
Explore Validate
Validate Learn
Learn16-7222-82
antibody from Invitrogen Antibodies
Targeting: IL22
IL-21, IL-22, IL-D110, IL-TIF, ILTIF, MGC79382, MGC79384, TIFa, TIFIL-23, zcyto18
 Blocking/Neutralizing
Blocking/Neutralizing Functional assay
Functional assayAntibody data
- Antibody Data
- Antigen structure
- References [31]
- Comments [0]
- Validations
- Functional assay [1]
- Other assay [28]
Submit
Validation data
Reference
Comment
Report error
- Product number
- 16-7222-82 - Provider product page

- Provider
- Invitrogen Antibodies
- Product name
- IL-22 Monoclonal Antibody (IL22JOP), Functional Grade, eBioscience™
- Antibody type
- Monoclonal
- Antigen
- Other
- Description
- Description: The monoclonal antibody IL22JOP reacts with and inhibits the bioactivity of human and mouse IL-22. IL-22 is a 20 kDa member of the IL-10 cytokine family that is secreted primarily by Th17 cells, NK cells, and other T cells. Compared to IL-6 or TGF beta, IL-23 can induce greater levels of IL-22 in in vitro-differentiated Th17 cells. This observation suggests that IL-22 may be secreted by more fully differentiated Th17 cells in vivo. Recently, it was demonstrated that IL-22 could protect hosts from bacterial infection of the lungs and gut. Moreover, it has been reported that anti-CD3/CD28-induced production of IL-22 by PBMCs was elevated significantly in asthma patients compared to control patients. Flow cytometric analysis also showed that the frequencies of IL-17+IL-22+ CD4 T cells were increased in PBMCs from patients with ankylosing spondylitis and rheumatoid arthritis. IL22JOP is published to recognize rhesus IL-22. Applications Reported: The monoclonal antibody IL22JOP reacts with and inhibits the bioactivity of human and mouse IL-22. Applications Tested: The ND50 of IL22JOP, as determined by the inhibition of IL-10 induction in COLO205 cells by IL-22, is 10-40 ng/mL, in the presence of 0.2 ng/mL recombinant human or mouse IL-22. Neutralization dose will vary depending on assay method, cytokine concentration, and cell type. Storage and handling: Use in a sterile environment. Filtration: 0.2 µm post-manufacturing filtered. Purity: Greater than 90%, as determined by SDS-PAGE. Endotoxin Level: Less than 0.001 ng/µg antibody, as determined by LAL assay. Aggregation: Less than 10%, as determined by HPLC.
- Reactivity
- Human, Mouse
- Host
- Rat
- Isotype
- IgG
- Antibody clone number
- IL22JOP
- Vial size
- 100 μg
- Concentration
- 1 mg/mL
- Storage
- 4°C
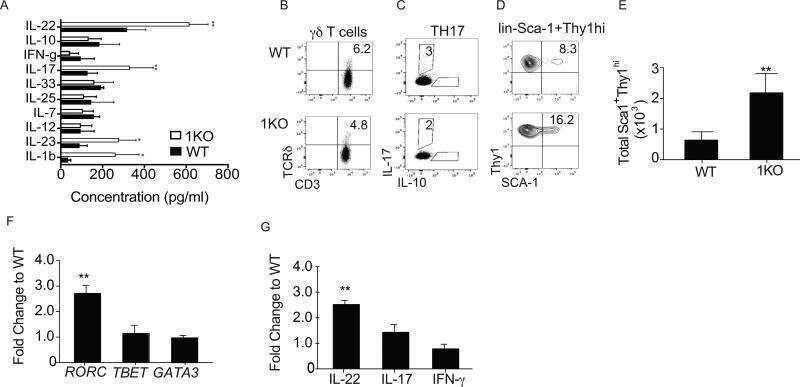
Submitted references Mitochondrial transcription factor A in RORγt(+) lymphocytes regulate small intestine homeostasis and metabolism.
Expanded IL-22(+) Group 3 Innate Lymphoid Cells and Role of Oxidized LDL-C in the Pathogenesis of Axial Spondyloarthritis with Dyslipidaemia.
Chlamydia muridarum Alleviates Colitis via the IL-22/Occludin Signal Pathway.
A Central Role for Lipocalin-2 in the Adaptation to Short-Bowel Syndrome Through Down-Regulation of IL22 in Mice.
T cell-intrinsic role for Nod2 in protection against Th17-mediated uveitis.
Differential Activation of Unconventional T Cells, Including iNKT Cells, in Alcohol-Related Liver Disease.
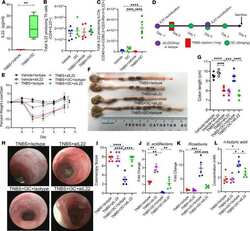
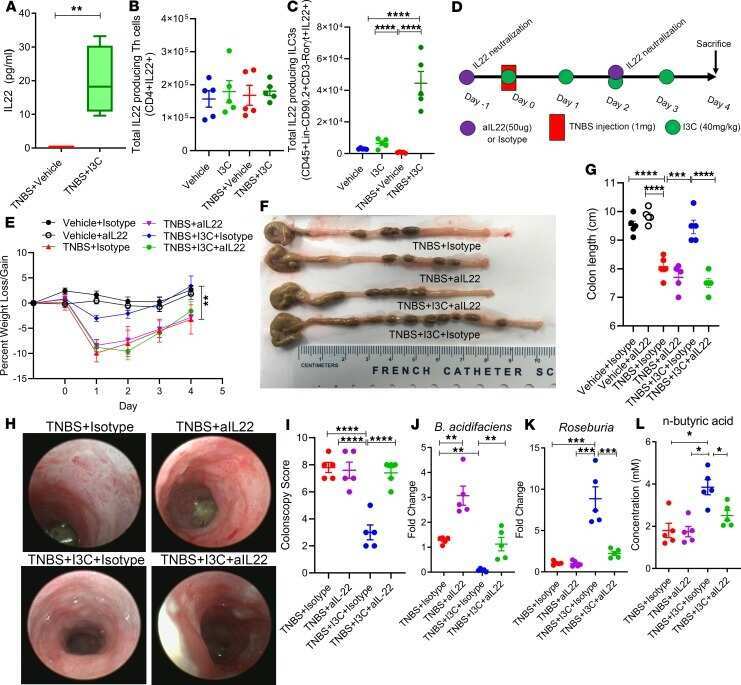
Indole-3-carbinol prevents colitis and associated microbial dysbiosis in an IL-22-dependent manner.
Single-cell multi-omics analysis presents the landscape of peripheral blood T-cell subsets in human chronic prostatitis/chronic pelvic pain syndrome.
CD4 T cell-intrinsic role for the T helper 17 signature cytokine IL-17: Effector resistance to immune suppression.
Induction of autophagy in Cx3cr1(+) mononuclear cells limits IL-23/IL-22 axis-mediated intestinal fibrosis.
The Intestine Harbors Functionally Distinct Homeostatic Tissue-Resident and Inflammatory Th17 Cells.
Suppressive and Gut-Reparative Functions of Human Type 1 T Regulatory Cells.
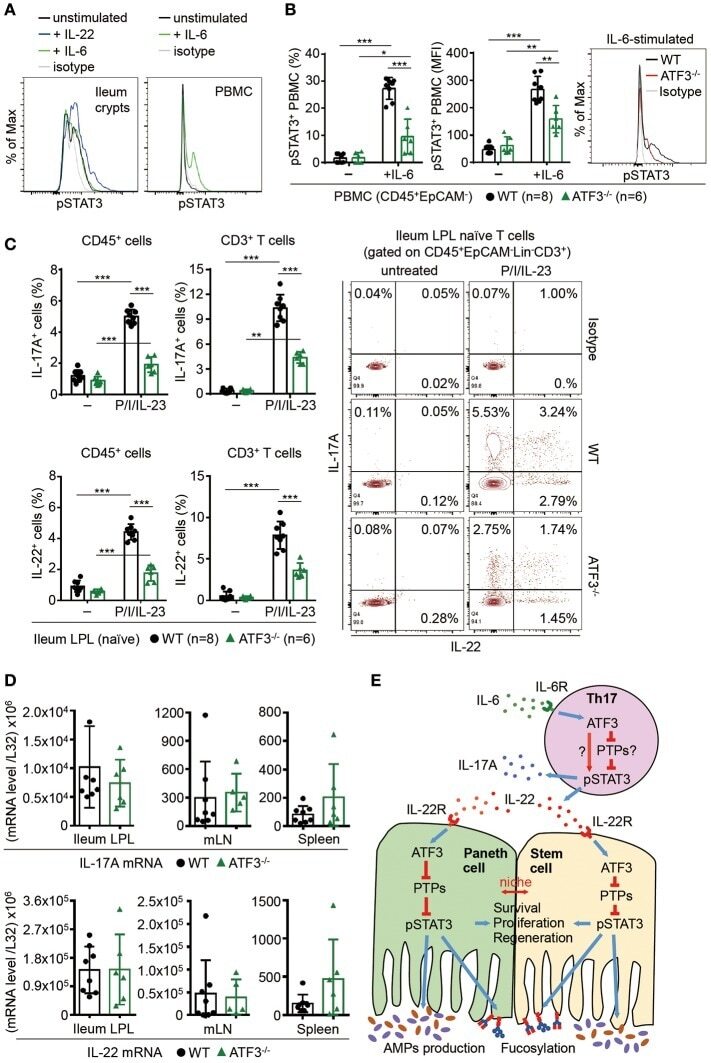
ATF3 Sustains IL-22-Induced STAT3 Phosphorylation to Maintain Mucosal Immunity Through Inhibiting Phosphatases.
CXCL4 is a novel inducer of human Th17 cells and correlates with IL-17 and IL-22 in psoriatic arthritis.
Innate Recognition of the Microbiota by TLR1 Promotes Epithelial Homeostasis and Prevents Chronic Inflammation.
Flt3 ligand expands bona fide innate lymphoid cell precursors in vivo.
WASH maintains NKp46(+) ILC3 cells by promoting AHR expression.
Proteomics analysis reveals a Th17-prone cell population in presymptomatic graft-versus-host disease.
A TLR9 agonist promotes IL-22-dependent pancreatic islet allograft survival in type 1 diabetic mice.
CD69 controls the uptake of L-tryptophan through LAT1-CD98 and AhR-dependent secretion of IL-22 in psoriasis.
Antibiotics in neonatal life increase murine susceptibility to experimental psoriasis.
Joint production of IL-22 participates in the initial phase of antigen-induced arthritis through IL-1β production.
The secretion of IL-22 from mucosal NKp44⁺ NK cells is associated with microbial translocation and virus infection in SIV/SHIV-infected Chinese macaques.
Commensal bacteria protect against food allergen sensitization.
Interleukin-22 promotes T helper 1 (Th1)/Th17 immunity in chlamydial lung infection.
OMIP-022: Comprehensive assessment of antigen-specific human T-cell functionality and memory.
Mechanisms of NDV-3 vaccine efficacy in MRSA skin versus invasive infection.
Rate of AIDS progression is associated with gastrointestinal dysfunction in simian immunodeficiency virus-infected pigtail macaques.
Retinoic acid expression associates with enhanced IL-22 production by γδ T cells and innate lymphoid cells and attenuation of intestinal inflammation.
Gluten-sensitive enteropathy coincides with decreased capability of intestinal T cells to secrete IL-17 and IL-22 in a macaque model for celiac disease.
Inpatient capillary glucose monitoring: a useful adjunct to the management of diabetes in community hospitals.
Fu Z, Dean JW, Xiong L, Dougherty MW, Oliff KN, Chen ZE, Jobin C, Garrett TJ, Zhou L
Nature communications 2021 Jul 22;12(1):4462
Nature communications 2021 Jul 22;12(1):4462
Expanded IL-22(+) Group 3 Innate Lymphoid Cells and Role of Oxidized LDL-C in the Pathogenesis of Axial Spondyloarthritis with Dyslipidaemia.
Min HK, Moon J, Lee SY, Lee AR, Lee CR, Lee J, Kwok SK, Cho ML, Park SH
Immune network 2021 Dec;21(6):e43
Immune network 2021 Dec;21(6):e43
Chlamydia muridarum Alleviates Colitis via the IL-22/Occludin Signal Pathway.
Wang X, Zeng HC, Huang YR, He QZ
BioMed research international 2020;2020:8894331
BioMed research international 2020;2020:8894331
A Central Role for Lipocalin-2 in the Adaptation to Short-Bowel Syndrome Through Down-Regulation of IL22 in Mice.
Zhang A, Sodhi CP, Wang M, Shores DR, Fulton W, Prindle T, Brosten S, O'Hare E, Lau A, Ding H, Jia H, Lu P, White JR, Hui J, Sears CL, Hackam DJ, Alaish SM
Cellular and molecular gastroenterology and hepatology 2020;10(2):309-326
Cellular and molecular gastroenterology and hepatology 2020;10(2):309-326
T cell-intrinsic role for Nod2 in protection against Th17-mediated uveitis.
Napier RJ, Lee EJ, Davey MP, Vance EE, Furtado JM, Snow PE, Samson KA, Lashley SJ, Brown BR, Horai R, Mattapallil MJ, Xu B, Callegan MC, Uebelhoer LS, Lancioni CL, Vehe RK, Binstadt BA, Smith JR, Caspi RR, Rosenzweig HL
Nature communications 2020 Oct 26;11(1):5406
Nature communications 2020 Oct 26;11(1):5406
Differential Activation of Unconventional T Cells, Including iNKT Cells, in Alcohol-Related Liver Disease.
Marrero I, Maricic I, Morgan TR, Stolz AA, Schnabl B, Liu ZX, Tsukamoto H, Kumar V
Alcoholism, clinical and experimental research 2020 May;44(5):1061-1074
Alcoholism, clinical and experimental research 2020 May;44(5):1061-1074
Indole-3-carbinol prevents colitis and associated microbial dysbiosis in an IL-22-dependent manner.
Busbee PB, Menzel L, Alrafas HR, Dopkins N, Becker W, Miranda K, Tang C, Chatterjee S, Singh U, Nagarkatti M, Nagarkatti PS
JCI insight 2020 Jan 16;5(1)
JCI insight 2020 Jan 16;5(1)
Single-cell multi-omics analysis presents the landscape of peripheral blood T-cell subsets in human chronic prostatitis/chronic pelvic pain syndrome.
Zhang M, Liu Y, Chen J, Chen L, Meng J, Yang C, Yin S, Zhang X, Zhang L, Hao Z, Chen X, Liang C
Journal of cellular and molecular medicine 2020 Dec;24(23):14099-14109
Journal of cellular and molecular medicine 2020 Dec;24(23):14099-14109
CD4 T cell-intrinsic role for the T helper 17 signature cytokine IL-17: Effector resistance to immune suppression.
Crawford MP, Sinha S, Renavikar PS, Borcherding N, Karandikar NJ
Proceedings of the National Academy of Sciences of the United States of America 2020 Aug 11;117(32):19408-19414
Proceedings of the National Academy of Sciences of the United States of America 2020 Aug 11;117(32):19408-19414
Induction of autophagy in Cx3cr1(+) mononuclear cells limits IL-23/IL-22 axis-mediated intestinal fibrosis.
Mathur R, Alam MM, Zhao XF, Liao Y, Shen J, Morgan S, Huang T, Lee H, Lee E, Huang Y, Zhu X
Mucosal immunology 2019 May;12(3):612-623
Mucosal immunology 2019 May;12(3):612-623
The Intestine Harbors Functionally Distinct Homeostatic Tissue-Resident and Inflammatory Th17 Cells.
Omenetti S, Bussi C, Metidji A, Iseppon A, Lee S, Tolaini M, Li Y, Kelly G, Chakravarty P, Shoaie S, Gutierrez MG, Stockinger B
Immunity 2019 Jul 16;51(1):77-89.e6
Immunity 2019 Jul 16;51(1):77-89.e6
Suppressive and Gut-Reparative Functions of Human Type 1 T Regulatory Cells.
Cook L, Stahl M, Han X, Nazli A, MacDonald KN, Wong MQ, Tsai K, Dizzell S, Jacobson K, Bressler B, Kaushic C, Vallance BA, Steiner TS, Levings MK
Gastroenterology 2019 Dec;157(6):1584-1598
Gastroenterology 2019 Dec;157(6):1584-1598
ATF3 Sustains IL-22-Induced STAT3 Phosphorylation to Maintain Mucosal Immunity Through Inhibiting Phosphatases.
Glal D, Sudhakar JN, Lu HH, Liu MC, Chiang HY, Liu YC, Cheng CF, Shui JW
Frontiers in immunology 2018;9:2522
Frontiers in immunology 2018;9:2522
CXCL4 is a novel inducer of human Th17 cells and correlates with IL-17 and IL-22 in psoriatic arthritis.
Affandi AJ, Silva-Cardoso SC, Garcia S, Leijten EFA, van Kempen TS, Marut W, van Roon JAG, Radstake TRDJ
European journal of immunology 2018 Mar;48(3):522-531
European journal of immunology 2018 Mar;48(3):522-531
Innate Recognition of the Microbiota by TLR1 Promotes Epithelial Homeostasis and Prevents Chronic Inflammation.
Kamdar K, Johnson AMF, Chac D, Myers K, Kulur V, Truevillian K, DePaolo RW
Journal of immunology (Baltimore, Md. : 1950) 2018 Jul 1;201(1):230-242
Journal of immunology (Baltimore, Md. : 1950) 2018 Jul 1;201(1):230-242
Flt3 ligand expands bona fide innate lymphoid cell precursors in vivo.
Parigi SM, Czarnewski P, Das S, Steeg C, Brockmann L, Fernandez-Gaitero S, Yman V, Forkel M, Höög C, Mjösberg J, Westerberg L, Färnert A, Huber S, Jacobs T, Villablanca EJ
Scientific reports 2018 Jan 9;8(1):154
Scientific reports 2018 Jan 9;8(1):154
WASH maintains NKp46(+) ILC3 cells by promoting AHR expression.
Xia P, Liu J, Wang S, Ye B, Du Y, Xiong Z, Han ZG, Tong L, Fan Z
Nature communications 2017 Jun 7;8:15685
Nature communications 2017 Jun 7;8:15685
Proteomics analysis reveals a Th17-prone cell population in presymptomatic graft-versus-host disease.
Li W, Liu L, Gomez A, Zhang J, Ramadan A, Zhang Q, Choi SW, Zhang P, Greenson JK, Liu C, Jiang D, Virts E, Kelich SL, Chu HW, Flynn R, Blazar BR, Hanenberg H, Hanash S, Paczesny S
JCI insight 2016 May 5;1(6)
JCI insight 2016 May 5;1(6)
A TLR9 agonist promotes IL-22-dependent pancreatic islet allograft survival in type 1 diabetic mice.
Tripathi D, Venkatasubramanian S, Cheekatla SS, Paidipally P, Welch E, Tvinnereim AR, Vankayalapati R
Nature communications 2016 Dec 16;7:13896
Nature communications 2016 Dec 16;7:13896
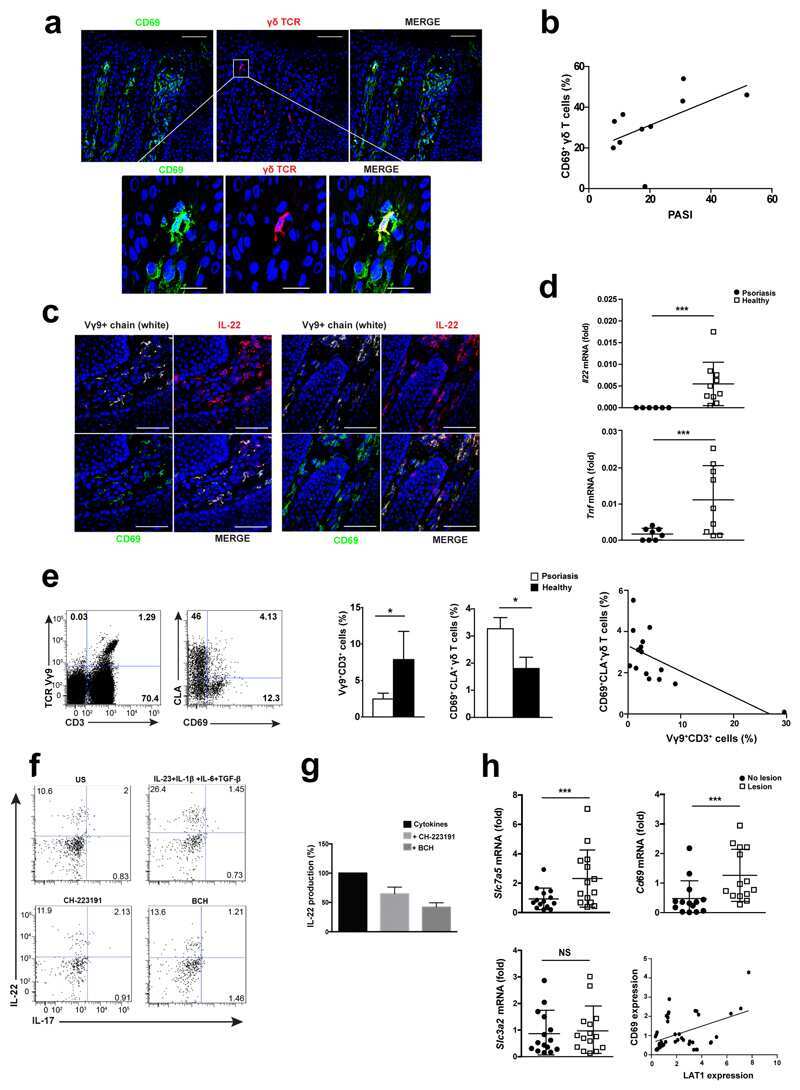
CD69 controls the uptake of L-tryptophan through LAT1-CD98 and AhR-dependent secretion of IL-22 in psoriasis.
Cibrian D, Saiz ML, de la Fuente H, Sánchez-Díaz R, Moreno-Gonzalo O, Jorge I, Ferrarini A, Vázquez J, Punzón C, Fresno M, Vicente-Manzanares M, Daudén E, Fernández-Salguero PM, Martín P, Sánchez-Madrid F
Nature immunology 2016 Aug;17(8):985-96
Nature immunology 2016 Aug;17(8):985-96
Antibiotics in neonatal life increase murine susceptibility to experimental psoriasis.
Zanvit P, Konkel JE, Jiao X, Kasagi S, Zhang D, Wu R, Chia C, Ajami NJ, Smith DP, Petrosino JF, Abbatiello B, Nakatsukasa H, Chen Q, Belkaid Y, Chen ZJ, Chen W
Nature communications 2015 Sep 29;6:8424
Nature communications 2015 Sep 29;6:8424
Joint production of IL-22 participates in the initial phase of antigen-induced arthritis through IL-1β production.
Pinto LG, Talbot J, Peres RS, Franca RF, Ferreira SH, Ryffel B, Aves-Filho JC, Figueiredo F, Cunha TM, Cunha FQ
Arthritis research & therapy 2015 Sep 2;17(1):235
Arthritis research & therapy 2015 Sep 2;17(1):235
The secretion of IL-22 from mucosal NKp44⁺ NK cells is associated with microbial translocation and virus infection in SIV/SHIV-infected Chinese macaques.
Wang W, Wu F, Cong Z, Liu K, Qin C, Wei Q
Journal of immunology research 2014;2014:387950
Journal of immunology research 2014;2014:387950
Commensal bacteria protect against food allergen sensitization.
Stefka AT, Feehley T, Tripathi P, Qiu J, McCoy K, Mazmanian SK, Tjota MY, Seo GY, Cao S, Theriault BR, Antonopoulos DA, Zhou L, Chang EB, Fu YX, Nagler CR
Proceedings of the National Academy of Sciences of the United States of America 2014 Sep 9;111(36):13145-50
Proceedings of the National Academy of Sciences of the United States of America 2014 Sep 9;111(36):13145-50
Interleukin-22 promotes T helper 1 (Th1)/Th17 immunity in chlamydial lung infection.
Peng Y, Gao X, Yang J, Shekhar S, Wang S, Fan Y, Zhao W, Yang X
Molecular medicine (Cambridge, Mass.) 2014 Mar 20;20(1):109-19
Molecular medicine (Cambridge, Mass.) 2014 Mar 20;20(1):109-19
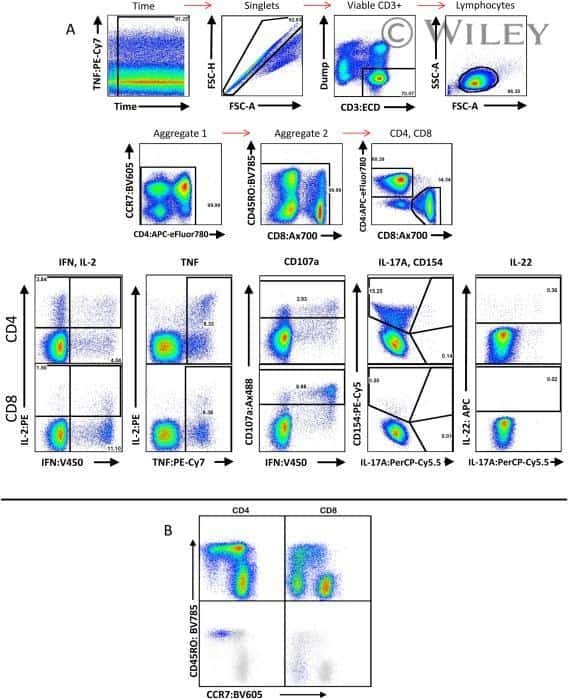
OMIP-022: Comprehensive assessment of antigen-specific human T-cell functionality and memory.
Graves AJ, Padilla MG, Hokey DA
Cytometry. Part A : the journal of the International Society for Analytical Cytology 2014 Jul;85(7):576-9
Cytometry. Part A : the journal of the International Society for Analytical Cytology 2014 Jul;85(7):576-9
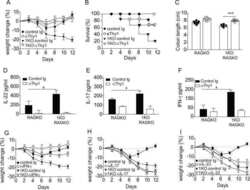
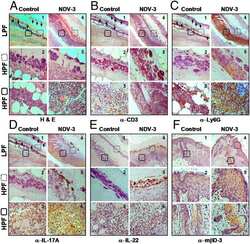
Mechanisms of NDV-3 vaccine efficacy in MRSA skin versus invasive infection.
Yeaman MR, Filler SG, Chaili S, Barr K, Wang H, Kupferwasser D, Hennessey JP Jr, Fu Y, Schmidt CS, Edwards JE Jr, Xiong YQ, Ibrahim AS
Proceedings of the National Academy of Sciences of the United States of America 2014 Dec 23;111(51):E5555-63
Proceedings of the National Academy of Sciences of the United States of America 2014 Dec 23;111(51):E5555-63
Rate of AIDS progression is associated with gastrointestinal dysfunction in simian immunodeficiency virus-infected pigtail macaques.
Canary LA, Vinton CL, Morcock DR, Pierce JB, Estes JD, Brenchley JM, Klatt NR
Journal of immunology (Baltimore, Md. : 1950) 2013 Mar 15;190(6):2959-65
Journal of immunology (Baltimore, Md. : 1950) 2013 Mar 15;190(6):2959-65
Retinoic acid expression associates with enhanced IL-22 production by γδ T cells and innate lymphoid cells and attenuation of intestinal inflammation.
Mielke LA, Jones SA, Raverdeau M, Higgs R, Stefanska A, Groom JR, Misiak A, Dungan LS, Sutton CE, Streubel G, Bracken AP, Mills KH
The Journal of experimental medicine 2013 Jun 3;210(6):1117-24
The Journal of experimental medicine 2013 Jun 3;210(6):1117-24
Gluten-sensitive enteropathy coincides with decreased capability of intestinal T cells to secrete IL-17 and IL-22 in a macaque model for celiac disease.
Xu H, Feely SL, Wang X, Liu DX, Borda JT, Dufour J, Li W, Aye PP, Doxiadis GG, Khosla C, Veazey RS, Sestak K
Clinical immunology (Orlando, Fla.) 2013 Apr;147(1):40-49
Clinical immunology (Orlando, Fla.) 2013 Apr;147(1):40-49
Inpatient capillary glucose monitoring: a useful adjunct to the management of diabetes in community hospitals.
Borst GC, Rice S, Robinson WL
The Journal of the Kentucky Medical Association 1989 Sep;87(9):451-5
The Journal of the Kentucky Medical Association 1989 Sep;87(9):451-5
No comments: Submit comment
Supportive validation
- Submitted by
- Invitrogen Antibodies (provider)
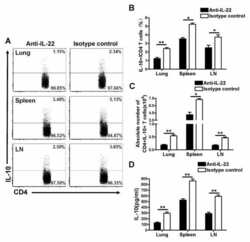
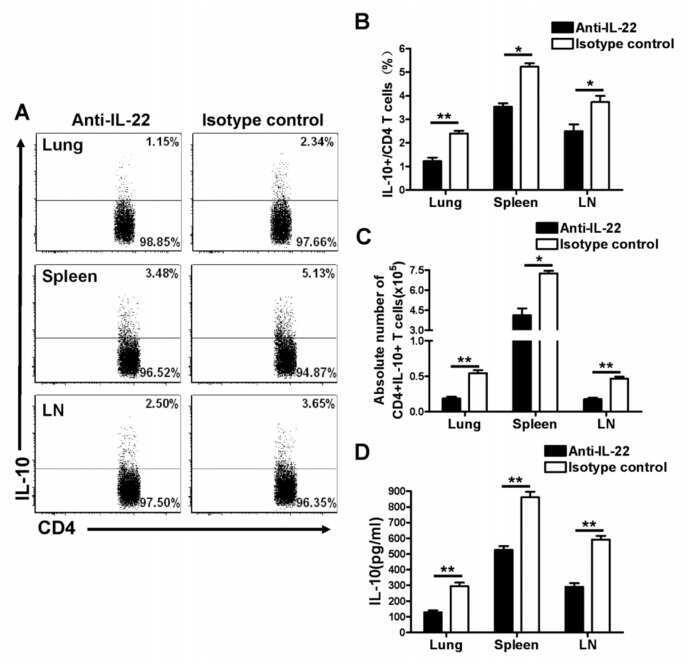
- Main image
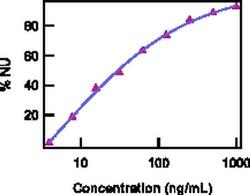
- Experimental details
- Neutralization of IL-22 Recombinant Protein activity as measured by IL-10 induction in COLO205 cells.
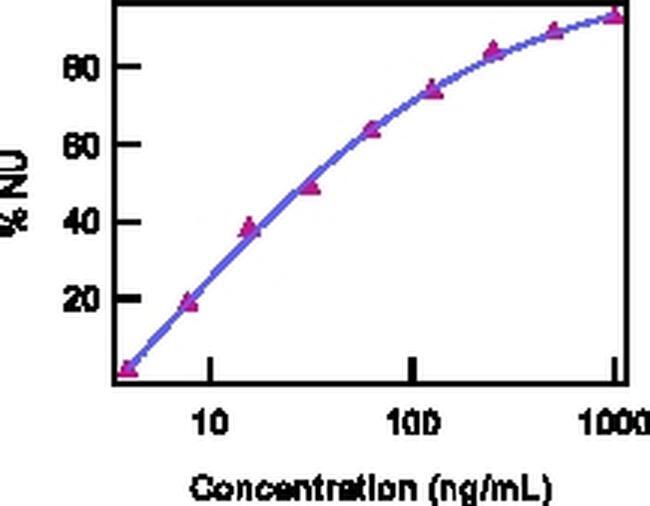
Supportive validation
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
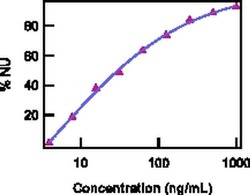
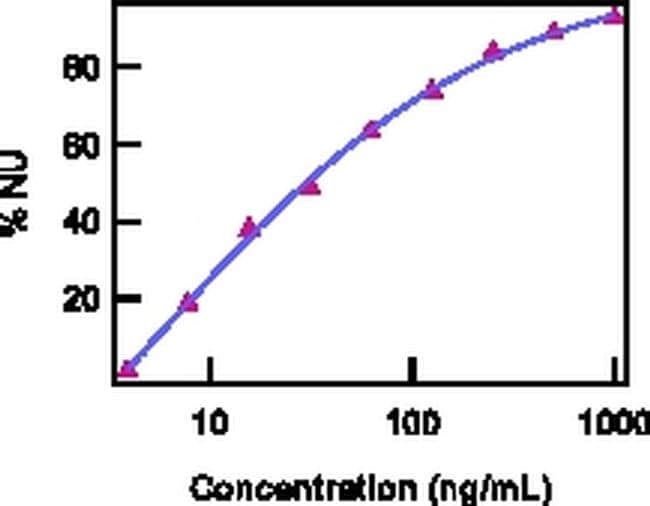
- Neutralization of IL-22 Recombinant Protein activity as measured by IL-10 induction in COLO205 cells.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
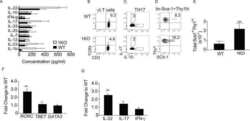
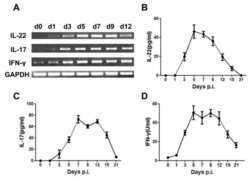
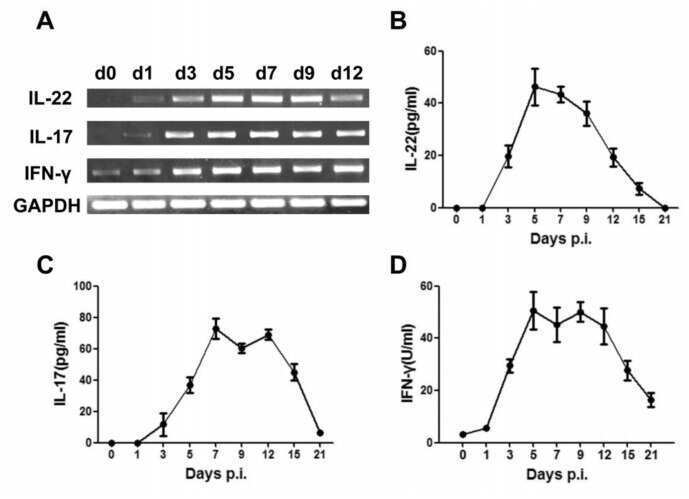
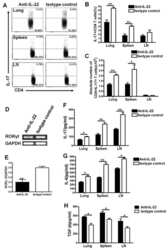
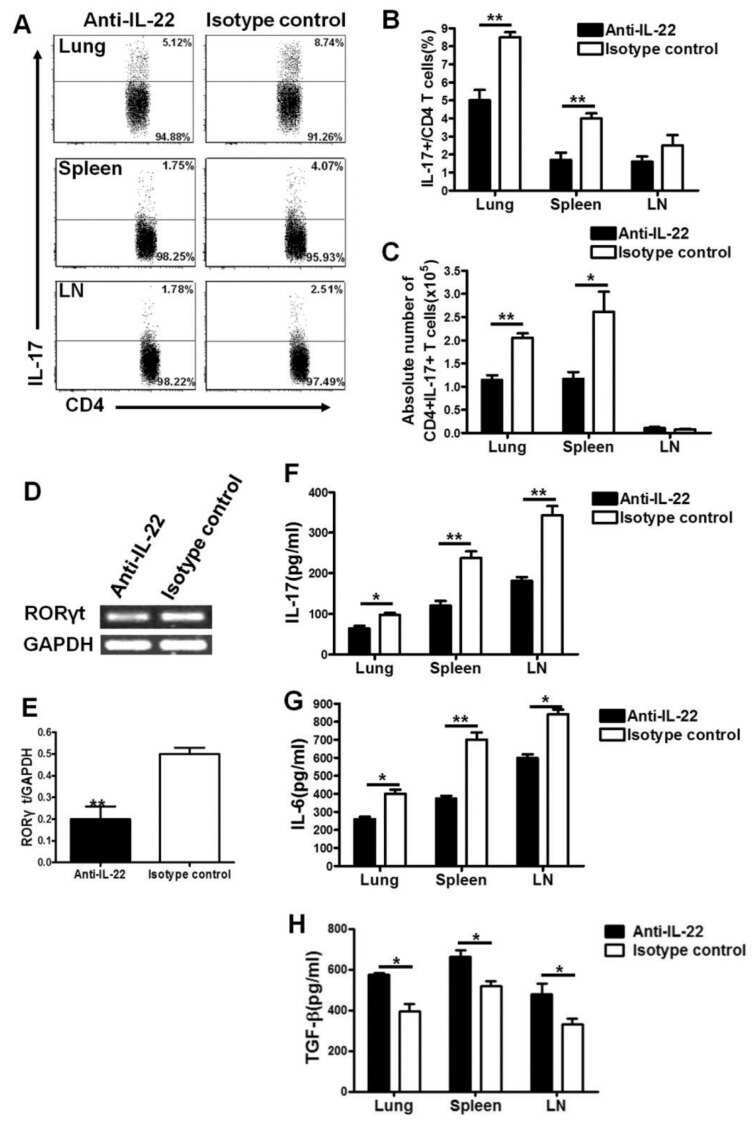
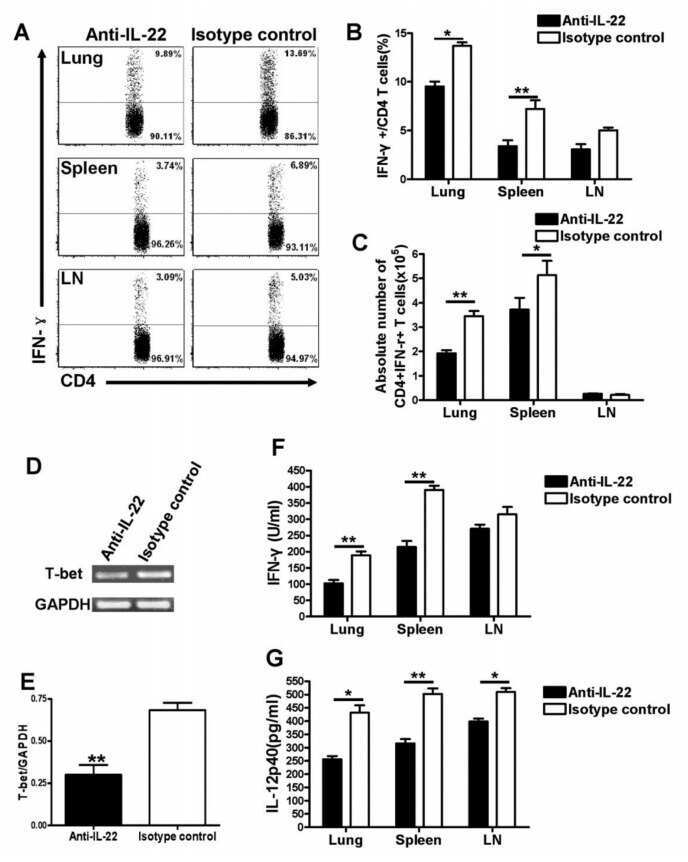
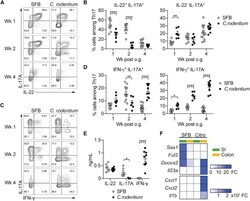
- Figure 2 SFB- and C. - rodentium -Elicited Th17 Cells Have Different Cytokine Profiles (A-D) Representative intracellular staining for IL-17A and IL-22 (A) or IFN-gamma (C) and corresponding quantifications in (B) and (D) in Th17 cells from the SI of mice colonized with SFB and colon of mice infected with C. rodentium at 1 (n = 9 and 6), 2 (n = 11 and 6), and 4 (n = 14 and 9) weeks after gavage. Lamina propria cells were isolated at the indicated time point, re-stimulated with PMA and ionomycin and Brefeldin A for 2 h, and analyzed by FACS. Bars show the mean +- SEM for the indicated populations, and each symbol represents an individual mouse from two pooled independent experiments. (E) Cytokine concentration in supernatants of FACS-purified Th17 cells from the SI of mice colonized with SFB or colon of mice infected with C. rodentium . In the graphs, bars show the mean +- SEM, and each symbol represents a technical replicate. Cells were FACS-purified from one sample per group, obtained by pooling 3-6 mice. Results are representative of three independent experiments. (F) Fold-change (FC) induction of listed genes quantified by qPCR in the SI (green) and colon (orange) of mice colonized with SFB or infected with C. rodentium , 1 week after gavage. Fold change is calculated on the matching organs from untreated mice and gene expression is normalized against beta-2-microglobulin. Data are from two pooled independent experiments. * p < 0.05, ** p < 0.01, **** p < 0.0001 by two-way
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
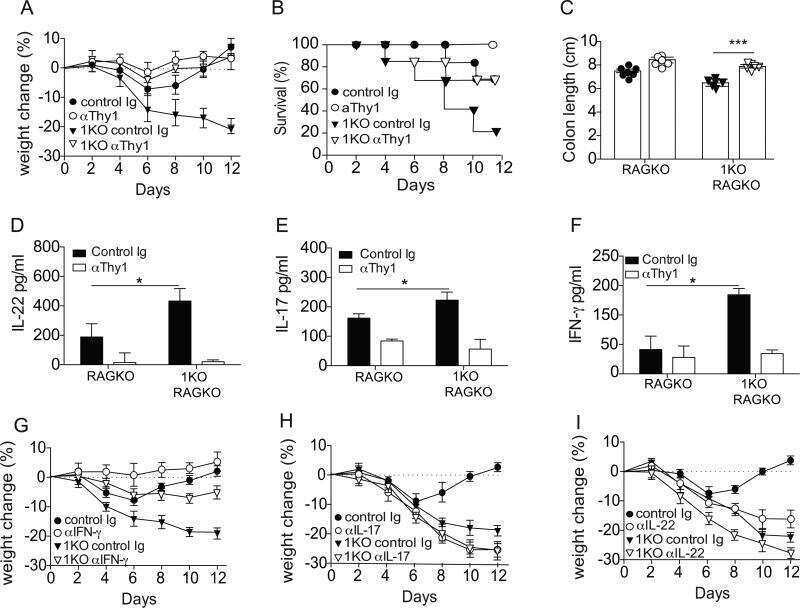
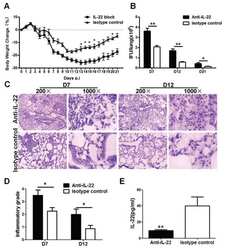
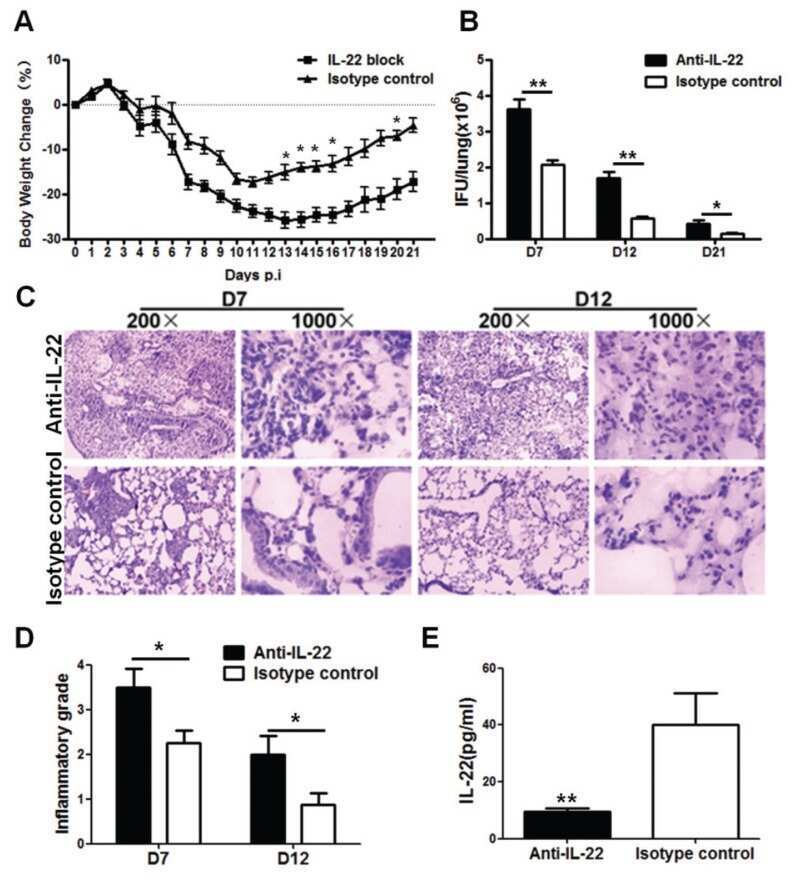
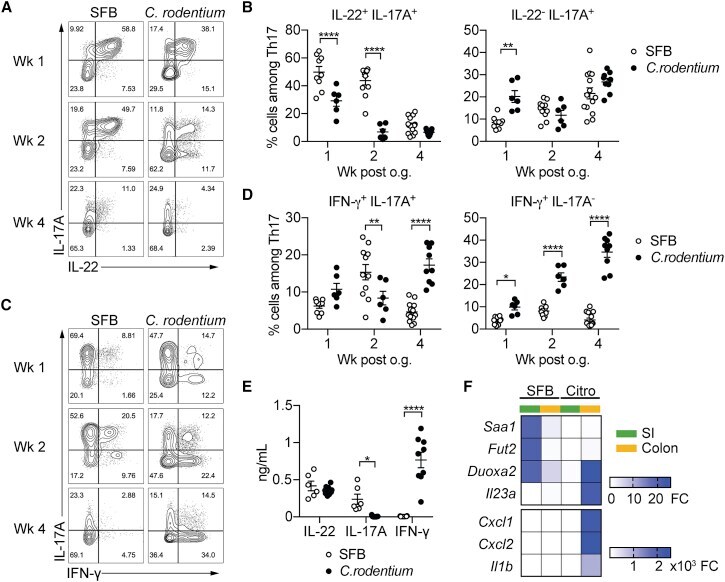
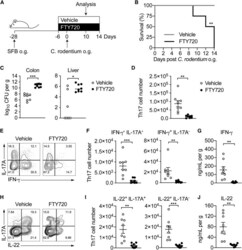
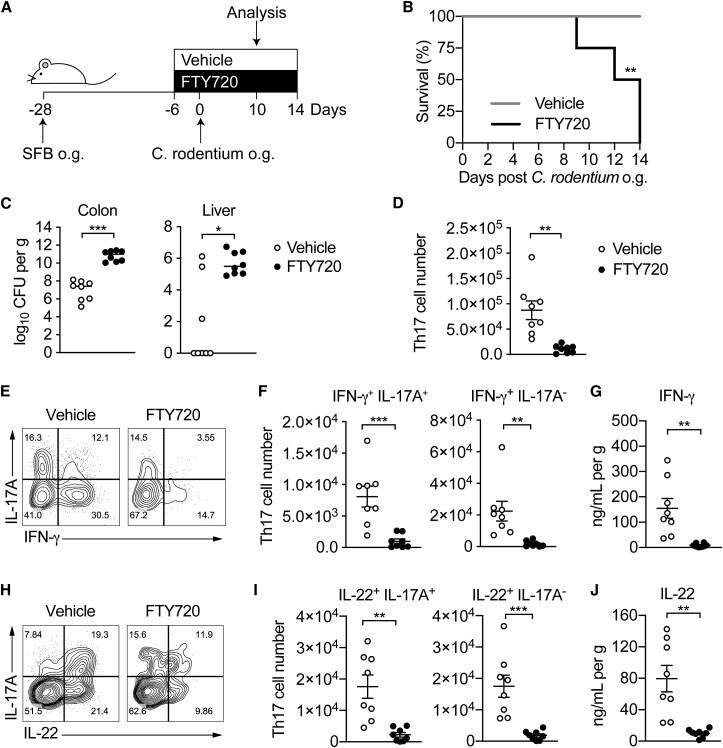
- Figure 4 Homeostatic Tissue-Resident Intestinal Th17 Cells Do Not Clear C. rodentium Infection (A) Schematic drawing of the experimental protocol. Mice were orally gavaged with SFB + feces 4 weeks before the infection with C. rodentium (at day 0). Six days before the infection with C. rodentium (day -6), mice were intra-peritoneally injected with FTY720 (3 mg/kg) or control vehicle every day. FTY720 and control vehicle administration continued after the infection every other day until the end of the experiment. (B) Survival curve of C. - rodentium -infected mice treated with vehicle or FTY720. (C) C. rodentium burdens in colon and liver. (D-J) Absolute numbers of colonic Th17 cells. Representative intracellular staining for IL-17A and IFN-gamma (E) or IL-22 (H) and quantification (F and I) in colonic Th17 cells. IFN-gamma (G) and IL-22 (J) protein content in colon explant cultures. Bars show median (C) or mean +- SEM (D, F, G, I, and J) and each symbol represents an individual mouse (n = 8). Results are representative of three independent experiments. * p < 0.05, ** p < 0.01, *** p < 0.001 by Mantel-Cox (B), Mann-Whitney (C) or Student''s t test (D, F, G, I, and J). See also Figure S4 .
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
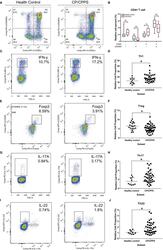
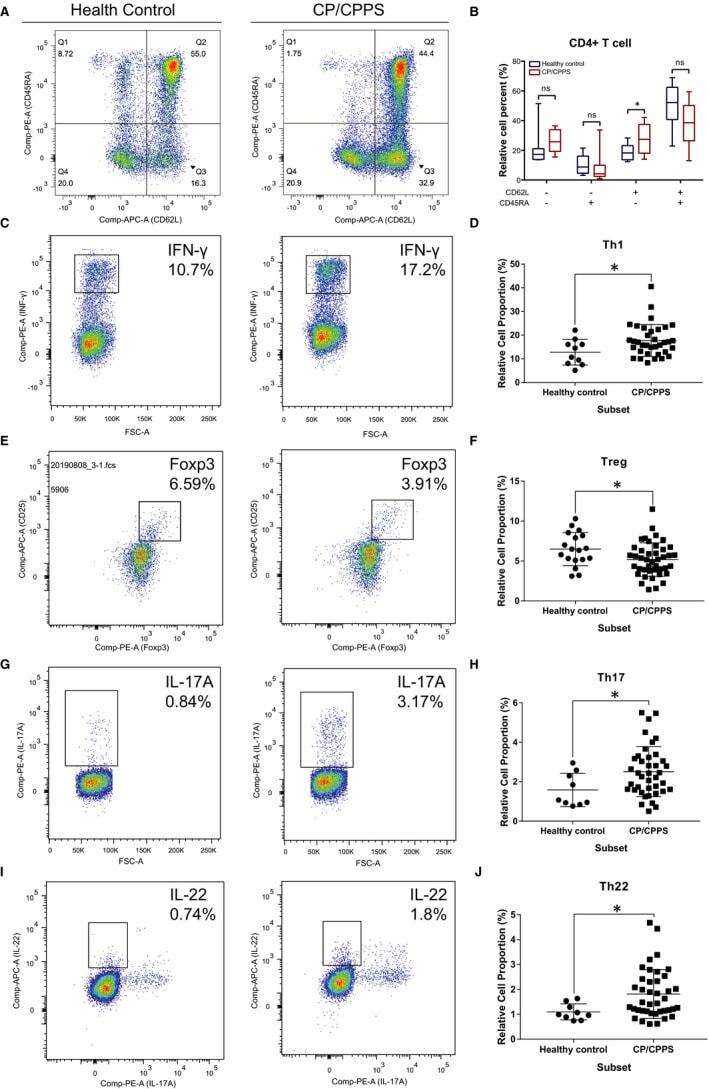
- FIGURE 5 Flow cytometry revealed central memory T cell, Th1, Th17, Th22 and Treg proportions increased in PBMCs derived from CP/CPPS patients than healthy controls. PBMCs were incubated with various fluorescein-labelled antigens for surface staining. PerCP/Cyanine5.5-conjugated CD3 and FITC-conjugated CD4 were used for the central memory T cell, Th1, Th17 and Th22 cell staining, and PE-conjugated CD25 and FITC-conjugated CD4 were incubated for staining Treg cells. After fixing and permeabilizing with cell fixation/permeabilization kit, (A) for samples staining central memory T cells were incubated with PE-conjugated CD45RA and APC-conjugated CD62L; (C) for samples staining, Th1 cells were incubated with PE-conjugated IFN-gamma; (E) for samples staining, Treg cells were incubated with eFluor 660-conjugated FoxP3; (G) for samples staining, Th17 cells were incubated with PE-conjugated IL-17A; and (I) for samples staining, Th22 cells were incubated with PE-conjugated IL-17A and APC-conjugated IL-22. The quantification data were presented in B, D, F, H and J. * P < 0.05; PBMC, peripheral blood mononuclear cell
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
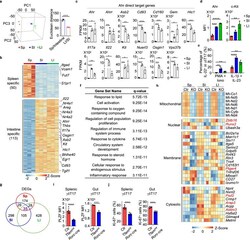
- Fig. 4 Gene profiling reveals tissue-specific transcriptomic signature of gammadeltaT17 cells. a Left panel, principal component analysis (PCA) plot of the RNA-seq of Tfam-sufficient splenic (Sp), small intestinal (SI), and large intestinal (LI) gammadeltaT17 cells from 3-week-old ( n = 3 for each group) mice. gammadeltaT17 cells of each sample were sorted from an individual mouse. Right panel, Euclidean distance comparison of the transcriptome of each splenic gammadeltaT17 cell sample and the mean intestine centroid vs. the mean small intestine centroid to each large intestinal gammadeltaT17 cell sample ( n = 3 for each group) (*** P = 0.0002). b Heatmap of tissue-specific genes (TSGs) identified by the RNA-seq in Sp, SI, and LI gammadeltaT17 cells. Genes were ranked in a descending order based on the fold changes of expression (Sp/[SI and LI average mean]; [SI and LI average mean]/Sp). c RNA-seq FPKM values of Ahr direct target genes of Sp, SI, and LI gammadeltaT17 cells (mean +- SD) ( n = 3 for each group) (Sp vs. SI: Ahr , *** P = 0.0005, Ahrr , ** P = 0.0037, Asb2 , *** P = 0.0004, Cd93 , **** P < 0.0001, Cd160 , *** P = 0.0002, Gem , ** P = 0.0096, Hic1 , *** P = 0.0002, Il17a , **** P < 0.0001, Il22 , * P = 0.0180, Kit , *** P = 0.0004, Nuerl3 , **** P < 0.0001, Osgin1 , *** P = 0.0002, Vps37b , **** P < 0.0001; Sp vs. LI: Ahr , * P = 0.0460, Ahrr , P = 0.0506, Asb2 , ** P = 0.0076, Cd93 , **** P < 0.0001, Cd160 , *** P = 0.0001, Gem , * P = 0.0108, Hic1 , *** P = 0.00
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
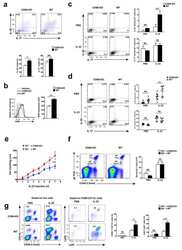
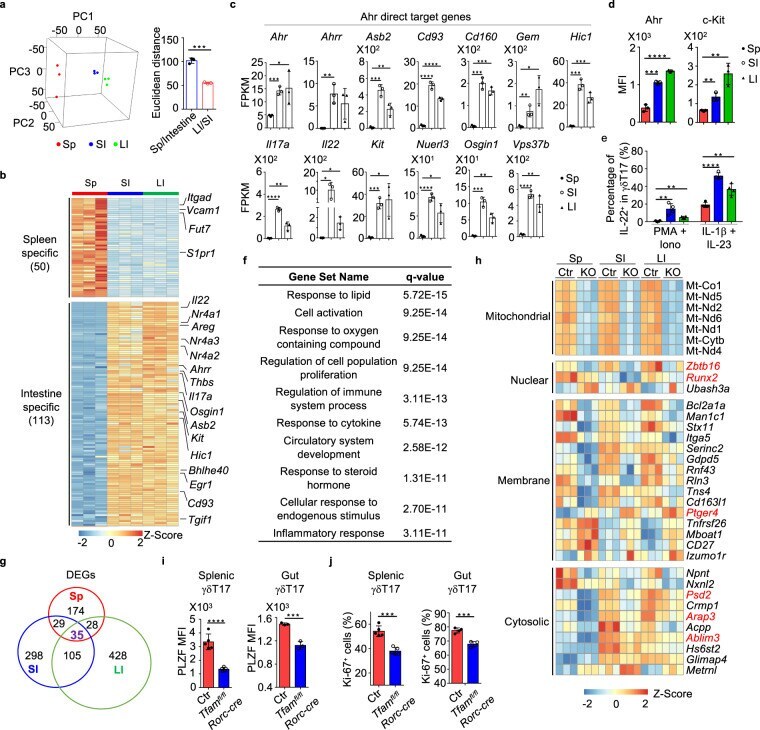
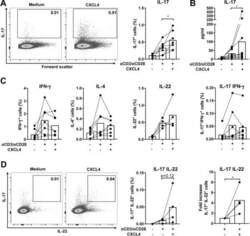
- Figure 1 CXCL4 increases the percentage of IL-17 producing cells in CD3/CD28-stimulated human CD4 + T cells. CD4 + T cells were isolated from healthy donors and cultured with CD3/CD28 coated Dynabeads and CXCL4 for five days. (A, B) The effect of CXCL4 on IL-17 production by CD4 + T cells was assessed by (A) flow cytometric intracellular cytokine staining and (B) enzyme-linked immunosorbent assay. (C) The percentage of of IFN-gamma-, IL-4- and IL-22-producing CD4 + T cells were measured by flow cytometry. (D, E) The amount of IL-17 producing cells co-expressing IFN-gamma (D) or IL-22 (E) were measured by flow cytometry. Cells were gated on live, single cells. Means (bars) and values from each donor are shown. Data are pooled from two to four independent experiments, except for panel B from 14 independent experiments, with one to four donor samples per experiment. Each dot on the bar graphs represent a single donor and paired t -test was used for statistical analysis. * p
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
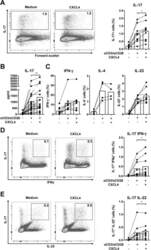
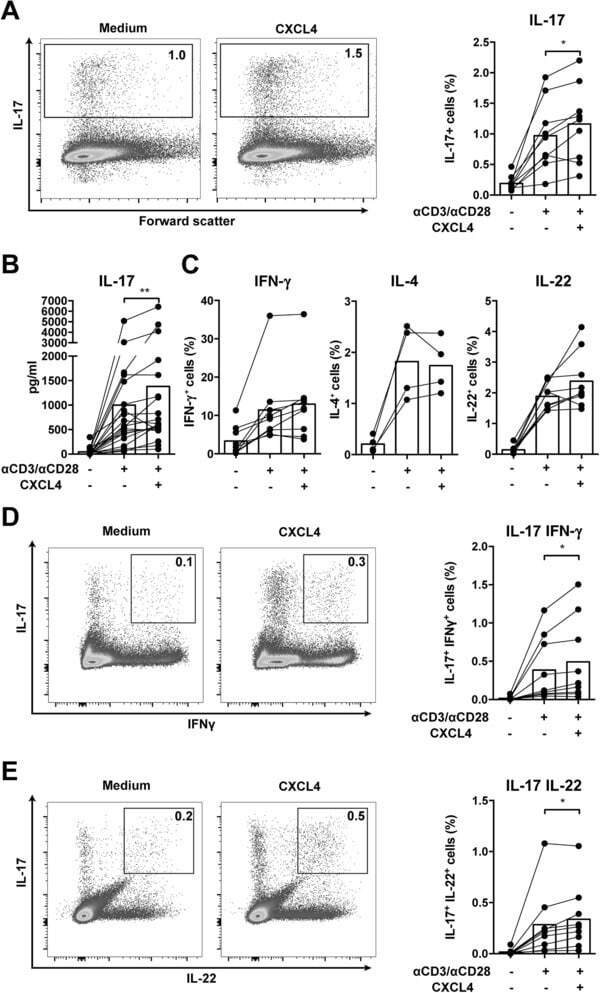
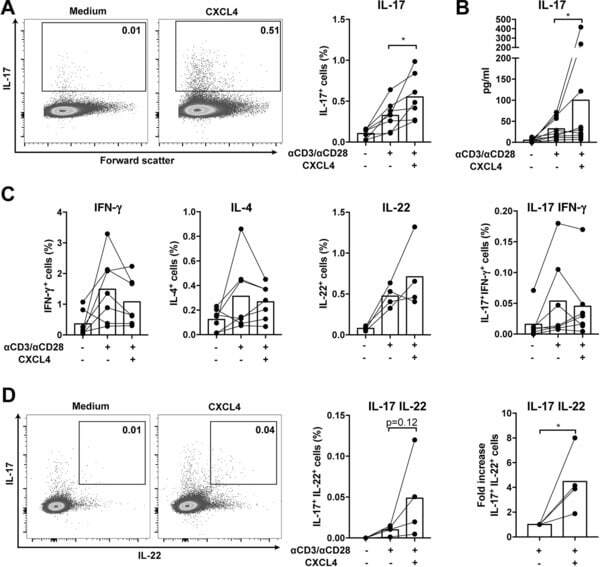
- Figure 2 CXCL4 induces IL-17 producing cells differentiated from naive human CD4 + T cells. Naive CD4 + T cells were purified by fluorescence-activated cell sorting and cultured with CD3/CD28 coated Dynabeads and CXCL4 for seven days. (A, B) The effect of CXCL4 on IL-17 production by CD4 + T cells was assessed by (A) flow cytometric intracellular cytokine staining and (B) enzyme-linked immunosorbent assay. (C) The levels of IFN-gamma-, IL-4-, IL-22-, and IL-17/IFN-gamma-producing CD4 + T cells were measured by flow cytometry. (D) The amount of IL-17 producing cells co-expressing or IL-22 was measured by flow cytometry. Cells were gated on live, single cells. Means (bars) and values from each donor are shown. Data are pooled from four to 10 independent experiments, with one to two donor samples per experiment. Each dot on the bar graphs represents a single donor and paired t -test was used for statistical analysis. * p
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
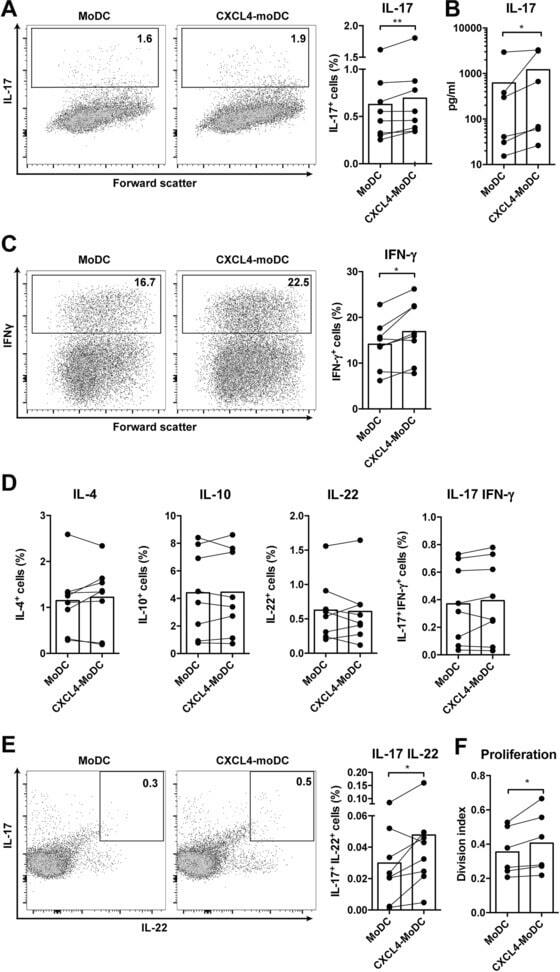
- Figure 4 CXCL4-differentiated monocyte-derived dendritic cells enhance pro-inflammatory cytokine production and proliferation by CD4 + T cells. Monocytes from healthy donors were isolated and differentiated into dendritic cells in the absence or presence of CXCL4 (moDCs or CXCL4-moDCs). moDCs were then co-cultured with autologous CD4 + T cells in the presence of superantigen from Staphylococcal Enterotoxin B (SEB) for three days and restimulated with PMA and ionomycin. (A-C) Comparison of co-culture with moDC or CXCL4-moDC on IL-17 or IFN-gamma production by CD4 + T cells was assessed by (A, C) intracellular cytokine staining and (B) enzyme-linked immunosorbent assay are shown. (D) Intracellular cytokine staining was performed for the measurement of IL-4 + , IL-10 + , IL-22 + , and IL-17 + IFN-gamma + cells gated on live CD4 + T cells. (E) The amount of IL-17 producing cells co-expressing IL-22 as measured by flow cytometry. (F) CD4 + T cells were labeled with CellTrace Violet prior co-culture and proliferation was analyzed as division index. Cells were gated on live, single, CD4 + T cells. Means (bars) and values from each donor are shown. Data are pooled from two to three independent experiments, with two to three donor samples per experiment. Each dot on the bar graphs represent a single donor and paired t -test was used for statistical analysis. * p
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
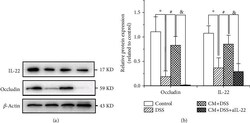
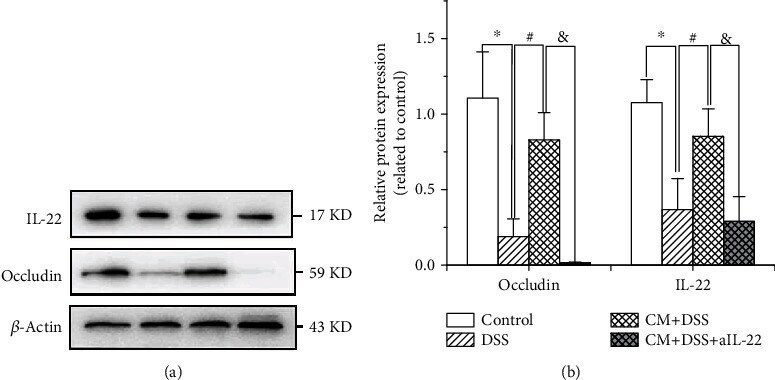
- Figure 3 Chlamydia muridarum upregulates IL-22 and occludin expression in mice with dextran sodium sulfate- (DSS-) induced colitis. (a) IL-22 and occludin expression in various groups assessed using western blotting. (b) Relative protein expression of IL-22 and occludin. ( n = 3 per group). * P < 0.05, DSS group compared with control group; # P < 0.05, CM+DSS group compared with DSS group; & P < 0.05, CM+DSS+aIL-22 group compared with CM+DSS group.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
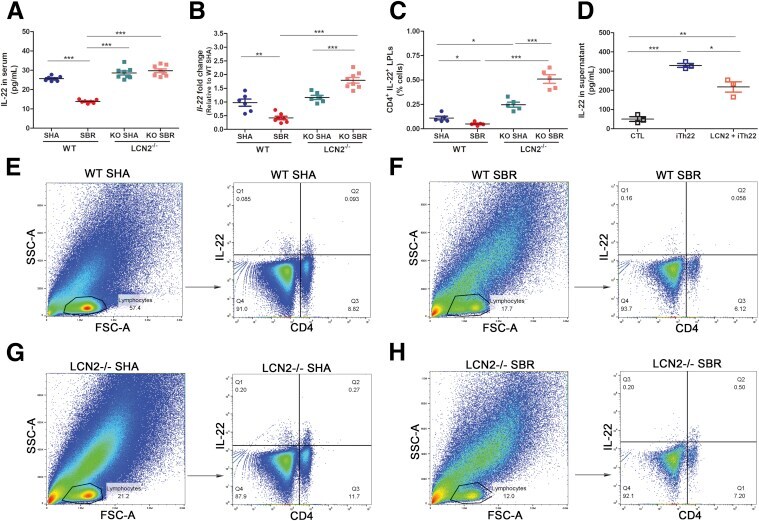
- Figure 9 LCN2 reduces intestinal adaptation by inhibiting IL22 expression after 75% SBR on POD 7 . ( A ) IL22 protein expression in the serum was measured by ELISA (n = 7 mice per group). ( B ) Transcriptional levels of the Il22 gene in the intestine were evaluated by quantitative PCR (n = 6 per group). ( C ) CD4 + IL22 + LPLs were quantified with flow cytometry analysis (n = 5 per group). ( D ) Representative IL22 protein expression in supernatant from Th22 cells in vitro with and without LCN2 stimulation as measured by ELISA (n = 3 per group/each experiment, each experiment was repeated 3 times). ( E-H ) Representative flow cytometry dot plots of the LPL population from small intestinal tissue. Single-cell suspensions prepared from the lamina propria were stained with CD4 and IL22 antibodies. Gate 1 identified lymphocytes based on FSC-A/SSC-A properties. The numbers within the quadrants represent the percentage of CD4 + IL22 - , CD4 + IL22 + , CD4 - IL22 - , and CD4 - IL22 + cells within the lymphocyte gate. Data are representative of analyses of 5 mice per group. Forward scatter area (FSC-A)/side scatter area (SSC-A). * P < .05, ** P < .01, and *** P < .001.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
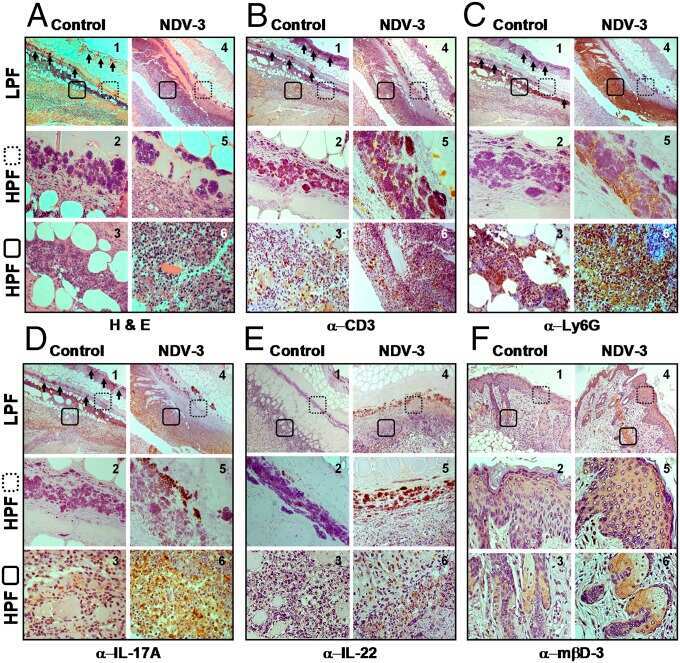
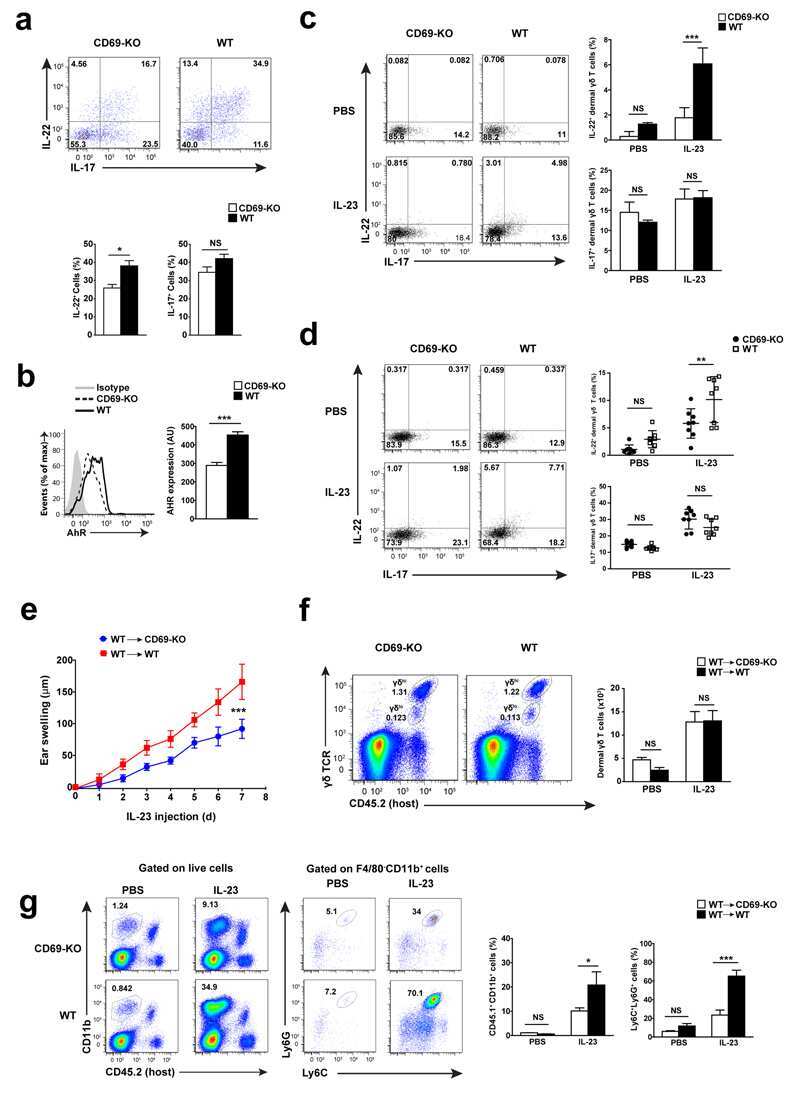
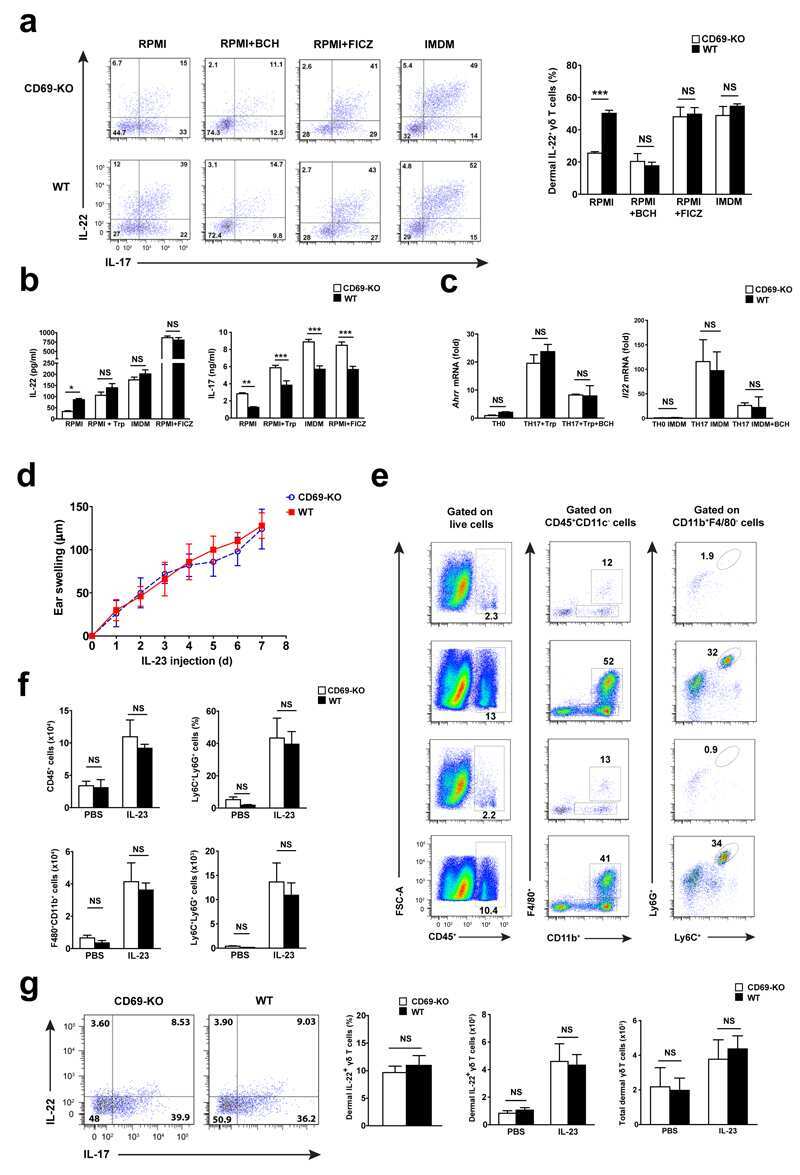
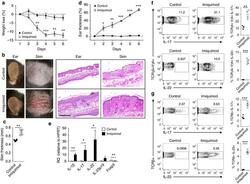
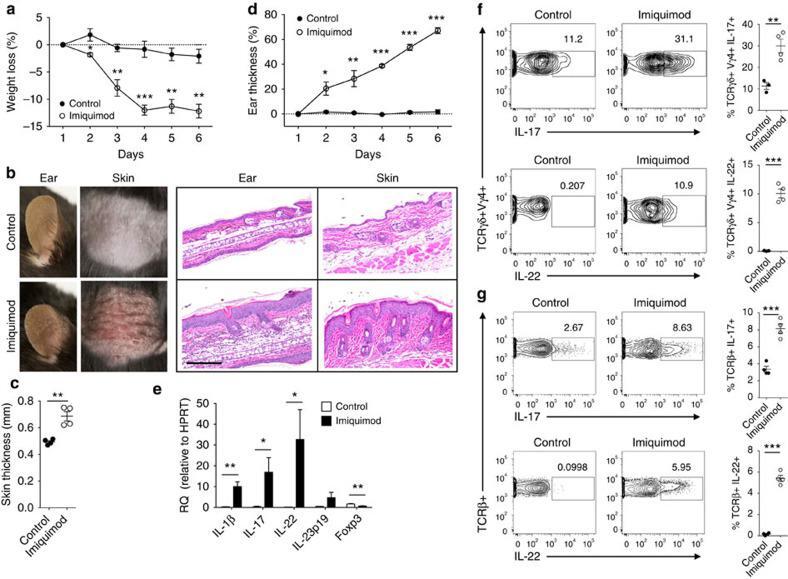
- Figure 1 Imiquimod increases IL-17 and IL-22 in both gammadelta and alphabeta T cells in skin. Skin and ear of wild type C57BL/6 mice were treated by topical application of imiquimod cream (Fougera, n =4) or control cream (Vaseline, n =3) for 6 consecutive days. ( a ) Weight loss of imiquimod and control cream treated mice monitored daily. ( b ) Photographs of imiquimod and control cream treated skin and ear; photos were taken at day 6. H&E stained ear and back skin sections of imiquimod treated and control mice. Scale bar, 200 mum. ( c ) Thickness of skin measured by Digimatic Caliper at day 6 in control and imiquimod treated mice. Data showed represents average of at least two measurements. ( d ) Ear thickness of imiquimod and control cream treated mice monitored daily. Ear thickness was measured using Digimatic Caliper. ( e ) Quantitative PCR analysis of Th17 associated cytokines and Foxp3 in skin after 6 days of imiquimod and control cream treatment. ( f ) Representative flow cytometric analysis of TCRgammadelta + Vgamma4 + IL-17 + (upper raw) and TCRgammadelta + Vgamma4 + IL-22 + cells (lower row) in the skin of control ( n =3) and imiquimod treated mice ( n =4). ( g ) Representative flow cytometric analysis of single-positive TCRbeta + IL-17 + (upper raw) or TCRbeta + IL-22 + (lower raw) cells in the skin of control ( n =3) and imiquimod treated mice ( n =4). Data representative of more than three experiments, results are shown as mean+-s.e.m., significance determined b
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
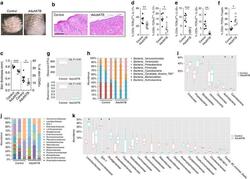
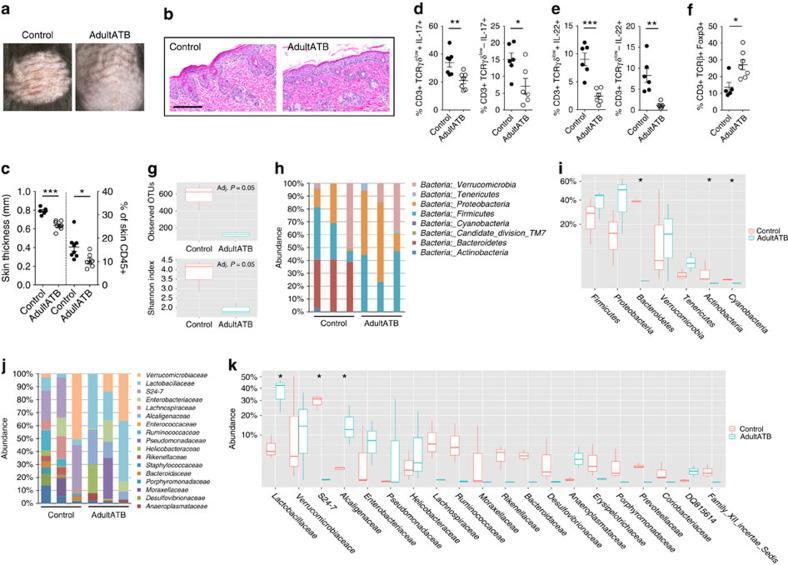
- Figure 2 Antibiotics in adult mice ameliorate psoriasis by reduction of IL-17 and IL-22 in T cells. Young adult mice (4-weeks-old) were exposed to antibiotics in drinking water (adultATB) for four weeks, and last six consecutive days followed by imiquimod treatment. ( a ) Representative photographs of back skin of control and adultATB mice followed by imiquimod treatment. Photos were taken at day 6. ( b ) H&E stained skin sections of control and adultATB mice after imiquimod treatment. Scale bar, 200 mum. ( c ) Thickness of skin measured by digimatic caliper at day 6 in control and imiquimod treated mice. Data showed represents average of at least two measurements. Increased skin infiltration in control compared adultATB mice based on staining of skin CD45 + cells. ( d ) Flow cytometric analysis of single-positive CD3 + TCRgammadelta low+ IL-17 + cells or CD3 + TCRgammadelta - IL-17 + cells in control ( n =6-7) and adultATB ( n =6-7) mice after imiquimod treatment. ( e ) Flow cytometric analysis of single-positive CD3 + TCRgammadelta low+ IL-22 + cells or CD3 + TCRgammadelta - IL-22 + cells in control ( n =6) and adultATB ( n =6) mice after imiquimod treatment. ( f ) Frequency of skin CD3 + TCRbeta + Foxp3 + cells in the skin of control ( n =5) and adultATB ( n =7) mice after imiquimod treatment. ( g ) Diversity of the microbiota (Observed OTUs, Shannon Index) in adultATB and control mice. Primary samples were collected per cage (Pooled samples from 3 independent experiments)
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
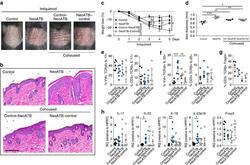
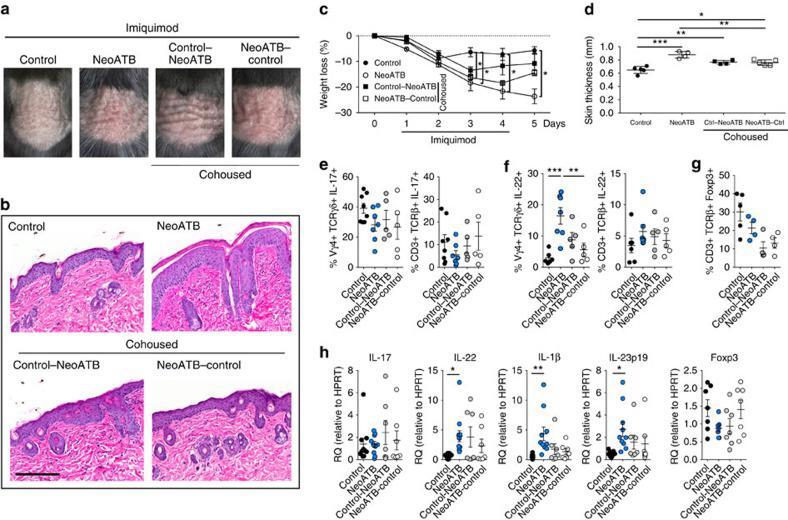
- Figure 3 Neonatal antibiotic treatment of mice increases severity of psoriasis. Breeding pairs were given antibiotics vancomycin+polymyxin B in drinking water immediately after pups delivery for 3 weeks of rearing (neoATB). After weaning, mice were housed or cohoused (mice without antibiotics treatment together with mice treated with antibiotics) in normal specific pathogen-free conditions for next 8 weeks. The mice were then treated with imiquimod cream for 4 consecutive days. ( a ) Representative photographs of back skin in controls (mice without antibiotic treatment); neonatally antibiotics treated mice (neoATB) or cohoused mice (control-neoATB resp. NeoATB-control) after imiquimod treatment. Photos were taken at day 5. ( b ) H&E stained skin sections of control, neoATB and cohoused mice after imiquimod treatment. Scale bar, 200 mum. ( c ) Weight loss of control, neoATB and cohoused mice during imiquimod treatment monitored daily. ( d ) Skin thickness of control, neoATB and cohoused mice measured at day 5. Skin thickness was measured by digimatic caliper by at least two investigators and data showed represents average of at least two measurements. ( e ) Flow cytometric analysis of single-positive TCRgammadelta + Vgamma4 + IL-17 + cells or CD3 + TCRbeta + IL-17 + cells in control ( n =8), neoATB ( n =7) and cohoused mice ( n =5) after imiquimod treatment. ( f ) Flow cytometric analysis of single-positive TCRgammadelta + Vgamma4 + IL-22 + cells or CD3 + TCRbeta + IL-22 + cel
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
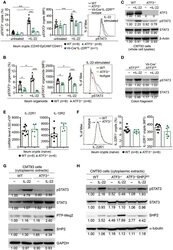
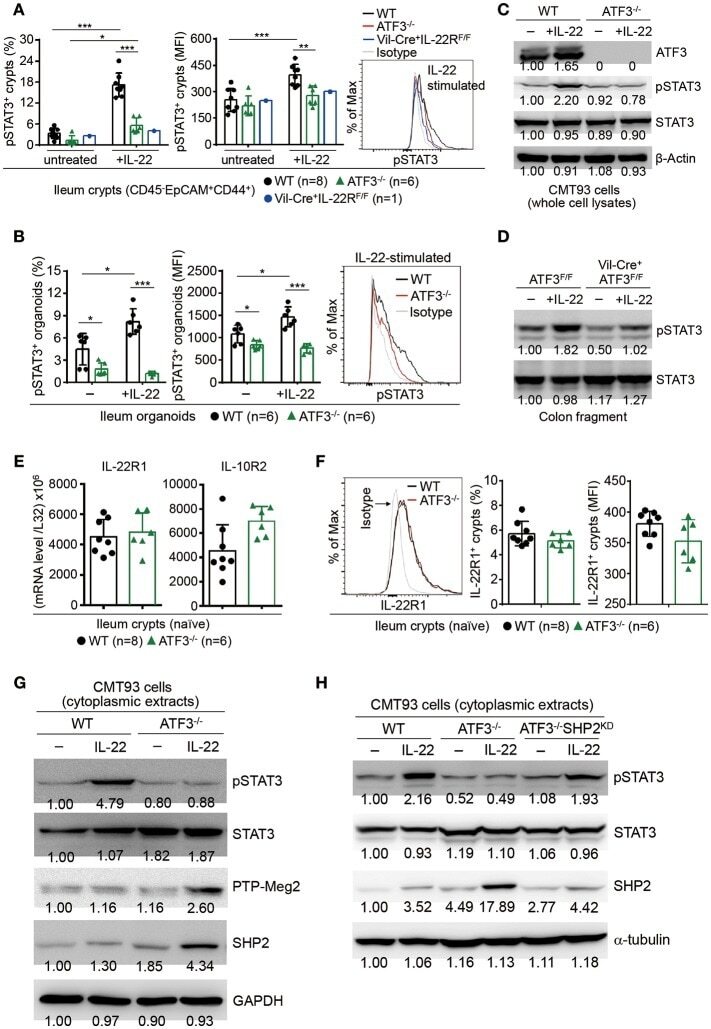
- Figure 7 ATF3 promotes IL-22-induced STAT3 phosphorylation by suppressing phosphatases. (A) Freshly isolated ileum crypts, or (B) ileum organoids at day 6 of culture, were stimulated with IL-22, followed by fixation and intracellular staining of phospho-STAT3, and analyzed by flow cytometry. Western blot analysis of (C) IL-22-stimulated CMT93 cells, or (D) IL-22-stimuated colon fragments isolated from the indicated mice, for the expression of the indicated proteins. (E) Quantitative real-time PCR analysis of IL-22R1 and IL-10R2 mRNA levels in freshly isolated ileum crypts from mice. (F) Flow cytometry analysis of IL-22R1 in freshly isolated ileum crypt cells gated on the CD45 - EpCAM + population. (G,H) Western blot analysis of unstimulated or IL-22-stimulated CMT93 cells for the indicated proteins. ATF3 -/- CMT93 cells with SHP2 knockdown (ATF3 -/- SHP2 KD ) were indicated. Images were representative of four independent experiments (G-H) . Results were from two independent experiments (A-F) . ""n"" refers to the number of mice analyzed (A,B,E,F) . Statistical analysis was done by multiple comparison in Two-way ANOVA test using Prism software. * P < 0.05, ** P < 0.005, *** P < 0.0005.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
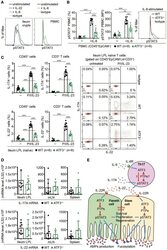
- Figure 8 ATF3 regulates IL-6-pSTAT3 signaling in intestinal Th17 cells. Flow cytometry analysis of phospho-STAT3 in (A) IL-6 or IL-22 stimulated freshly isolated ileum crypts or IL-6-stimulated peripheral blood mononuclear cell (PBMC) from wild-type mice, or in (B) IL-6-stimulated PBMC from wild-type and ATF3-deficient mice. (C) Flow cytometry analysis of intracellular IL-17A and IL-22 expression in naive lamina propria T cells from the indicated mice. Cells were treated with PMA, ionomycin and IL-23 in the presence of BFA for 4 h before analysis and gated on live CD45 + EpCAM - Lin - CD3 + population as shown. (D) Quantitative real-time PCR analysis of IL-17A and IL-22 mRNA levels in freshly isolated lamina propria (LPL) cells, mesenteric lymph nodes (mLN), or splenocytes. (E) Model of ATF3-mediated mucosal immunity via cross-regulation between IL-22-pSTAT3 signaling in epithelium (associated with AMP production and epithelial fucosylation) and IL-6-pSTAT3 signaling in Th17 cells (associated with signature IL-17A and IL-22 production). ""n"" refers to the number of mice analyzed. Statistical analysis was done by multiple comparison in Two-way ANOVA test using Prism software. * P < 0.05, ** P < 0.005, *** P < 0.0005.