Explore
Explore Validate
Validate Learn
Learn Western blot
Western blot Immunohistochemistry
ImmunohistochemistryAntibody data
- Antibody Data
- Antigen structure
- References [58]
- Comments [0]
- Validations
- Immunohistochemistry [2]
- Other assay [28]
Submit
Validation data
Reference
Comment
Report error
- Product number
- 36-4500 - Provider product page

- Provider
- Invitrogen Antibodies
- Product name
- Claudin 11 Polyclonal Antibody
- Antibody type
- Polyclonal
- Antigen
- Synthetic peptide
- Reactivity
- Human, Mouse
- Host
- Rabbit
- Isotype
- IgG
- Vial size
- 100 μg
- Concentration
- 0.25 mg/mL
- Storage
- -20°C
Submitted references Single-Cell RNA-Seq of Cisplatin-Treated Adult Stria Vascularis Identifies Cell Type-Specific Regulatory Networks and Novel Therapeutic Gene Targets.
Cullin4 E3 Ubiquitin Ligases Regulate Male Gonocyte Migration, Proliferation and Blood-Testis Barrier Homeostasis.
iPSC-derived myelinoids to study myelin biology of humans.
Fertility Preservation in Childhood Cancer: Endocrine Activity in Prepubertal Human Testis Xenografts Exposed to a Pubertal Hormone Environment.
Noncoding Microdeletion in Mouse Hgf Disrupts Neural Crest Migration into the Stria Vascularis, Reduces the Endocochlear Potential, and Suggests the Neuropathology for Human Nonsyndromic Deafness DFNB39.
Distinct Roles for Rac1 in Sertoli Cell Function during Testicular Development and Spermatogenesis.
Single Cell and Single Nucleus RNA-Seq Reveal Cellular Heterogeneity and Homeostatic Regulatory Networks in Adult Mouse Stria Vascularis.
The cell-cell junctions of mammalian testes: II. The lamellar smooth muscle monolayer cells of the peritubular wall are laterally connected by vertical adherens junctions-a novel architectonic cell-cell junction system.
Planar cell polarity protein Dishevelled 3 (Dvl3) regulates ectoplasmic specialization (ES) dynamics in the testis through changes in cytoskeletal organization.
Deafness and loss of cochlear hair cells in the absence of thyroid hormone transporters Slc16a2 (Mct8) and Slc16a10 (Mct10).
ESRP1 Mutations Cause Hearing Loss due to Defects in Alternative Splicing that Disrupt Cochlear Development.
Regulation of spermatogenesis by a local functional axis in the testis: role of the basement membrane-derived noncollagenous 1 domain peptide.
Highly Conserved Testicular Localization of Claudin-11 in Normal and Impaired Spermatogenesis.
Interleukin 1alpha-induced disruption of the Sertoli cell cytoskeleton affects gap junctional communication.
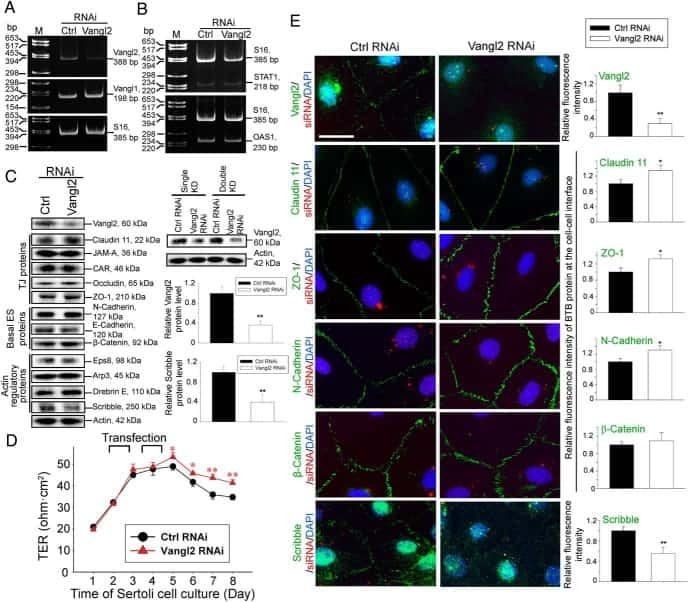
Planar Cell Polarity (PCP) Protein Vangl2 Regulates Ectoplasmic Specialization Dynamics via Its Effects on Actin Microfilaments in the Testes of Male Rats.
Increased expression of dermatopontin and its implications for testicular dysfunction in mice.
Claudin-11 and occludin are major contributors to Sertoli cell tight junction function, in vitro.
Rescue of perfluorooctanesulfonate (PFOS)-mediated Sertoli cell injury by overexpression of gap junction protein connexin 43.
Modeling immune functions of the mouse blood-cerebrospinal fluid barrier in vitro: primary rather than immortalized mouse choroid plexus epithelial cells are suited to study immune cell migration across this brain barrier.
Overexpression of PRL7D1 in Leydig Cells Causes Male Reproductive Dysfunction in Mice.
Overexpression of plastin 3 in Sertoli cells disrupts actin microfilament bundle homeostasis and perturbs the tight junction barrier.
Gonadotropin suppression in men leads to a reduction in claudin-11 at the Sertoli cell tight junction.
Actin-bundling protein plastin 3 is a regulator of ectoplasmic specialization dynamics during spermatogenesis in the rat testis.
Musashi-1 maintains blood-testis barrier structure during spermatogenesis and regulates stress granule formation upon heat stress.
Dermatopontin is a novel regulator of the CdCl2-induced decrease in claudin-11 expression.
rpS6 regulates blood-testis barrier dynamics through Akt-mediated effects on MMP-9.
Differential permeability of the blood-testis barrier during reinitiation of spermatogenesis in adult male rats.
Claudin-11 and connexin-43 display altered spatial patterns of organization in men with primary seminiferous tubule failure compared with controls.
Retinoic acid promotes Sertoli cell differentiation and antagonises activin-induced proliferation.
Nf1 loss and Ras hyperactivation in oligodendrocytes induce NOS-driven defects in myelin and vasculature.
SOX8 regulates permeability of the blood-testes barrier that affects adult male fertility in the mouse.
Intercellular adhesion molecule-2 is involved in apical ectoplasmic specialization dynamics during spermatogenesis in the rat.
A survey of Sertoli cell differentiation in men after gonadotropin suppression and in testicular cancer.
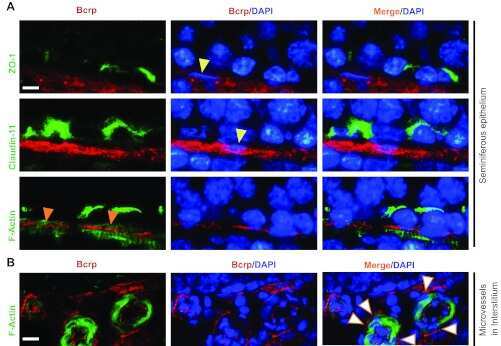
Breast cancer resistance protein regulates apical ectoplasmic specialization dynamics stage specifically in the rat testis.
Regulation of the blood-testis barrier by coxsackievirus and adenovirus receptor.
Lentiviral transduction of rat Sertoli cells as a means to modify gene expression.
Focal adhesion kinase-Tyr407 and -Tyr397 exhibit antagonistic effects on blood-testis barrier dynamics in the rat.
Spermatogonial stem cells alone are not sufficient to re-initiate spermatogenesis in the rat testis following adjudin-induced infertility.
Activin signaling regulates Sertoli cell differentiation and function.
Androgen initiates Sertoli cell tight junction formation in the hypogonadal (hpg) mouse.
Enhanced immunohistochemical resolution of claudin proteins in glycolmethacrylate-embedded tissue biopsies.
Postnatal changes in the expression of claudin-11 in the testes and excurrent ducts of the domestic rabbit (Oryctolagus cuniculus domesticus).
A study to assess the assembly of a functional blood-testis barrier in developing rat testes.
Claudin-11 expression increased in spermatogenic defect in human testes.
Pou3f4 deficiency causes defects in otic fibrocytes and stria vascularis by different mechanisms.
Expression and immunohistochemical localization of TMEM16A/anoctamin 1, a calcium-activated chloride channel in the mouse cochlea.
Differential effects of testosterone and TGF-β3 on endocytic vesicle-mediated protein trafficking events at the blood-testis barrier.
Adjudin-mediated Sertoli-germ cell junction disassembly affects Sertoli cell barrier function in vitro and in vivo.
Gonadotropins regulate rat testicular tight junctions in vivo.
Differential expression of claudin tight junction proteins in the human cortical nephron.
Claudin-1 has tumor suppressive activity and is a direct target of RUNX3 in gastric epithelial cells.
Tight junction proteins in human Schwann cell autotypic junctions.
Drug transporter, P-glycoprotein (MDR1), is an integrated component of the mammalian blood-testis barrier.
Transcriptomic screening of microvascular endothelial cells implicates novel molecular regulators of vascular dysfunction after spinal cord injury.
Regulation of testicular tight junctions by gonadotrophins in the adult Djungarian hamster in vivo.
Adult sertoli cells are not terminally differentiated in the Djungarian hamster: effect of FSH on proliferation and junction protein organization.
Androgens regulate the permeability of the blood-testis barrier.
Distribution of the tight junction proteins ZO-1, occludin, and claudin-4, -8, and -12 in bladder epithelium.
Taukulis IA, Olszewski RT, Korrapati S, Fernandez KA, Boger ET, Fitzgerald TS, Morell RJ, Cunningham LL, Hoa M
Frontiers in molecular neuroscience 2021;14:718241
Frontiers in molecular neuroscience 2021;14:718241
Cullin4 E3 Ubiquitin Ligases Regulate Male Gonocyte Migration, Proliferation and Blood-Testis Barrier Homeostasis.
Yin Y, Zhu L, Li Q, Zhou P, Ma L
Cells 2021 Oct 13;10(10)
Cells 2021 Oct 13;10(10)
iPSC-derived myelinoids to study myelin biology of humans.
James OG, Selvaraj BT, Magnani D, Burr K, Connick P, Barton SK, Vasistha NA, Hampton DW, Story D, Smigiel R, Ploski R, Brophy PJ, Ffrench-Constant C, Lyons DA, Chandran S
Developmental cell 2021 May 3;56(9):1346-1358.e6
Developmental cell 2021 May 3;56(9):1346-1358.e6
Fertility Preservation in Childhood Cancer: Endocrine Activity in Prepubertal Human Testis Xenografts Exposed to a Pubertal Hormone Environment.
Hutka M, Kadam P, Van Saen D, Homer NZM, Onofre J, Wallace WHB, Smith LB, Stukenborg JB, Goossens E, Mitchell RT
Cancers 2020 Sep 30;12(10)
Cancers 2020 Sep 30;12(10)
Noncoding Microdeletion in Mouse Hgf Disrupts Neural Crest Migration into the Stria Vascularis, Reduces the Endocochlear Potential, and Suggests the Neuropathology for Human Nonsyndromic Deafness DFNB39.
Morell RJ, Olszewski R, Tona R, Leitess S, Wafa TT, Taukulis I, Schultz JM, Thomason EJ, Richards K, Whitley BN, Hill C, Saunders T, Starost MF, Fitzgerald T, Wilson E, Ohyama T, Friedman TB, Hoa M
The Journal of neuroscience : the official journal of the Society for Neuroscience 2020 Apr 8;40(15):2976-2992
The Journal of neuroscience : the official journal of the Society for Neuroscience 2020 Apr 8;40(15):2976-2992
Distinct Roles for Rac1 in Sertoli Cell Function during Testicular Development and Spermatogenesis.
Heinrich A, Potter SJ, Guo L, Ratner N, DeFalco T
Cell reports 2020 Apr 14;31(2):107513
Cell reports 2020 Apr 14;31(2):107513
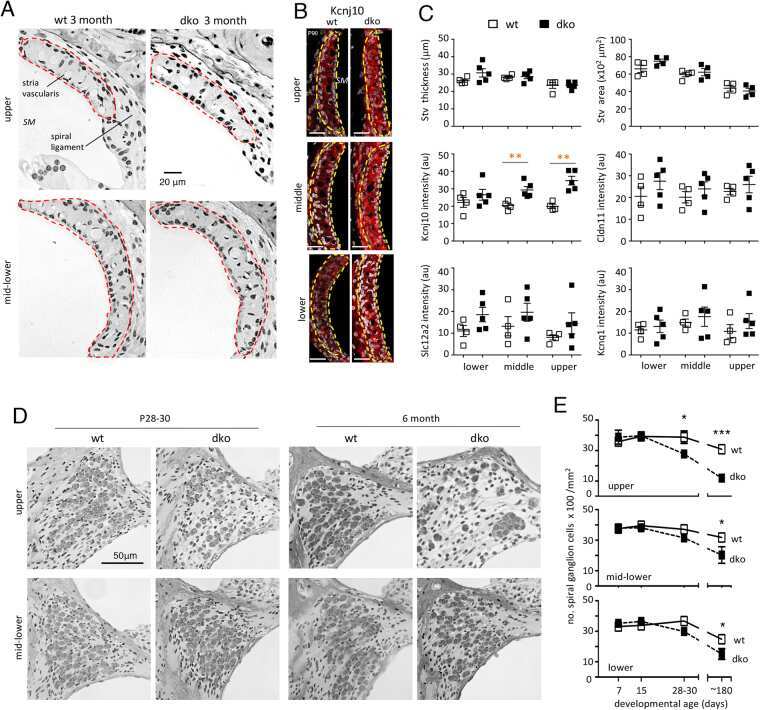
Single Cell and Single Nucleus RNA-Seq Reveal Cellular Heterogeneity and Homeostatic Regulatory Networks in Adult Mouse Stria Vascularis.
Korrapati S, Taukulis I, Olszewski R, Pyle M, Gu S, Singh R, Griffiths C, Martin D, Boger E, Morell RJ, Hoa M
Frontiers in molecular neuroscience 2019;12:316
Frontiers in molecular neuroscience 2019;12:316
The cell-cell junctions of mammalian testes: II. The lamellar smooth muscle monolayer cells of the peritubular wall are laterally connected by vertical adherens junctions-a novel architectonic cell-cell junction system.
Domke LM, Franke WW
Cell and tissue research 2019 Feb;375(2):451-482
Cell and tissue research 2019 Feb;375(2):451-482
Planar cell polarity protein Dishevelled 3 (Dvl3) regulates ectoplasmic specialization (ES) dynamics in the testis through changes in cytoskeletal organization.
Li L, Mao B, Yan M, Wu S, Ge R, Lian Q, Cheng CY
Cell death & disease 2019 Feb 26;10(3):194
Cell death & disease 2019 Feb 26;10(3):194
Deafness and loss of cochlear hair cells in the absence of thyroid hormone transporters Slc16a2 (Mct8) and Slc16a10 (Mct10).
Sharlin DS, Ng L, Verrey F, Visser TJ, Liu Y, Olszewski RT, Hoa M, Heuer H, Forrest D
Scientific reports 2018 Mar 13;8(1):4403
Scientific reports 2018 Mar 13;8(1):4403
ESRP1 Mutations Cause Hearing Loss due to Defects in Alternative Splicing that Disrupt Cochlear Development.
Rohacek AM, Bebee TW, Tilton RK, Radens CM, McDermott-Roe C, Peart N, Kaur M, Zaykaner M, Cieply B, Musunuru K, Barash Y, Germiller JA, Krantz ID, Carstens RP, Epstein DJ
Developmental cell 2017 Nov 6;43(3):318-331.e5
Developmental cell 2017 Nov 6;43(3):318-331.e5
Regulation of spermatogenesis by a local functional axis in the testis: role of the basement membrane-derived noncollagenous 1 domain peptide.
Chen H, Mruk DD, Lee WM, Cheng CY
FASEB journal : official publication of the Federation of American Societies for Experimental Biology 2017 Aug;31(8):3587-3607
FASEB journal : official publication of the Federation of American Societies for Experimental Biology 2017 Aug;31(8):3587-3607
Highly Conserved Testicular Localization of Claudin-11 in Normal and Impaired Spermatogenesis.
Stammler A, Lüftner BU, Kliesch S, Weidner W, Bergmann M, Middendorff R, Konrad L
PloS one 2016;11(8):e0160349
PloS one 2016;11(8):e0160349
Interleukin 1alpha-induced disruption of the Sertoli cell cytoskeleton affects gap junctional communication.
Chojnacka K, Bilinska B, Mruk DD
Cellular signalling 2016 May;28(5):469-480
Cellular signalling 2016 May;28(5):469-480
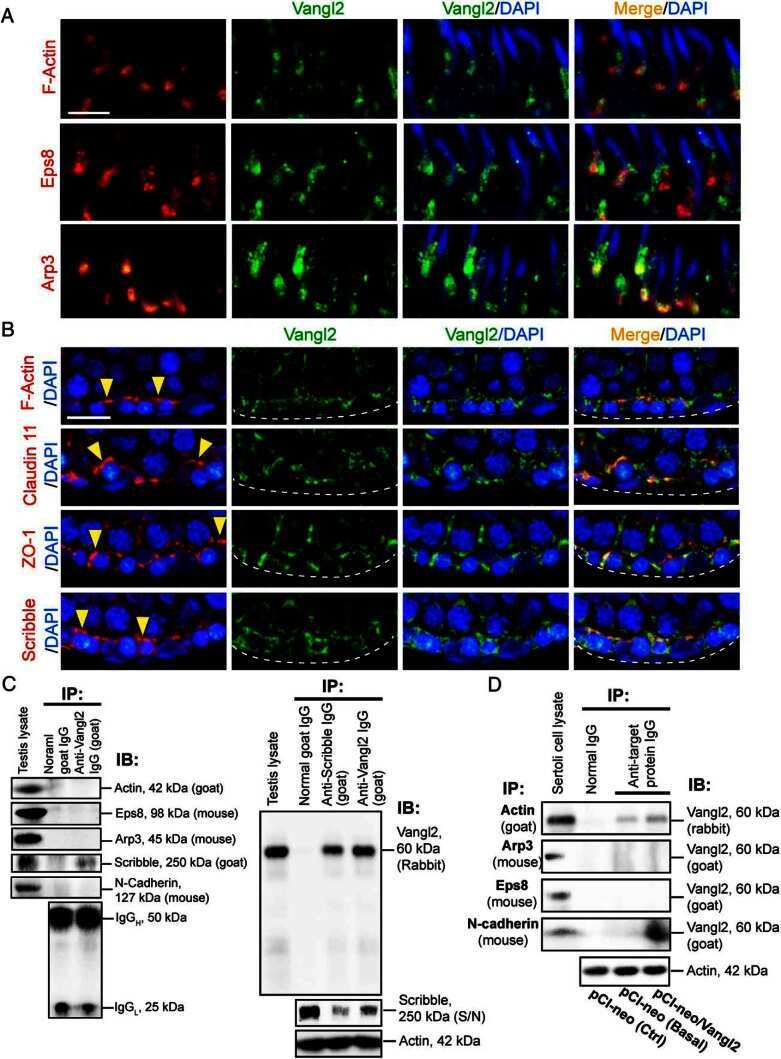
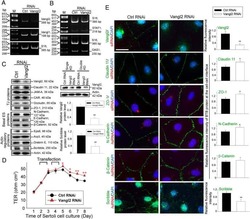
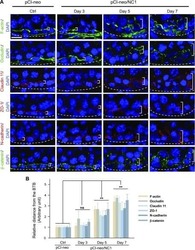
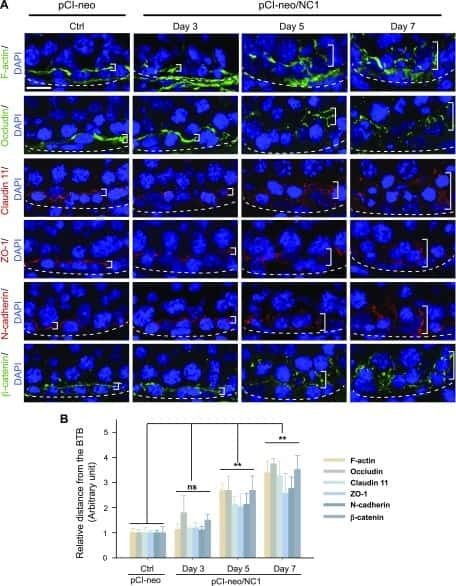
Planar Cell Polarity (PCP) Protein Vangl2 Regulates Ectoplasmic Specialization Dynamics via Its Effects on Actin Microfilaments in the Testes of Male Rats.
Chen H, Mruk DD, Lee WM, Cheng CY
Endocrinology 2016 May;157(5):2140-59
Endocrinology 2016 May;157(5):2140-59
Increased expression of dermatopontin and its implications for testicular dysfunction in mice.
Cai J, Liu W, Hao J, Chen M, Li G
Molecular medicine reports 2016 Mar;13(3):2431-8
Molecular medicine reports 2016 Mar;13(3):2431-8
Claudin-11 and occludin are major contributors to Sertoli cell tight junction function, in vitro.
McCabe MJ, Foo CF, Dinger ME, Smooker PM, Stanton PG
Asian journal of andrology 2016 Jul-Aug;18(4):620-6
Asian journal of andrology 2016 Jul-Aug;18(4):620-6
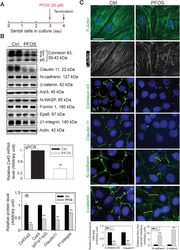
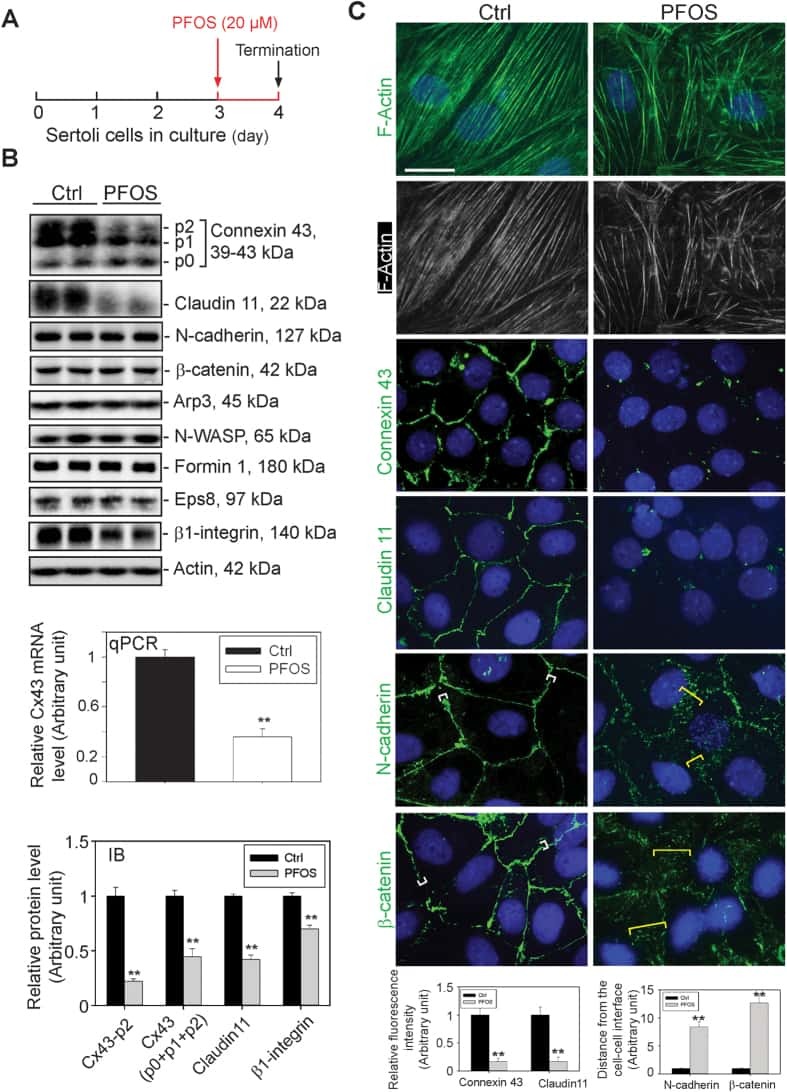
Rescue of perfluorooctanesulfonate (PFOS)-mediated Sertoli cell injury by overexpression of gap junction protein connexin 43.
Li N, Mruk DD, Chen H, Wong CK, Lee WM, Cheng CY
Scientific reports 2016 Jul 20;6:29667
Scientific reports 2016 Jul 20;6:29667
Modeling immune functions of the mouse blood-cerebrospinal fluid barrier in vitro: primary rather than immortalized mouse choroid plexus epithelial cells are suited to study immune cell migration across this brain barrier.
Lazarevic I, Engelhardt B
Fluids and barriers of the CNS 2016 Jan 29;13:2
Fluids and barriers of the CNS 2016 Jan 29;13:2
Overexpression of PRL7D1 in Leydig Cells Causes Male Reproductive Dysfunction in Mice.
Liu Y, Su X, Hao J, Chen M, Liu W, Liao X, Li G
International journal of molecular sciences 2016 Jan 13;17(1)
International journal of molecular sciences 2016 Jan 13;17(1)
Overexpression of plastin 3 in Sertoli cells disrupts actin microfilament bundle homeostasis and perturbs the tight junction barrier.
Li N, Lee WM, Cheng CY
Spermatogenesis 2016 Apr;6(1):e1206353
Spermatogenesis 2016 Apr;6(1):e1206353
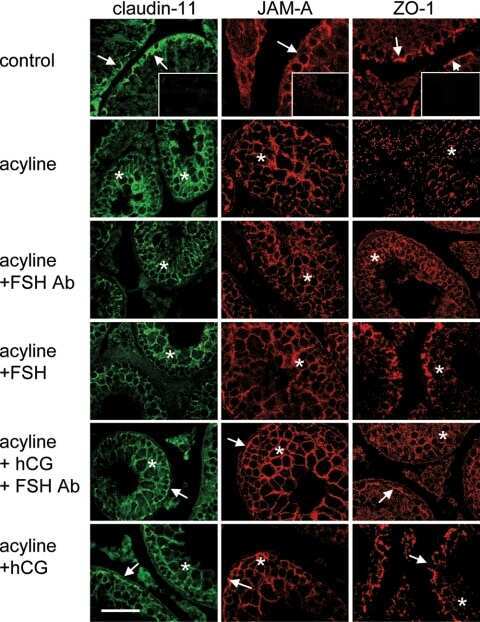
Gonadotropin suppression in men leads to a reduction in claudin-11 at the Sertoli cell tight junction.
McCabe MJ, Tarulli GA, Laven-Law G, Matthiesson KL, Meachem SJ, McLachlan RI, Dinger ME, Stanton PG
Human reproduction (Oxford, England) 2016 Apr;31(4):875-86
Human reproduction (Oxford, England) 2016 Apr;31(4):875-86
Actin-bundling protein plastin 3 is a regulator of ectoplasmic specialization dynamics during spermatogenesis in the rat testis.
Li N, Mruk DD, Wong CK, Lee WM, Han D, Cheng CY
FASEB journal : official publication of the Federation of American Societies for Experimental Biology 2015 Sep;29(9):3788-805
FASEB journal : official publication of the Federation of American Societies for Experimental Biology 2015 Sep;29(9):3788-805
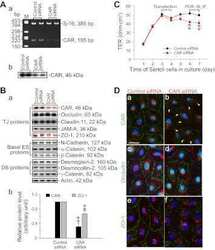
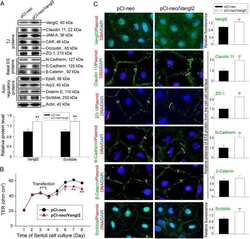
Musashi-1 maintains blood-testis barrier structure during spermatogenesis and regulates stress granule formation upon heat stress.
ErLin S, WenJie W, LiNing W, BingXin L, MingDe L, Yan S, RuiFa H
Molecular biology of the cell 2015 May 15;26(10):1947-56
Molecular biology of the cell 2015 May 15;26(10):1947-56
Dermatopontin is a novel regulator of the CdCl2-induced decrease in claudin-11 expression.
Yang Q, Hao J, Chen M, Li G
Toxicology in vitro : an international journal published in association with BIBRA 2014 Sep;28(6):1158-64
Toxicology in vitro : an international journal published in association with BIBRA 2014 Sep;28(6):1158-64
rpS6 regulates blood-testis barrier dynamics through Akt-mediated effects on MMP-9.
Mok KW, Mruk DD, Cheng CY
Journal of cell science 2014 Nov 15;127(Pt 22):4870-82
Journal of cell science 2014 Nov 15;127(Pt 22):4870-82
Differential permeability of the blood-testis barrier during reinitiation of spermatogenesis in adult male rats.
Haverfield JT, Meachem SJ, Nicholls PK, Rainczuk KE, Simpson ER, Stanton PG
Endocrinology 2014 Mar;155(3):1131-44
Endocrinology 2014 Mar;155(3):1131-44
Claudin-11 and connexin-43 display altered spatial patterns of organization in men with primary seminiferous tubule failure compared with controls.
Haverfield JT, Meachem SJ, O'Bryan MK, McLachlan RI, Stanton PG
Fertility and sterility 2013 Sep;100(3):658-66
Fertility and sterility 2013 Sep;100(3):658-66
Retinoic acid promotes Sertoli cell differentiation and antagonises activin-induced proliferation.
Nicholls PK, Harrison CA, Rainczuk KE, Wayne Vogl A, Stanton PG
Molecular and cellular endocrinology 2013 Sep 5;377(1-2):33-43
Molecular and cellular endocrinology 2013 Sep 5;377(1-2):33-43
Nf1 loss and Ras hyperactivation in oligodendrocytes induce NOS-driven defects in myelin and vasculature.
Mayes DA, Rizvi TA, Titus-Mitchell H, Oberst R, Ciraolo GM, Vorhees CV, Robinson AP, Miller SD, Cancelas JA, Stemmer-Rachamimov AO, Ratner N
Cell reports 2013 Sep 26;4(6):1197-212
Cell reports 2013 Sep 26;4(6):1197-212
SOX8 regulates permeability of the blood-testes barrier that affects adult male fertility in the mouse.
Singh AP, Cummings CA, Mishina Y, Archer TK
Biology of reproduction 2013 May;88(5):133
Biology of reproduction 2013 May;88(5):133
Intercellular adhesion molecule-2 is involved in apical ectoplasmic specialization dynamics during spermatogenesis in the rat.
Xiao X, Cheng CY, Mruk DD
The Journal of endocrinology 2013 Jan;216(1):73-86
The Journal of endocrinology 2013 Jan;216(1):73-86
A survey of Sertoli cell differentiation in men after gonadotropin suppression and in testicular cancer.
Tarulli GA, Stanton PG, Loveland KL, Rajpert-De Meyts E, McLachlan RI, Meachem SJ
Spermatogenesis 2013 Jan 1;3(1):e24014
Spermatogenesis 2013 Jan 1;3(1):e24014
Breast cancer resistance protein regulates apical ectoplasmic specialization dynamics stage specifically in the rat testis.
Qian X, Mruk DD, Wong EW, Cheng CY
American journal of physiology. Endocrinology and metabolism 2013 Apr 1;304(7):E757-69
American journal of physiology. Endocrinology and metabolism 2013 Apr 1;304(7):E757-69
Regulation of the blood-testis barrier by coxsackievirus and adenovirus receptor.
Su L, Mruk DD, Cheng CY
American journal of physiology. Cell physiology 2012 Oct 15;303(8):C843-53
American journal of physiology. Cell physiology 2012 Oct 15;303(8):C843-53
Lentiviral transduction of rat Sertoli cells as a means to modify gene expression.
Nicholls PK, Stanton PG, Rainczuk KE, Qian H, Gregorevic P, Harrison CA
Spermatogenesis 2012 Oct 1;2(4):279-284
Spermatogenesis 2012 Oct 1;2(4):279-284
Focal adhesion kinase-Tyr407 and -Tyr397 exhibit antagonistic effects on blood-testis barrier dynamics in the rat.
Lie PP, Mruk DD, Mok KW, Su L, Lee WM, Cheng CY
Proceedings of the National Academy of Sciences of the United States of America 2012 Jul 31;109(31):12562-7
Proceedings of the National Academy of Sciences of the United States of America 2012 Jul 31;109(31):12562-7
Spermatogonial stem cells alone are not sufficient to re-initiate spermatogenesis in the rat testis following adjudin-induced infertility.
Mok KW, Mruk DD, Lee WM, Cheng CY
International journal of andrology 2012 Feb;35(1):86-101
International journal of andrology 2012 Feb;35(1):86-101
Activin signaling regulates Sertoli cell differentiation and function.
Nicholls PK, Stanton PG, Chen JL, Olcorn JS, Haverfield JT, Qian H, Walton KL, Gregorevic P, Harrison CA
Endocrinology 2012 Dec;153(12):6065-77
Endocrinology 2012 Dec;153(12):6065-77
Androgen initiates Sertoli cell tight junction formation in the hypogonadal (hpg) mouse.
McCabe MJ, Allan CM, Foo CF, Nicholls PK, McTavish KJ, Stanton PG
Biology of reproduction 2012 Aug;87(2):38
Biology of reproduction 2012 Aug;87(2):38
Enhanced immunohistochemical resolution of claudin proteins in glycolmethacrylate-embedded tissue biopsies.
Collins JE, Kirk A, Campbell SK, Mason J, Wilson SJ
Methods in molecular biology (Clifton, N.J.) 2011;762:371-82
Methods in molecular biology (Clifton, N.J.) 2011;762:371-82
Postnatal changes in the expression of claudin-11 in the testes and excurrent ducts of the domestic rabbit (Oryctolagus cuniculus domesticus).
Park CJ, Lee JE, Oh YS, Shim S, Kim DM, Park NC, Park HJ, Gye MC
Journal of andrology 2011 May-Jun;32(3):295-306
Journal of andrology 2011 May-Jun;32(3):295-306
A study to assess the assembly of a functional blood-testis barrier in developing rat testes.
Mok KW, Mruk DD, Lee WM, Cheng CY
Spermatogenesis 2011 Jul;1(3):270-280
Spermatogenesis 2011 Jul;1(3):270-280
Claudin-11 expression increased in spermatogenic defect in human testes.
Nah WH, Lee JE, Park HJ, Park NC, Gye MC
Fertility and sterility 2011 Jan;95(1):385-8
Fertility and sterility 2011 Jan;95(1):385-8
Pou3f4 deficiency causes defects in otic fibrocytes and stria vascularis by different mechanisms.
Song MH, Choi SY, Wu L, Oh SK, Lee HK, Lee DJ, Shim DB, Choi JY, Kim UK, Bok J
Biochemical and biophysical research communications 2011 Jan 7;404(1):528-33
Biochemical and biophysical research communications 2011 Jan 7;404(1):528-33
Expression and immunohistochemical localization of TMEM16A/anoctamin 1, a calcium-activated chloride channel in the mouse cochlea.
Jeon JH, Park JW, Lee JW, Jeong SW, Yeo SW, Kim IB
Cell and tissue research 2011 Aug;345(2):223-30
Cell and tissue research 2011 Aug;345(2):223-30
Differential effects of testosterone and TGF-β3 on endocytic vesicle-mediated protein trafficking events at the blood-testis barrier.
Su L, Mruk DD, Lee WM, Cheng CY
Experimental cell research 2010 Oct 15;316(17):2945-60
Experimental cell research 2010 Oct 15;316(17):2945-60
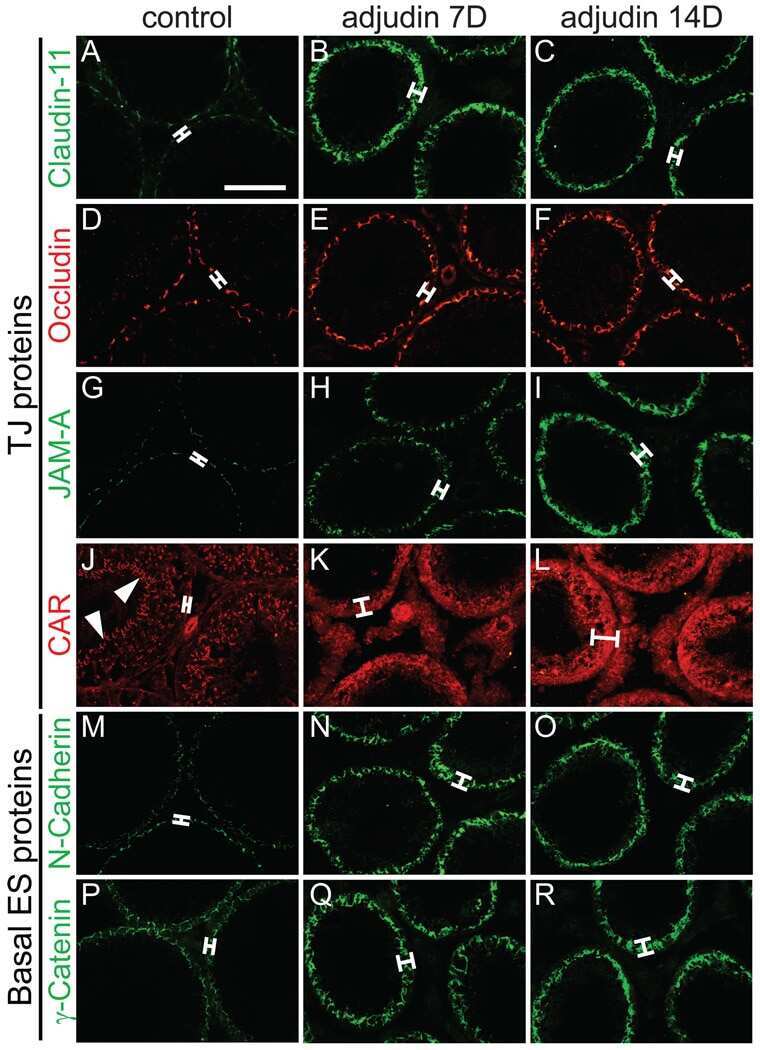
Adjudin-mediated Sertoli-germ cell junction disassembly affects Sertoli cell barrier function in vitro and in vivo.
Su L, Cheng CY, Mruk DD
The international journal of biochemistry & cell biology 2010 Nov;42(11):1864-75
The international journal of biochemistry & cell biology 2010 Nov;42(11):1864-75
Gonadotropins regulate rat testicular tight junctions in vivo.
McCabe MJ, Tarulli GA, Meachem SJ, Robertson DM, Smooker PM, Stanton PG
Endocrinology 2010 Jun;151(6):2911-22
Endocrinology 2010 Jun;151(6):2911-22
Differential expression of claudin tight junction proteins in the human cortical nephron.
Kirk A, Campbell S, Bass P, Mason J, Collins J
Nephrology, dialysis, transplantation : official publication of the European Dialysis and Transplant Association - European Renal Association 2010 Jul;25(7):2107-19
Nephrology, dialysis, transplantation : official publication of the European Dialysis and Transplant Association - European Renal Association 2010 Jul;25(7):2107-19
Claudin-1 has tumor suppressive activity and is a direct target of RUNX3 in gastric epithelial cells.
Chang TL, Ito K, Ko TK, Liu Q, Salto-Tellez M, Yeoh KG, Fukamachi H, Ito Y
Gastroenterology 2010 Jan;138(1):255-65.e1-3
Gastroenterology 2010 Jan;138(1):255-65.e1-3
Tight junction proteins in human Schwann cell autotypic junctions.
Alanne MH, Pummi K, Heape AM, Grènman R, Peltonen J, Peltonen S
The journal of histochemistry and cytochemistry : official journal of the Histochemistry Society 2009 Jun;57(6):523-9
The journal of histochemistry and cytochemistry : official journal of the Histochemistry Society 2009 Jun;57(6):523-9
Drug transporter, P-glycoprotein (MDR1), is an integrated component of the mammalian blood-testis barrier.
Su L, Cheng CY, Mruk DD
The international journal of biochemistry & cell biology 2009 Dec;41(12):2578-87
The international journal of biochemistry & cell biology 2009 Dec;41(12):2578-87
Transcriptomic screening of microvascular endothelial cells implicates novel molecular regulators of vascular dysfunction after spinal cord injury.
Benton RL, Maddie MA, Worth CA, Mahoney ET, Hagg T, Whittemore SR
Journal of cerebral blood flow and metabolism : official journal of the International Society of Cerebral Blood Flow and Metabolism 2008 Nov;28(11):1771-85
Journal of cerebral blood flow and metabolism : official journal of the International Society of Cerebral Blood Flow and Metabolism 2008 Nov;28(11):1771-85
Regulation of testicular tight junctions by gonadotrophins in the adult Djungarian hamster in vivo.
Tarulli GA, Meachem SJ, Schlatt S, Stanton PG
Reproduction (Cambridge, England) 2008 Jun;135(6):867-77
Reproduction (Cambridge, England) 2008 Jun;135(6):867-77
Adult sertoli cells are not terminally differentiated in the Djungarian hamster: effect of FSH on proliferation and junction protein organization.
Tarulli GA, Stanton PG, Lerchl A, Meachem SJ
Biology of reproduction 2006 May;74(5):798-806
Biology of reproduction 2006 May;74(5):798-806
Androgens regulate the permeability of the blood-testis barrier.
Meng J, Holdcraft RW, Shima JE, Griswold MD, Braun RE
Proceedings of the National Academy of Sciences of the United States of America 2005 Nov 15;102(46):16696-700
Proceedings of the National Academy of Sciences of the United States of America 2005 Nov 15;102(46):16696-700
Distribution of the tight junction proteins ZO-1, occludin, and claudin-4, -8, and -12 in bladder epithelium.
Acharya P, Beckel J, Ruiz WG, Wang E, Rojas R, Birder L, Apodaca G
American journal of physiology. Renal physiology 2004 Aug;287(2):F305-18
American journal of physiology. Renal physiology 2004 Aug;287(2):F305-18
No comments: Submit comment
Supportive validation
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
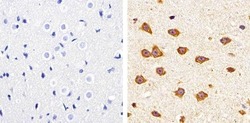
- Experimental details
- Immunohistochemistry analysis of Claudin-11 showing staining in the membrane of paraffin-embedded mouse brain tissue (right) compared to a negative control without primary antibody (left). To expose target proteins, antigen retrieval was performed using 10mM sodium citrate (pH 6.0), microwaved for 8-15 min. Following antigen retrieval, tissues were blocked in 3% H2O2-methanol for 15 min at room temperature, washed with ddH2O and PBS, and then probed with a Claudin-11 polyclonal antibody (Product # 36-4500) diluted in 3% BSA-PBS at a dilution of 1:100 overnight at 4ºC in a humidified chamber. Tissues were washed extensively in PBST and detection was performed using an HRP-conjugated secondary antibody followed by colorimetric detection using a DAB kit. Tissues were counterstained with hematoxylin and dehydrated with ethanol and xylene to prep for mounting.
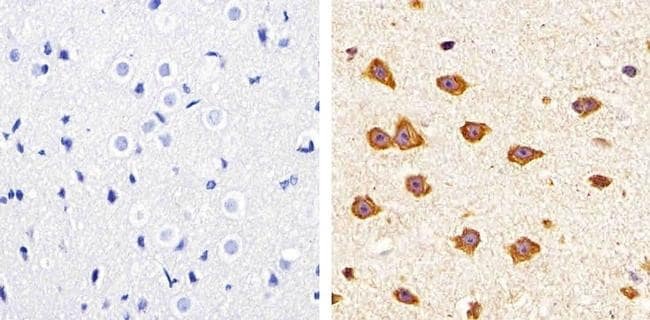
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
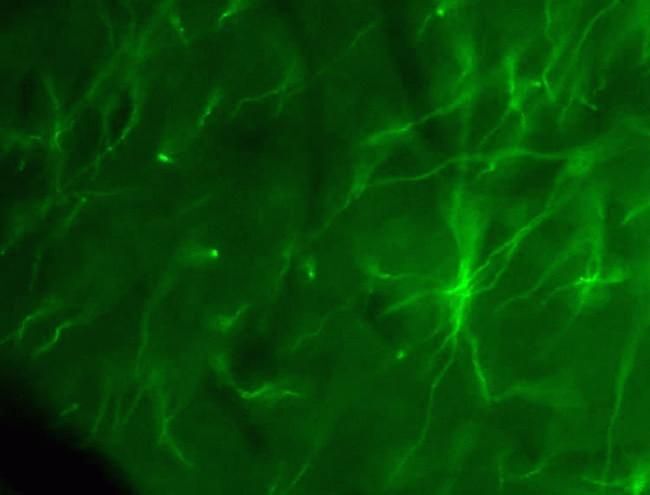
- Immunofluorescent staining of mouse brain using Rb anti-Claudin-11/OSP (Product # 36-4500). Image courtesy of Christina Van Itallie and Dr. James Anderson, University of North Carolina at Chapel Hill, NC. Immunofluorescent staining of mouse brain using Claudin-11/OSP Polyclonal Antibody, Rabbit (Product # 36-4500). Image courtesy of Christina Van Itallie and Dr. James Anderson, University of North Carolina at Chapel Hill, NC.
Supportive validation
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
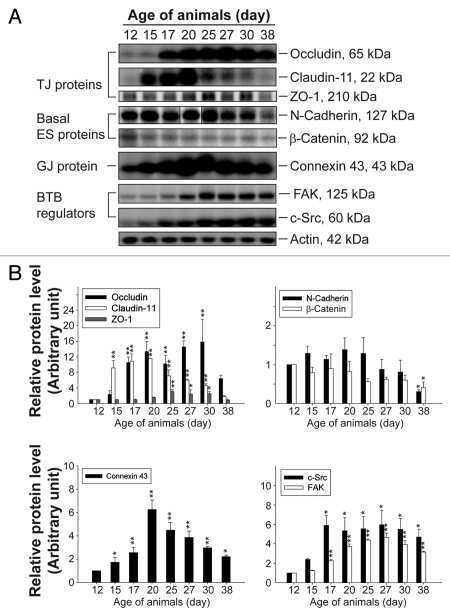
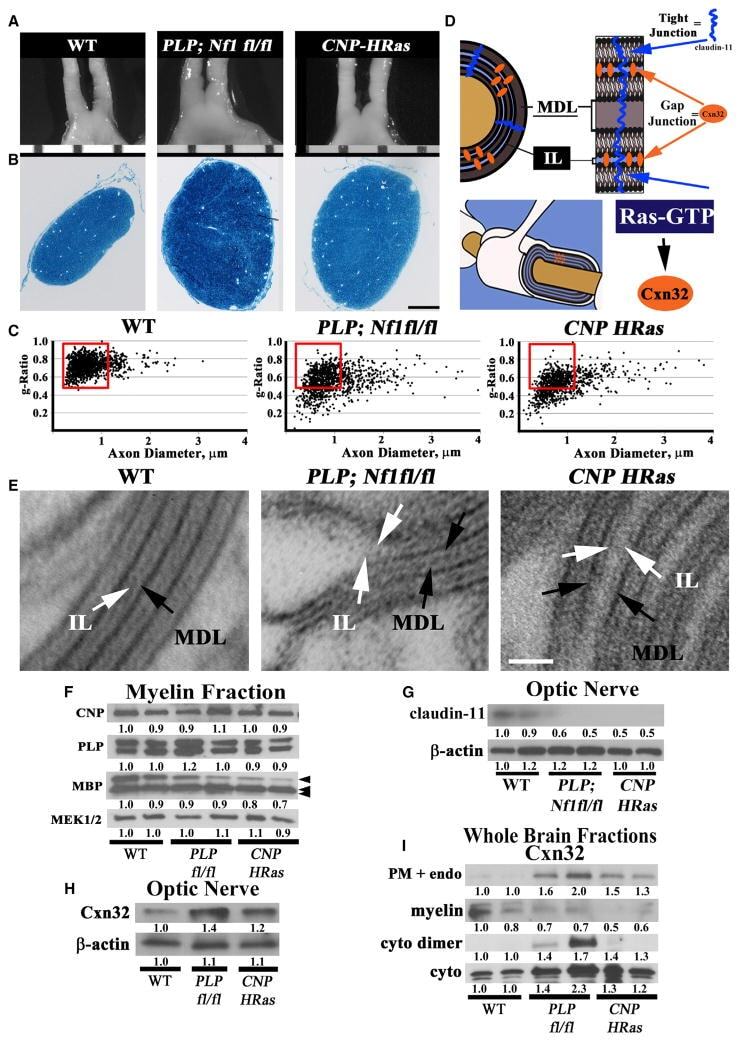
- Figure 1 Nf1 Loss or HRas Activation in Oligodendrocytes Causes Optic-Nerve Enlargement, Myelin Decompaction, Loss of Claudin-11, and Altered Cx32 Localization (A) Gross micrographs of optic nerves at the level of the chiasm in 12-month-old animals. Ruler shows 1 mm markings. (B) Toluidine-blue-stained semithin optic-nerve cross-sections, 1 mm rostral to the chiasm. (C) Optic-nerve g-ratio scatterplots (~5,000 axons/graph; measurements 1 mm rostral to the chiasm in three to five electron micrographs at 10,000x). Red box: 99% of WT g-ratios superimposed upon graphs of other genotypes. ANOVA, Tukey post hoc, p = 2.2 x 10 -16 . (D) Diagram of myelin compaction. Brown circle: axon cross-section; gray: major dense lines (MDLs); orange: Cx32 GJs between compact myelin layers; blue lines: claudin-11+ TJs in compact myelin. (E) Electron micrographs (scale bar, 10 nm) of WT, PLPCre;Nf1 fl / fl , and CNP-HRas optic-nerve myelin. White arrows: intraperiod lines (ILs); black arrows: MDLs. (F-I) Western blots of total optic-nerve lysate or whole-brain cytosolic, endosomal, or myelin fractions showing myelin proteins (F), claudin-11 (G), or Cx32 cellular localization (H and I). n = 3-5 animals/genotype/experiment. See also Figures S1, S2, S3, and S4 .
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
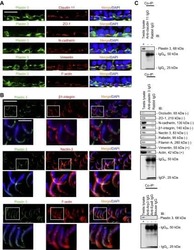
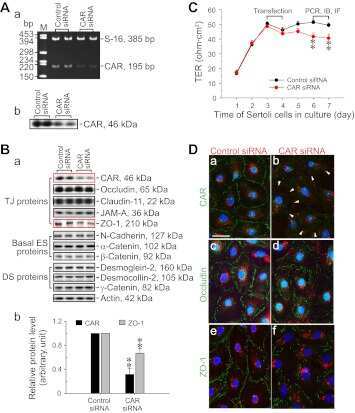
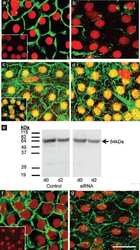
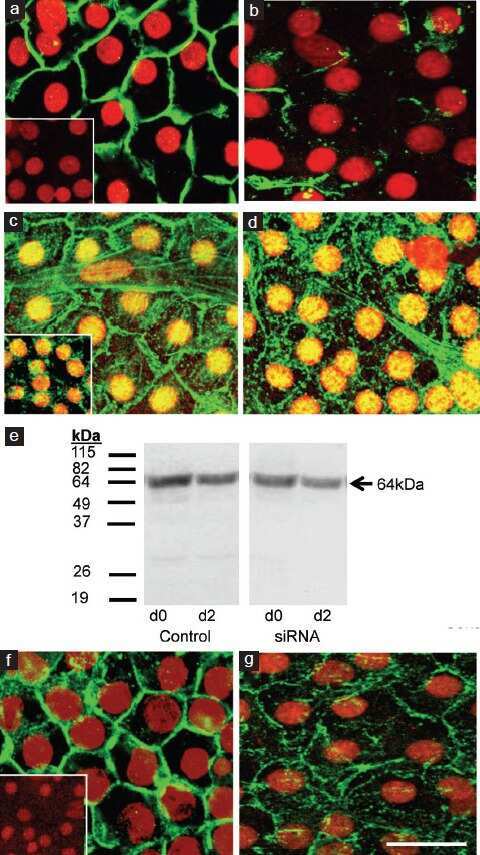
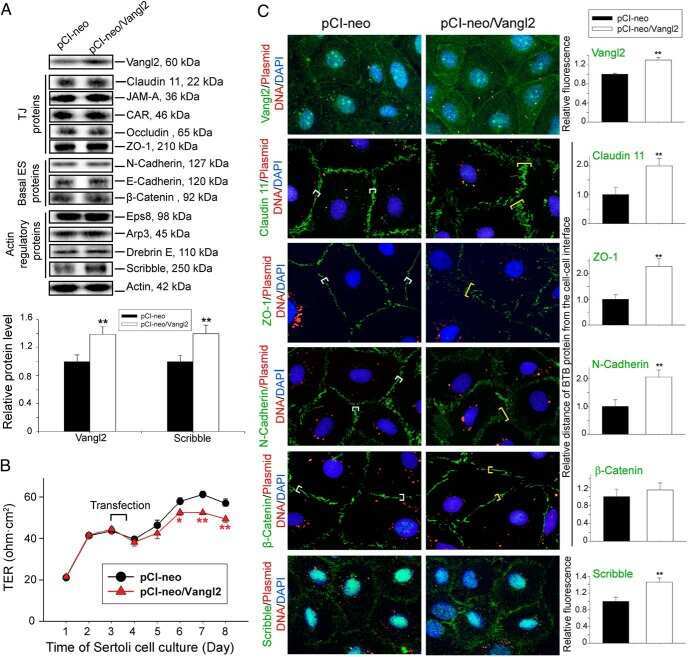
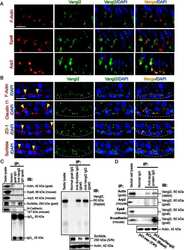
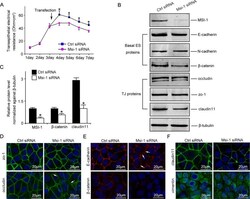
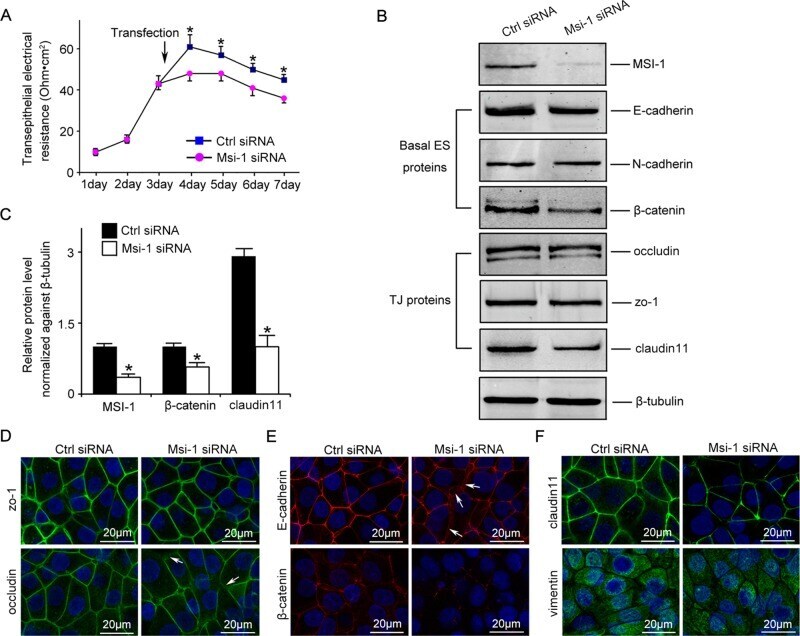
- FIGURE 2: Msi-1 knockdown by RNAi in Sertoli cells perturbs Sertoli cell-cell junctions via changes in BTB-associated proteins. (A) On day 3, Sertoli cells were transfected with Msi-1-specific siRNA duplexes and nontargeting control siRNA duplexes. The TER was detected until day 7, and each data point is the mean +- SD of n = 5 replicates from a representative experiment. *, p < 0.05. (B) Immunoblot showing the steady-state levels of Msi-1, basal ectoplasmic specialization (ES) proteins, and TJ proteins in lysates of Sertoli cells lysed 48 h after transfection; beta-tubulin served as a loading control. (C) Histogram summarizing selected immunoblotting results in B and normalized against beta-tubulin. Each bar represents the mean +- SD of n = 3 experiments. *, p < 0.05. (D-F) Changes in the localization of TJ proteins, basal ES proteins, and the framework protein vimentin at Sertoli cell-cell interfaces after Msi-1 knockdown were assessed 48 h after transfection. Sertoli cell nuclei were stained with DAPI. Note that considerably less beta-catenin and claudin-11 were detected, and the distributions of occludin and E-cadherin (white arrows) were changed at Sertoli cell-cell interfaces after Msi-1 knockdown. Scale bars: 20 mum.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
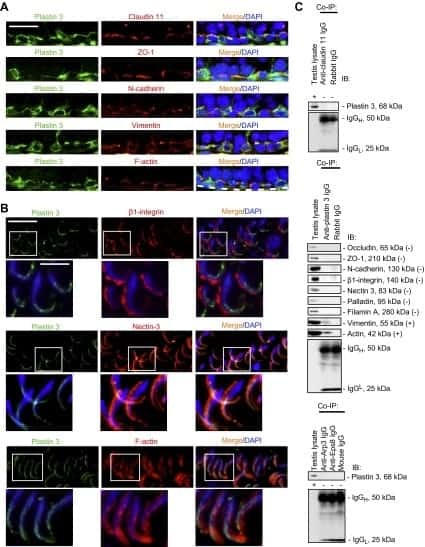
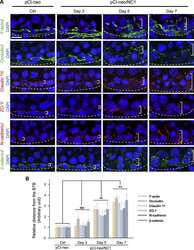
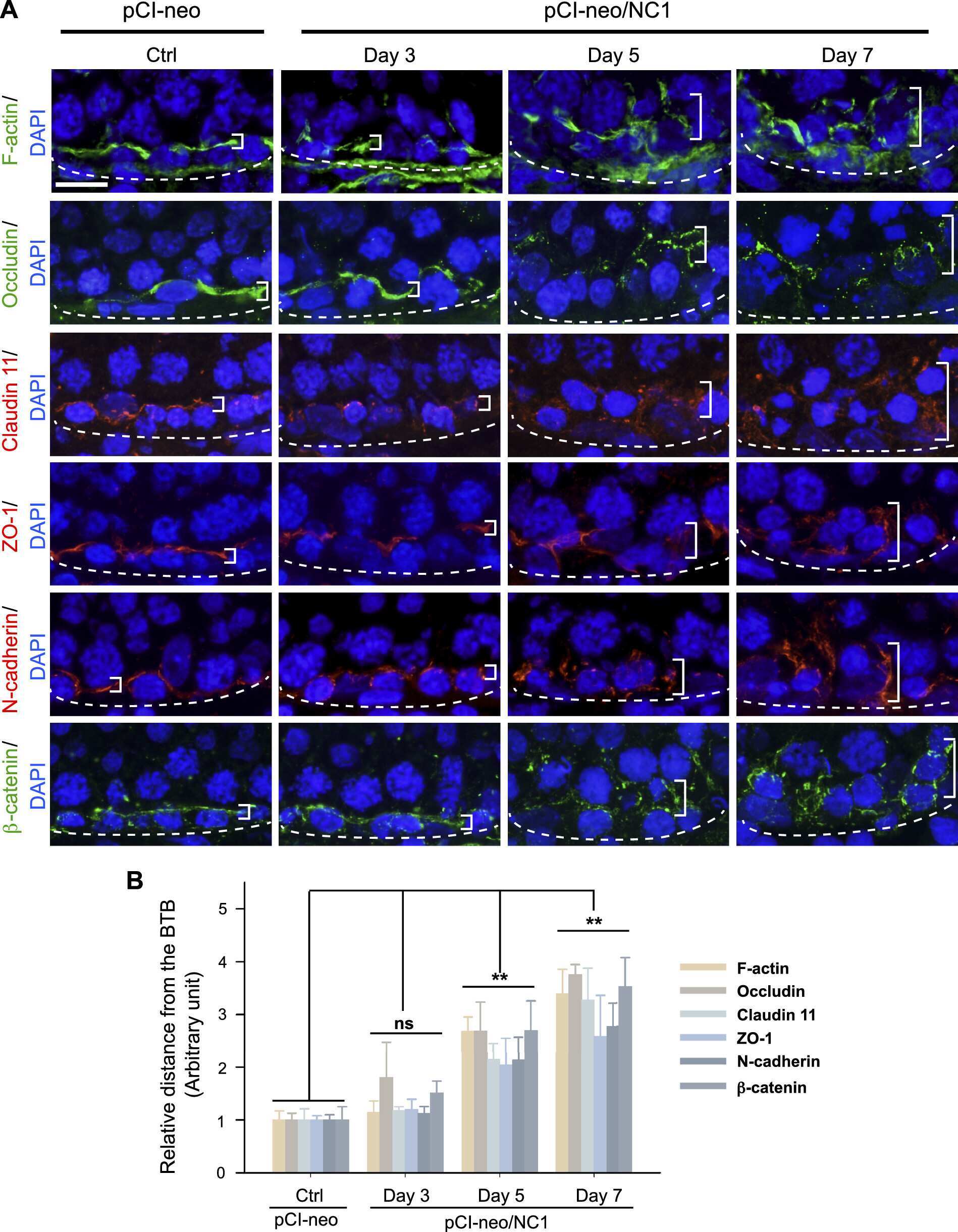
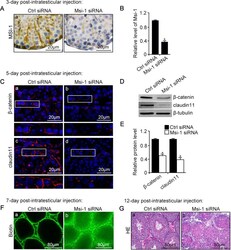
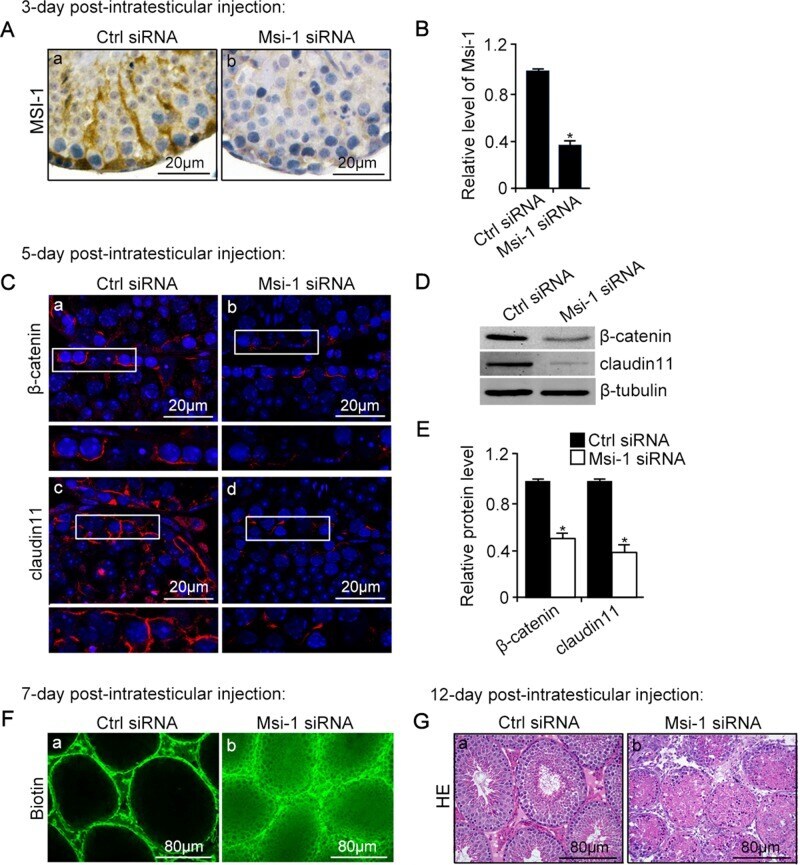
- FIGURE 3: An in vivo study assessing the role of Msi-1 in BTB function and spermatogenesis by RNAi intratesticular injection. Msi-1-targeting siRNA duplexes and nontargeting duplexes were administered to each testis in adult mice as a single treatment. The knockdown efficiency, BTB-associated protein expression, biotin tracer experiment, and hematoxylin-eosin (HE) staining at 3, 5, 9, and 12 d postintratesticular injection, respectively. (A) Considerably more intense Msi-1 staining was observed in the Sertoli cells from testes transfected with nontargeting control (a) versus testes transfected with Msi-1-specific siRNA duplexes (b). Scale bars: 20 mum. (B) Histogram summarizing immunoblotting results of Msi-1 and normalized against beta-tubulin. Each bar represents the mean +- SD of n = 3 experiments. *, p < 0.05. (C) Paraffin sections were used to study changes in the expression of beta-catenin (a and b) and claudin-11 (c and d) after Msi-1 knockdown in vivo. The boxed area was randomly selected. Scale bars: 20 mum. (D) The protein levels of beta-catenin and claudin-11 in nontargeting control and Msi-1 knockdown testes. beta-tubulin served as a loading control. (E) Histogram summarizing selected immunoblotting results in D and normalized against beta-tubulin. Each bar represents the mean +- SD of n = 3 experiments. *, p < 0.05. (F) We injected a biotin tracer into the testes of live anesthetized mice ( n = 3) and examined the subsequent changes in BTB integrity after siRNA d
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
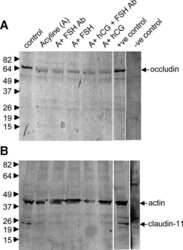
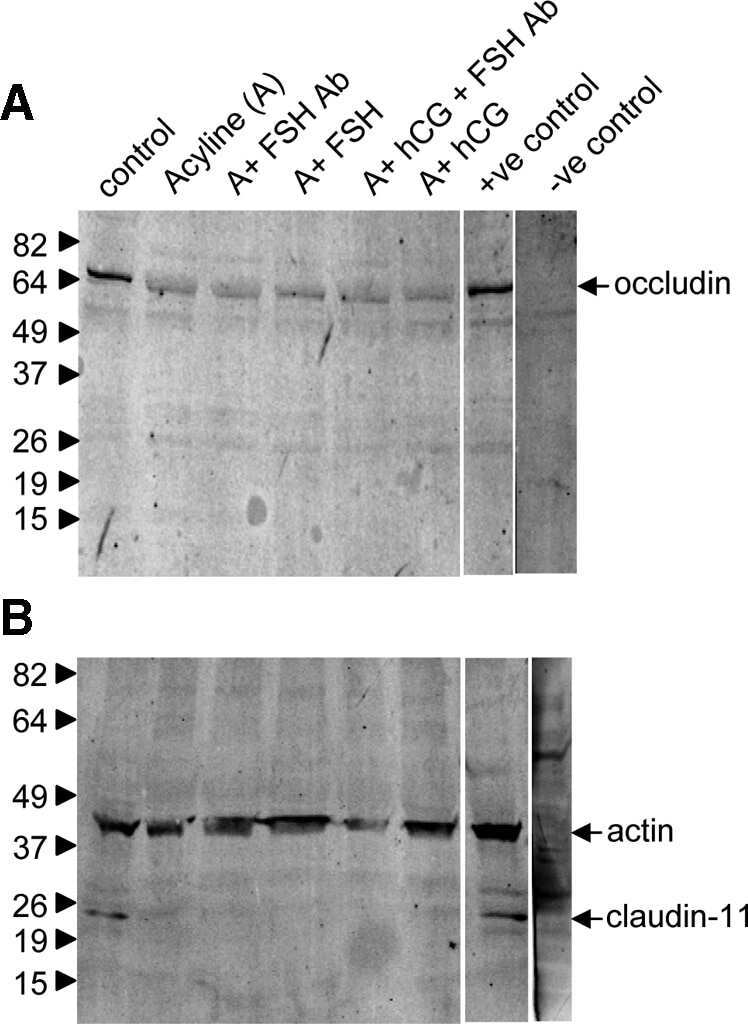
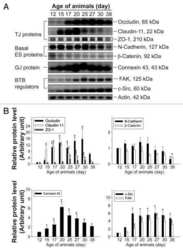
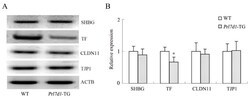
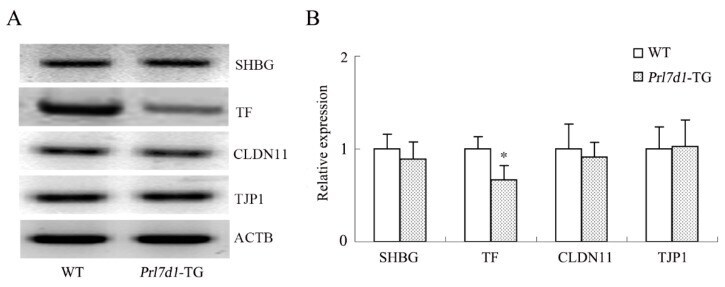
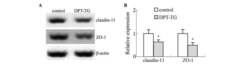
- Figure 6 Western blot analyses of the SHBG, TF, CLDN11 and TJP1 in the testes of wild-type (WT) and Prl7d1 transgenic ( Prl7d1 -TG) mice. ( A ) Representative western immunoblots; ( B ) The relative expression levels of the proteins were determined using densitometric analyses of the data that were normalized against the ACTB signal ( n = 6). The protein level in the control group was set to one. Each bar represents the mean +- SD. * p < 0.01 compared to the WT group.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
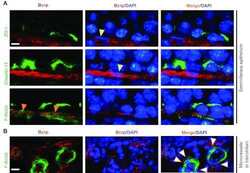
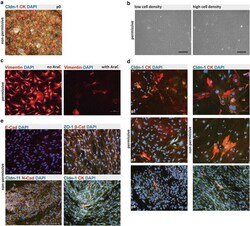
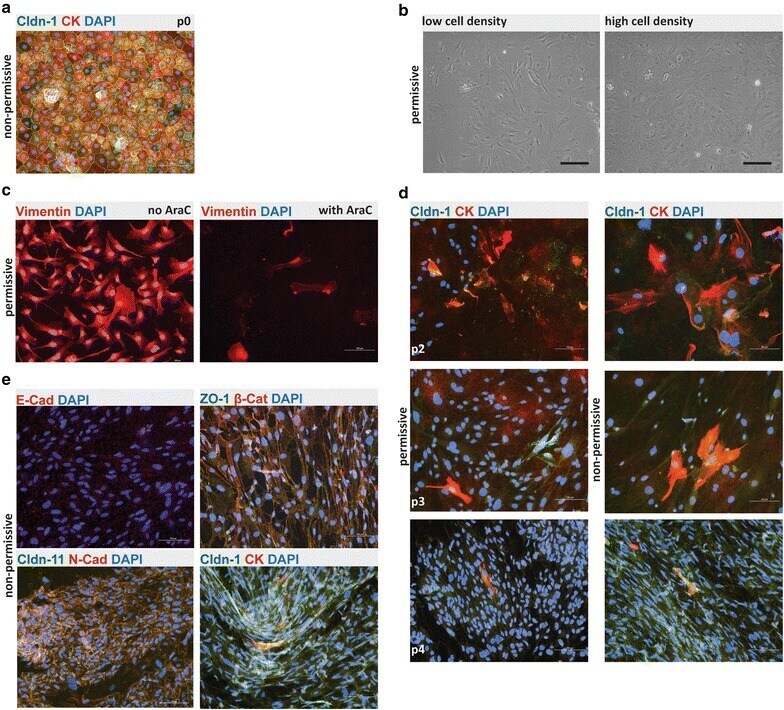
- Fig. 2 Phenotype and irreversible de-differentiation of Immortomouse (r) CPECs. a Primary Immortomouse (r) choroid plexus epithelial cells (p0) grown at 37 degC for 7 days showed the same staining patterns for cytokeratin (CK) and claudin-1 (Cldn-1) as pmCPECs isolated from wild-type mice. b When Immortomouse (r) CPECs were grown under permissive conditions (33 degC, 10U IFNgamma) the heterogenous de-differentiation process started in areas with low cell density, whereas CPECs kept their cuboidal shape longer in areas with high cell density. The contrast of the pictures in a and b was enhanced using Adobe Photoshop software. c Vimentin staining, rather than CK staining was observed at 33 degC. Cell death took place at 33 degC upon Ara-C addition to culture. d The irreversible loss of CPEC specific markers Cldn-1 and CK and an increasing proliferation rate of de-differentiated Immortomouse (r) CPECs was observed with increased passage. The first row is passage 2 (p2), the second row is p3, the third row is p4. e The cells failed to form confluent monolayers or display the correct expression pattern of epithelial markers upon shift to non-permissive temperature and IFNgamma withdrawal. Immortomouse (r) CPECs in e were stained after 7 days of non-permissive growth; the passage numbers (p) were E-cad/DAPI: p8, ZO1/beta-Cat/DAPI: p5, Cldn11/N-Cad/DAPI: p6, Cldn-1/CK/DAPI: p4. Scale bar in all immunofluorescent and phase contrast pictures = 100 mum. p0 primary culture, E-Cad E-cadh
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
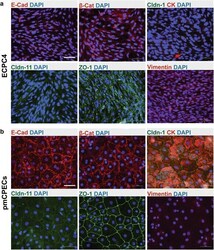
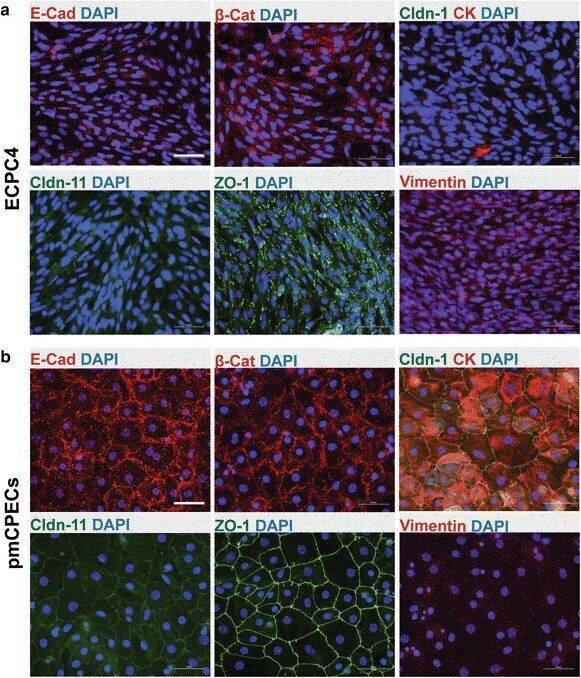
- Fig. 4 Phenotype of ECPC4 cells and primary mouse choroid plexus epithelial cells (pmCPECs). Immunofluorescence staining for CPEC specific proteins is shown in ECPC4 cells ( a ) and pmCPECs ( b ). a There is weak staining for the adhesion junction (AJ) protein E-Cadherin (E-Cad) and its cytoskeleton linker beta-catenin (beta-Cat) of ECPC4 cells and their localization is not specifically at the plasma membrane. Staining for tight junctional (TJ) claudins-1 and -11 was absent or showed a weak cytosolic pattern, respectively. The scaffolding protein ZO1 staining was disrupted. Additionally, the cell line failed to stain for the early epithelial marker cytokeratin but rather was positive for the mesenchymal intermediate filament protein vimentin. ECPC4 cells from passage 41 were stained on d4 in culture. b In contrast, the staining of pmCPECs stained on d7 in culture, revealed a proper distribution of all epithelial markers. pmCPECs. All staining was performed at least 3 times. Scale bars 50 mum. E-Cad E-cadherin, Cldn-1 Claudin 1, Cldn-11 claudin 11, CK cytokeratin, beta-Cat beta-catenin, ZO-1 zona occludens protein 1
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
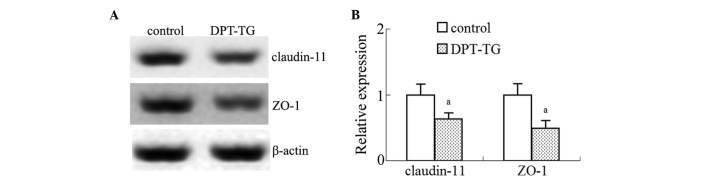
- Figure 5 Overexpression of DPT in the Sertoli cells reduces the expression levels of ZO-1 and claudin-11 in the testes. (A) Representative western blotting analyses of ZO-1 and claudin-11 in the control mice and the mice with DPT overexpression. (B) The relative expression levels of ZO-1 and claudin-11 were determined using densitometric analysis of the data normalized against the beta-actin from five different samples. The data are presented as the mean +- standard error of the mean ( a P
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
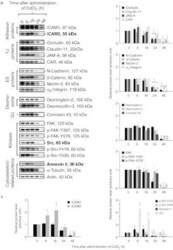
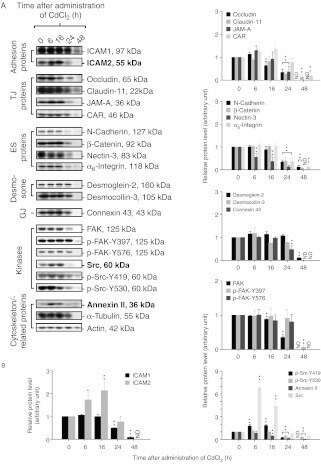
- Figure 4 ICAM2, Src, and annexin II increase during CdCl 2 -induced testis damage. Immunoblotting experiments showing changes in the levels of several proteins (A). Proteins whose levels increased following CdCl 2 treatment are labeled as bold. Actin was used as an internal control. Histograms (B) summarizing immunoblotting results. Each data point was normalized against its corresponding actin data point and then against '0 h', which was arbitrarily set at 1. Each bar represents mean+- s.d. of n =3-6 rats. * P
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
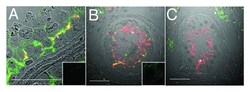
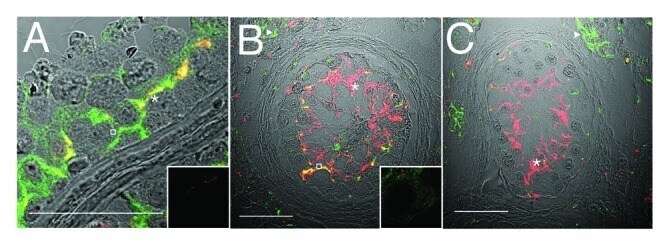
- Figure 3. Claudin-11 and Connexin 43 are disorganized in testicular dysgenesis. Confocal immunofluorescence of normal ( A ) and carcinoma in situ ( B and C ) human testis sections detecting claudin-11 (red, tight junctions, asterisks) and connexin 43 (green, gap junctions, open squares and triangles). Co-localization appears in yellow. Inserts are negative controls. (Bar = 50 um).
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
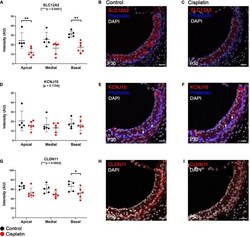
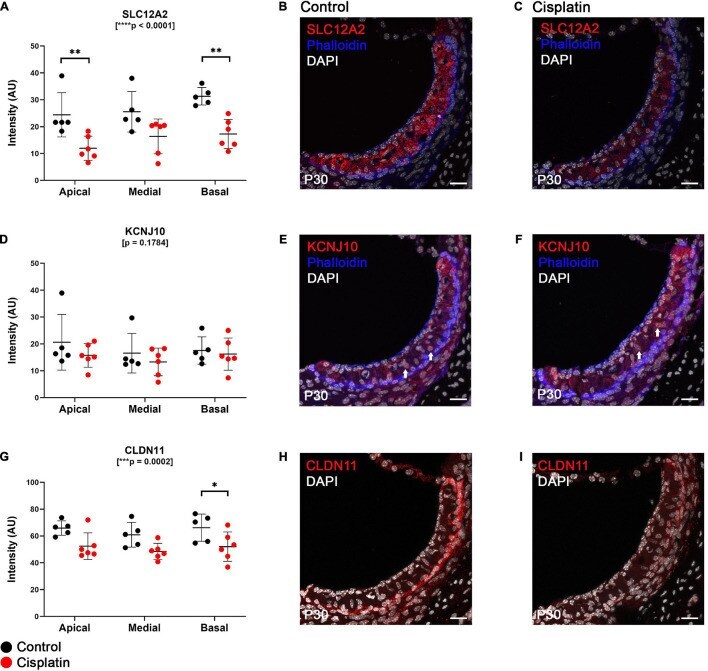
- FIGURE 2 Expression of key EP-related cell-type specific proteins is reduced in the SV after cisplatin treatment. (A) Twenty four hours after cisplatin injection, SLC12A2 immunofluorescence intensity was significantly reduced in the apical and basal turns of the cochlea (two-way ANOVA, p < 0.0001; Sidak's multiple comparisons test, q = 0.0068 and q = 0.0022, respectively). (B,C) Representative SLC12A2 immunostaining of SV cross sections of control and cisplatin-treated mice. (D) Immunofluorescence staining intensity of KCNJ10 was unchanged between control and cisplatin-treated mice (two-way ANOVA, p = 0.1784). (E,F) Representative KCNJ10 immunostaining of SV cross sections of control and cisplatin-treated mice. White arrows point to representative intermediate cell nuclei. (G) Immunofluorescence staining intensity of CLDN11 was reduced in the basal turn of cisplatin-treated compared to control mice (two-way ANOVA, p = 0.0002; Sidak's multiple comparisons test q = 0.0418). (H,I) Representative CLDN11 immunostaining of SV cross sections of control and cisplatin-treated mice. Scale bars: 20 mum. * p < 0.05; ** p < 0.01; *** p < 0.001; **** p < 0.0001.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
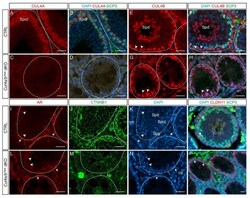
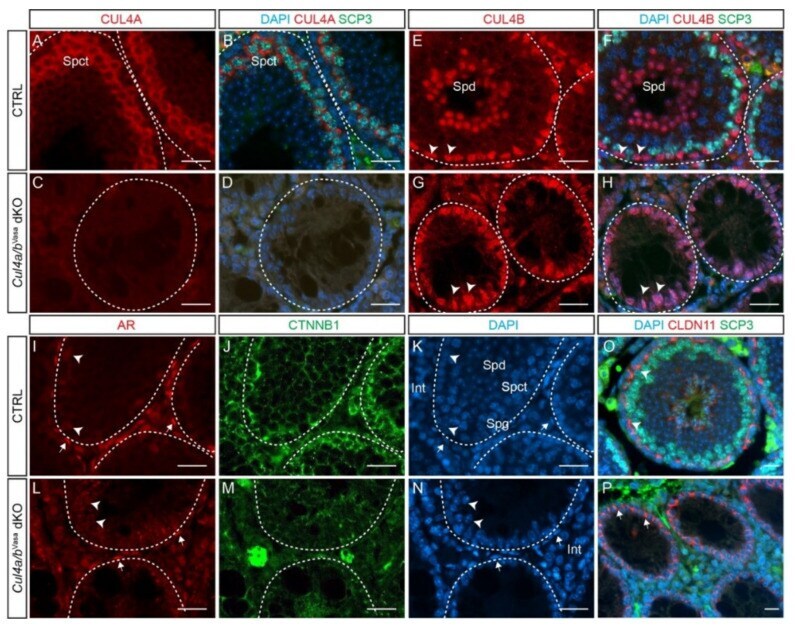
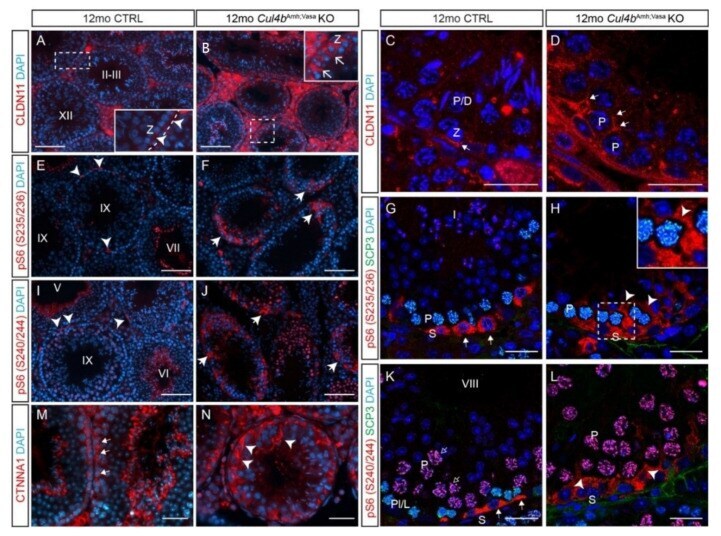
- Figure 4 Cul4 genes are required for male germ cell survival. ( A - D ) IF staining of CUL4A (red) and SCP3 (green) of P28 testis. While primary spermatocytes expressing both CUL4A ( A ) and SCP3 ( B ) are present in the CTRL testis, this population is completely missing in the dKO mutant testis ( C , D ). ( E - H ) IF staining of CUL4B (red) and SCP3 (green) of P28 testis. CUL4B is highly expressed in the Sertoli cells ( E , arrowheads) and round spermatids (Spd) of CTRL testis ( E , F ); whereas in the Cul4a/b Vasa dKO, CUL4B-positive Sertoli cells are the only cell type remaining in the seminiferous tubules ( G , H arrowheads). ( I - N ) IF staining of AR (red) and CTNNB1 (green) of P28 testicular sections. Wild-type testis show AR-high myoid cells ( I , arrows) and AR-low Sertoli cells ( I , arrowheads), and well organized germ cell/Sertoli cell network outlined by CTNNB1 staining ( J ). The dKO mutant testis retains myoid and Sertoli cells ( L , arrows and arrowheads, respectively), with disorganized residual CTNNB1 staining ( M ). DAPI counterstain is shown in ( K ) and ( N ). Spd, spermatids; Spct, spermatocytes; Spg, spermatogonia; Int, interstitial cells. ( O , P ) IF staining of CLDN11 (red) and SCP3 (green) of P28 testis. At this stage in the wildtype testis, tight junctions have formed apical to the spermatogonial stem cells to separate the basal and adluminal compartments ( O , arrowheads). CLDN11 was only detected in the basal-lateral membranes of dKO Sertoli ce
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
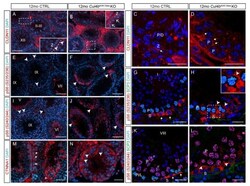
- Figure 6 Ablation of Cul4b in both germ cells and Sertoli cells leads to BTB defects. ( A , B ) IF staining and ( C , D ) confocal microscopy of tight junction marker CLDN11 in CTRL and Cul4b Amh;Vasa testis. The insets in A and B are magnified views of boxed areas. Basement membrane outlined by dashed lines in insets. ( E , F ) IF staining of pS6 (S235/236) showing its accumulation in the mutant tubules. ( G , H ) Confocal IF of pS6 (S235/236) (red) and SCP3 (green) showing localization of pS6 (S235/236) in CTRL spermatogonia ( G , arrows), and ectopic activation in the mutant Sertoli cells ( H , arrowheads). ( I , J ) IF staining of pS6 (S240/244) showing its accumulation in the mutant tubules. ( K , L ) Confocal IF of pS6(S240/244) (red) and SCP3 (green) showing its normal expression in spermatogonia ( K , arrows) and ectopic activation in the mutant germ cells ( L , arrowheads). ( M , N ) IF of alpha-catenin (CTNNA1) showing its accumulation in the mutant testis. S, spermatogonia; P, pachytene spermatocytes; Z, zygotene spermatocytes; Spg,. White dashed lines outline the seminiferous tubules. Scale bars: 200 um in ( A , B ), ( E , F ), ( I , J ); 50 um in the rest.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
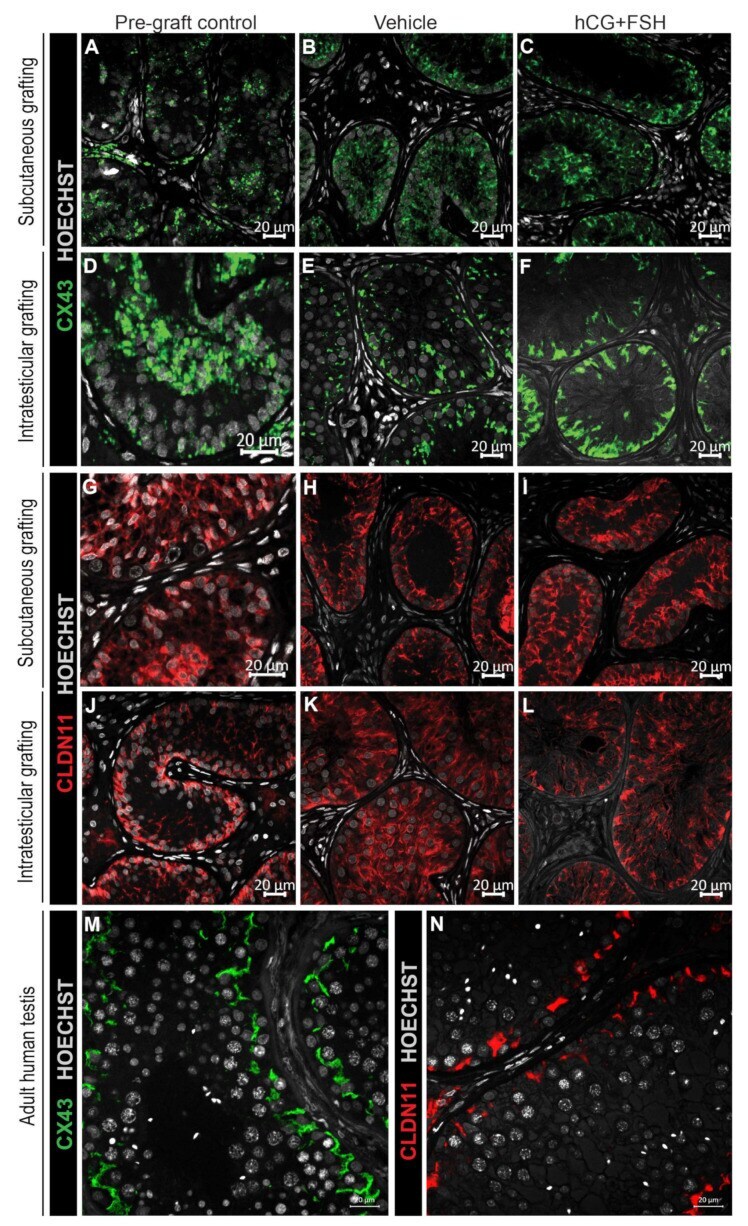
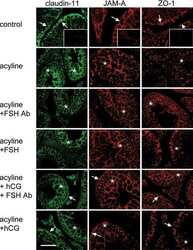
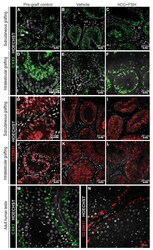
- Figure 6 Expression of CX43 and CLDN11 in prepubertal human testis grafts. ( A - C ) Immature CX43 expression profile in subcutaneous grafts (pre-graft control; 1-year-old). ( D - F ) Partially mature CX43 expression profile in intratesticular grafts (pre-graft control; 14-year-old). ( G - I ) Immature CLDN11 expression pattern in subcutaneous grafts (pre-graft control; 1-year-old). ( J ) Partially mature CLDN11 expression pattern (pre-graft control; 14-year-old). ( K , L ) Immature CLDN11 expression pattern in intratesticular grafts. ( M , N ) Expression of CX43 and CLDN11 in adult human testis tissue.