Explore
Explore Validate
Validate Learn
Learn Western blot
Western blot ELISA
ELISA Other assay
Other assayAntibody data
- Antibody Data
- Antigen structure
- References [13]
- Comments [0]
- Validations
- Other assay [8]
Submit
Validation data
Reference
Comment
Report error
- Product number
- 33-5000 - Provider product page

- Provider
- Invitrogen Antibodies
- Product name
- GAP43 Monoclonal Antibody (7B10)
- Antibody type
- Monoclonal
- Antigen
- Purifed from natural sources
- Description
- This antibody can recognize both the phosphorylated and unphosphorylated target.
- Reactivity
- Human, Mouse, Rat, Chicken/Avian, Rabbit
- Host
- Mouse
- Isotype
- IgG
- Antibody clone number
- 7B10
- Vial size
- 100 μg
- Concentration
- 0.5 mg/mL
- Storage
- -20°C
Submitted references Effects of amyloid precursor protein peptide APP96-110, alone or with human mesenchymal stromal cells, on recovery after spinal cord injury.
Optimised techniques for high-throughput screening of differentiated SH-SY5Y cells and application for neurite outgrowth assays.
Overexpression of Reticulon 3 Enhances CNS Axon Regeneration and Functional Recovery after Traumatic Injury.
The superiority of conditioned medium derived from rapidly expanded mesenchymal stem cells for neural repair.
Rescue of the genetically engineered Cul4b mutant mouse as a potential model for human X-linked mental retardation.
Microglial involvement in neuroplastic changes following focal brain ischemia in rats.
Fezf1 is required for penetration of the basal lamina by olfactory axons to promote olfactory development.
Localization of adenylate kinase 4 in mouse tissues.
Gradual nerve elongation affects nerve cell bodies and neuro-muscular junctions.
Gene discovery in genetically labeled single dopaminergic neurons of the retina.
In vitro development of mouse embryonic stem cells lacking JNK/stress-activated protein kinase-associated protein 1 (JSAP1) scaffold protein revealed its requirement during early embryonic neurogenesis.
Recovery from spinal cord injury: a new transection model in the C57Bl/6 mouse.
Binding of the Ras activator son of sevenless to insulin receptor substrate-1 signaling complexes.
Hodgetts SI, Lovett SJ, Baron-Heeris D, Fogliani A, Sturm M, Van den Heuvel C, Harvey AR
Neural regeneration research 2022 Jun;17(6):1376-1386
Neural regeneration research 2022 Jun;17(6):1376-1386
Optimised techniques for high-throughput screening of differentiated SH-SY5Y cells and application for neurite outgrowth assays.
Dravid A, Raos B, Svirskis D, O'Carroll SJ
Scientific reports 2021 Dec 14;11(1):23935
Scientific reports 2021 Dec 14;11(1):23935
Overexpression of Reticulon 3 Enhances CNS Axon Regeneration and Functional Recovery after Traumatic Injury.
Alhajlah S, Thompson AM, Ahmed Z
Cells 2021 Aug 6;10(8)
Cells 2021 Aug 6;10(8)
The superiority of conditioned medium derived from rapidly expanded mesenchymal stem cells for neural repair.
Chen YT, Tsai MJ, Hsieh N, Lo MJ, Lee MJ, Cheng H, Huang WC
Stem cell research & therapy 2019 Dec 16;10(1):390
Stem cell research & therapy 2019 Dec 16;10(1):390
Rescue of the genetically engineered Cul4b mutant mouse as a potential model for human X-linked mental retardation.
Chen CY, Tsai MS, Lin CY, Yu IS, Chen YT, Lin SR, Juan LW, Chen YT, Hsu HM, Lee LJ, Lin SW
Human molecular genetics 2012 Oct 1;21(19):4270-85
Human molecular genetics 2012 Oct 1;21(19):4270-85
Microglial involvement in neuroplastic changes following focal brain ischemia in rats.
Madinier A, Bertrand N, Mossiat C, Prigent-Tessier A, Beley A, Marie C, Garnier P
PloS one 2009 Dec 1;4(12):e8101
PloS one 2009 Dec 1;4(12):e8101
Fezf1 is required for penetration of the basal lamina by olfactory axons to promote olfactory development.
Watanabe Y, Inoue K, Okuyama-Yamamoto A, Nakai N, Nakatani J, Nibu K, Sato N, Iiboshi Y, Yusa K, Kondoh G, Takeda J, Terashima T, Takumi T
The Journal of comparative neurology 2009 Aug 10;515(5):565-84
The Journal of comparative neurology 2009 Aug 10;515(5):565-84
Localization of adenylate kinase 4 in mouse tissues.
Miyoshi K, Akazawa Y, Horiguchi T, Noma T
Acta histochemica et cytochemica 2009 Apr 28;42(2):55-64
Acta histochemica et cytochemica 2009 Apr 28;42(2):55-64
Gradual nerve elongation affects nerve cell bodies and neuro-muscular junctions.
Kazuo Ikeda KI, Masaki Matsuda MM, Daisuke Yamauchi DY, Katsuro Tomita KT, Shigenori Tanaka ST
Hand surgery : an international journal devoted to hand and upper limb surgery and related research : journal of the Asia-Pacific Federation of Societies for Surgery of the Hand 2005 Jul;10(1):7-15
Hand surgery : an international journal devoted to hand and upper limb surgery and related research : journal of the Asia-Pacific Federation of Societies for Surgery of the Hand 2005 Jul;10(1):7-15
Gene discovery in genetically labeled single dopaminergic neurons of the retina.
Gustincich S, Contini M, Gariboldi M, Puopolo M, Kadota K, Bono H, LeMieux J, Walsh P, Carninci P, Hayashizaki Y, Okazaki Y, Raviola E
Proceedings of the National Academy of Sciences of the United States of America 2004 Apr 6;101(14):5069-74
Proceedings of the National Academy of Sciences of the United States of America 2004 Apr 6;101(14):5069-74
In vitro development of mouse embryonic stem cells lacking JNK/stress-activated protein kinase-associated protein 1 (JSAP1) scaffold protein revealed its requirement during early embryonic neurogenesis.
Xu P, Yoshioka K, Yoshimura D, Tominaga Y, Nishioka T, Ito M, Nakabeppu Y
The Journal of biological chemistry 2003 Nov 28;278(48):48422-33
The Journal of biological chemistry 2003 Nov 28;278(48):48422-33
Recovery from spinal cord injury: a new transection model in the C57Bl/6 mouse.
Seitz A, Aglow E, Heber-Katz E
Journal of neuroscience research 2002 Feb 1;67(3):337-45
Journal of neuroscience research 2002 Feb 1;67(3):337-45
Binding of the Ras activator son of sevenless to insulin receptor substrate-1 signaling complexes.
Baltensperger K, Kozma LM, Cherniack AD, Klarlund JK, Chawla A, Banerjee U, Czech MP
Science (New York, N.Y.) 1993 Jun 25;260(5116):1950-2
Science (New York, N.Y.) 1993 Jun 25;260(5116):1950-2
No comments: Submit comment
Supportive validation
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
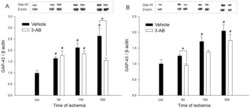
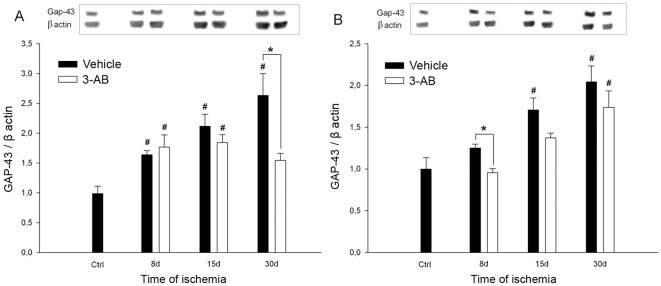
- Figure 6 Time course of GAP-43 expression after ischemia: effect of 3-AB treatment. Western blots were prepared after 8 d, 15 d and 30 d of ischemia from P1 (A) and P2 (B) samples. Representative immunoblots are shown and densitometric analysis was performed after normalization of GAP-43 expression on beta-actin levels. Photothrombotic ischemia induces an increase in GAP-43 expression from 8 d of ischemia in P1 and 15 d in P2. 3-AB treatment repressed GAP-43 expression in P1 at 30 d. In P2, the overall GAP-43 expression was lower in 3-AB treated animals but a significant decrease was found at 8 d only. Values are expressed as means+-S.E.M. and are representative of 5 to 6 animals. ( # P
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
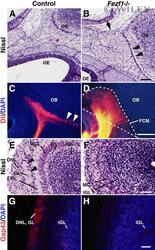
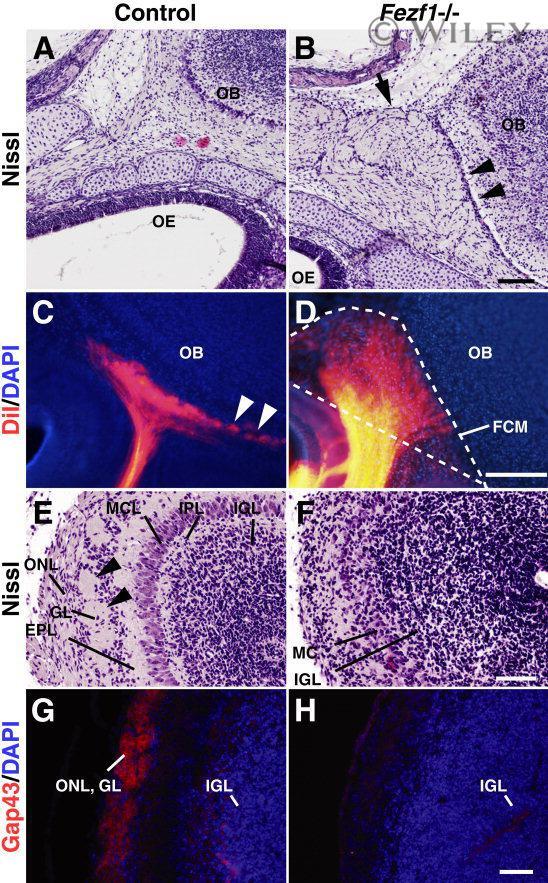
- 3 Defects of olfactory axonal connectivity in Fezf1 -deficient mice at P0. A - D : Parasagittal sections of the heads of control (A,C) and Fezf1 -deficient mice (B,D) at P0. Anterior is to the left and posterior is to the right. Nissl-stained sections (A,B) show a disconnected space (arrowheads in B) between the OB and fibrocellular mass (FCM) (arrow in B) in the Fezf1 -deficient mice. Placement of DiI in the OE of aldehyde-fixed mice resulted in postmortem labeling of olfactory axons (orange; C,D). Sections are counterstained with DAPI (blue; C,D). Olfactory axons enter the OB and form glomeruli (arrowheads in C) in control. However, they terminate in the FCM at a space between the cribriform plate and OB in Fezf1 -deficient mice (D). The FCM is outlined by a dotted line in D. E - H : Coronal sections of the control (E,G) and Fezf1 -deficient (F,H) OB. Nissl-stained sections show the absence of glomeruli (arrowheads in E) in the Fezf1 -deficient OB (F). G,H: Immunostaining of olfactory axons using anti-Gap43 antibody (red) in coronal sections of OB, showing olfactory nerve and glomerular layer in control (G). Sections are counterstained with DAPI (blue). Olfactory nerve layer and glomerular layer are not formed in the Fezf1 -deficient OB (F,H). ONL, olfactory nerve layer; GL, glomerular layer; EPL, external plexiform layer; MCL, mitral cell layer; IPL, internal plexiform layer; IGL, internal granular layer. Scale bars = 100 mum in B (applies to A); 200 mum in D (applies
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
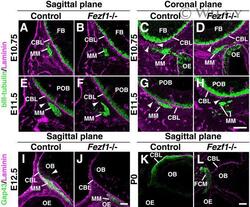
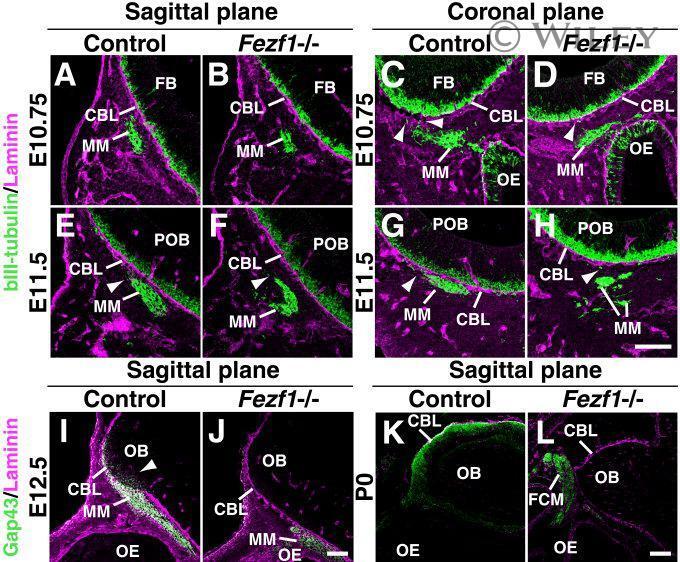
- 8 Pioneer olfactory axons access but do not penetrate the CNS basal lamina in Fezf1 -deficient mice. A - L : Immunofluorescence analyses of olfactory axonal projections at E10.75 (A-D), E11.5 (E-H), E12.5 (I,J), and P0 (K,L). Parasagittal (A,B,E,F,I-L) and coronal (C,D,G,H) sections of control (Control; A,C,E,G,I,K) and Fezf1 -deficient ( Fezf1 -/-; B,D,F,H,J,L) mice were analyzed. In the parasagittal sections (A,B,E,F,I-L), anterior is to the left and posterior is to the right. In the coronal sections (C,D,G,H), ventral is to the left and lateral is to the right. Olfactory axons (green signals in the migratory mass (MM) in A-H) are stained with anti-betaIII-tubulin antibody at E10.75 and E11.5. At E12.5 and P0, olfactory axons are stained with the anti-Gap43 antibody (green or white signals in I-L). The basal lamina is stained in magenta with the anti-laminin antibody (A-L). At E10.75, while several olfactory axons access the CNS basal lamina (CBL) in the controls (arrowheads in C), only a few of them access the CBL in Fezf1 -deficient mice (arrowhead in D). Note that in the parasagittal section the MM is localized further away from the CBL in Fezf1 -deficient mice (B) relative to the control (A). At E11.5 the MM is attached to the CBL in the control (E,G), whereas it is not in Fezf1 -deficient mice (F,H). However, some axons attach to the CBL in Fezf1 -deficient mice (arrowhead in H). The MM is disintegrated in Fezf1 -deficient mice at E11.5 (F,H). Pioneer axons of the
- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
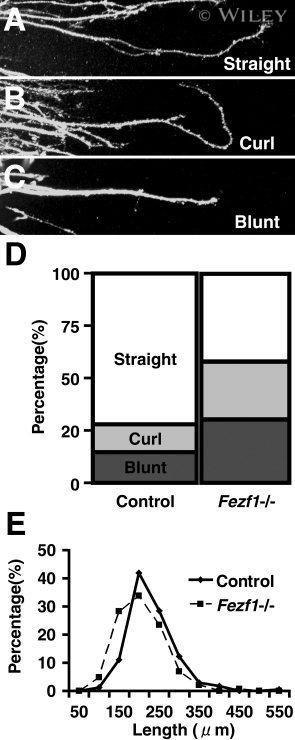
- 9 Abnormal phenotype of axonal elongation in the Fezf1 -deficient ORNs. A - C : Axonal shapes of 2-day OE culture in 80% Matrigel basement matrix. Axons are immunostained with the anti-Gap43 antibody. Elongation pattern of olfactory axons are classified into three types, i.e., straight (A), curl (B), and blunt (C). The ratio of curl and blunt types of axonal elongation is higher in Fezf1 -deficient mice (n = 124 explants) compared with their normal counterparts (n = 146) ( D ). Length of axons (mean +- SEM) elongating from the explant tissue is shorter in Fezf1 -deficient mice (178.94 +- 4.75 mum, n = 145 axons) than that in the control (207.77 +- 4.48 mum, n = 172) ( E ).
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
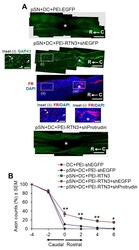
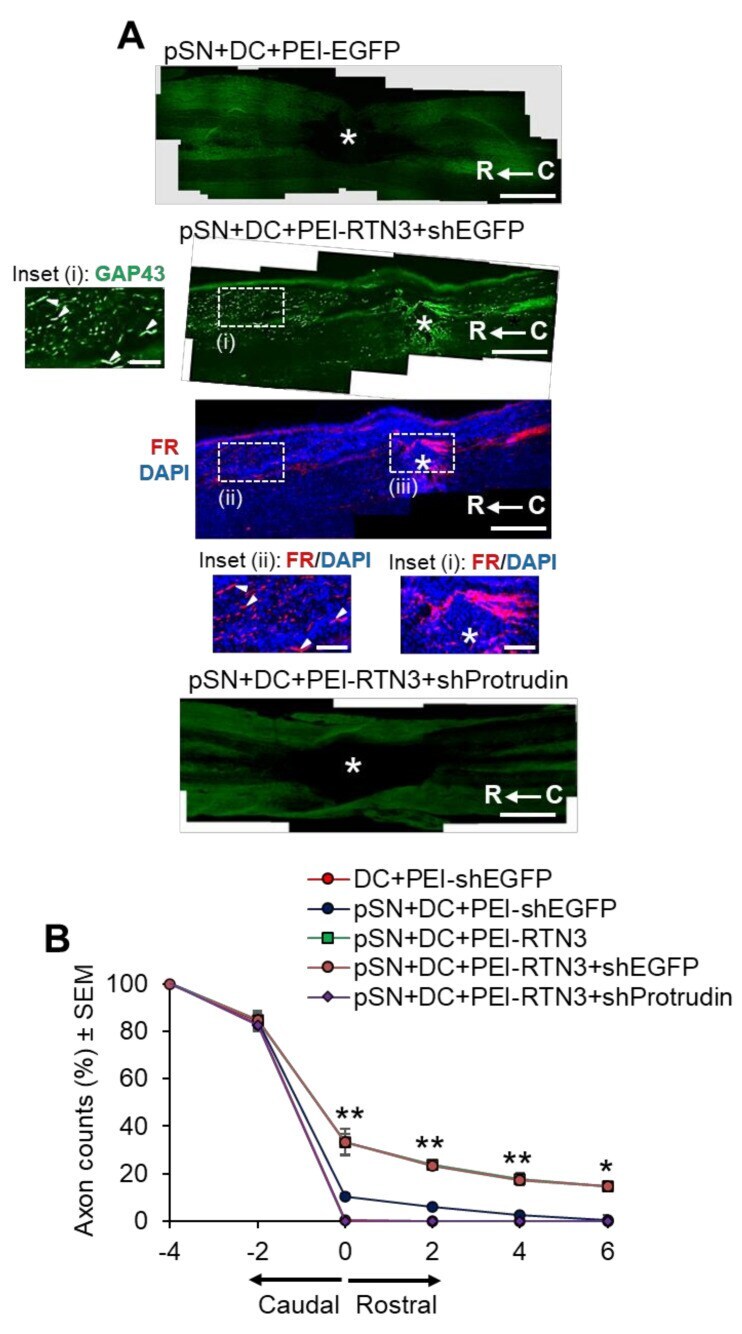
- Figure 6 RTN3 overexpression promotes Protrudin-dependent DC axon regeneration. ( A ) Representative longitudinal sections of the spinal cord immunostained with GAP43 antibodies after treatment with pSN + DC + PEI-EGFP, pSN + DC + PEI-RTN3 + shEGFP and pSN + DC + PEI-RTN3 + shProtrudin (* = lesion site; C to R = caudal to rostral). FluoroRuby (FR) traced axons in the same sections as pSN + DC + PEI-RTN3 + shEGFP confirms overlap of GAP43 + axons (arrowheads), unequivocally demonstrating axon regeneration. Inset (i) shows high power images of GAP43 + axons in the boxed region (i). Inset (ii) and (iii) show high power images of FR + axons in boxed regions (ii) and (iii). Note the overlap (arrowheads) between GAP43 + axons in inset (i) and FR+ axons in inset (ii). ( B ) Quantification of the % of GAP43 + axons at different distances caudal and rostral to the lesion site. Scale bars in ( A ) = 200 um. n = 18 rats/group. * p = 0.05, ** p = 0.001, one-way ANOVA with Dunnett's post hoc test.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
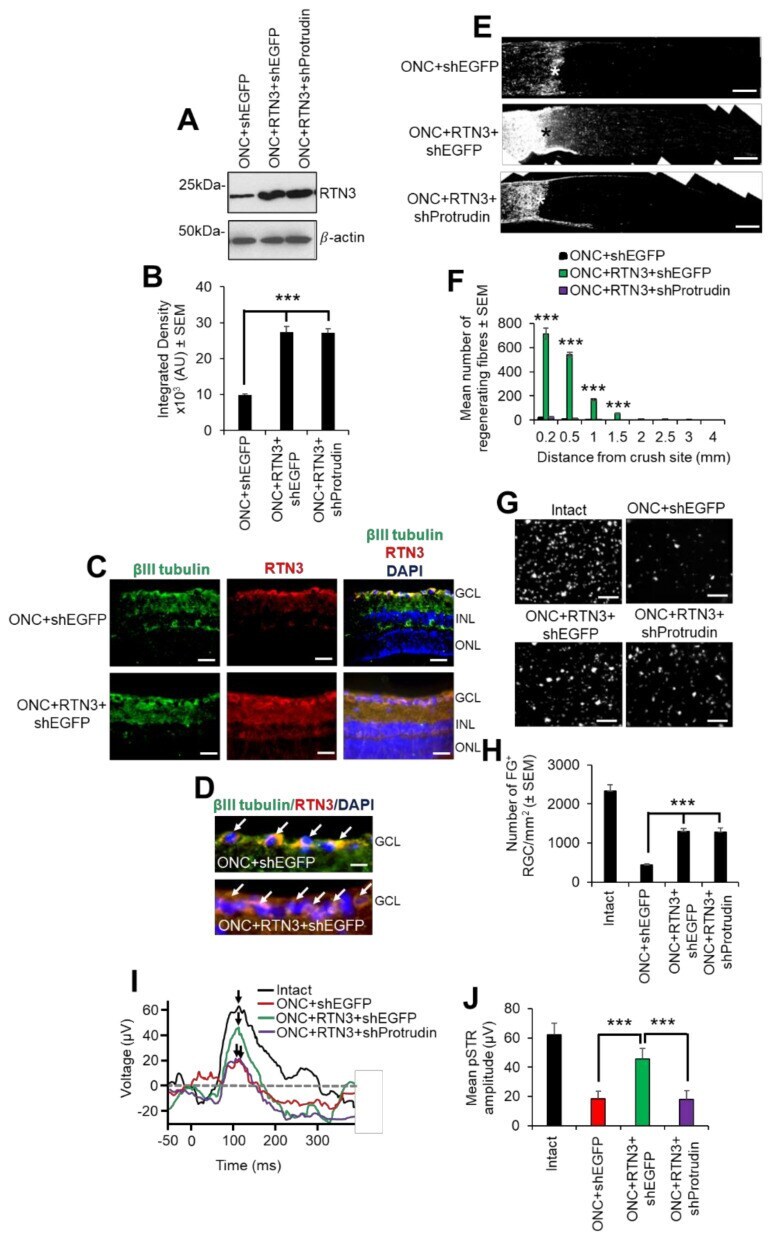
- Figure 8 RTN3 overexpression promotes RGC axon regeneration and functional recovery after ONC, which is also protrudin-dependent. ( A ) Western blot and ( B ) quantification to show that RTN3 plasmids significantly upregulate RTN3 protein in the retina after intravitreal injection in vivo. ( C ) Immunohistochemistry to localise RTN3 protein to the RGCs (green) in the ganglion cell layer (GCL) and the inner plexiform and inner nuclear layer (INL). ONL = outer nuclear layer. ( D ) High power images to show RTN3 protein localised to betaIII-tubulin + RGCs (arrows) in the GCL. ( E ) GAP43 immunohistochemistry and ( F ) quantification to show that in control optic nerve, few if any GAP43 + axons pass beyond the lesion site (*) whilst in RTN3 overexpressed eyes, significant GAP43 + axons are present beyond the lesion site, an effect which is obliterated when Protrudin is knocked out at the same time as RTN3 overexpression. ( G ) Representative images to show FluorGold (FG) backfilled RGC in retinal wholemounts after RTN3 overexpression. ( H ) Quantification of the number of FG + RGC in retinal wholemounts shows that overexpression of RTN3 is significantly neuroprotective, an effect that is independent of protrudin. ( I ) Representative ERG traces and ( J ) quantification of the pSTR amplitude shows significant improvements in RTN3 overexpressed eyes which are ablated after protrudin knockdown. Arrows show the peak of the pSTR. Data are means +- SEM. n = 12 eyes/optic nerves/treatme
- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
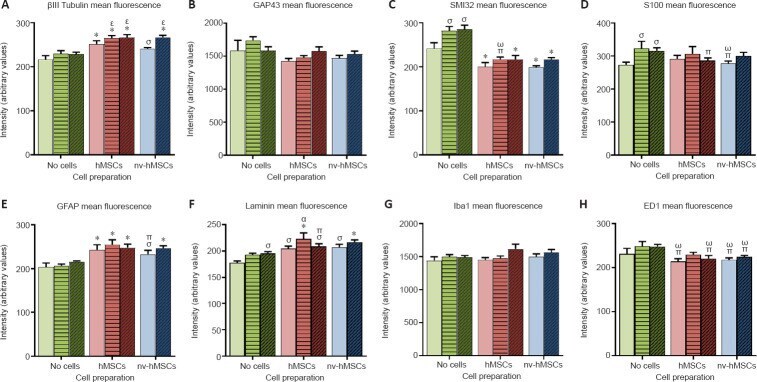
- Figure 6 Mean fluorescence intensities of various immunostained sections . Cell transplantation (hMSCs, red bars; nv-hMSCs, blue bars) affected fluorescence intensity more than treatment with mAPP96-110 (+- viable/nv-hMSCs; horizontal striped bars) or APP96-110 (+- viable/nv-hMSCs; diagonal striped bars). Cell transplantation significantly increased the mean fluorescence intensity of betaIII tubulin (A), GFAP (E) and laminin (F), decreased SMI32 (C), S100 (D; APP96-110 + hMSCs and nv-hMSCs only) and ED1 (H), and had no effect on GAP43 (B) or Iba1 (G) fluorescence intensity. mAPP96-110 and APP96-110 significantly increased the mean fluorescence intensity of SMI32 (C), S100 (D) and laminin (F; APP96-110 only). Significance to SCI, mAPP, APP (*), SCI (sigma), mAPP96-110 (pi), APP96-110 (omega), hMSCs (alpha) and nv-hMSCs (epsilon); ( P < 0.05, two-way analysis of variance followed by Fisher's least significant difference test). Data are expressed as the mean +- SEM. APP96-110: Amyloid precursor protein peptide; GAP43: growth associated protein-43; GFAP: glial fibrillary acidic protein; hMSCs: human mesenchymal stromal cells; mAPP96-100: mutant amyloid precursor protein; nv: non-viable.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
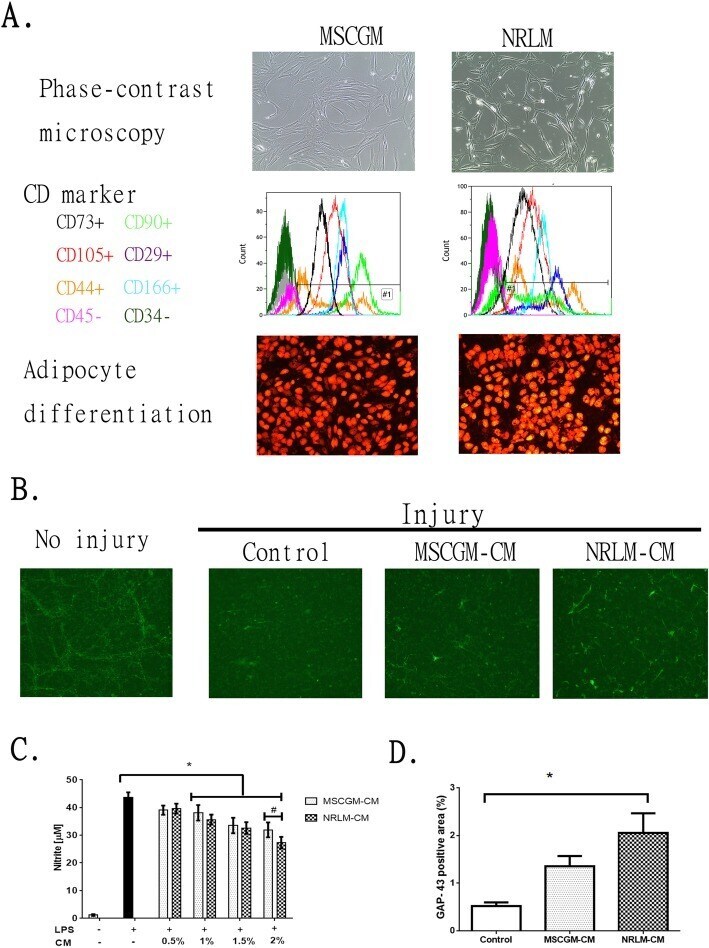
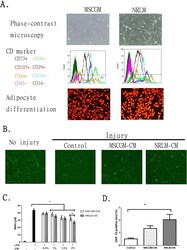
- Fig. 1 Comparative characterization of commercial mesenchymal stem cells (MSCs) cultured in MSCGM or NRLM and the function of the conditioned medium (CM). a MSC characteristics, including morphology (phase-contrast micrograph, upper panel), cell surface expression of CD markers (by flow cytometry, middle panel), and adipocyte differentiation (oil-positive signal after oil staining, lower panel), are shown. b The effect of MSCGM-CM and NRLM-CM on the extension of regenerated neurites (GAP-43-positive neurite). c Quantitative results from b . d The effect of MSCGM-CM and NRLM-CM on LPS stimulation in BV2 microglia. Nitrite release in the medium was measured after microglia were treated with LPS in the presence of CM (0-2% v/v). Data represent the mean +- SEM. * p < 0.05 indicates statistical significance using one-way ANOVA with Tukey's multiple comparisons test