Explore
Explore Validate
Validate Learn
Learn Flow cytometry
Flow cytometryAntibody data
- Antibody Data
- Antigen structure
- References [42]
- Comments [0]
- Validations
- Flow cytometry [1]
- Other assay [36]
Submit
Validation data
Reference
Comment
Report error
- Product number
- 16-9917-025 - Provider product page

- Provider
- Invitrogen Antibodies
- Product name
- CD284 (TLR4) Monoclonal Antibody (HTA125), Functional Grade, eBioscience™
- Antibody type
- Monoclonal
- Antigen
- Other
- Description
- Description: The HTA125 monoclonal antibody reacts with human Toll-like receptor 4 (TLR4). So far, at least ten members of the Toll family have been identified in humans. This family of type I transmembrane proteins is characterized by an extracellular domain with leucine-rich repeats and a cytoplasmic domain with homology to the type I IL-1 receptor. Two of these receptors, TLR2 and TLR4, are pattern recognition receptors and signaling molecules in response to bacterial lipoproteins and have been implicated in innate immunity and inflammation. TLR4 physically associates with another molecule called MD-2, and together with CD14, this complex is responsible for LPS recognition and signaling. TLR4 is expressed by peripheral blood monocytes. HTA125 has been reported to immunoprecipitate human TLR4 (~100 kDa) from transfected cells. Most TLR cell surface expression, especially TLR1 and TLR4, occurs at low levels on monocytes and at even lower levels on other cell types including granulocytes and immature dendritic cells (iDC). Furthermore, a relatively high degree of variability in TLR surface expression has been reported among normal donors.
- Antibody clone number
- HTA125
- Concentration
- 1 mg/mL
Submitted references Mechanical Compression by Simulating Orthodontic Tooth Movement in an In Vitro Model Modulates Phosphorylation of AKT and MAPKs via TLR4 in Human Periodontal Ligament Cells.
Helicobacter pylori Neutrophil-Activating Protein Directly Interacts with and Activates Toll-like Receptor 2 to Induce the Secretion of Interleukin-8 from Neutrophils and ATRA-Induced Differentiated HL-60 Cells.
Lys694Arg polymorphism leads to blunted responses to LPS by interfering TLR4 with recruitment of MyD88.
Inflammatory signaling in dengue-infected platelets requires translation and secretion of nonstructural protein 1.
Bacterial lipopolysaccharide and antimicrobial LL-37 enhance ICAM-1 expression and NF-κB p65 phosphorylation in senescent endothelial cells.
Bushfire smoke is pro-inflammatory and suppresses macrophage phagocytic function.
Butyrate upregulates the TLR4 expression and the phosphorylation of MAPKs and NK-κB in colon cancer cell in vitro.
Toll‑Like receptor 4 promotes the phosphorylation of CRMP2 via the activation of Rho‑kinase in MCAO rats.
n-butanol extract from Folium isatidis inhibits the lipopolysaccharide-induced downregulation of CXCR1 and CXCR2 on human neutrophils.
Altered Toll-like receptor expression and function in HPV-associated oropharyngeal carcinoma.
Platelet proteome reveals novel pathways of platelet activation and platelet-mediated immunoregulation in dengue.
Suppression of αvβ6 Integrin Expression by Polymicrobial Oral Biofilms in Gingival Epithelial Cells.
Comparative miRNA-Based Fingerprinting Reveals Biological Differences in Human Olfactory Mucosa- and Bone-Marrow-Derived Mesenchymal Stromal Cells.
Picfeltarraenin IA inhibits lipopolysaccharide-induced inflammatory cytokine production by the nuclear factor-κB pathway in human pulmonary epithelial A549 cells.
Lipopolysaccharide induces SBD-1 expression via the P38 MAPK signaling pathway in ovine oviduct epithelial cells.
Toll-Like Receptor Activation by Generalized Modules for Membrane Antigens from Lipid A Mutants of Salmonella enterica Serovars Typhimurium and Enteritidis.
TLR4 Expression Is Associated with Left Ventricular Dysfunction in Patients Undergoing Coronary Artery Bypass Surgery.
Biologically active polymers from spontaneous carotenoid oxidation: a new frontier in carotenoid activity.
Biomechanical loading modulates proinflammatory and bone resorptive mediators in bacterial-stimulated PDL cells.
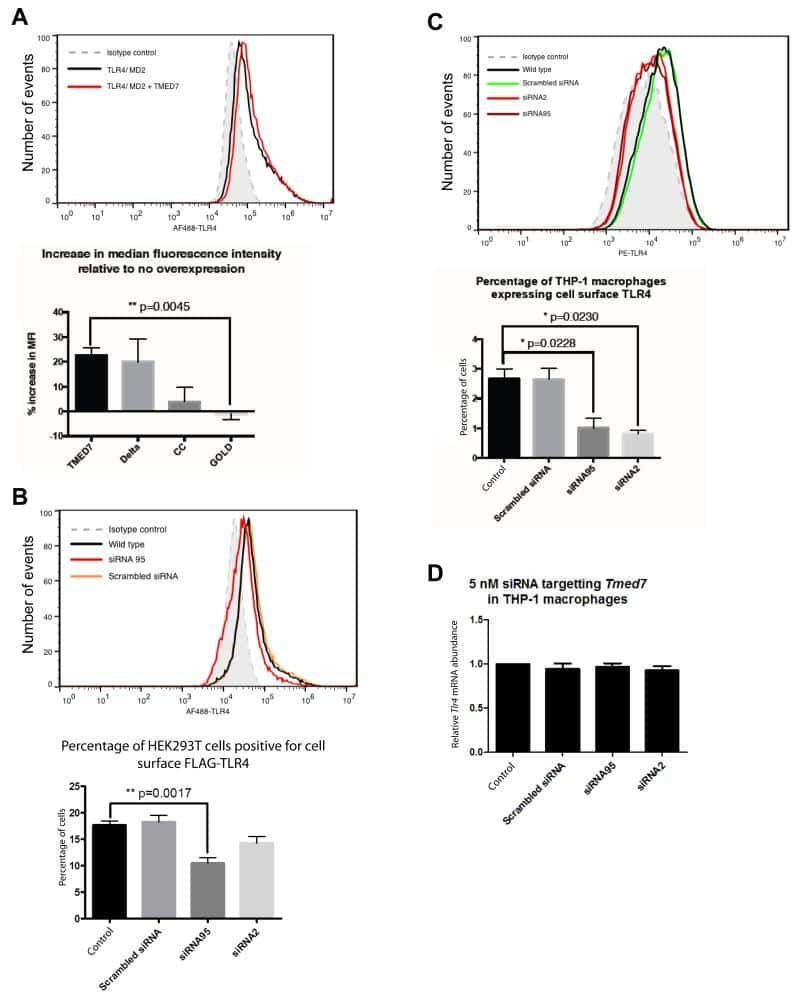
The COP II adaptor protein TMED7 is required to initiate and mediate the delivery of TLR4 to the plasma membrane.
HIV-1 gp120 induces TLR2- and TLR4-mediated innate immune activation in human female genital epithelium.
Body fluid exosomes promote secretion of inflammatory cytokines in monocytic cells via Toll-like receptor signaling.
Impaired clearance of apoptotic cells leads to HMGB1 release in the bone marrow of patients with myelodysplastic syndromes and induces TLR4-mediated cytokine production.
Monocytes from tuberculosis patients that exhibit cleaved caspase 9 and denaturalized cytochrome c are more susceptible to death mediated by Toll-like receptor 2.
Late repression of NF-κB activity by invasive but not non-invasive meningococcal isolates is required to display apoptosis of epithelial cells.
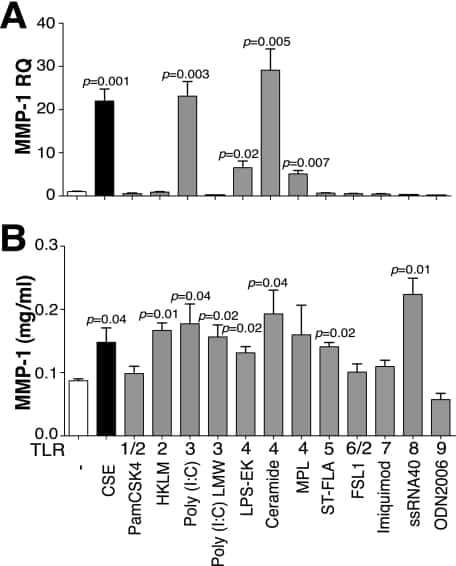
TLR4 protein contributes to cigarette smoke-induced matrix metalloproteinase-1 (MMP-1) expression in chronic obstructive pulmonary disease.
Donor Toll-like receptor 4 contributes to ischemia and reperfusion injury following human kidney transplantation.
Aggregation and microparticle production through toll-like receptor 4 activation in platelets from recently menopausal women.
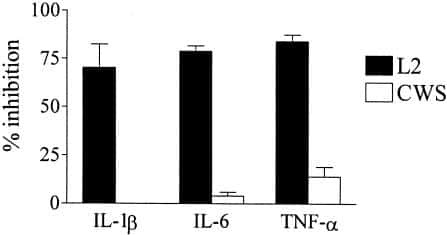
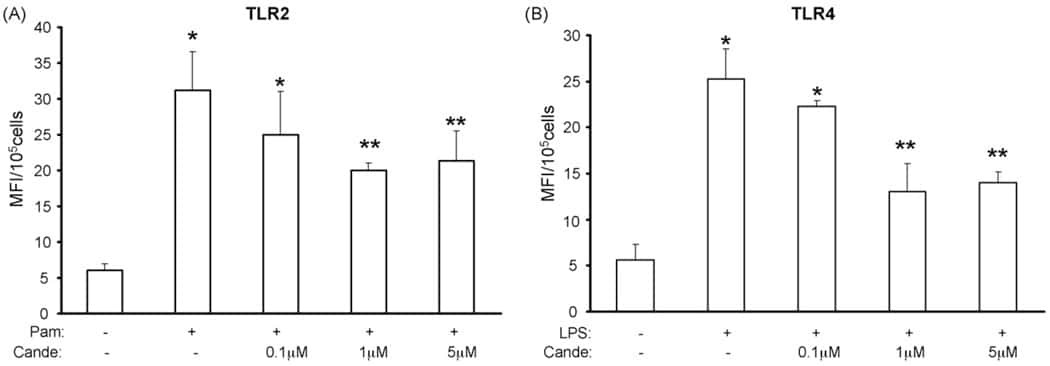
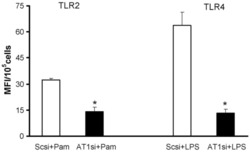
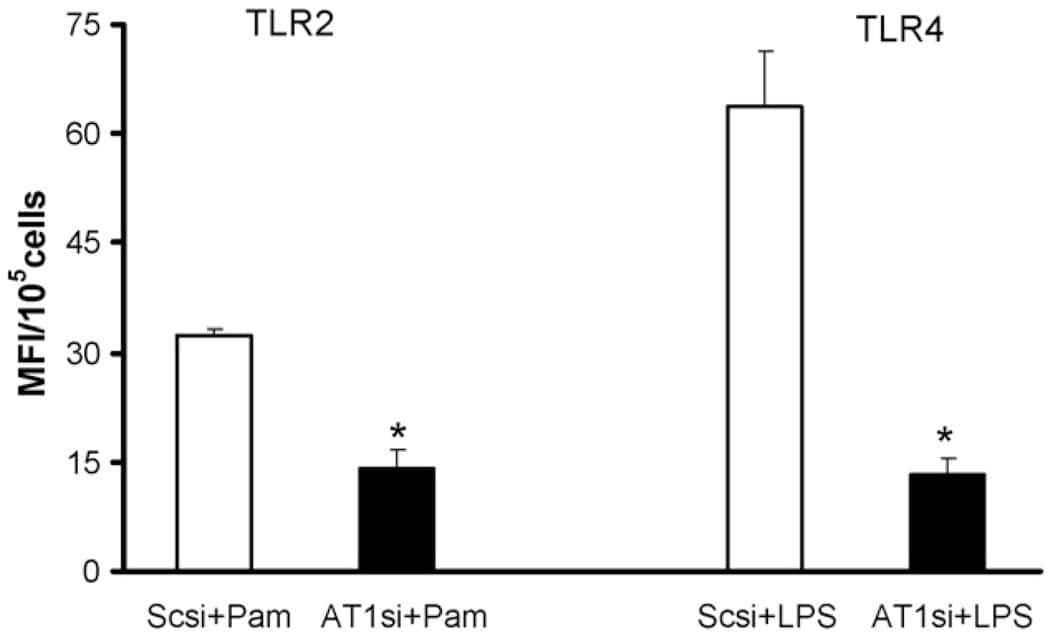
Candesartan inhibits Toll-like receptor expression and activity both in vitro and in vivo.
High-mobility group box 1 protein induces tissue factor expression in vascular endothelial cells via activation of NF-kappaB and Egr-1.
Mycobacterium tuberculosis-induced gamma interferon production by natural killer cells requires cross talk with antigen-presenting cells involving Toll-like receptors 2 and 4 and the mannose receptor in tuberculous pleurisy.
Human heat shock protein 60 stimulates vascular smooth muscle cell proliferation through Toll-like receptors 2 and 4.
Role of dendritic cell-derived CXCL13 in the pathogenesis of Bartonella henselae B-rich granuloma.
The uptake of a Klebsiella pneumoniae capsule polysaccharide mutant triggers an inflammatory response by human airway epithelial cells.
Respiratory epithelial cells require Toll-like receptor 4 for induction of human beta-defensin 2 by lipopolysaccharide.
Distinct strains of Propionibacterium acnes induce selective human beta-defensin-2 and interleukin-8 expression in human keratinocytes through toll-like receptors.
Surface calreticulin mediates muramyl dipeptide-induced apoptosis in RK13 cells.
TLR9 is localized in the endoplasmic reticulum prior to stimulation.
Differential regulation of inflammatory cytokine secretion by human dendritic cells upon Chlamydia trachomatis infection.
Expression and involvement of Toll-like receptors (TLR)2, TLR4, and CD14 in monocyte TNF-alpha production induced by lipopolysaccharides from Neisseria meningitidis.
Toll-like receptors confer responsiveness to lipopolysaccharide from Porphyromonas gingivalis in human gingival fibroblasts.
MD-2, a molecule that confers lipopolysaccharide responsiveness on Toll-like receptor 4.
Roth CE, Craveiro RB, Niederau C, Malyaran H, Neuss S, Jankowski J, Wolf M
International journal of molecular sciences 2022 Jul 22;23(15)
International journal of molecular sciences 2022 Jul 22;23(15)
Helicobacter pylori Neutrophil-Activating Protein Directly Interacts with and Activates Toll-like Receptor 2 to Induce the Secretion of Interleukin-8 from Neutrophils and ATRA-Induced Differentiated HL-60 Cells.
Wen SH, Hong ZW, Chen CC, Chang HW, Fu HW
International journal of molecular sciences 2021 Oct 26;22(21)
International journal of molecular sciences 2021 Oct 26;22(21)
Lys694Arg polymorphism leads to blunted responses to LPS by interfering TLR4 with recruitment of MyD88.
Yang Y, Hu Y, Zhou Y, Liang T, Tang H, Ju H, Shi Q, Fang H
Innate immunity 2021 Aug;27(6):483-492
Innate immunity 2021 Aug;27(6):483-492
Inflammatory signaling in dengue-infected platelets requires translation and secretion of nonstructural protein 1.
Quirino-Teixeira AC, Rozini SV, Barbosa-Lima G, Coelho DR, Carneiro PH, Mohana-Borges R, Bozza PT, Hottz ED
Blood advances 2020 May 12;4(9):2018-2031
Blood advances 2020 May 12;4(9):2018-2031
Bacterial lipopolysaccharide and antimicrobial LL-37 enhance ICAM-1 expression and NF-κB p65 phosphorylation in senescent endothelial cells.
Suzuki K, Ohkuma M, Nagaoka I
International journal of molecular medicine 2019 Oct;44(4):1187-1196
International journal of molecular medicine 2019 Oct;44(4):1187-1196
Bushfire smoke is pro-inflammatory and suppresses macrophage phagocytic function.
Hamon R, Tran HB, Roscioli E, Ween M, Jersmann H, Hodge S
Scientific reports 2018 Sep 7;8(1):13424
Scientific reports 2018 Sep 7;8(1):13424
Butyrate upregulates the TLR4 expression and the phosphorylation of MAPKs and NK-κB in colon cancer cell in vitro.
Xiao T, Wu S, Yan C, Zhao C, Jin H, Yan N, Xu J, Wu Y, Li C, Shao Q, Xia S
Oncology letters 2018 Oct;16(4):4439-4447
Oncology letters 2018 Oct;16(4):4439-4447
Toll‑Like receptor 4 promotes the phosphorylation of CRMP2 via the activation of Rho‑kinase in MCAO rats.
Li XB, Ding MX, Ding CL, Li LL, Feng J, Yu XJ
Molecular medicine reports 2018 Jul;18(1):342-348
Molecular medicine reports 2018 Jul;18(1):342-348
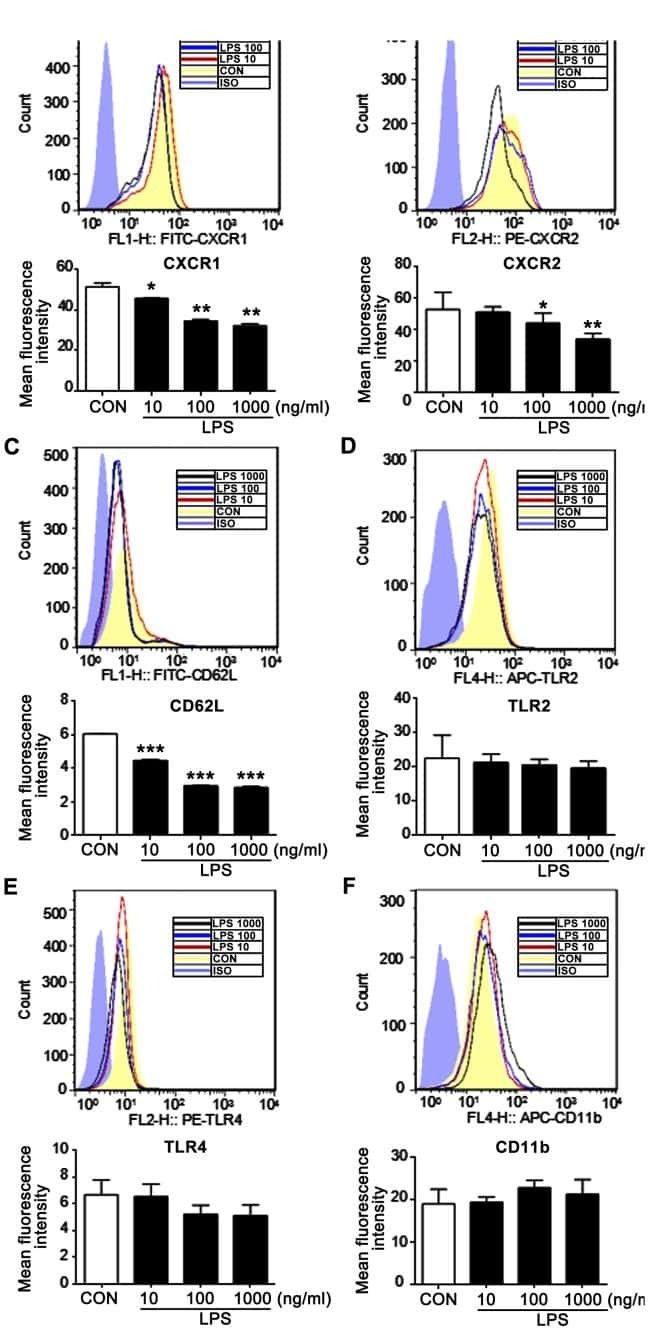
n-butanol extract from Folium isatidis inhibits the lipopolysaccharide-induced downregulation of CXCR1 and CXCR2 on human neutrophils.
Wu B, Wang L, Jiang L, Dong L, Xu F, Lu Y, Jin J, Wang Z, Liang G, Shan X
Molecular medicine reports 2018 Jan;17(1):179-185
Molecular medicine reports 2018 Jan;17(1):179-185
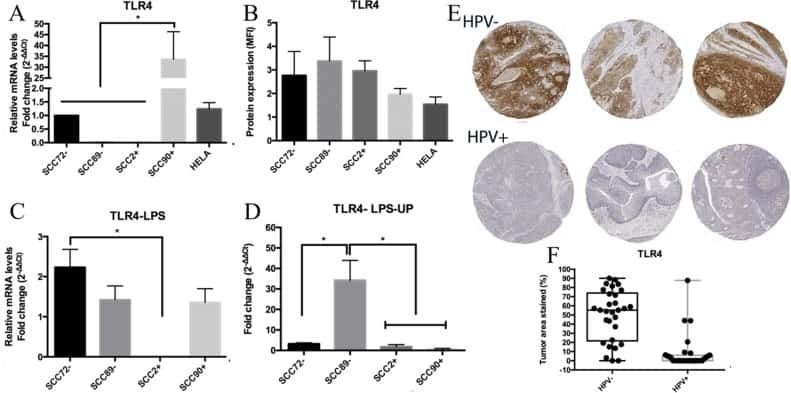
Altered Toll-like receptor expression and function in HPV-associated oropharyngeal carcinoma.
Tobouti PL, Bolt R, Radhakrishnan R, de Sousa SCOM, Hunter KD
Oncotarget 2018 Jan 2;9(1):236-248
Oncotarget 2018 Jan 2;9(1):236-248
Platelet proteome reveals novel pathways of platelet activation and platelet-mediated immunoregulation in dengue.
Trugilho MRO, Hottz ED, Brunoro GVF, Teixeira-Ferreira A, Carvalho PC, Salazar GA, Zimmerman GA, Bozza FA, Bozza PT, Perales J
PLoS pathogens 2017 May;13(5):e1006385
PLoS pathogens 2017 May;13(5):e1006385
Suppression of αvβ6 Integrin Expression by Polymicrobial Oral Biofilms in Gingival Epithelial Cells.
Bi J, Koivisto L, Pang A, Li M, Jiang G, Aurora S, Wang Z, Owen GR, Dai J, Shen Y, Grenier D, Haapasalo M, Häkkinen L, Larjava H
Scientific reports 2017 Jun 30;7(1):4411
Scientific reports 2017 Jun 30;7(1):4411
Comparative miRNA-Based Fingerprinting Reveals Biological Differences in Human Olfactory Mucosa- and Bone-Marrow-Derived Mesenchymal Stromal Cells.
Lindsay SL, Johnstone SA, McGrath MA, Mallinson D, Barnett SC
Stem cell reports 2016 May 10;6(5):729-742
Stem cell reports 2016 May 10;6(5):729-742
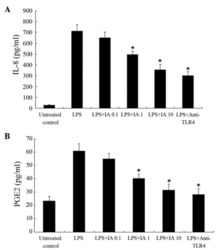
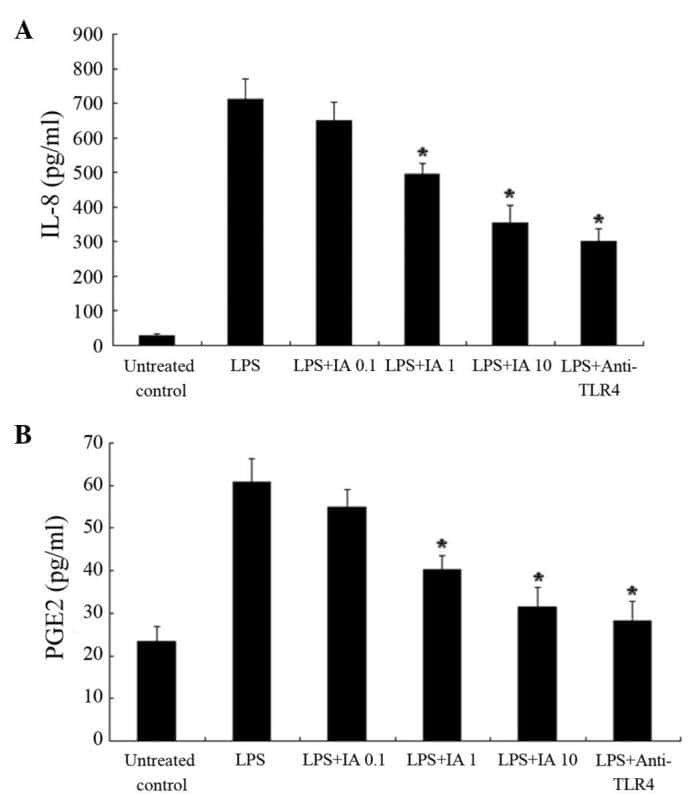
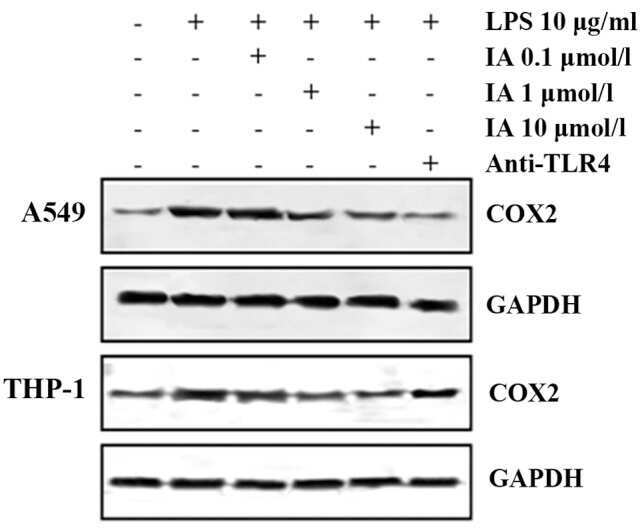
Picfeltarraenin IA inhibits lipopolysaccharide-induced inflammatory cytokine production by the nuclear factor-κB pathway in human pulmonary epithelial A549 cells.
Shi R, Wang Q, Ouyang Y, Wang Q, Xiong X
Oncology letters 2016 Feb;11(2):1195-1200
Oncology letters 2016 Feb;11(2):1195-1200
Lipopolysaccharide induces SBD-1 expression via the P38 MAPK signaling pathway in ovine oviduct epithelial cells.
Li Q, Bao F, Zhi D, Liu M, Yan Q, Zheng X, Ren L, Cong S, Li Y, Cao G
Lipids in health and disease 2016 Aug 11;15(1):127
Lipids in health and disease 2016 Aug 11;15(1):127
Toll-Like Receptor Activation by Generalized Modules for Membrane Antigens from Lipid A Mutants of Salmonella enterica Serovars Typhimurium and Enteritidis.
Rossi O, Caboni M, Negrea A, Necchi F, Alfini R, Micoli F, Saul A, MacLennan CA, Rondini S, Gerke C
Clinical and vaccine immunology : CVI 2016 Apr;23(4):304-14
Clinical and vaccine immunology : CVI 2016 Apr;23(4):304-14
TLR4 Expression Is Associated with Left Ventricular Dysfunction in Patients Undergoing Coronary Artery Bypass Surgery.
Avlas O, Bragg A, Fuks A, Nicholson JD, Farkash A, Porat E, Aravot D, Levy-Drummer RS, Cohen C, Shainberg A, Arad M, Hochhauser E
PloS one 2015;10(6):e0120175
PloS one 2015;10(6):e0120175
Biologically active polymers from spontaneous carotenoid oxidation: a new frontier in carotenoid activity.
Johnston JB, Nickerson JG, Daroszewski J, Mogg TJ, Burton GW
PloS one 2014;9(10):e111346
PloS one 2014;9(10):e111346
Biomechanical loading modulates proinflammatory and bone resorptive mediators in bacterial-stimulated PDL cells.
Nogueira AV, Nokhbehsaim M, Eick S, Bourauel C, Jäger A, Jepsen S, Rossa C Jr, Deschner J, Cirelli JA
Mediators of inflammation 2014;2014:425421
Mediators of inflammation 2014;2014:425421
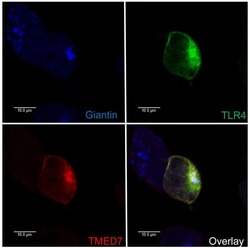
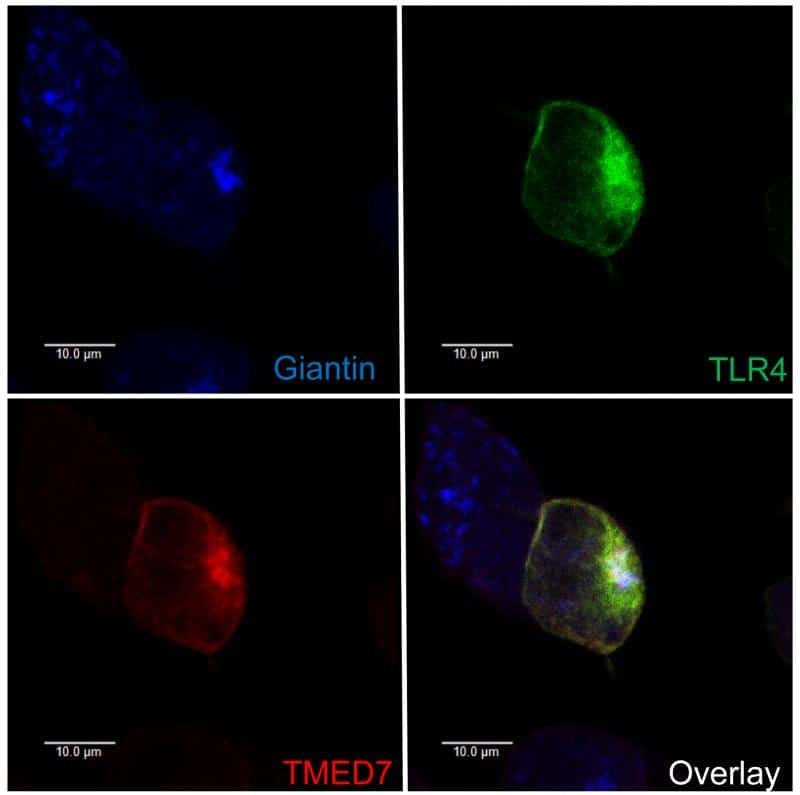
The COP II adaptor protein TMED7 is required to initiate and mediate the delivery of TLR4 to the plasma membrane.
Liaunardy-Jopeace A, Bryant CE, Gay NJ
Science signaling 2014 Jul 29;7(336):ra70
Science signaling 2014 Jul 29;7(336):ra70
HIV-1 gp120 induces TLR2- and TLR4-mediated innate immune activation in human female genital epithelium.
Nazli A, Kafka JK, Ferreira VH, Anipindi V, Mueller K, Osborne BJ, Dizzell S, Chauvin S, Mian MF, Ouellet M, Tremblay MJ, Mossman KL, Ashkar AA, Kovacs C, Bowdish DM, Snider DP, Kaul R, Kaushic C
Journal of immunology (Baltimore, Md. : 1950) 2013 Oct 15;191(8):4246-58
Journal of immunology (Baltimore, Md. : 1950) 2013 Oct 15;191(8):4246-58
Body fluid exosomes promote secretion of inflammatory cytokines in monocytic cells via Toll-like receptor signaling.
Bretz NP, Ridinger J, Rupp AK, Rimbach K, Keller S, Rupp C, Marmé F, Umansky L, Umansky V, Eigenbrod T, Sammar M, Altevogt P
The Journal of biological chemistry 2013 Dec 20;288(51):36691-702
The Journal of biological chemistry 2013 Dec 20;288(51):36691-702
Impaired clearance of apoptotic cells leads to HMGB1 release in the bone marrow of patients with myelodysplastic syndromes and induces TLR4-mediated cytokine production.
Velegraki M, Papakonstanti E, Mavroudi I, Psyllaki M, Tsatsanis C, Oulas A, Iliopoulos I, Katonis P, Papadaki HA
Haematologica 2013 Aug;98(8):1206-15
Haematologica 2013 Aug;98(8):1206-15
Monocytes from tuberculosis patients that exhibit cleaved caspase 9 and denaturalized cytochrome c are more susceptible to death mediated by Toll-like receptor 2.
Chávez-Galán L, Sada-Ovalle I, Baez-Saldaña R, Chávez R, Lascurain R
Immunology 2012 Apr;135(4):299-311
Immunology 2012 Apr;135(4):299-311
Late repression of NF-κB activity by invasive but not non-invasive meningococcal isolates is required to display apoptosis of epithelial cells.
Deghmane AE, El Kafsi H, Giorgini D, Abaza A, Taha MK
PLoS pathogens 2011 Dec;7(12):e1002403
PLoS pathogens 2011 Dec;7(12):e1002403
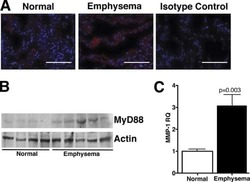
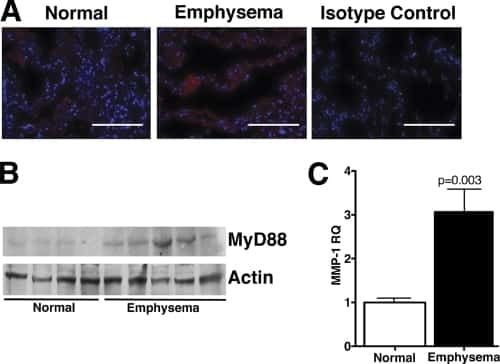
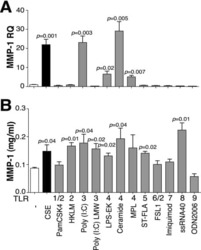
TLR4 protein contributes to cigarette smoke-induced matrix metalloproteinase-1 (MMP-1) expression in chronic obstructive pulmonary disease.
Geraghty P, Dabo AJ, D'Armiento J
The Journal of biological chemistry 2011 Aug 26;286(34):30211-8
The Journal of biological chemistry 2011 Aug 26;286(34):30211-8
Donor Toll-like receptor 4 contributes to ischemia and reperfusion injury following human kidney transplantation.
Krüger B, Krick S, Dhillon N, Lerner SM, Ames S, Bromberg JS, Lin M, Walsh L, Vella J, Fischereder M, Krämer BK, Colvin RB, Heeger PS, Murphy BT, Schröppel B
Proceedings of the National Academy of Sciences of the United States of America 2009 Mar 3;106(9):3390-5
Proceedings of the National Academy of Sciences of the United States of America 2009 Mar 3;106(9):3390-5
Aggregation and microparticle production through toll-like receptor 4 activation in platelets from recently menopausal women.
Hashimoto K, Jayachandran M, Owen WG, Miller VM
Journal of cardiovascular pharmacology 2009 Jul;54(1):57-62
Journal of cardiovascular pharmacology 2009 Jul;54(1):57-62
Candesartan inhibits Toll-like receptor expression and activity both in vitro and in vivo.
Dasu MR, Riosvelasco AC, Jialal I
Atherosclerosis 2009 Jan;202(1):76-83
Atherosclerosis 2009 Jan;202(1):76-83
High-mobility group box 1 protein induces tissue factor expression in vascular endothelial cells via activation of NF-kappaB and Egr-1.
Lv B, Wang H, Tang Y, Fan Z, Xiao X, Chen F
Thrombosis and haemostasis 2009 Aug;102(2):352-9
Thrombosis and haemostasis 2009 Aug;102(2):352-9
Mycobacterium tuberculosis-induced gamma interferon production by natural killer cells requires cross talk with antigen-presenting cells involving Toll-like receptors 2 and 4 and the mannose receptor in tuberculous pleurisy.
Schierloh P, Yokobori N, Alemán M, Landoni V, Geffner L, Musella RM, Castagnino J, Baldini M, Abbate E, de la Barrera SS, Sasiain MC
Infection and immunity 2007 Nov;75(11):5325-37
Infection and immunity 2007 Nov;75(11):5325-37
Human heat shock protein 60 stimulates vascular smooth muscle cell proliferation through Toll-like receptors 2 and 4.
de Graaf R, Kloppenburg G, Kitslaar PJ, Bruggeman CA, Stassen F
Microbes and infection 2006 Jun;8(7):1859-65
Microbes and infection 2006 Jun;8(7):1859-65
Role of dendritic cell-derived CXCL13 in the pathogenesis of Bartonella henselae B-rich granuloma.
Vermi W, Facchetti F, Riboldi E, Heine H, Scutera S, Stornello S, Ravarino D, Cappello P, Giovarelli M, Badolato R, Zucca M, Gentili F, Chilosi M, Doglioni C, Ponzi AN, Sozzani S, Musso T
Blood 2006 Jan 15;107(2):454-62
Blood 2006 Jan 15;107(2):454-62
The uptake of a Klebsiella pneumoniae capsule polysaccharide mutant triggers an inflammatory response by human airway epithelial cells.
Regueiro V, Campos MA, Pons J, Albertí S, Bengoechea JA
Microbiology (Reading, England) 2006 Feb;152(Pt 2):555-566
Microbiology (Reading, England) 2006 Feb;152(Pt 2):555-566
Respiratory epithelial cells require Toll-like receptor 4 for induction of human beta-defensin 2 by lipopolysaccharide.
MacRedmond R, Greene C, Taggart CC, McElvaney N, O'Neill S
Respiratory research 2005 Oct 12;6(1):116
Respiratory research 2005 Oct 12;6(1):116
Distinct strains of Propionibacterium acnes induce selective human beta-defensin-2 and interleukin-8 expression in human keratinocytes through toll-like receptors.
Nagy I, Pivarcsi A, Koreck A, Széll M, Urbán E, Kemény L
The Journal of investigative dermatology 2005 May;124(5):931-8
The Journal of investigative dermatology 2005 May;124(5):931-8
Surface calreticulin mediates muramyl dipeptide-induced apoptosis in RK13 cells.
Chen D, Texada DE, Duggan C, Liang C, Reden TB, Kooragayala LM, Langford MP
The Journal of biological chemistry 2005 Jun 10;280(23):22425-36
The Journal of biological chemistry 2005 Jun 10;280(23):22425-36
TLR9 is localized in the endoplasmic reticulum prior to stimulation.
Leifer CA, Kennedy MN, Mazzoni A, Lee C, Kruhlak MJ, Segal DM
Journal of immunology (Baltimore, Md. : 1950) 2004 Jul 15;173(2):1179-83
Journal of immunology (Baltimore, Md. : 1950) 2004 Jul 15;173(2):1179-83
Differential regulation of inflammatory cytokine secretion by human dendritic cells upon Chlamydia trachomatis infection.
Gervassi A, Alderson MR, Suchland R, Maisonneuve JF, Grabstein KH, Probst P
Infection and immunity 2004 Dec;72(12):7231-9
Infection and immunity 2004 Dec;72(12):7231-9
Expression and involvement of Toll-like receptors (TLR)2, TLR4, and CD14 in monocyte TNF-alpha production induced by lipopolysaccharides from Neisseria meningitidis.
Mirlashari MR, Lyberg T
Medical science monitor : international medical journal of experimental and clinical research 2003 Aug;9(8):BR316-24
Medical science monitor : international medical journal of experimental and clinical research 2003 Aug;9(8):BR316-24
Toll-like receptors confer responsiveness to lipopolysaccharide from Porphyromonas gingivalis in human gingival fibroblasts.
Tabeta K, Yamazaki K, Akashi S, Miyake K, Kumada H, Umemoto T, Yoshie H
Infection and immunity 2000 Jun;68(6):3731-5
Infection and immunity 2000 Jun;68(6):3731-5
MD-2, a molecule that confers lipopolysaccharide responsiveness on Toll-like receptor 4.
Shimazu R, Akashi S, Ogata H, Nagai Y, Fukudome K, Miyake K, Kimoto M
The Journal of experimental medicine 1999 Jun 7;189(11):1777-82
The Journal of experimental medicine 1999 Jun 7;189(11):1777-82
No comments: Submit comment
Supportive validation
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
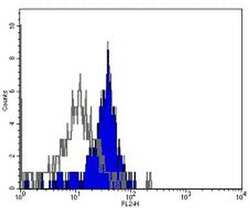
- Staining of normal human peripheral blood cells with Mouse IgG2a Isotype Control (Product # 16-4724-85) or Anti-Human CD284 (TLR4) Functional Grade Purified. Cells in the monocyte population were used for analysis.
Supportive validation
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
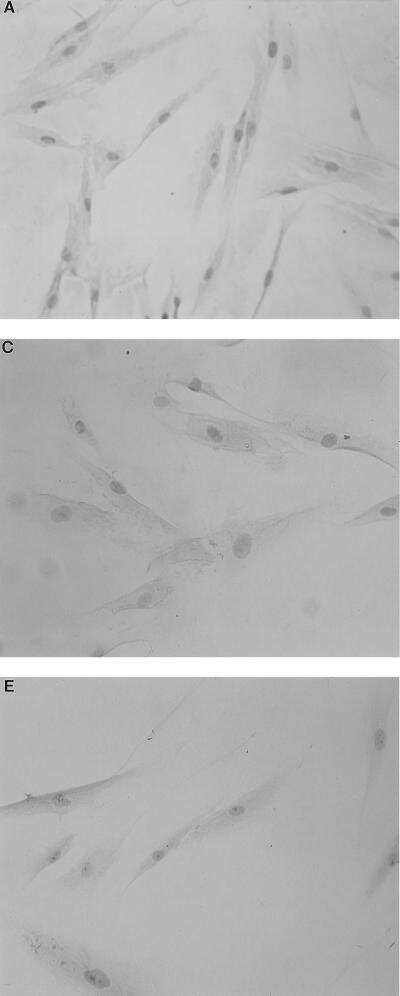
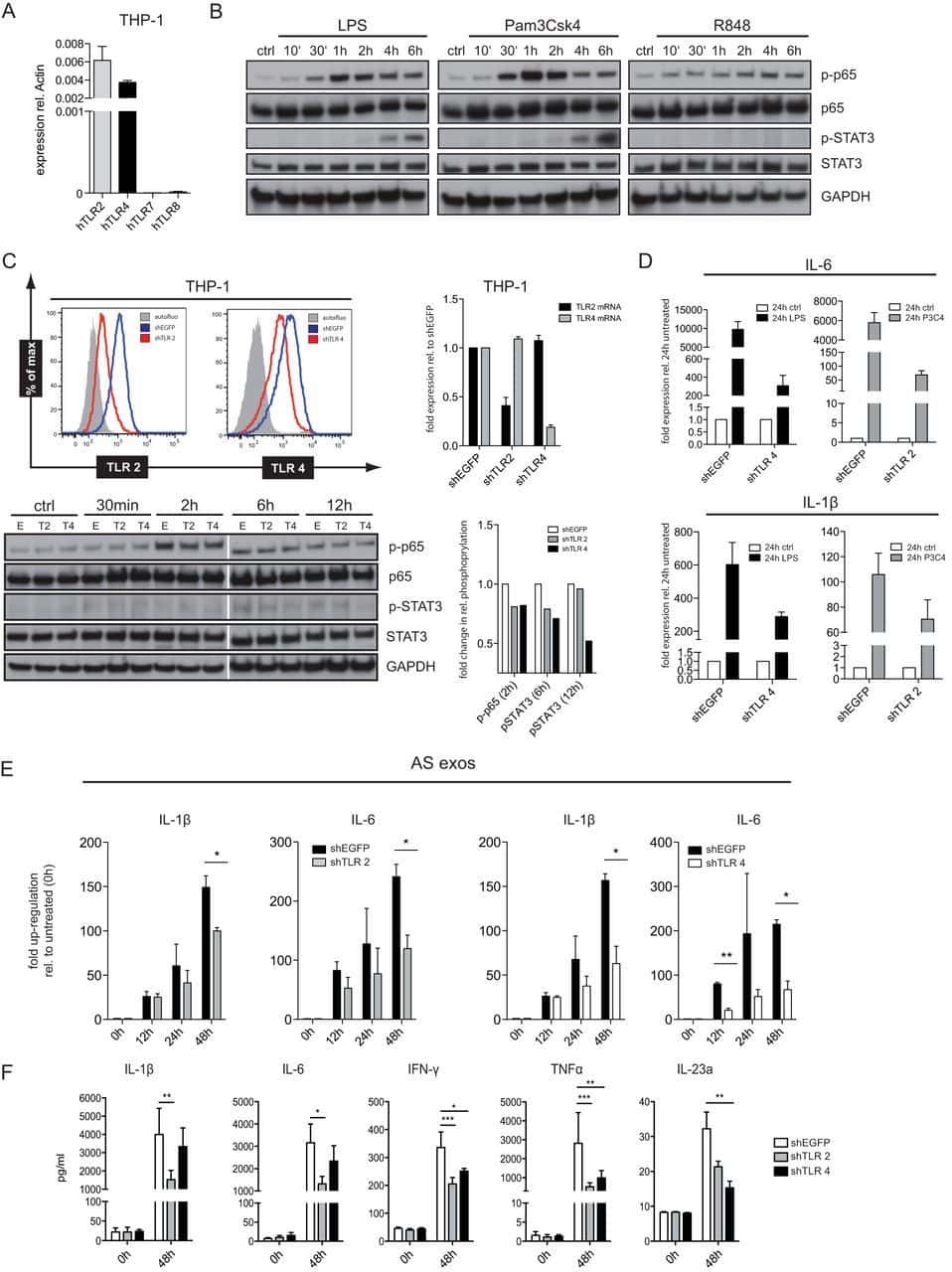
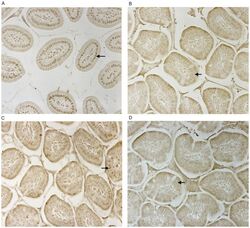
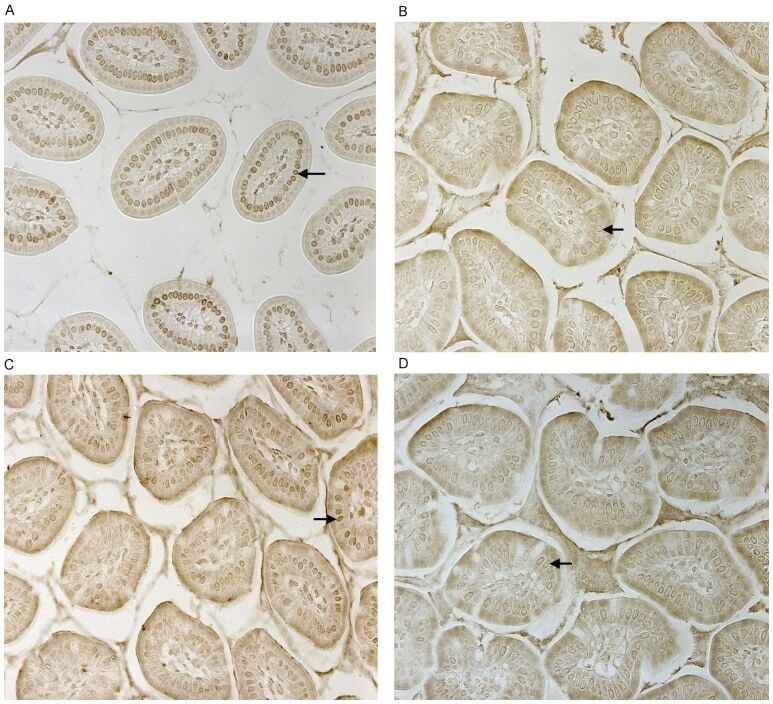
- Figure 2 CD14 and TLR-4 staining in gut epithelial cells. Balb/c mice were not supplemented (control) or supplemented daily by oral gavage with OxC-beta (10 mg/kg). After 4 weeks, intestinal tissues were harvested and CD14 and TLR-4 expression was determined by immunocytochemistry. Increased CD14 (A) and TLR-4 (C) expression is readily apparent in epithelial cells in the OxC-beta-supplemented animals compared to the controls receiving vehicle alone (B and D, respectively). Arrows indicate the location of enterocytes within the cross section of microvilli. Magnification 40x.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
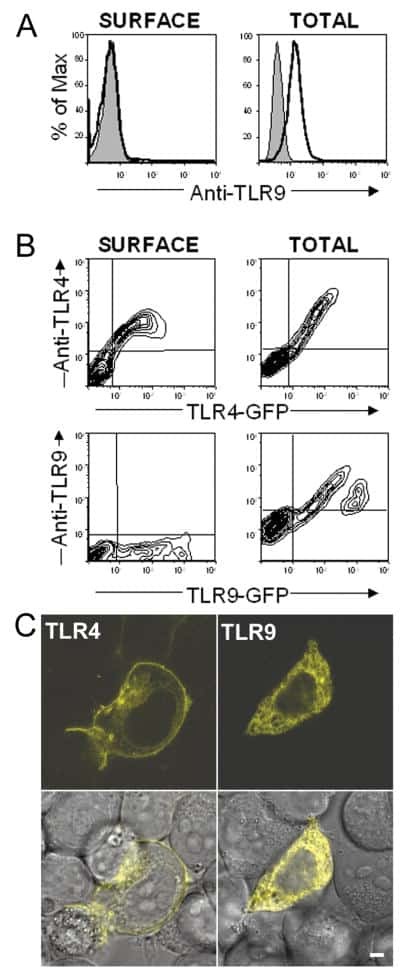
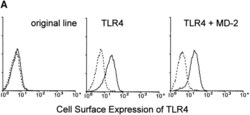
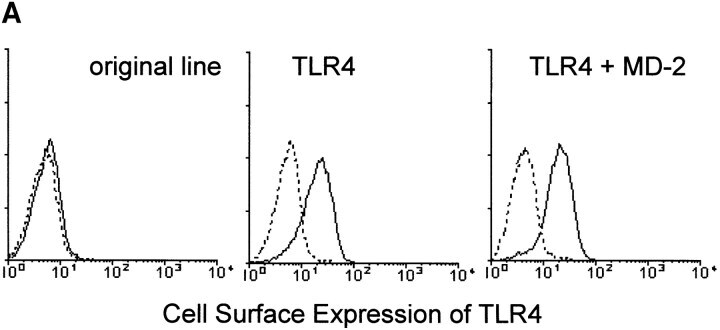
- Figure 4 MD-2 is coprecipitated with TLR4. (A) Stable transfectants expressing MD-2 alone or with TLR4 were stained with or without cell permeabilization. The intracellular MD-2 precursor was stained with the anti-protein C mAb. The specificity of the anti-TLR4 mAb was shown by cell surface staining of stable transfectants. Goat anti- mouse IgG-FITC was used as the second reagent. Dotted lines correspond to histograms stained with the second reagent alone. Panels correspond to the original Ba/F3 line, a transfectant expressing TLR4 alone, and a transfectant expressing TLR4 + MD-2, respectively. (B) Stable transfectants expressing TLR4 and MD-2 were subjected to immunoprecipitation with control mouse IgG (lane 1) or the anti-TLR4 mAb, HTA125 (lanes 2 and 3). After blotting, precipitated molecules were detected with an anti-flag mAb, M2. The precipitates shown in lanes 1 and 2 are from the transfectant expressing TLR4 and MD-2, both of which were tagged with the flag epitope. In lane 3, we used the control line in which the flag epitope was on TLR4 but not on MD-2.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
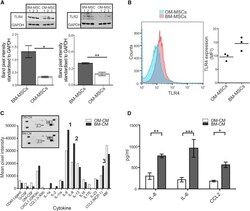
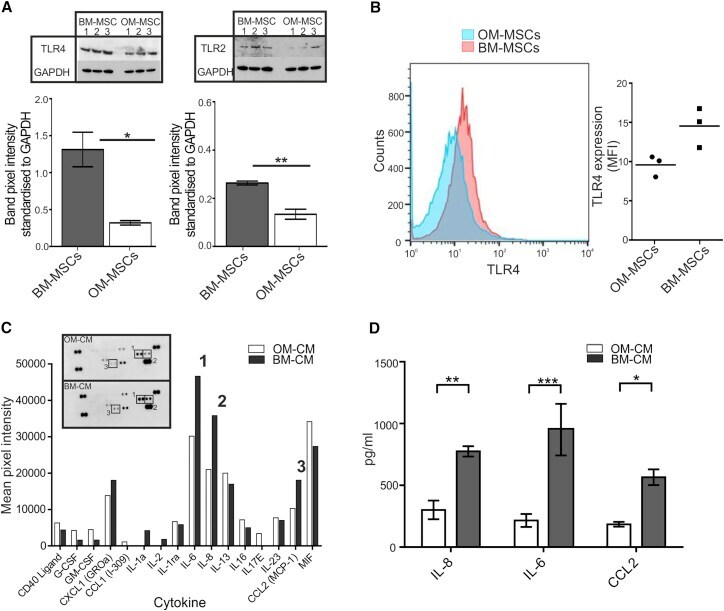
- Figure 5 MiR-146-5p Differentially Regulates TLR4, TLR2, and Cytokine Secretion (A) Graphical representation of TLR4 and TLR2 (western blot shown as inserts) in BM-MSCs (n = 6 patient samples) and OM-MSCs (n = 6 patient samples). BM-MSCs have significantly higher levels compared with OM-MSCs (mean +- SEM, * p < 0.05, ** p < 0.01, Students unpaired t test). (B) FACS analysis of TLR4 expression on OM- and BM-MSCs (n = 3, patient samples both). (C) OM-MSC (n = 1 patient sample) and BM-MSC (n = 1 patient sample) cytokine profile. Insert shows dot plot while the graph illustrates the mean pixel intensity of the blot. IL-6 (1) IL-8 (2), and CCL2 (3) were expressed 1.5-fold higher in BM-MSC-CM. (D) ELISA analysis of IL-8, IL-6, and CCL2 in BM-MSC-CM (n = 3 patient samples) and OM-MSC-CM (n = 3 patient samples, mean +- SEM, * p < 0.05, ** p < 0.01, *** p < 0.001, Students unpaired t test).
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
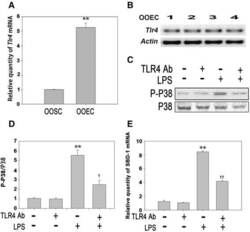
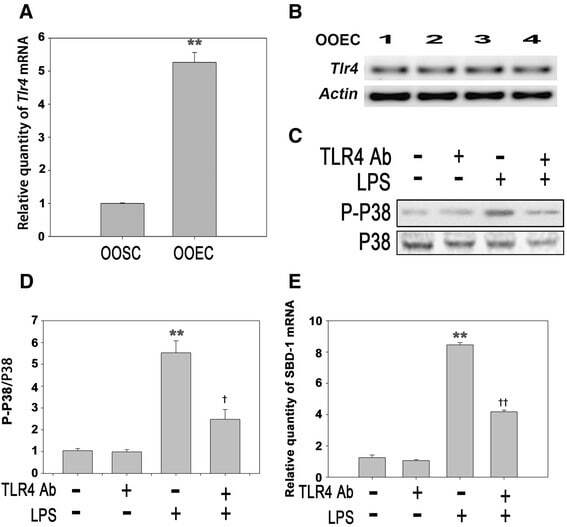
- Fig. 4 LPS stimulates the expression of SBD-1 through TLR4. a Tlr4 mRNA expression of ovine oviduct stromal cells (OOSCs) and epithelial cells (OOECs) was assessed by qRT-PCR. b PCR was conducted to examine the expression of TLR4 in the ovine oviduct epithelial cells harvested at different times. c Western blotting was performed to examine the expression levels of P38 MAPK after treatment with LPS (100 ng/mL) or a TLR4 neutralizing antibody for 12 h. d The densitometric analysis of the bands on the western blotting showed that LPS could markedly activate P38 MAPK, while the separate addition of the TLR4 neutralizing antibody had no effect. However, treatment with the TLR4 neutralizing antibody could significantly decrease the levels of phosphorylated P38 induced by LPS. e QRT-PCR analysis was used to examine the mRNA levels of SBD-1 after treatment with the TLR4 neutralizing antibody for 12 h. Blocking TLR4 activity could inhibit the expression of SBD-1. All of the experiments were repeated at least three times. * p < 0.05, ** p < 0.01 ( t -test) vs. Control. + p < 0.05, ++ p < 0.01 ( t -test) vs. LPS
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
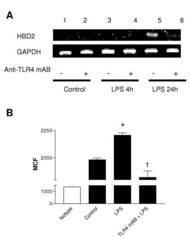
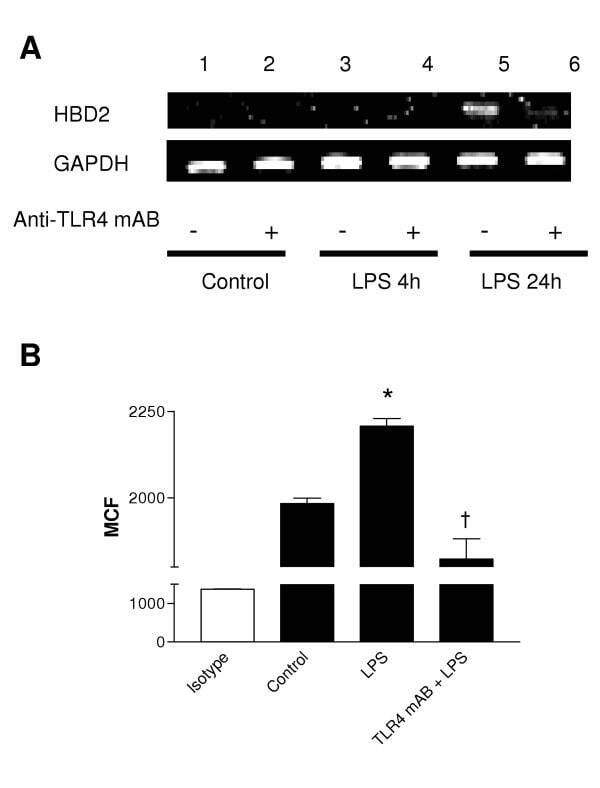
- Figure 3 LPS-induced HBD2 expression in A549 cells requires TLR4. A459 cells were incubated with an isotype control or anti-TLR4 neutralizing antibody (Anti-TLR4 mAB 5 mug/ml, 30 min) then, (A) left untreated or stimulated with LPS (10 mug) for 4 or 24 hours. Total RNA was extracted, reverse transcribed into cDNA and used as a template in PCR reactions using HBD2 gene-specific primers. Products were electrophoresed in 1.5% TBE agarose gels containing 0.5 mug/ml ethidium bromide and visualized under UV. Gels are representative of three independent experiments or (B) left untreated or stimulated with LPS (10 mug) for 24 hours, Fc-blocked and labeled with anti-HBD2 (solid) or isotype control antibodies (clear) and fluorophore-conjugated detection antibodies. HBD2 expression was quantified by laser scanning cytometry, as described, and data from three experiments is presented. HBD2 expression is expressed as Mean Channel Fluorescence (MCF) + SEM. (* P < 0.05 vs control, + P < 0.05 vs control + LPS).
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
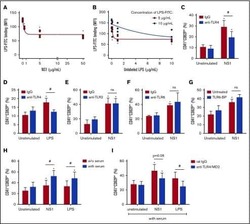
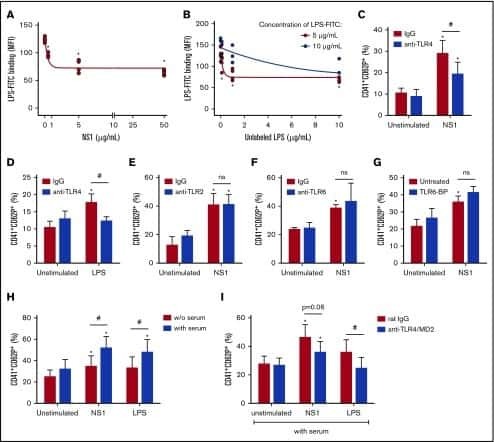
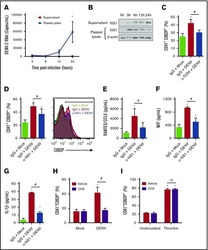
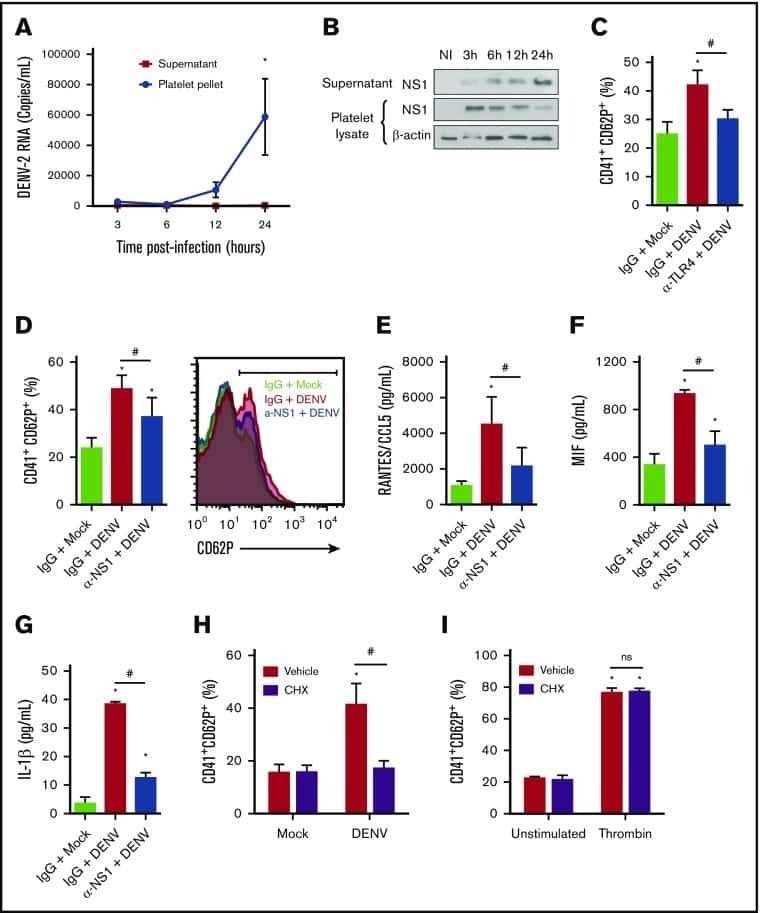
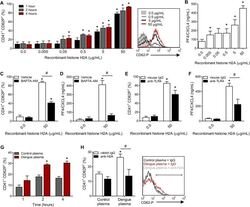
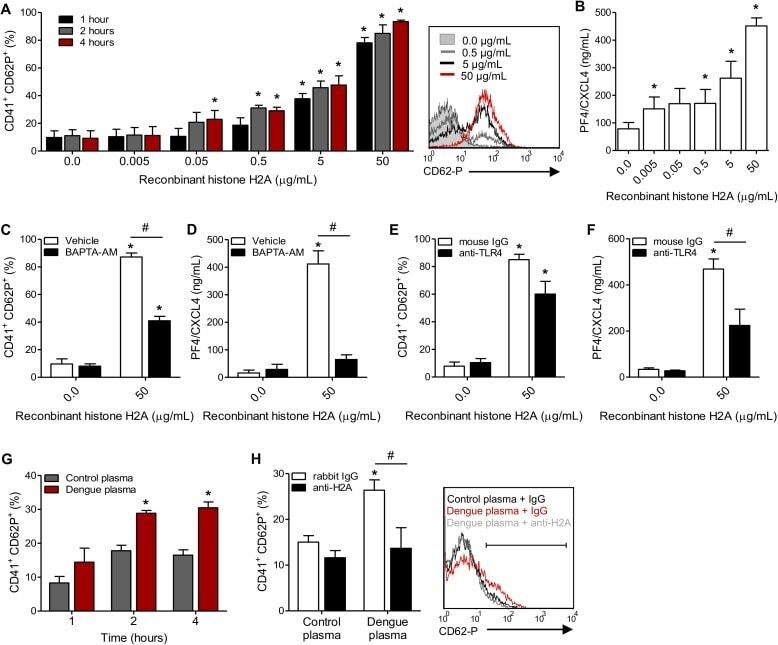
- Fig 6 Circulating histone H2A in plasma from dengue-infected patients activates platelets. ( A-B ) Platelets isolated from healthy volunteers were stimulated with recombinant human histone H2A at the indicated concentrations. ( A ) Platelet surface P-selectin (CD62-P) was evaluated 1, 2 and 4 hours post stimulation by flow cytometry and ( B ) PF4/CXCL4 concentration was measured in supernatants 4 hours post stimulation. ( C-F ) Surface P-selectin and PF4/CXCL4 concentration in the supernatants of platelets stimulated with recombinant histone H2A for 2 hour in the presence of ( C-D ) the calcium chelator BAPTA-AM (20 muM) or vehicle (DMSO); or ( E-F ) blocking antibody against TLR4 (20 mug/mL) or isotype matched IgG. ( G-H ) P-selectin expression on platelets exposed to ( G ) plasma from six dengue-infected patients (dengue plasma) or four heterologous healthy volunteers (control plasma) for the indicated time-points; and ( H ) platelets exposed to dengue plasma or control plasma for 4 hours in the presence of anti-histone H2A (20 mug/mL) or isotype matched IgG. Bars represent mean +- standard error of the mean of 3 independent experiments ( A-F ) and of 4 to 6 independent plasma donors ( G-H ). * indicates p
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
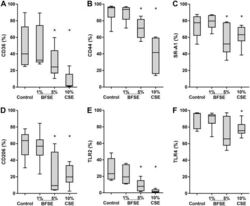
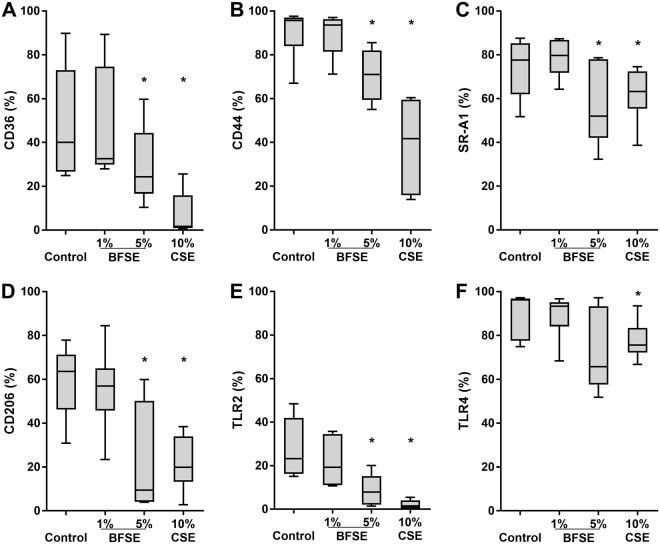
- Figure 2 BFSE alters phagocytic recognition receptors. MDM were exposure to air control, 1% or 5% BFSE or 10% CSE for 24 h. Cell surface markers: CD36 ( A ), CD44 ( B ), SR-A1 ( C ), CD206 ( D ), TLR-2 ( E ) and TLR-4 ( F ) were detected by immunofluorescence on a FACScanto II flow cytometer and expressed as % positive cells compared to negative controls. (n = 5) *p < 0.05.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
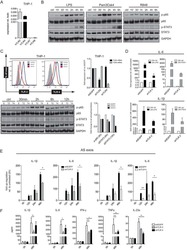
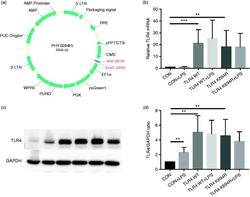
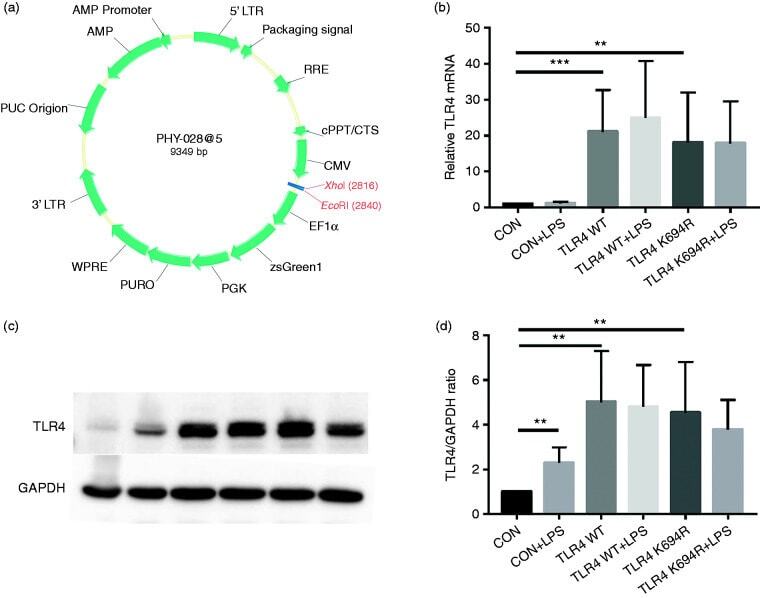
- Figure 2. The K694R polymorphism does not affect TLR4 expression levels. (a) Diagram of wild type (WT) and K694R (K694G) TLR4 plasmid vector. (b) mRNA expression levels for the genes TLR4 in human embryonic kidney 293T (HEK293T) cells transiently transfected with WT or K694R TLR4 plasmid or empty vector as control, treated with or without LPS for 6 h. (c) Western blot (WB) analysis of TLR4 expression levels in HEK293T cells transiently transfected with WT, K694R TLR4 plasmid or empty vector treated with or without LPS for 6h. (d) Quantification of TLR4 expression levels in (c). Graphs are the mean +- SD, n = 6, * P < 0.05, Student's t -test.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
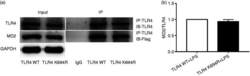
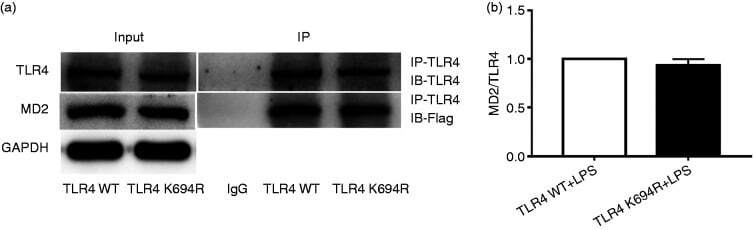
- Figure 5. The K694R polymorphism does not affect TLR4 interaction with myeloid differentiation factor 2 (MD2). (a) HEK293T cells transiently transfected with WT or K694R TLR4, CD14 and Flag-MD2 plasmids, treated with LPS, were subjected to co-immunoprecipitation with anti-TLR4 Ab, and analysed by WB with anti-Flag Ab. (b) Densitometric quantification of the data shown in (a). The results are presented as ratios of MD2 and TLR4. Graphs are the mean +- SD, n = 3, * P < 0.05, Student's t -test.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
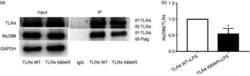
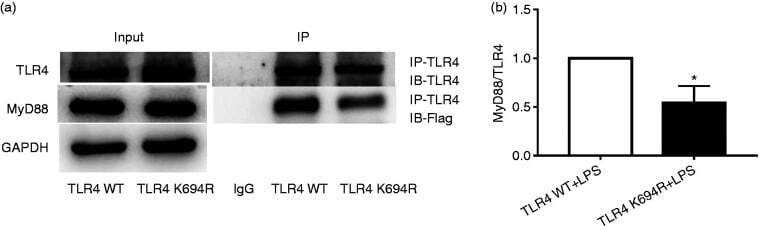
- Figure 6. The K694R polymorphism impairs the ability of TLR4 to recruit MyD88. (a) HEK293T cells transiently transfected with WT or K694R TLR4, CD14, MD2 and Flag-MyD88 plasmids, treated with LPS, were subjected to co-immunoprecipitation with anti-TLR4 Ab, and analysed by WB with anti-Flag Ab. (b) Densitometric quantification of the data shown in (a) The results are presented as ratios of MyD88 and TLR4. Graphs are the mean +- SD, n = 3, * P < 0.05, Student's t -test.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
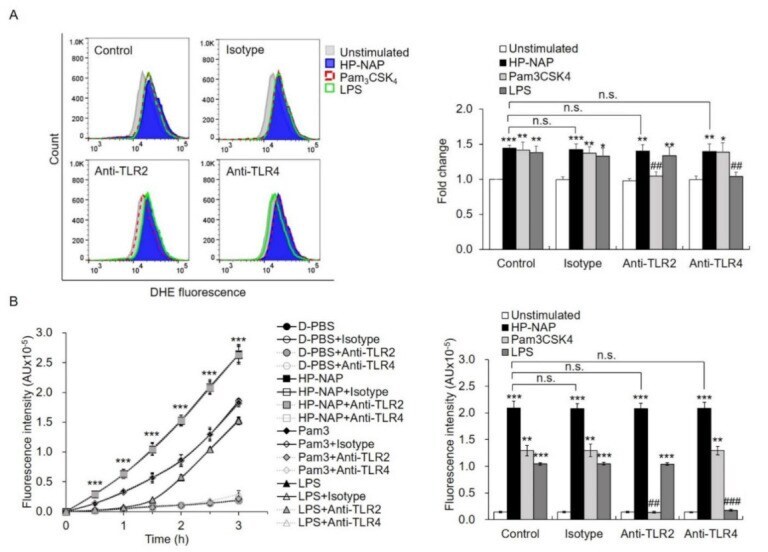
- Figure 5 The effect of TLR2-neutralizing antibodies on HP-NAP-induced ROS production by ATRA-induced differentiated HL-60 cells and neutrophils. ( A ) TLR2-independent ROS production in ATRA-induced differentiated HL-60 cells induced by HP-NAP. ATRA-induced differentiated HL-60 cells at a density of 4 x 10 6 cell/mL were pretreated with 10 mug/mL of anti-TLR2, anti-TLR4, or IgG2a isotype antibodies at 37 degC for 30 min and followed by the stimulation with 1 muM HP-NAP, 1 mug/mL Pam 3 CSK 4 , 10 mug/mL LPS, or D-PBS, pH 7.2, as the unstimulated control at 37 degC for 30 min. ROS production was measured as DHE-derived fluorescence by flow cytometry as described in Figure 2 C. Representative histograms are shown in the left panel. Data in the right panel are expressed as the fold change as described in Figure 2 C and as mean +- SD of three independent experiments. ( B ) TLR2-independent ROS production in neutrophils induced by HP-NAP. Neutrophils at a density of 2 x 10 6 cell/mL were pretreated with 10 mug/mL of anti-TLR2, anti-TLR4 or IgG2a isotype antibodies at 37 degC for 30 min and followed by the stimulation with 1 muM HP-NAP, 1 mug/mL Pam 3 CSK 4 , 10 mug/mL LPS or D-PBS, pH 7.2, as the unstimulated control for the indicated time. ROS production by neutrophils was determined by H 2 DCF-DA-derived derived fluorescence assay as described in Figure 2 A. Data from neutrophils stimulated with HP-NAP for 2.5 h are shown as bar graph and expressed as mean +- SD of four independe
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
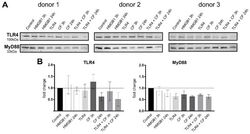
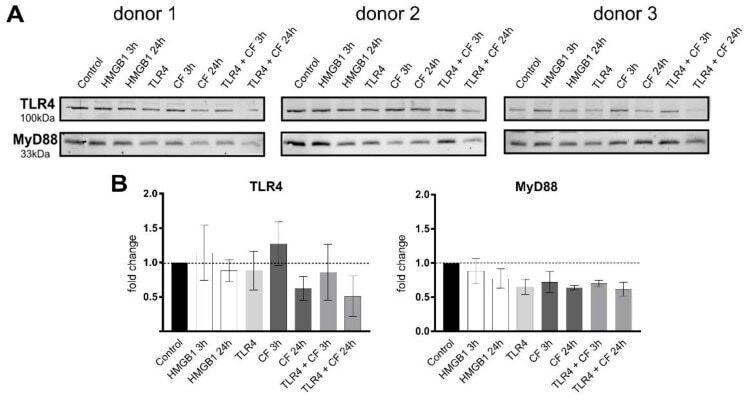
- Compressive forces and TLR4 blocking antibody modulate TLR4 production and MyD88 in primary PDL cells. ( A ) Protein levels of TLR4 and MyD88 were determined by Western blot in different conditions: HMGB1 (100 ng/mL), TLR4 blocking antibody (5 ug/mL) (TLR4: TLR4 blocking antibody) and compressive force (CF) 2 g/cm 2 for 3 and 24 h. Three different donors showed similar patterns with reduction of TLR4 antibody. TLR4 production was upregulated under 3 h compression forces. MyD88 production was reduced in all conditions. ( B ) Quantification of three donors, normalized to the control with stain-free technology. Data were tested for normal distribution by Shapiro-Wilk test. Afterward, a one-way analysis of variance (ANOVA) followed by Tukeys' post hoc test was performed.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
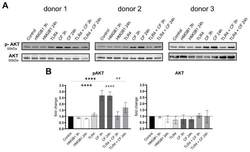
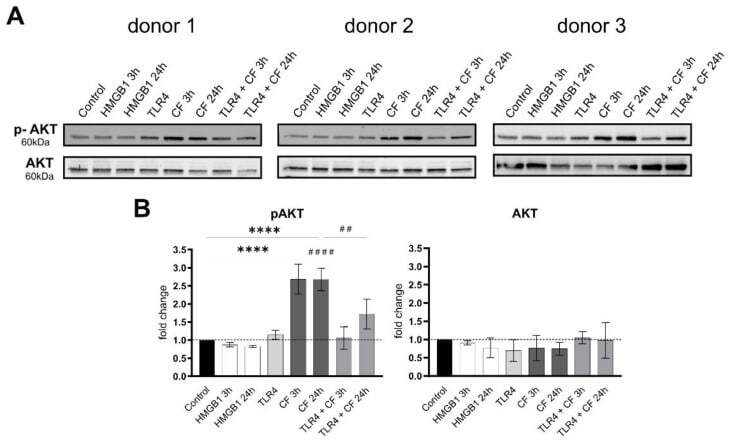
- Compressive forces upregulate phosphor AKT in primary hPDL cells. ( A ) Protein production levels and phospho-AKT were determined by Western blot in different conditions: HMGB1 (100 ng/mL), TLR4 blocking antibody (5 ug/mL) (TLR4: TLR4 blocking antibody) and compressive force (CF) 2 g/cm 2 at 3 and 24 h. Three different donors showed similar patterns. Under all conditions, AKT showed no differences, but the phosphorylated AKT was significantly upregulated with compressive forces (CF) for 3 h and 24 h. Compressive forces with additional blocking TLR4 monoclonal antibody led to significant downregulation comparable to the control. ( B ) Quantification of three donors, normalized to the control with stain-free technology. CF conditions without TLR4 blocking antibody showed a significant upregulation. Data were tested for normal distribution by Shapiro-Wilk test. Afterward, a one-way analysis of variance (ANOVA) followed by Tukeys' post hoc test was performed. Statistically significant differences to control are marked by asterisks (**** p < 0.0001) and hashtag show significant differences between CF and TLR4 +CF (## p < 0.01; #### p < 0.0001).
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
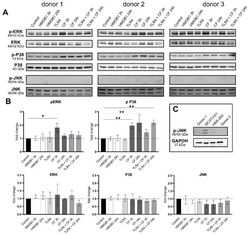
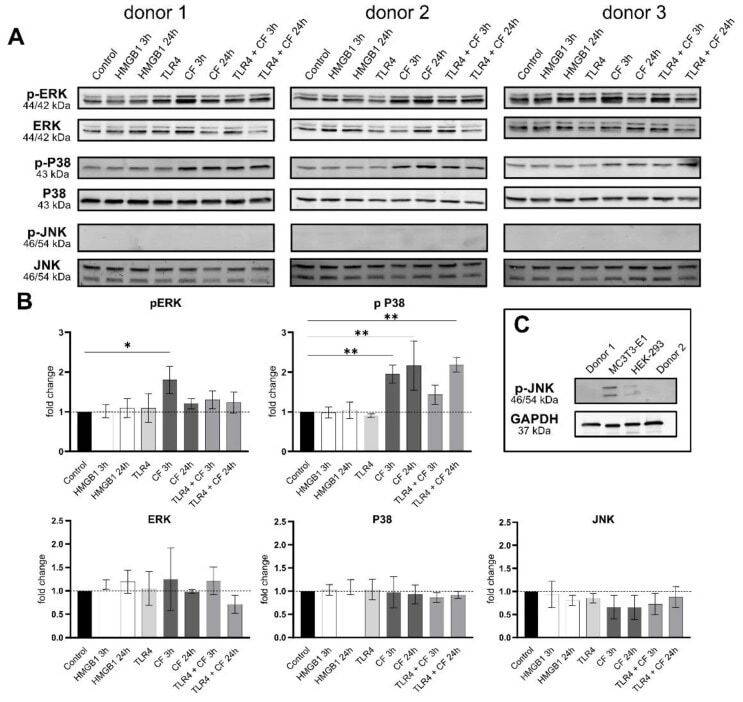
- Compressive forces upregulate pERK and p P38 in primary hPDL cells. ( A ) Protein levels and phospho-MAPK (ERK, p38 and JNK) were determined by Western blot in different conditions: HMGB1 (100 ng/mL), TLR4 blocking antibody (5 ug/mL) (TLR4: TLR4 blocking antibody) and compressive force 2 g/cm 2 at 3 and 24 h. Three different donors showed similar patterns. Under all conditions, ERK, p38 and JNK showed no differences, but the phosphorylated ERK and p38 were significantly upregulated with compressive forces (CF) as follows: ERK for 3 h and p38 for 3 h and 24 h. ( B ) Quantification of three donors, normalized to the control with stain-free technology. CF conditions without TLR4 blocking antibody showed a significant upregulation for phospho-ERK and phospho-p38. ( C ) HEK-293 and MC3T3 cells were used as a positive control for phospho-JNK antibody. Data were tested for normal distribution by Shapiro-Wilk test. Afterward, a one-way analysis of variance (ANOVA) followed by Tukeys' post hoc test was performed. Statistically significant differences to control are marked by asterisks (* p < 0.05; ** p < 0.01).
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
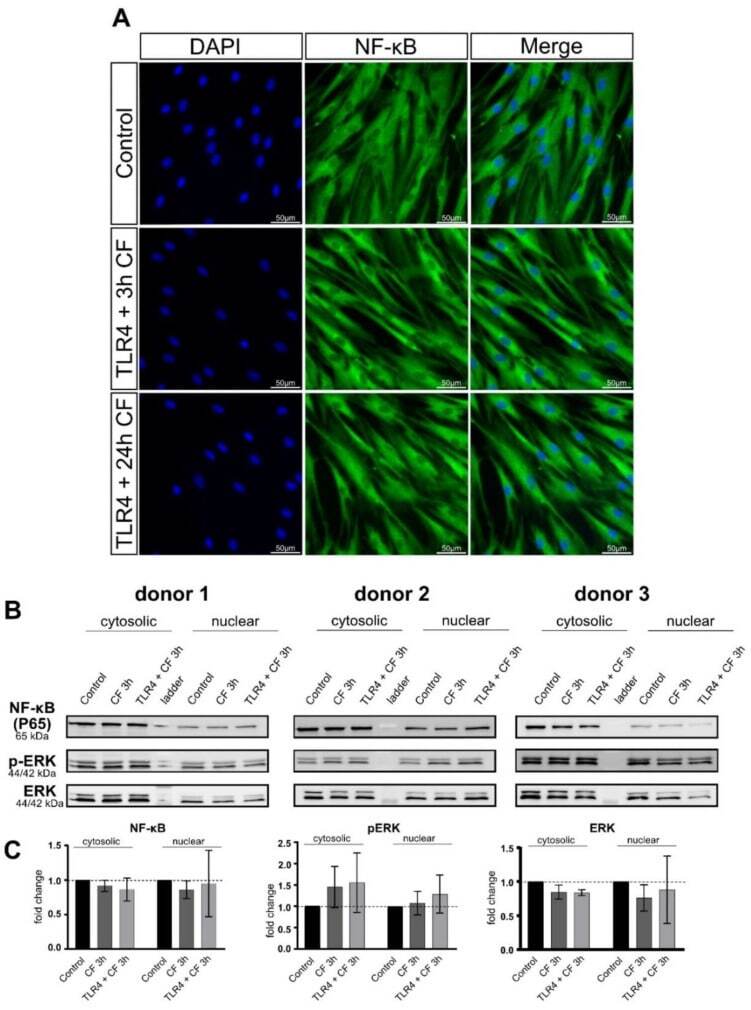
- Fluorescence images of CF and TLR4 blocking antibody on PDL cells. ( A ) PDL cells treaded with and without TLR4 blocking antibody (5 ug/mL) and compressive force 2 g/cm 2 for 24 h. NF-kB was stained green (Alexa 488), blue areas represent nuclei (DAPI); scalebar 50 um, CF: compressive force NF-kB was not translocated to the nucleus in any condition. ( B ) Protein production of NF-kB, ERK and phospho-ERK were determined by Western blot. ( C ) Quantification of three donors, normalized to the control with stain-free technology. CF conditions without TLR4 antibody showed a significant upregulation for phospho-ERK and phospho-p38.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
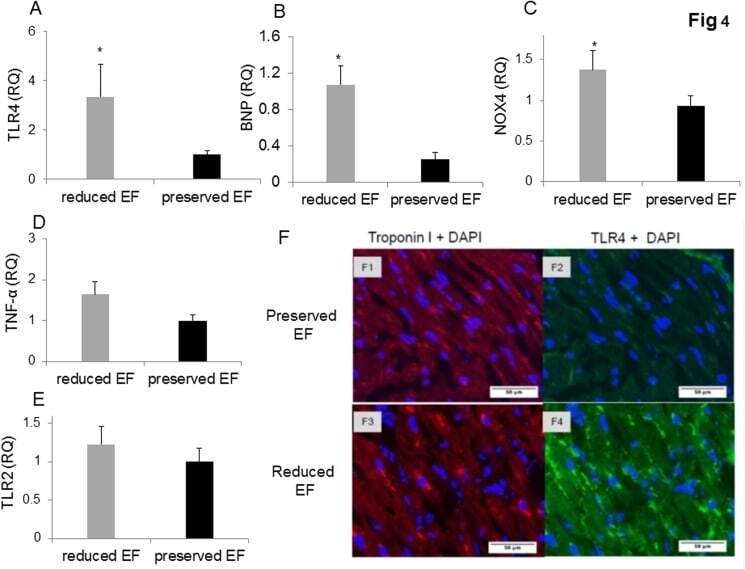
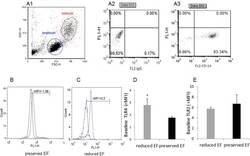
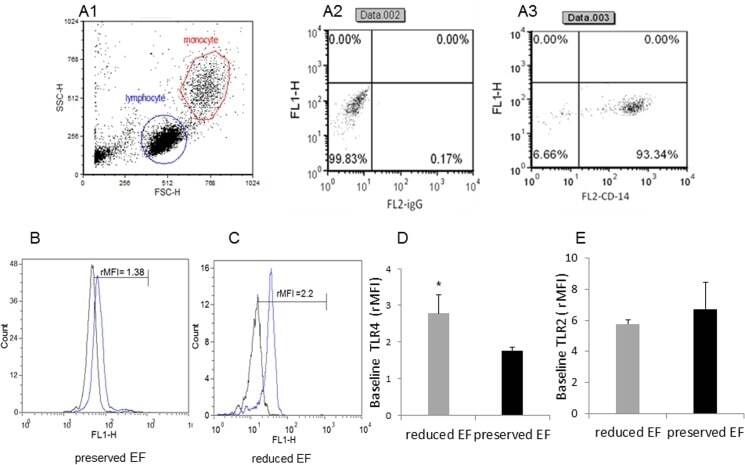
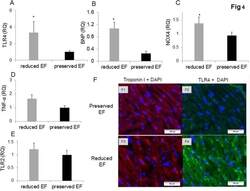
- Fig 2 Dot plot and histograms representing TLR4 and TLR2 levels in monocytes. (A1-A3) Monocytes/lymphocyte cell population after CD14-positive cell staining. (B-C) Shift to the right demonstrating an increase in the amount of TLR4 staining in the monocytes. Specific mean fluorescence (MFI) can be quantified from these histograms for each case. (D) TLR4 levels on the population of monocytes as measured using relative mean fluorescence intensity (rMFI). (C) 'reduced EF' patients (EF55%). (E) TLR2 levels on the population of monocytes as measured using rMFI. FACS analysis,* P
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
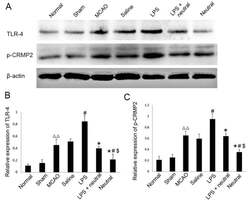
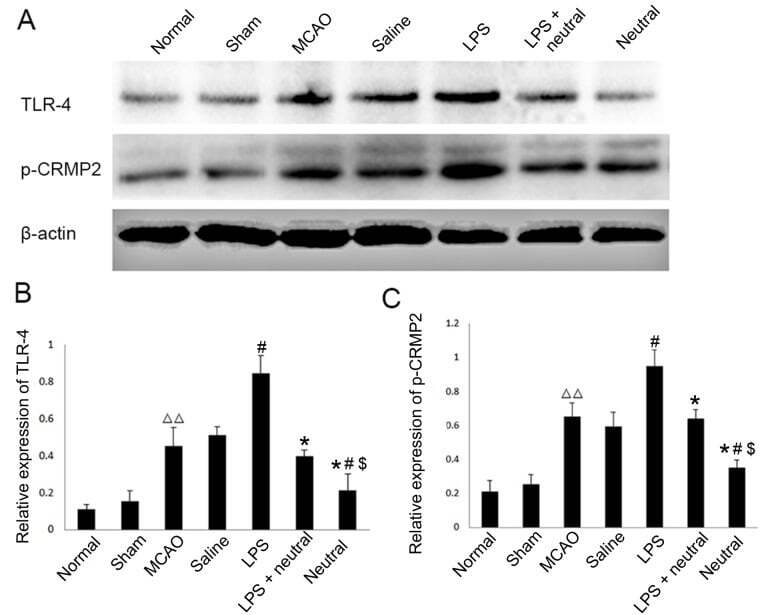
- Figure 1. Expression levels of TLR4 and phosphorylation of CRMP2 in the cortex via western blotting. (A) Western blotting was performed to reveal the expression levels of TLR4 and p-CRMP2 in each group. (B) Quantitative analysis of TLR4 expression relative to beta-actin from western blotting as presented in (A). (C) Quantitative analysis of p-CRMP2 expression relative to beta-actin from western blotting a presented in (A). Data are expressed as the mean +- standard deviation (n=5). P
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
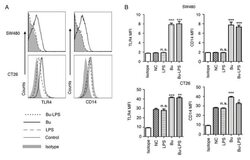
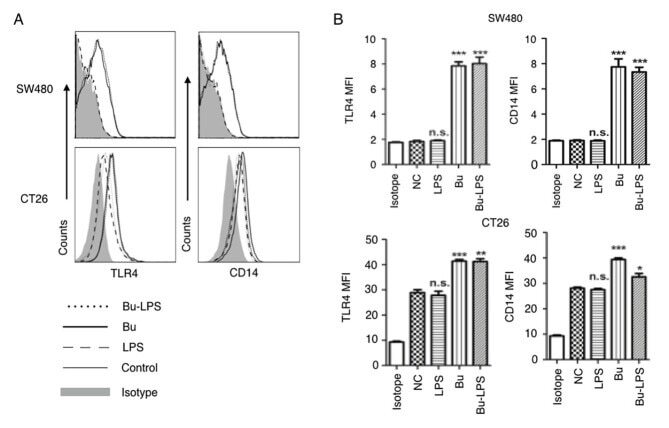
- Figure 4. Butyrate upregulates the levels of TLR4 and CD14 on colon cancer cells. The expression levels of TLR4 and CD14 on the membrane of SW480 cells and CT26 cells treated by butyrate and/or LPS were (A) analyzed using a flow cytometer, and (B) the MFI values of TLR4 and CD14 were quantitative analyzed. These experiments were repeated >=3 times, and representative graphs are presented. Compared with the NC group, *P
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
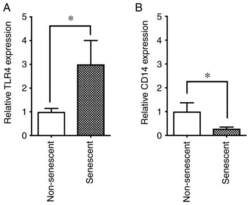
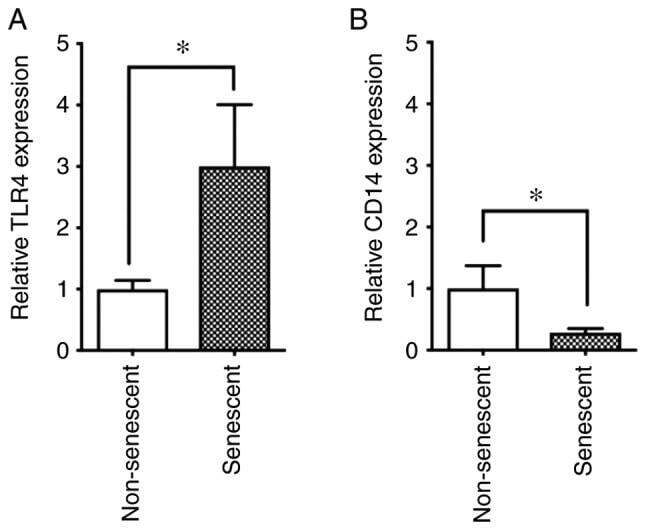
- Figure 4 Expression levels of TLR4 and CD14 in senescent and non-senescent endothelial cells. Cell surface expression levels of the LPS receptors TLR4 and CD14 were analyzed with phycoerythrin-conjugated specific antibodies in senescent and non-senescent HUVECs by flow cytometry. (A) Relative expression of TLR4 and (B) CD14 in senescent cells was expressed as a ratio to non-senescent cells. Data are presented as the mean +- SD of at least three independent experiments. * P
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- Fig 4 Gene expression of biomarkers of injury and immunostaining of TLR4 in patient auricles. (A-D) TLR4 and NOX4 are activated resulting in elevated TNF-alpha in the auricles. Auricles obtained during CABG surgery presented higher expression of TLR4 (P