Explore
Explore Validate
Validate Learn
Learn Western blot
Western blot Immunoprecipitation
ImmunoprecipitationAntibody data
- Antibody Data
- Antigen structure
- References [50]
- Comments [0]
- Validations
- Western blot [1]
- Immunocytochemistry [2]
- Immunohistochemistry [1]
- Other assay [15]
Submit
Validation data
Reference
Comment
Report error
- Product number
- MA1-411 - Provider product page

- Provider
- Invitrogen Antibodies
- Product name
- Progesterone Receptor Monoclonal Antibody (Alpha PR6)
- Antibody type
- Monoclonal
- Antigen
- Purifed from natural sources
- Description
- MA1-411 reacts with high affinity to the B form of the progesterone receptor from human, mouse, rat, rabbit, bovine, ovine, chicken, and guinea pig tissues. This antibody does not cross-react with estrogen receptor or glucocorticoid receptor. MA1-411 has been successfully used in Western blot, immunocytochemical, immunohistochemical and immunoprecipitation procedures. By Western blot, this antibody detects a 110 kDa protein representing the B form of the PR in chickens." The MA1-411 immunogen is progesterone receptor purified from chick oviduct cytosol. Reconstitute with PBS.
- Reactivity
- Human, Mouse, Rat, Bovine, Chicken/Avian, Guinea Pig, Rabbit
- Host
- Mouse
- Isotype
- IgG
- Antibody clone number
- Alpha PR6
- Vial size
- 100 μg
- Concentration
- 1 mg/mL
- Storage
- -20°C, Avoid Freeze/Thaw Cycles
Submitted references Macrophages in bovine term placenta: An ultrastructural and molecular study.
Pre-implantation exogenous progesterone and pregnancy in sheep: I. polyamines, nutrient transport, and progestamedins.
Levels of sex steroid hormones and their receptors in women with preeclampsia.
NF1 deficiency correlates with estrogen receptor signaling and diminished survival in breast cancer.
Lack of cortistatin or somatostatin differentially influences DMBA-induced mammary gland tumorigenesis in mice in an obesity-dependent mode.
Progesterone and Wnt4 control mammary stem cells via myoepithelial crosstalk.
No direct nuclear effect of progesterone in the inner ear: other possible pathways.
Endometrial population of oestrogen receptors alpha and beta and progesterone receptors A and B during the different phases of the follicular wave of llamas (Lama glama).
Prostaglandin treatment is associated with a withdrawal of progesterone and androgen at the receptor level in the uterine cervix.
Conditional deletion of the Lkb1 gene in the mouse mammary gland induces tumour formation.
Developmental programming: prenatal androgen excess disrupts ovarian steroid receptor balance.
A composite intronic element directs dynamic binding of the progesterone receptor and GATA-2.
Steroid receptor expression and morphology in provoked vestibulodynia.
Effects of testosterone and estrogen treatment on the distribution of sex hormone receptors in the endometrium of postmenopausal women.
Effects of tibolone and conventional HRT on the expression of estrogen and progesterone receptors in the breast.
Pelvic floor sex steroid hormone receptors, distribution and expression in pre- and postmenopausal stress urinary incontinent women.
Steroid receptor expression in the vulvar vestibular mucosa--effects of oral contraceptives and menstrual cycle.
A mouse model of basal-like breast carcinoma with metaplastic elements.
Tissue- and hormone-dependent progesterone receptor distribution in the rat uterus.
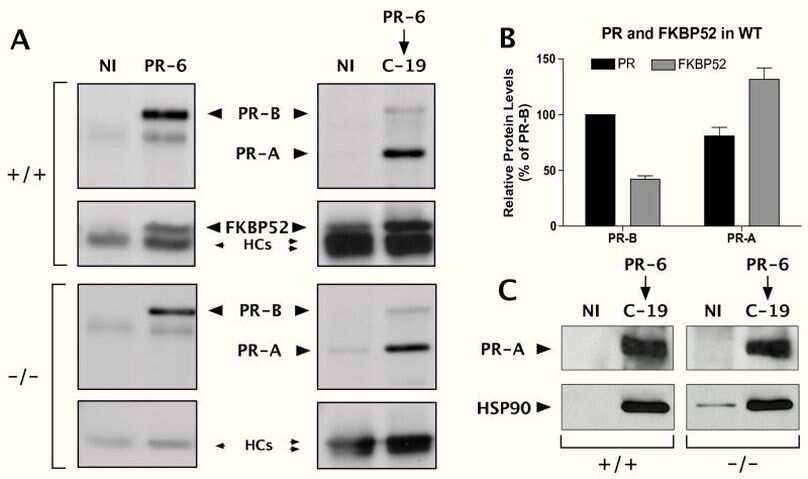
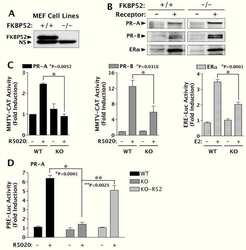
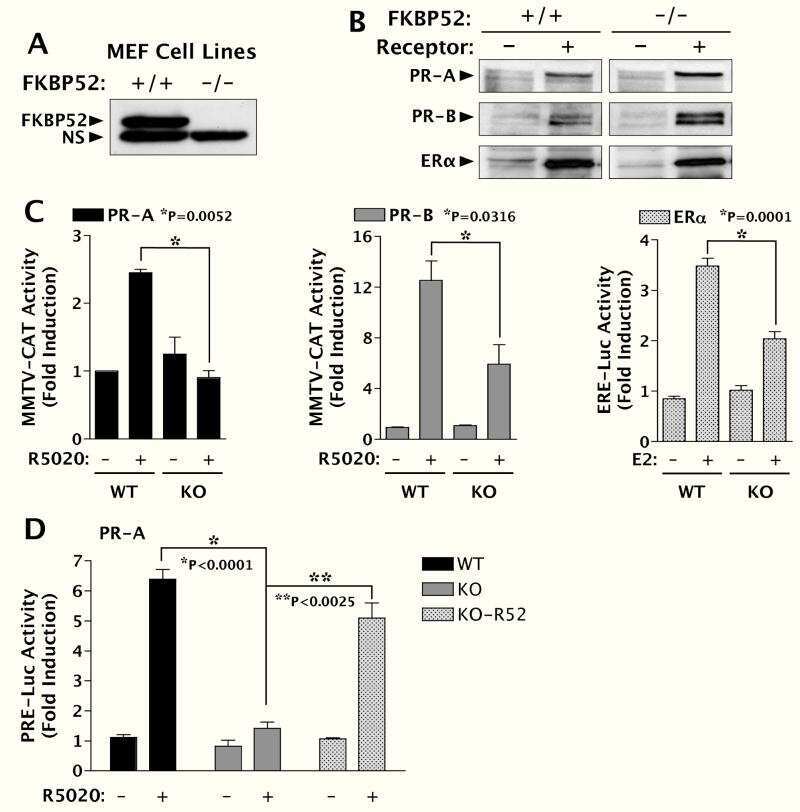
FK506-binding protein 52 is essential to uterine reproductive physiology controlled by the progesterone receptor A isoform.
Oestrogen receptor alpha and beta, androgen receptor and progesterone receptor mRNA and protein localisation within the developing ovary and in small growing follicles of sheep.
Characterization of ovarian surface epithelial cells from the hen: a unique model for ovarian cancer.
Overlapping and distinct expression of progesterone receptors A and B in mouse uterus and mammary gland during the estrous cycle.
Progesterone regulation of preimplantation conceptus growth and galectin 15 (LGALS15) in the ovine uterus.
Progesterone receptor deficient in chromatin binding has an altered cellular state.
Development and characterization of an endometrial tissue culture system.
Development and characterization of an endometrial tissue culture system.
Effects of long-term HRT and tamoxifen on the expression of progesterone receptors A and B in breast tissue from surgically postmenopausal cynomolgus macaques.
Postpartum uterine involution in sheep: histoarchitecture and changes in endometrial gene expression.
Effect of long-term treatment with steroid hormones or tamoxifen on the progesterone receptor and androgen receptor in the endometrium of ovariectomized cynomolgus macaques.
Myometrial effects of selective estrogen receptor modulators on estradiol-responsive gene expression are gene and cell-specific.
Endometrial effects of selective estrogen receptor modulators (SERMs) on estradiol-responsive gene expression are gene and cell-specific.
Expression of progesterone receptors A and B and insulin-like growth factor-I in human myometrium and fibroids after treatment with a gonadotropin-releasing hormone analogue.
Expression of progesterone receptors A and B and insulin-like growth factor-I in human myometrium and fibroids after treatment with a gonadotropin-releasing hormone analogue.
Formation of pinopodes in human endometrium is associated with the concentrations of progesterone and progesterone receptors.
Formation of pinopodes in human endometrium is associated with the concentrations of progesterone and progesterone receptors.
Neonatal ovine uterine development involves alterations in expression of receptors for estrogen, progesterone, and prolactin.
Progesterone modulation of osteopontin gene expression in the ovine uterus.
Interferon-tau and progesterone regulate ubiquitin cross-reactive protein expression in the ovine uterus.
Ovine uterine gland knock-out model: effects of gland ablation on the estrous cycle.
Ovine uterine gland knock-out model: effects of gland ablation on the estrous cycle.
Effects of recombinant ovine interferon tau, placental lactogen, and growth hormone on the ovine uterus.
Estradiol up-regulates estrogen receptor and progesterone receptor gene expression in specific ovine uterine cells.
Localization of progesterone receptors in pre- and postovulatory follicles of the domestic hen.
Progesterone-induced avidin as a marker of cytodifferentiation in the oviduct: comparison to ovalbumin.
Progesterone receptor concentration differences in the chick oviduct cells and apparent down-regulation by ligand. A semiquantitative immunohistochemical study.
Immunocytochemical study of progesterone receptors in hyperplastic and neoplastic endometrial tissues.
Interaction of murine progesterone receptors with specific monoclonal antibodies to the avian progesterone receptor.
Immunocytochemical study of progesterone receptors in the human endometrium during the menstrual cycle.
Immunologic analysis of human breast cancer progesterone receptors. 1. Immunoaffinity purification of transformed receptors and production of monoclonal antibodies.
Hooshmandabbasi R, Kazemian A, Zerbe H, Kowalewski MP, Klisch K
Reproduction in domestic animals = Zuchthygiene 2021 Sep;56(9):1243-1253
Reproduction in domestic animals = Zuchthygiene 2021 Sep;56(9):1243-1253
Pre-implantation exogenous progesterone and pregnancy in sheep: I. polyamines, nutrient transport, and progestamedins.
Hoskins EC, Halloran KM, Stenhouse C, Moses RM, Dunlap KA, Satterfield MC, Seo H, Johnson GA, Wu G, Bazer FW
Journal of animal science and biotechnology 2021 Mar 5;12(1):39
Journal of animal science and biotechnology 2021 Mar 5;12(1):39
Levels of sex steroid hormones and their receptors in women with preeclampsia.
Lan KC, Lai YJ, Cheng HH, Tsai NC, Su YT, Tsai CC, Hsu TY
Reproductive biology and endocrinology : RB&E 2020 Feb 18;18(1):12
Reproductive biology and endocrinology : RB&E 2020 Feb 18;18(1):12
NF1 deficiency correlates with estrogen receptor signaling and diminished survival in breast cancer.
Dischinger PS, Tovar EA, Essenburg CJ, Madaj ZB, Gardner EE, Callaghan ME, Turner AN, Challa AK, Kempston T, Eagleson B, Kesterson RA, Bronson RT, Bowman MJ, Graveel CR, Steensma MR
NPJ breast cancer 2018;4:29
NPJ breast cancer 2018;4:29
Lack of cortistatin or somatostatin differentially influences DMBA-induced mammary gland tumorigenesis in mice in an obesity-dependent mode.
Luque RM, Villa-Osaba A, L-López F, Pozo-Salas AI, Sánchez-Sánchez R, Ortega-Salas R, de Lecea L, Álvarez-Benito M, López-Miranda J, Gahete MD, Castaño JP
Breast cancer research : BCR 2016 Mar 8;18(1):29
Breast cancer research : BCR 2016 Mar 8;18(1):29
Progesterone and Wnt4 control mammary stem cells via myoepithelial crosstalk.
Rajaram RD, Buric D, Caikovski M, Ayyanan A, Rougemont J, Shan J, Vainio SJ, Yalcin-Ozuysal O, Brisken C
The EMBO journal 2015 Mar 4;34(5):641-52
The EMBO journal 2015 Mar 4;34(5):641-52
No direct nuclear effect of progesterone in the inner ear: other possible pathways.
Bonnard A, Sahlin L, Hultcrantz M, Simonoska R
Acta oto-laryngologica 2013 Dec;133(12):1250-7
Acta oto-laryngologica 2013 Dec;133(12):1250-7
Endometrial population of oestrogen receptors alpha and beta and progesterone receptors A and B during the different phases of the follicular wave of llamas (Lama glama).
Bianchi C, Sahlin L, Meikle A, Masironi B, Cavilla M, Aba M
Reproduction in domestic animals = Zuchthygiene 2010 Oct;45(5):872-80
Reproduction in domestic animals = Zuchthygiene 2010 Oct;45(5):872-80
Prostaglandin treatment is associated with a withdrawal of progesterone and androgen at the receptor level in the uterine cervix.
Vladic-Stjernholm Y, Vladic T, Blesson CS, Ekman-Ordeberg G, Sahlin L
Reproductive biology and endocrinology : RB&E 2009 Oct 23;7:116
Reproductive biology and endocrinology : RB&E 2009 Oct 23;7:116
Conditional deletion of the Lkb1 gene in the mouse mammary gland induces tumour formation.
McCarthy A, Lord CJ, Savage K, Grigoriadis A, Smith DP, Weigelt B, Reis-Filho JS, Ashworth A
The Journal of pathology 2009 Nov;219(3):306-16
The Journal of pathology 2009 Nov;219(3):306-16
Developmental programming: prenatal androgen excess disrupts ovarian steroid receptor balance.
Ortega HH, Salvetti NR, Padmanabhan V
Reproduction (Cambridge, England) 2009 May;137(5):865-77
Reproduction (Cambridge, England) 2009 May;137(5):865-77
A composite intronic element directs dynamic binding of the progesterone receptor and GATA-2.
Magklara A, Smith CL
Molecular endocrinology (Baltimore, Md.) 2009 Jan;23(1):61-73
Molecular endocrinology (Baltimore, Md.) 2009 Jan;23(1):61-73
Steroid receptor expression and morphology in provoked vestibulodynia.
Johannesson U, Sahlin L, Masironi B, Hilliges M, Blomgren B, Rylander E, Bohm-Starke N
American journal of obstetrics and gynecology 2008 Mar;198(3):311.e1-6
American journal of obstetrics and gynecology 2008 Mar;198(3):311.e1-6
Effects of testosterone and estrogen treatment on the distribution of sex hormone receptors in the endometrium of postmenopausal women.
Zang H, Sahlin L, Masironi B, Hirschberg AL
Menopause (New York, N.Y.) 2008 Mar-Apr;15(2):233-9
Menopause (New York, N.Y.) 2008 Mar-Apr;15(2):233-9
Effects of tibolone and conventional HRT on the expression of estrogen and progesterone receptors in the breast.
Ma L, Hofling M, Masironi B, von Schoultz B, Cline JM, Sahlin L
Maturitas 2008 Dec 20;61(4):345-9
Maturitas 2008 Dec 20;61(4):345-9
Pelvic floor sex steroid hormone receptors, distribution and expression in pre- and postmenopausal stress urinary incontinent women.
Söderberg MW, Johansson B, Masironi B, Byström B, Falconer C, Sahlin L, Ordeberg GE
Acta obstetricia et gynecologica Scandinavica 2007;86(11):1377-84
Acta obstetricia et gynecologica Scandinavica 2007;86(11):1377-84
Steroid receptor expression in the vulvar vestibular mucosa--effects of oral contraceptives and menstrual cycle.
Johannesson U, Sahlin L, Masironi B, Rylander E, Bohm-Starke N
Contraception 2007 Oct;76(4):319-25
Contraception 2007 Oct;76(4):319-25
A mouse model of basal-like breast carcinoma with metaplastic elements.
McCarthy A, Savage K, Gabriel A, Naceur C, Reis-Filho JS, Ashworth A
The Journal of pathology 2007 Mar;211(4):389-98
The Journal of pathology 2007 Mar;211(4):389-98
Tissue- and hormone-dependent progesterone receptor distribution in the rat uterus.
Sahlin L, Masironi B, Akerberg S, Eriksson H
Reproductive biology and endocrinology : RB&E 2006 Sep 11;4:47
Reproductive biology and endocrinology : RB&E 2006 Sep 11;4:47
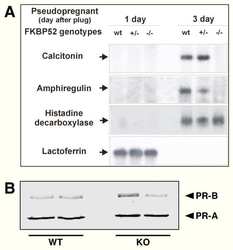
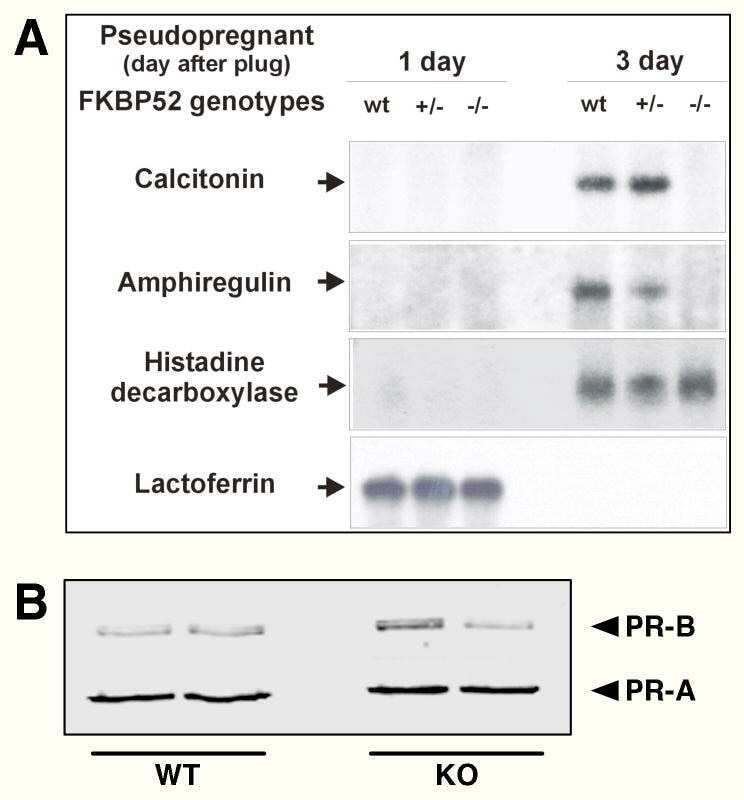
FK506-binding protein 52 is essential to uterine reproductive physiology controlled by the progesterone receptor A isoform.
Yang Z, Wolf IM, Chen H, Periyasamy S, Chen Z, Yong W, Shi S, Zhao W, Xu J, Srivastava A, Sánchez ER, Shou W
Molecular endocrinology (Baltimore, Md.) 2006 Nov;20(11):2682-94
Molecular endocrinology (Baltimore, Md.) 2006 Nov;20(11):2682-94
Oestrogen receptor alpha and beta, androgen receptor and progesterone receptor mRNA and protein localisation within the developing ovary and in small growing follicles of sheep.
Juengel JL, Heath DA, Quirke LD, McNatty KP
Reproduction (Cambridge, England) 2006 Jan;131(1):81-92
Reproduction (Cambridge, England) 2006 Jan;131(1):81-92
Characterization of ovarian surface epithelial cells from the hen: a unique model for ovarian cancer.
Giles JR, Olson LM, Johnson PA
Experimental biology and medicine (Maywood, N.J.) 2006 Dec;231(11):1718-25
Experimental biology and medicine (Maywood, N.J.) 2006 Dec;231(11):1718-25
Overlapping and distinct expression of progesterone receptors A and B in mouse uterus and mammary gland during the estrous cycle.
Mote PA, Arnett-Mansfield RL, Gava N, deFazio A, Mulac-Jericevic B, Conneely OM, Clarke CL
Endocrinology 2006 Dec;147(12):5503-12
Endocrinology 2006 Dec;147(12):5503-12
Progesterone regulation of preimplantation conceptus growth and galectin 15 (LGALS15) in the ovine uterus.
Satterfield MC, Bazer FW, Spencer TE
Biology of reproduction 2006 Aug;75(2):289-96
Biology of reproduction 2006 Aug;75(2):289-96
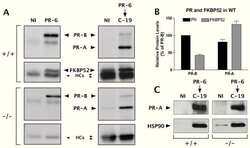
Progesterone receptor deficient in chromatin binding has an altered cellular state.
Botos J, Xian W, Smith DF, Smith CL
The Journal of biological chemistry 2004 Apr 9;279(15):15231-9
The Journal of biological chemistry 2004 Apr 9;279(15):15231-9
Development and characterization of an endometrial tissue culture system.
Stavreus-Evers A, Hovatta O, Eriksson H, Landgren BM
Reproductive biomedicine online 2003 Sep;7(2):243-9
Reproductive biomedicine online 2003 Sep;7(2):243-9
Development and characterization of an endometrial tissue culture system.
Stavreus-Evers A, Hovatta O, Eriksson H, Landgren BM
Reproductive biomedicine online 2003 Sep;7(2):243-9
Reproductive biomedicine online 2003 Sep;7(2):243-9
Effects of long-term HRT and tamoxifen on the expression of progesterone receptors A and B in breast tissue from surgically postmenopausal cynomolgus macaques.
Isaksson E, Wang H, Sahlin L, von Schoultz B, Cline JM, von Schoultz E
Breast cancer research and treatment 2003 May;79(2):233-9
Breast cancer research and treatment 2003 May;79(2):233-9
Postpartum uterine involution in sheep: histoarchitecture and changes in endometrial gene expression.
Gray CA, Stewart MD, Johnson GA, Spencer TE
Reproduction (Cambridge, England) 2003 Feb;125(2):185-98
Reproduction (Cambridge, England) 2003 Feb;125(2):185-98
Effect of long-term treatment with steroid hormones or tamoxifen on the progesterone receptor and androgen receptor in the endometrium of ovariectomized cynomolgus macaques.
Wang H, Isaksson E, von Schoultz B, Cline JM, Sahlin L
Reproductive biology and endocrinology : RB&E 2003 Feb 5;1:7
Reproductive biology and endocrinology : RB&E 2003 Feb 5;1:7
Myometrial effects of selective estrogen receptor modulators on estradiol-responsive gene expression are gene and cell-specific.
Farnell YZ, Ing NH
The Journal of steroid biochemistry and molecular biology 2003 Apr;84(5):527-36
The Journal of steroid biochemistry and molecular biology 2003 Apr;84(5):527-36
Endometrial effects of selective estrogen receptor modulators (SERMs) on estradiol-responsive gene expression are gene and cell-specific.
Farnell YZ, Ing NH
The Journal of steroid biochemistry and molecular biology 2003 Apr;84(5):513-26
The Journal of steroid biochemistry and molecular biology 2003 Apr;84(5):513-26
Expression of progesterone receptors A and B and insulin-like growth factor-I in human myometrium and fibroids after treatment with a gonadotropin-releasing hormone analogue.
Wu X, Wang H, Englund K, Blanck A, Lindblom B, Sahlin L
Fertility and sterility 2002 Nov;78(5):985-93
Fertility and sterility 2002 Nov;78(5):985-93
Expression of progesterone receptors A and B and insulin-like growth factor-I in human myometrium and fibroids after treatment with a gonadotropin-releasing hormone analogue.
Wu X, Wang H, Englund K, Blanck A, Lindblom B, Sahlin L
Fertility and sterility 2002 Nov;78(5):985-93
Fertility and sterility 2002 Nov;78(5):985-93
Formation of pinopodes in human endometrium is associated with the concentrations of progesterone and progesterone receptors.
Stavreus-Evers A, Nikas G, Sahlin L, Eriksson H, Landgren BM
Fertility and sterility 2001 Oct;76(4):782-91
Fertility and sterility 2001 Oct;76(4):782-91
Formation of pinopodes in human endometrium is associated with the concentrations of progesterone and progesterone receptors.
Stavreus-Evers A, Nikas G, Sahlin L, Eriksson H, Landgren BM
Fertility and sterility 2001 Oct;76(4):782-91
Fertility and sterility 2001 Oct;76(4):782-91
Neonatal ovine uterine development involves alterations in expression of receptors for estrogen, progesterone, and prolactin.
Taylor KM, Gray CA, Joyce MM, Stewart MD, Bazer FW, Spencer TE
Biology of reproduction 2000 Oct;63(4):1192-204
Biology of reproduction 2000 Oct;63(4):1192-204
Progesterone modulation of osteopontin gene expression in the ovine uterus.
Johnson GA, Spencer TE, Burghardt RC, Taylor KM, Gray CA, Bazer FW
Biology of reproduction 2000 May;62(5):1315-21
Biology of reproduction 2000 May;62(5):1315-21
Interferon-tau and progesterone regulate ubiquitin cross-reactive protein expression in the ovine uterus.
Johnson GA, Spencer TE, Burghardt RC, Joyce MM, Bazer FW
Biology of reproduction 2000 Mar;62(3):622-7
Biology of reproduction 2000 Mar;62(3):622-7
Ovine uterine gland knock-out model: effects of gland ablation on the estrous cycle.
Allison Gray C, Bartol FF, Taylor KM, Wiley AA, Ramsey WS, Ott TL, Bazer FW, Spencer TE
Biology of reproduction 2000 Feb;62(2):448-56
Biology of reproduction 2000 Feb;62(2):448-56
Ovine uterine gland knock-out model: effects of gland ablation on the estrous cycle.
Allison Gray C, Bartol FF, Taylor KM, Wiley AA, Ramsey WS, Ott TL, Bazer FW, Spencer TE
Biology of reproduction 2000 Feb;62(2):448-56
Biology of reproduction 2000 Feb;62(2):448-56
Effects of recombinant ovine interferon tau, placental lactogen, and growth hormone on the ovine uterus.
Spencer TE, Gray A, Johnson GA, Taylor KM, Gertler A, Gootwine E, Ott TL, Bazer FW
Biology of reproduction 1999 Dec;61(6):1409-18
Biology of reproduction 1999 Dec;61(6):1409-18
Estradiol up-regulates estrogen receptor and progesterone receptor gene expression in specific ovine uterine cells.
Ing NH, Tornesi MB
Biology of reproduction 1997 May;56(5):1205-15
Biology of reproduction 1997 May;56(5):1205-15
Localization of progesterone receptors in pre- and postovulatory follicles of the domestic hen.
Yoshimura Y, Bahr JM
Endocrinology 1991 Jan;128(1):323-30
Endocrinology 1991 Jan;128(1):323-30
Progesterone-induced avidin as a marker of cytodifferentiation in the oviduct: comparison to ovalbumin.
Joensuu TK, Ylikomi TJ, Toft DO, Keinanen RA, Kulomaa MS, Tuohimaa PJ
Endocrinology 1990 Feb;126(2):1143-55
Endocrinology 1990 Feb;126(2):1143-55
Progesterone receptor concentration differences in the chick oviduct cells and apparent down-regulation by ligand. A semiquantitative immunohistochemical study.
Pekki A, Joensuu T, Vidqvist K, Toft D, Tuohimaa P
Journal of steroid biochemistry 1989;34(1-6):351-4
Journal of steroid biochemistry 1989;34(1-6):351-4
Immunocytochemical study of progesterone receptors in hyperplastic and neoplastic endometrial tissues.
Bergeron C, Ferenczy A, Toft DO, Shyamala G
Cancer research 1988 Nov 1;48(21):6132-6
Cancer research 1988 Nov 1;48(21):6132-6
Interaction of murine progesterone receptors with specific monoclonal antibodies to the avian progesterone receptor.
Schneider W, Toft DO, Sullivan WP, Shyamala G
Journal of steroid biochemistry 1988 Mar;29(3):297-306
Journal of steroid biochemistry 1988 Mar;29(3):297-306
Immunocytochemical study of progesterone receptors in the human endometrium during the menstrual cycle.
Bergeron C, Ferenczy A, Toft DO, Schneider W, Shyamala G
Laboratory investigation; a journal of technical methods and pathology 1988 Dec;59(6):862-9
Laboratory investigation; a journal of technical methods and pathology 1988 Dec;59(6):862-9
Immunologic analysis of human breast cancer progesterone receptors. 1. Immunoaffinity purification of transformed receptors and production of monoclonal antibodies.
Estes PA, Suba EJ, Lawler-Heavner J, Elashry-Stowers D, Wei LL, Toft DO, Sullivan WP, Horwitz KB, Edwards DP
Biochemistry 1987 Sep 22;26(19):6250-62
Biochemistry 1987 Sep 22;26(19):6250-62
No comments: Submit comment
Supportive validation
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
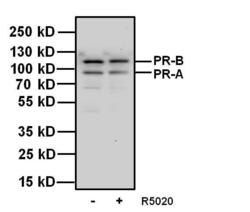
- Western blot analysis of Progesterone Receptor was performed by loading 20 µg of T47D cell lysates, untreated (-) or stimulated (+) with 100 nm promegestone (R5020) for 1 hour and 10 µL PageRuler Plus Prestained Protein Ladder (Product # 26619) per well onto a 4-20% Tris-Glycine polyacrylamide gel. Proteins were transferred to a nitrocellulose membrane using the G2 Fast Blotter (Product # 62288) and blocked with 5% Milk/TBST for at least 1 hour at room temperature. Progesterone Receptor was detected using a Progesterone Receptor mouse monoclonal antibody, Product # MA1-411, at a concentration of 1 µg/mL in blocking buffer overnight at 4°C on a rocking platform, followed by a Superclonal goat anti-Mouse IgG-HRP secondary antibody (Product # A28177) at a dilution of 1:2,000 for at least 1 hour at room temperature. Chemiluminescent detection was performed using SuperSignal West Pico (Product # 34078) and the myECL Imager (Product # 62236).
Supportive validation
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
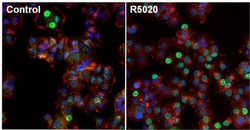
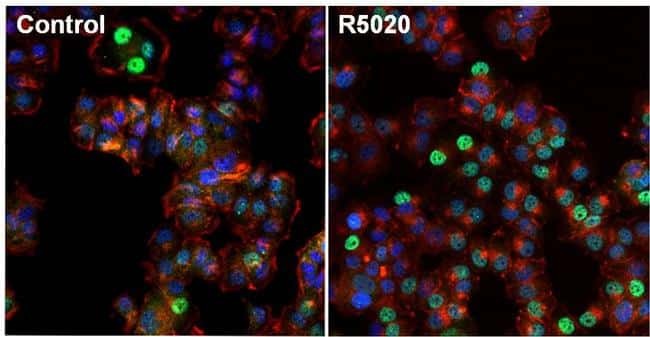
- Immunofluorescent analysis of Progesterone Receptor (green) in T47D cells untreated (left) or stimulated with 100nm promegestone, R5020, for 1 hour (right). The cells were fixed with formalin for 15 minutes, permeabilized with 0.1% Triton X-100 in TBS for 10 minutes, and blocked with 3% Blocker BSA (Product # 37525) for 15 minutes at room temperature. Cells were stained with Progesterone Receptor mouse monoclonal antibody (Product # MA1-411), at a dilution of 1:100 for 1 hour at 37C, and then incubated with a Alexa Fluor 488 Superclonal goat anti-mouse IgG secondary antibody (Product # A28175) at a dilution of 1:1000 for 30 minutes at room temperature (both panels, green). Nuclei (both panels, blue) were stained with Hoechst 33342 dye (Product # 62249). Images were taken on a Thermo Scientific ToxInsight at 20X magnification.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
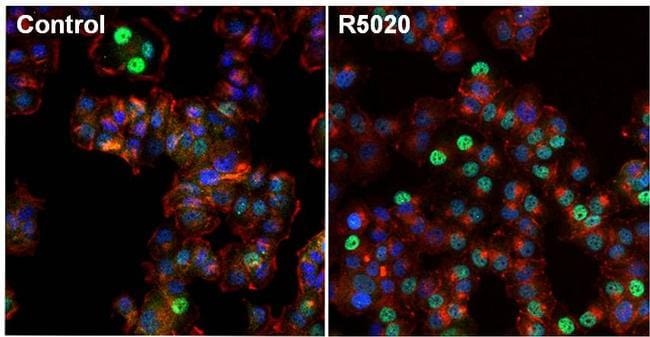
- Immunofluorescent analysis of Progesterone Receptor (green) in T47D cells untreated (left) or stimulated with 100nm promegestone, R5020, for 1 hour (right). The cells were fixed with formalin for 15 minutes, permeabilized with 0.1% Triton X-100 in TBS for 10 minutes, and blocked with 3% Blocker BSA (Product # 37525) for 15 minutes at room temperature. Cells were stained with Progesterone Receptor mouse monoclonal antibody (Product # MA1-411), at a dilution of 1:100 for 1 hour at 37C, and then incubated with a Alexa Fluor 488 Superclonal goat anti-mouse IgG secondary antibody (Product # A28175) at a dilution of 1:1000 for 30 minutes at room temperature (both panels, green). Nuclei (both panels, blue) were stained with Hoechst 33342 dye (Product # 62249). Images were taken on a Thermo Scientific ToxInsight at 20X magnification.
Supportive validation
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
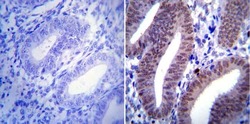
- Immunohistochemistry was performed on human uterus tissue. To expose target protein, antigen was retreived using 10mM sodium citrate followed by microwave treatment for 8-15 minutes. Endogenous peroxidases were blocked in 3% H202-methanol for 15 minutes and tissues were blocked in 3% BSA-PBS for 30 minutes at room temperature. Cells were probed with a Progesterone Receptor mouse monoclonal antibody (Product # MA1-411) at a dilution of 1:20 overnight in a humidified chamber. Tissues were washed in PBST and detection was performed using a secondary antibody conjugated to HRP. DAB staining buffer was applied and tissues were counterstained with hematoxylin and prepped for mounting. Images were taken at 40X magnification.
Supportive validation
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
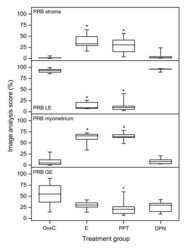
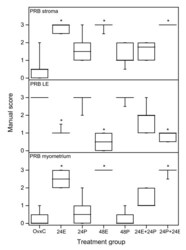
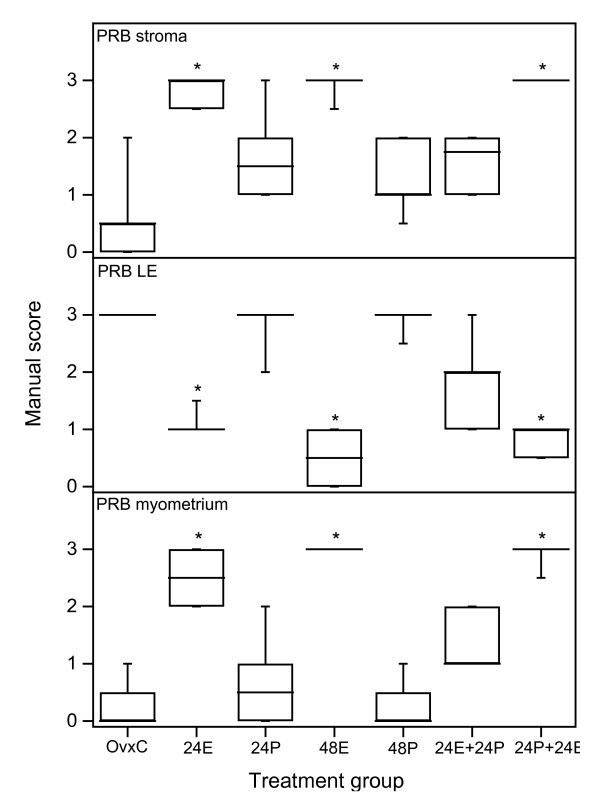
- Figure 3 Scoring results PRB experiment 1 . Results from manual scoring of PRB immunohistochemistry results in stroma (upper panel), luminal epithelium (LE, middle panel) and myometrium (bottom panel). The ""box-and-whisker plot"" represents the median value with 50% of all data falling within the box. The whiskers extend to the 5th and 95th percentiles. An asterisk indicates a significant (p < 0.05) difference compared to the OvxC group.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
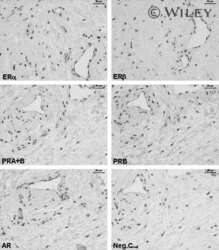
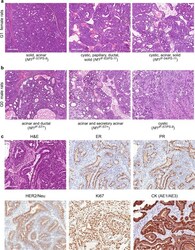
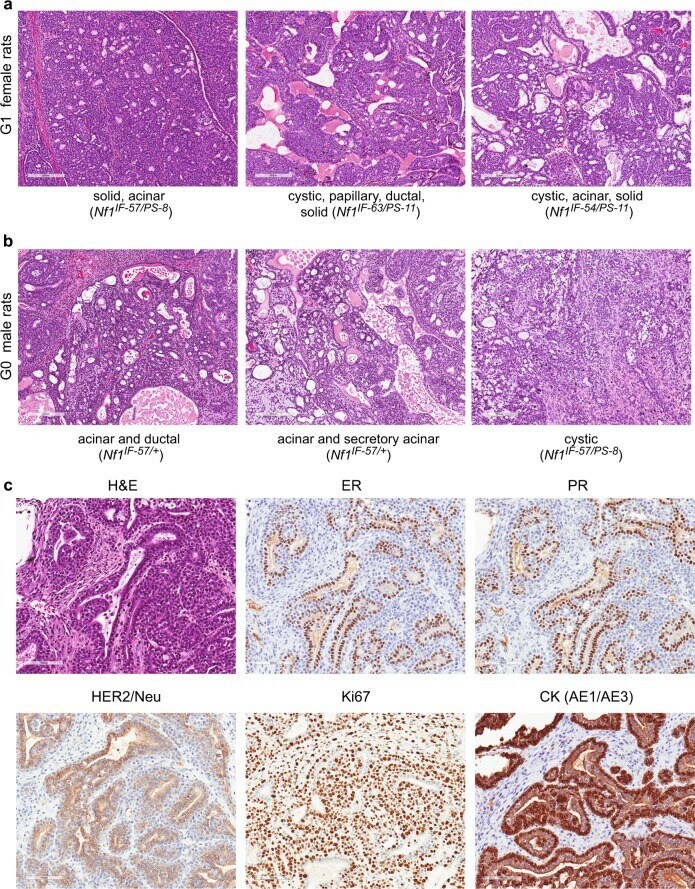
- Fig. 2 Nf1 deficiency induces ER + /PR + mammary tumors. We observed mammary tumors with diverse mammary histology in G0 - G2 animals. a Nf1 females and b Nf1 males developed mammary adenocarcinomas with mixed histology including acinar, cystic, papillary, ductal, and solid features. All images were taken at 100X magnification. c Immunostaining of Nf1 mammary tumors ( rNf1 #6) for estrogen receptor, progesterone receptor, HER2, Ki67, and pan-cytokeratin (AE1/AE3). All immunostaining images were taken at 200X magnification
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
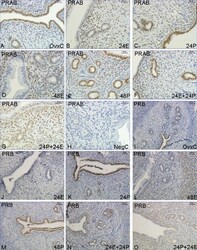
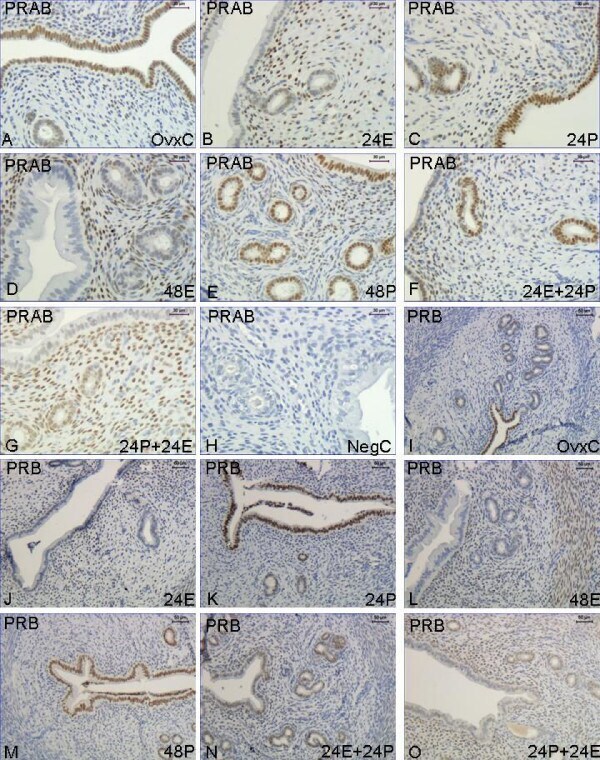
- Figure 1 Immunohistochemistry experiment 1 . Representative images of immunohistochemical results from each hormone treatment group in experiment 1, PRAB A-H , PRB I-O . Treatment groups are as follows: OvxC: A and I , 24E: B and J . 24P: C and K , 48E: D and L , 48P: E and M , 24E+24P: F and N , 24P+24E: G and O . Magnification bar 30 mum in images A-H and 50 mum in images I-O.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
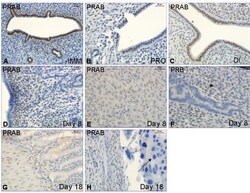
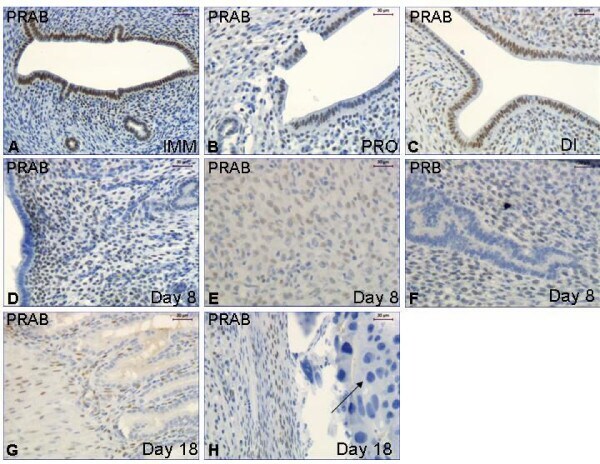
- Figure 4 Immunohistochemistry experiment 2 . Representative images of immunohistochemical results from the immature ( A ), cycling ( B,C ) and pregnant rats ( D-H ). Image F shows PRB immunostaining while all the others show PRAB. The arrow in image H indicates the negative cells in the placenta. Magnification bar 30 mum in all images.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
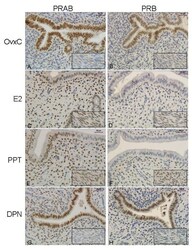
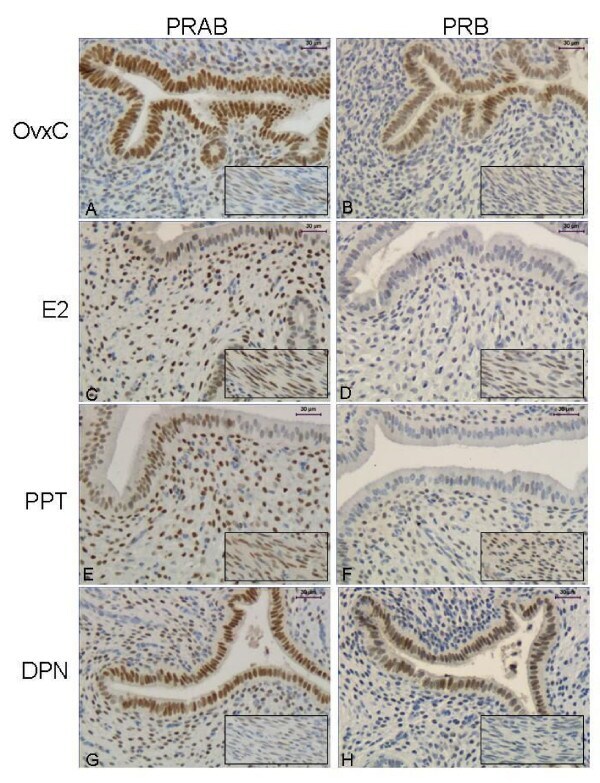
- Figure 5 Immunohistochemistry experiment 3 . Representative images of immunohistochemical results from the agonist treated rats, PRAB in left columnand PRB in right column. Representative images are shown from a rat in each treatment group as follows: OvxC ( A,B ), E2 ( C,D ), PPT ( E,F ) and DPN ( G,H ). The insert shows the myometrium from the same uterus. Magnification bar 30 mum.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
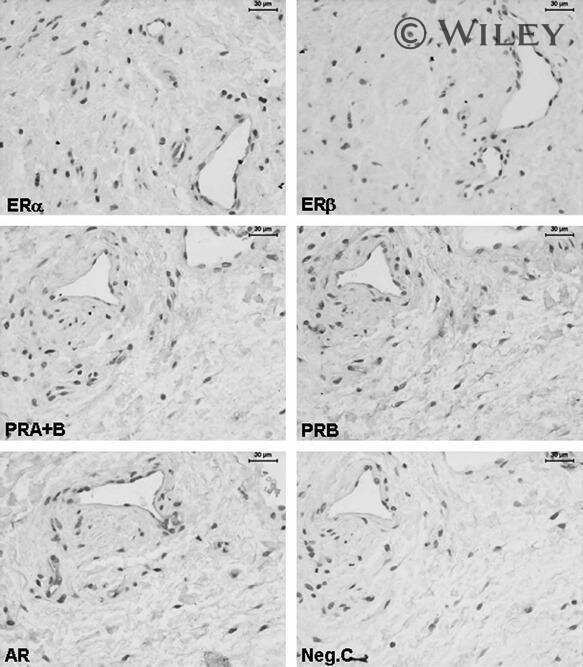
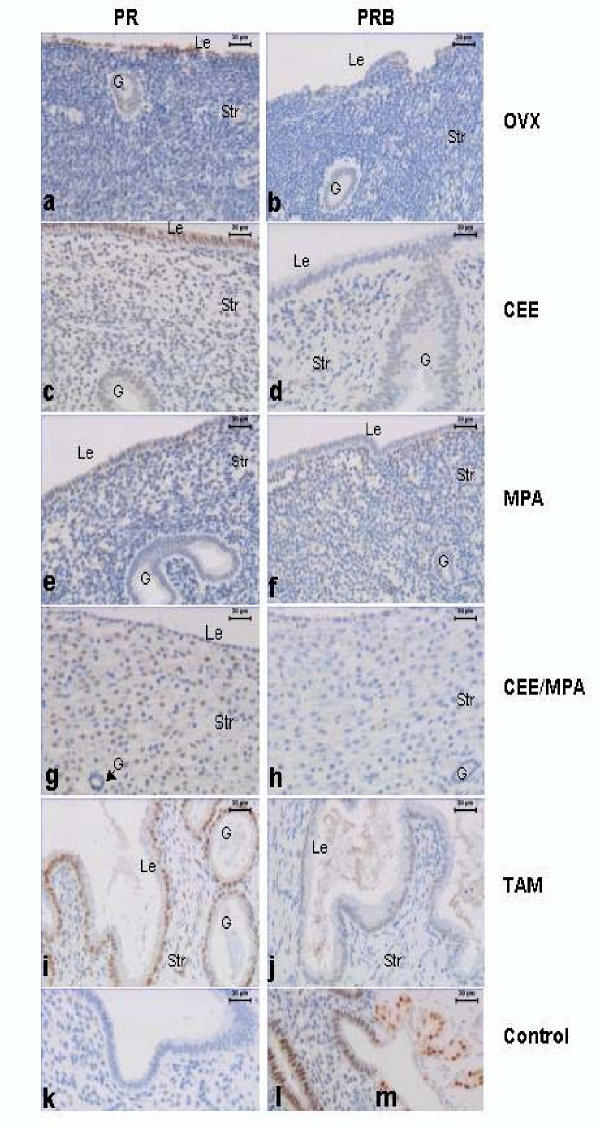
- Figure 1 Immunohistochemical localization of PR and PRB in the superficial endometrial stroma (Str), glands (G) and luminal epithelium (Le). Positive nuclear immunostaining for PR is present in the study groups as shown: OVX ( a ); CEE ( c ); MPA ( e ); CEE /MPA ( g ), and TAM ( i ). Nuclear PRB immunostaining is shown for: OVX ( b ); CEE ( d ); MPA ( f ); CEE /MPA ( h ) and TAM ( j ) groups, (bar = 30 mum). A negative control is shown for PR where the first antibody is replaced by mouse IgG ( k ). Positive controls for PRB immunostaining in human endometrium ( l ) and breast tissue from a CEE treated monkey ( m ), (bar = 30 mum).
- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
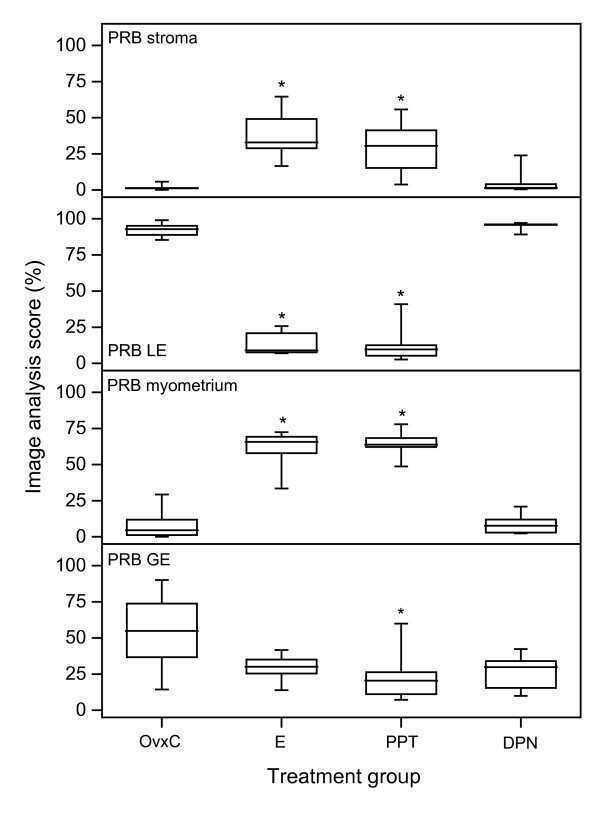
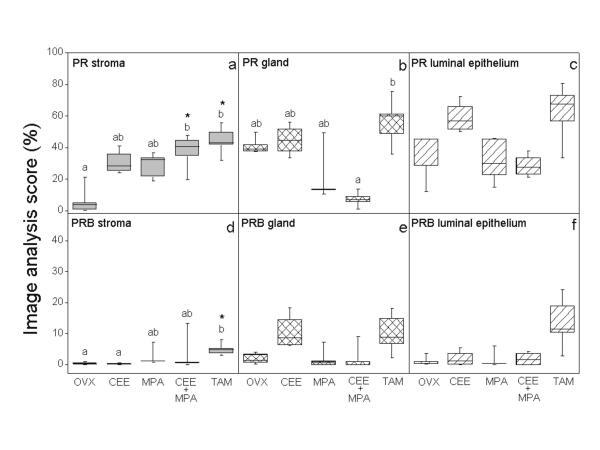
- Figure 3 Image analysis scores of positive PR immunoreactivity in superficial stroma ( a ), superficial glands ( b ) and luminal epithelium ( c ). PRB immunostaining is shown in superficial stroma ( d ), superficial glands ( e ) and luminal epithelium ( f ). Box and whisker plots representing the median value with 50% of all data falling within the box. The whiskers extend to the 5 th and 95 th percentiles. In OVX, MPA and CEE/MPA groups: n = 5, in CEE group n = 4 and in the TAM group n = 6. Values with different letter designations are significantly different (p < 0.05). Values with * is significantly different (p < 0.05) to OVX control.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
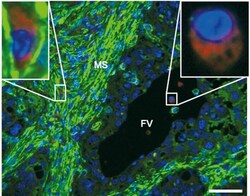
- FIGURE 5 Composition and distribution of the fetal and maternal macrophage population in bovine placenta. Placental sections of a healthy term cow were stained with LAMP-1 (red), alpha-SMA (green, cytoplasm) and PGR (green, nucleus). MS: maternal stroma and FV: fetal villi. Scale bar =50mum
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
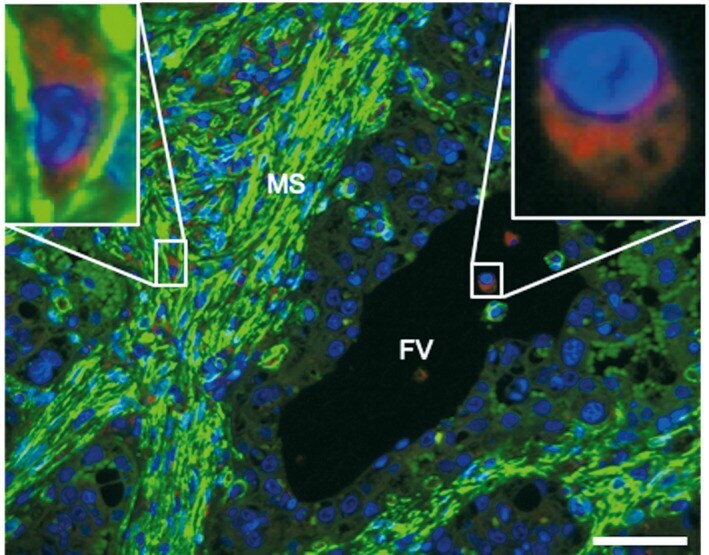
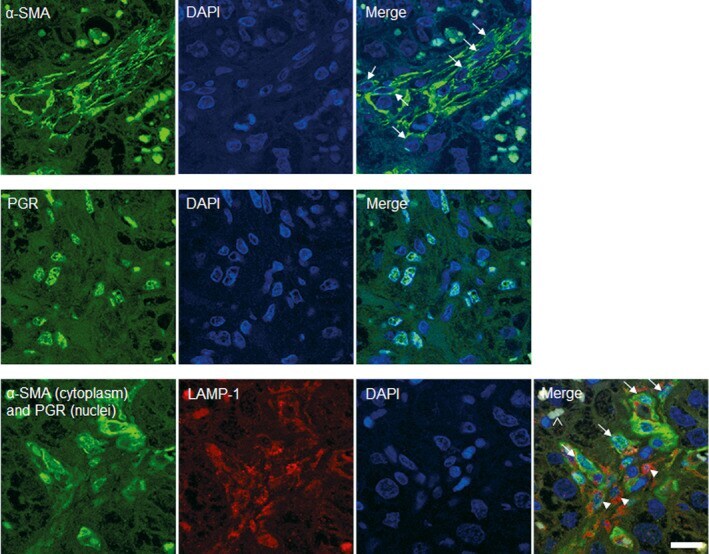
- FIGURE 4 Immunofluorescent labeling of cell populations located in maternal stroma. Placental sections from healthy term cows were stained with LAMP-1 (red), alpha-SMA (green, cytoplasm) and PGR (green, nucleus). The detection of PGR was performed to more easily differentiate cell types in maternal stroma. In the merged images maternal macrophages (arrowheads) and myofibroblasts (arrows) in maternal stroma (MS) adjacent to fetal villi (FV) are labelled. Nuclei were labeled with DAPI (blue). Scale bar = 15 mum
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
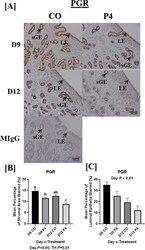
- Experimental details
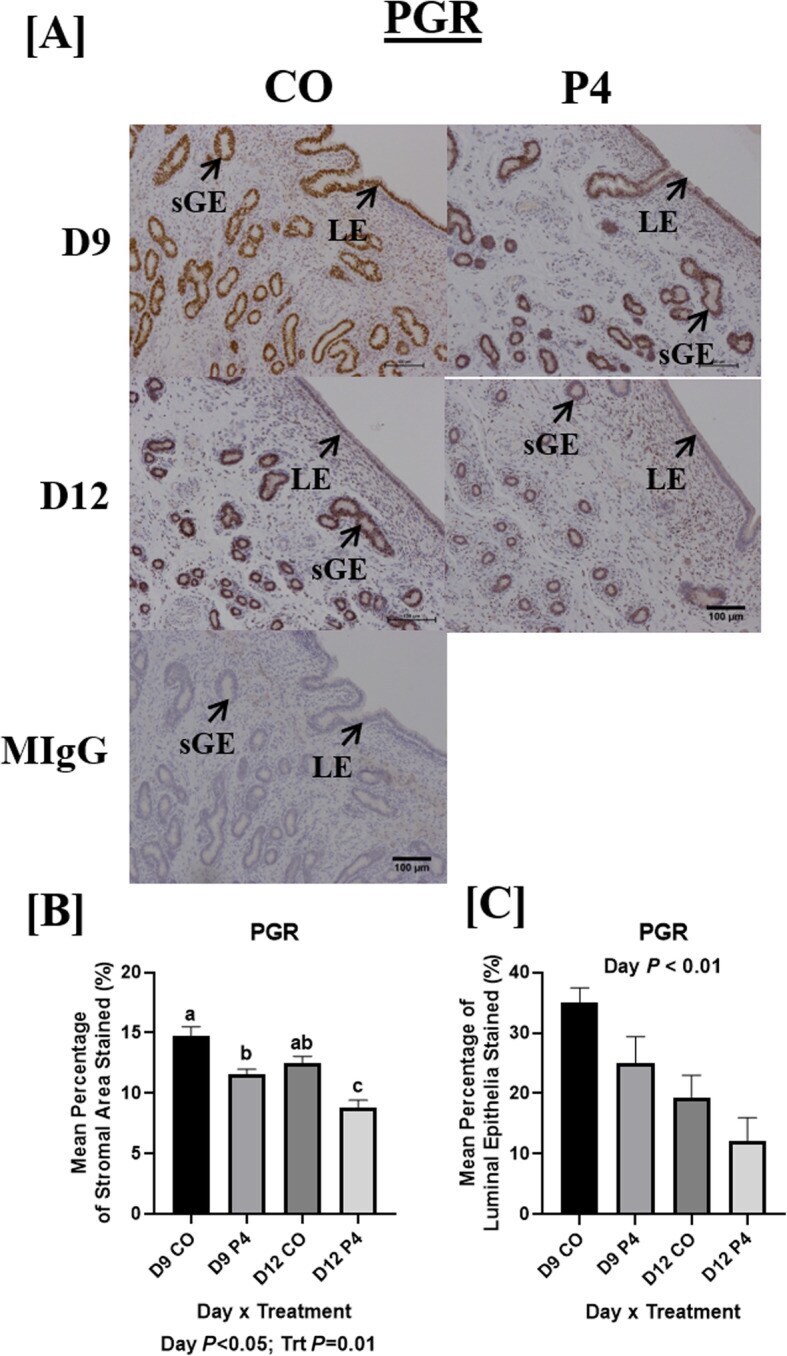
- Fig. 7 Localization of endometrial PGR. PGR staining was most abundant in endometrial luminal epithelium (LE) and superficial glandular epithelia (sGE) in CO-treated ewes necropsied on day 9 of pregnancy and was markedly reduced in endometria of P4-treated ewes necropsied on day 9 and 12 ( a and b ). Similar PGR localization and staining intensity was found in the LE of CO and P4-treated ewes necropsied on day 12. PGR in endometria of P4-treated ewes necropsied on day 12 was least abundant in both sGE and LE. Scale bar represents 100 mum. Only endometria from ewes which were considered pregnant with normally developed conceptuse were utilized for these analyses