Explore
Explore Validate
Validate Learn
Learn Western blot
Western blot ELISA
ELISAAntibody data
- Antibody Data
- Antigen structure
- References [16]
- Comments [0]
- Validations
- Western blot [1]
- Immunocytochemistry [2]
- Immunohistochemistry [2]
- Other assay [7]
Submit
Validation data
Reference
Comment
Report error
- Product number
- PA3-102 - Provider product page

- Provider
- Invitrogen Antibodies
- Product name
- NMDAR1 Polyclonal Antibody
- Antibody type
- Polyclonal
- Antigen
- Synthetic peptide
- Description
- PA3-102 detects N-methyl-D-aspartate (NMDA) receptor type 1. PA3-102 has been used successfully in Western blot, ELISA, immunoprecipitation, immunohistochemistry, and immunocytochemistry procedures. In Western blot analysis of rat brain synaptic membranes this antibody detects a ~120 kDa protein representing NMDA receptor type 1. The PA3-102 immunogen is a synthetic peptide corresponding to residues (195)N Y E N L D Q L S Y D N K R G P(210) C of Exon 5 of rat NMDA Receptor Type 1 (Isoforms B, F, G).
- Reactivity
- Human, Mouse, Rat
- Host
- Rabbit
- Isotype
- IgG
- Vial size
- 200 μL
- Concentration
- Conc. Not Determined
- Storage
- -20°C, Avoid Freeze/Thaw Cycles
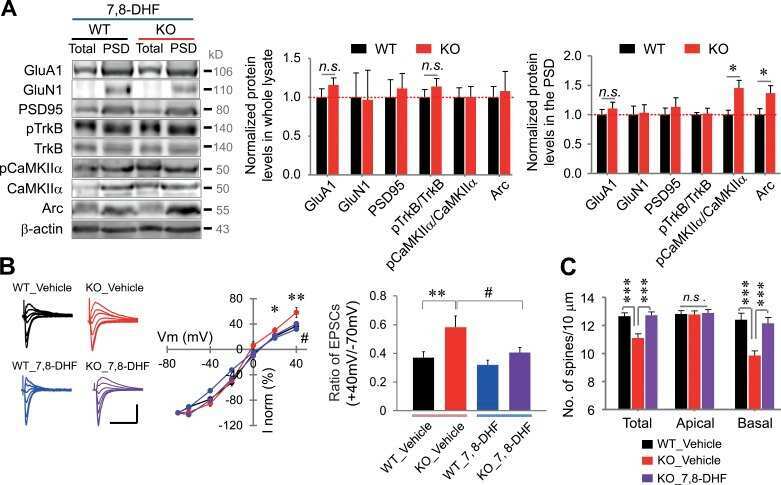
Submitted references Repeated Galvanic Vestibular Stimulation Modified the Neuronal Potential in the Vestibular Nucleus.
Autism-Misregulated eIF4G Microexons Control Synaptic Translation and Higher Order Cognitive Functions.
Glycogen synthase kinase-3 inhibition rescues sex-dependent contextual fear memory deficit in human immunodeficiency virus-1 transgenic mice.
Tissue-type plasminogen activator selectively inhibits multiple toll-like receptors in CSF-1-differentiated macrophages.
SK Channel Modulates Synaptic Plasticity by Tuning CaMKIIα/β Dynamics.
Synaptic proximity enables NMDAR signalling to promote brain metastasis.
Hydrogen sulfide inhibits Ca(2+)-induced mitochondrial permeability transition pore opening in type-1 diabetes.
Cereblon Maintains Synaptic and Cognitive Function by Regulating BK Channel.
Glutamate Deregulation in Ketamine-Induced Psychosis-A Potential Role of PSD95, NMDA Receptor and PMCA Interaction.
Autism-like behavior caused by deletion of vaccinia-related kinase 3 is improved by TrkB stimulation.
Low levels of methyl β-cyclodextrin disrupt GluA1-dependent synaptic potentiation but not synaptic depression.
Sox18 preserves the pulmonary endothelial barrier under conditions of increased shear stress.
Hippocampal long-term potentiation is disrupted during expression and extinction but is restored after reinstatement of morphine place preference.
Hijacking the neuronal NMDAR signaling circuit to promote tumor growth and invasion.
Impacts of forebrain neuronal glycine transporter 1 disruption in the senescent brain: evidence for age-dependent phenotypes in Pavlovian learning.
Selective dendrite-targeting of mRNAs of NR1 splice variants without exon 5: identification of a cis-acting sequence and isolation of sequence-binding proteins.
Kim G, Lee S, Kim KS
Neural plasticity 2020;2020:5743972
Neural plasticity 2020;2020:5743972
Autism-Misregulated eIF4G Microexons Control Synaptic Translation and Higher Order Cognitive Functions.
Gonatopoulos-Pournatzis T, Niibori R, Salter EW, Weatheritt RJ, Tsang B, Farhangmehr S, Liang X, Braunschweig U, Roth J, Zhang S, Henderson T, Sharma E, Quesnel-Vallières M, Permanyer J, Maier S, Georgiou J, Irimia M, Sonenberg N, Forman-Kay JD, Gingras AC, Collingridge GL, Woodin MA, Cordes SP, Blencowe BJ
Molecular cell 2020 Mar 19;77(6):1176-1192.e16
Molecular cell 2020 Mar 19;77(6):1176-1192.e16
Glycogen synthase kinase-3 inhibition rescues sex-dependent contextual fear memory deficit in human immunodeficiency virus-1 transgenic mice.
Moidunny S, Benneyworth MA, Titus DJ, Beurel E, Kolli U, Meints J, Jalodia R, Ramakrishnan S, Atkins CM, Roy S
British journal of pharmacology 2020 Dec;177(24):5658-5676
British journal of pharmacology 2020 Dec;177(24):5658-5676
Tissue-type plasminogen activator selectively inhibits multiple toll-like receptors in CSF-1-differentiated macrophages.
Das L, Azmoon P, Banki MA, Mantuano E, Gonias SL
PloS one 2019;14(11):e0224738
PloS one 2019;14(11):e0224738
SK Channel Modulates Synaptic Plasticity by Tuning CaMKIIα/β Dynamics.
Shrestha A, Sultana R, Lee CC, Ogundele OM
Frontiers in synaptic neuroscience 2019;11:18
Frontiers in synaptic neuroscience 2019;11:18
Synaptic proximity enables NMDAR signalling to promote brain metastasis.
Zeng Q, Michael IP, Zhang P, Saghafinia S, Knott G, Jiao W, McCabe BD, Galván JA, Robinson HPC, Zlobec I, Ciriello G, Hanahan D
Nature 2019 Sep;573(7775):526-531
Nature 2019 Sep;573(7775):526-531
Hydrogen sulfide inhibits Ca(2+)-induced mitochondrial permeability transition pore opening in type-1 diabetes.
Papu John AS, Kundu S, Pushpakumar S, Amin M, Tyagi SC, Sen U
American journal of physiology. Endocrinology and metabolism 2019 Aug 1;317(2):E269-E283
American journal of physiology. Endocrinology and metabolism 2019 Aug 1;317(2):E269-E283
Cereblon Maintains Synaptic and Cognitive Function by Regulating BK Channel.
Choi TY, Lee SH, Kim YJ, Bae JR, Lee KM, Jo Y, Kim SJ, Lee AR, Choi S, Choi LM, Bang S, Song MR, Chung J, Lee KJ, Kim SH, Park CS, Choi SY
The Journal of neuroscience : the official journal of the Society for Neuroscience 2018 Apr 4;38(14):3571-3583
The Journal of neuroscience : the official journal of the Society for Neuroscience 2018 Apr 4;38(14):3571-3583
Glutamate Deregulation in Ketamine-Induced Psychosis-A Potential Role of PSD95, NMDA Receptor and PMCA Interaction.
Lisek M, Ferenc B, Studzian M, Pulaski L, Guo F, Zylinska L, Boczek T
Frontiers in cellular neuroscience 2017;11:181
Frontiers in cellular neuroscience 2017;11:181
Autism-like behavior caused by deletion of vaccinia-related kinase 3 is improved by TrkB stimulation.
Kang MS, Choi TY, Ryu HG, Lee D, Lee SH, Choi SY, Kim KT
The Journal of experimental medicine 2017 Oct 2;214(10):2947-2966
The Journal of experimental medicine 2017 Oct 2;214(10):2947-2966
Low levels of methyl β-cyclodextrin disrupt GluA1-dependent synaptic potentiation but not synaptic depression.
Choi TY, Jung S, Nah J, Ko HY, Jo SH, Chung G, Park K, Jung YK, Choi SY
Journal of neurochemistry 2015 Feb;132(3):276-85
Journal of neurochemistry 2015 Feb;132(3):276-85
Sox18 preserves the pulmonary endothelial barrier under conditions of increased shear stress.
Gross CM, Aggarwal S, Kumar S, Tian J, Kasa A, Bogatcheva N, Datar SA, Verin AD, Fineman JR, Black SM
Journal of cellular physiology 2014 Nov;229(11):1802-16
Journal of cellular physiology 2014 Nov;229(11):1802-16
Hippocampal long-term potentiation is disrupted during expression and extinction but is restored after reinstatement of morphine place preference.
Portugal GS, Al-Hasani R, Fakira AK, Gonzalez-Romero JL, Melyan Z, McCall JG, Bruchas MR, Morón JA
The Journal of neuroscience : the official journal of the Society for Neuroscience 2014 Jan 8;34(2):527-38
The Journal of neuroscience : the official journal of the Society for Neuroscience 2014 Jan 8;34(2):527-38
Hijacking the neuronal NMDAR signaling circuit to promote tumor growth and invasion.
Li L, Hanahan D
Cell 2013 Mar 28;153(1):86-100
Cell 2013 Mar 28;153(1):86-100
Impacts of forebrain neuronal glycine transporter 1 disruption in the senescent brain: evidence for age-dependent phenotypes in Pavlovian learning.
Dubroqua S, Singer P, Boison D, Feldon J, Möhler H, Yee BK
Behavioral neuroscience 2010 Dec;124(6):839-50
Behavioral neuroscience 2010 Dec;124(6):839-50
Selective dendrite-targeting of mRNAs of NR1 splice variants without exon 5: identification of a cis-acting sequence and isolation of sequence-binding proteins.
Pal R, Agbas A, Bao X, Hui D, Leary C, Hunt J, Naniwadekar A, Michaelis ML, Kumar KN, Michaelis EK
Brain research 2003 Dec 19;994(1):1-18
Brain research 2003 Dec 19;994(1):1-18
No comments: Submit comment
Supportive validation
- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
- Western blot was performed using Anti-NMDAR1 Polyclonal Antibody (Product # PA3-102) and a 110kDa band corresponding to NMDAR1 was observed across cell lines and tissues except liver and kidney tissues. Membrane enriched extracts (30 µg lysate) of (Fig. a) SH-SY5Y (Lane 1), U-87 MG (Lane 2), Neuro-2a (Lane 3), HeLa (Lane 4), (Fig. b) Mouse Brain (Lane 1), Rat Brain (Lane 2), Mouse Liver (Lane 3), Rat Liver (Lane 4), Mouse Kidney (Lane 5) were electrophoresed using NuPAGE™ 4-12% Bis-Tris Protein Gel (Product # NP0322BOX). Resolved proteins were then transferred onto a Nitrocellulose membrane (Product # IB23001) by iBlot® 2 Dry Blotting System (Product # IB21001). The blots were probed with the primary antibody (1:600 dilution) and detected by chemiluminescence with Goat anti-Rabbit IgG (Heavy Chain) Superclonal™ Recombinant Secondary Antibody, HRP (Product # A27036, 1:6000 dilution) using the iBright FL 1000 (Product # A32752). Chemiluminescent detection was performed using SuperSignal™ West Dura Extended Duration Substrate (Product # 34076). Few uncharacterized bands were observed between 30-50 kDa.

Supportive validation
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
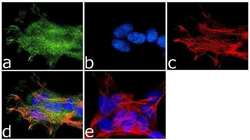
- Immunofluorescence analysis of NMDA Receptor 1 was done on 70% confluent log phase SH-SY5Y cells. The cells were fixed with 4% paraformaldehyde for 10 minutes, permeabilized with 0.1% Triton™ X-100 for 10 minutes, and blocked with 1% BSA for 1 hour at room temperature. The cells were labeled with NMDA Receptor 1 Rabbit Polyclonal Antibody (Product # PA3-102) at 1:250 dilution in 0.1% BSA and incubated for 3 hours at room temperature and then labeled with Goat anti-Rabbit IgG (H+L) Superclonal™ Secondary Antibody, Alexa Fluor® 488 conjugate (Product # A27034) at a dilution of 1:2000 for 45 minutes at room temperature (Panel a: green). Nuclei (Panel b: blue) were stained with SlowFade® Gold Antifade Mountant with DAPI (Product # S36938). F-actin (Panel c: red) was stained with Rhodamine Phalloidin (Product # R415, 1:300). Panel d is a merged image showing membranous localization. Panel e is a no primary antibody control. The images were captured at 60X magnification.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
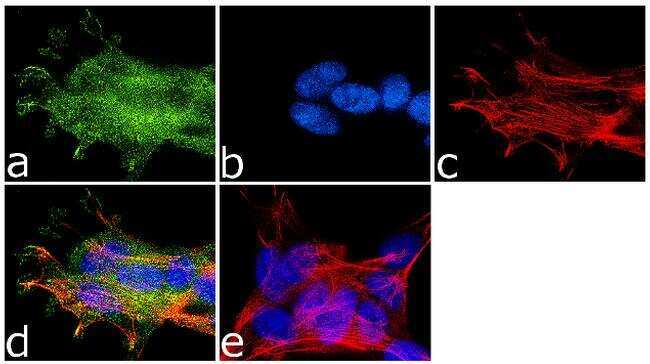
- Immunofluorescence analysis of NMDA Receptor 1 was done on 70% confluent log phase SH-SY5Y cells. The cells were fixed with 4% paraformaldehyde for 10 minutes, permeabilized with 0.1% Triton™ X-100 for 10 minutes, and blocked with 1% BSA for 1 hour at room temperature. The cells were labeled with NMDA Receptor 1 Rabbit Polyclonal Antibody (Product # PA3-102) at 1:250 dilution in 0.1% BSA and incubated for 3 hours at room temperature and then labeled with Goat anti-Rabbit IgG (Heavy Chain) Superclonal™ Secondary Antibody, Alexa Fluor® 488 conjugate (Product # A27034) at a dilution of 1:2000 for 45 minutes at room temperature (Panel a: green). Nuclei (Panel b: blue) were stained with SlowFade® Gold Antifade Mountant with DAPI (Product # S36938). F-actin (Panel c: red) was stained with Rhodamine Phalloidin (Product # R415, 1:300). Panel d is a merged image showing membranous localization. Panel e is a no primary antibody control. The images were captured at 60X magnification.

Supportive validation
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- Immunofluorescence image of NMDA receptor type 1 in rat brain tissue using Product # PA3-102.

- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- Immunofluorescence image of NMDA receptor type 1 in rat brain tissue using Product # PA3-102.

Supportive validation
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
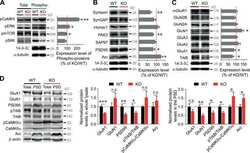
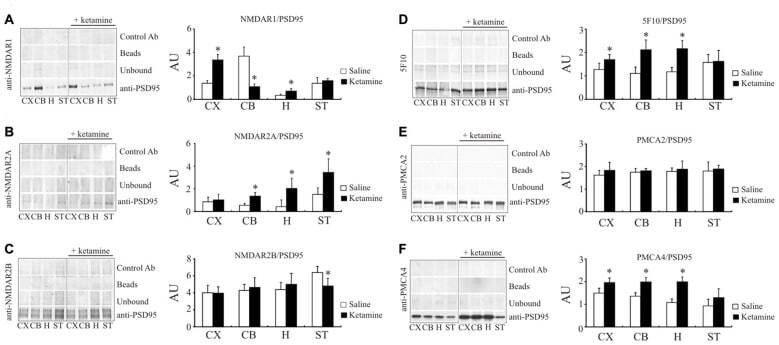
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
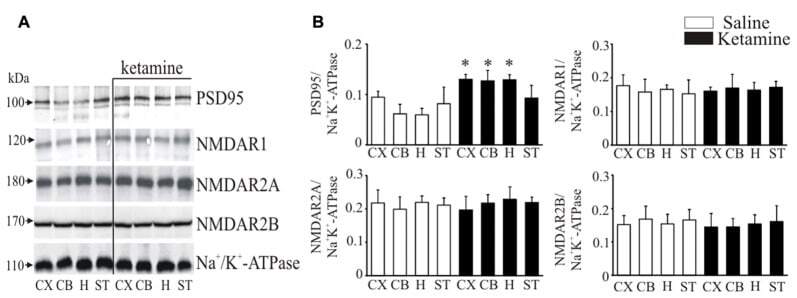
- Figure 6 Interaction of PSD95 with NMDAR subunits and PMCA isoforms. Brain lysates were incubated with anti-PSD95 antibodies conjugated to sepharose beads. Immunocomplexes were recovered and resolved by SDS-PAGE and the membranes were next probed with anti-NMDAR1 (A) , anti-NMDAR2A (B) , anti-NMDAR2B (C) , 5F10 (D) , anti-PMCA2 (E) or anti-PMCA4 (F) antibodies. Negative controls included sepharose-linked secondary antibodies (Control Ab) or sepharose beads only. PSD95 was also immunoprobed in beads-unbound fraction (unbound). Representative blots are shown. The bands intensity were densitometically quantified and the results were expressed as AU defined as the optical density per mg of protein (OD/mg protein). * P < 0.05 ketamine treated vs. saline, n = 5. CX, cortex; CB, cerebellum; H, hippocampus; ST, striatum.
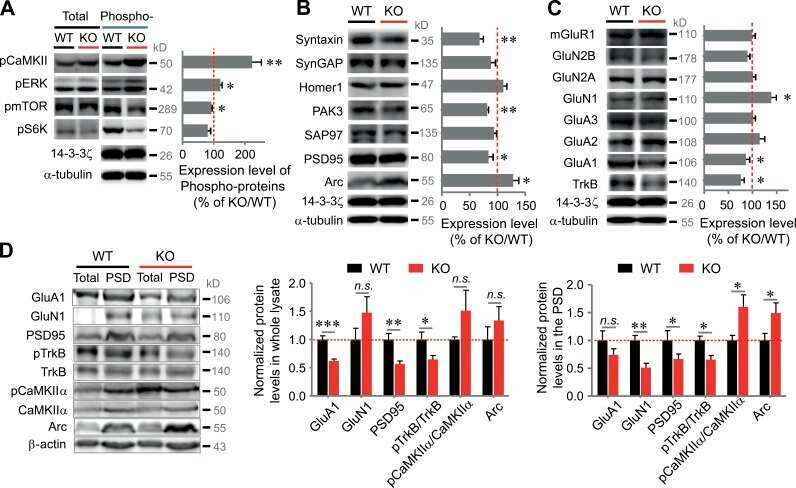
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
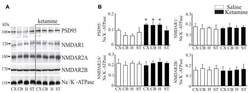
- Experimental details
- Figure 5 Western blot analysis of PSD95 and N-methyl-D-aspartic acid receptor (NMDAR) subunit protein level in synaptosomal membranes. The protein level of PSD95, NMDAR1, NMDAR2A and NMDAR2B was determined by immunoblotting (A) and the bands intensity corresponding to the target protein was densitometrically analyzed (B) . The results are expressed as AU obtained following normalization to endogenous Na + /K + -ATPase level. Representative blots are shown. * P < 0.05 ketamine treated vs. saline, n = 5. CX, cortex; CB, cerebellum; H, hippocampus; ST, striatum.
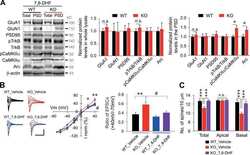
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
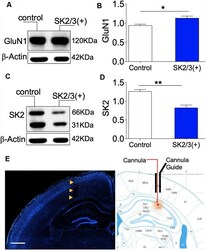
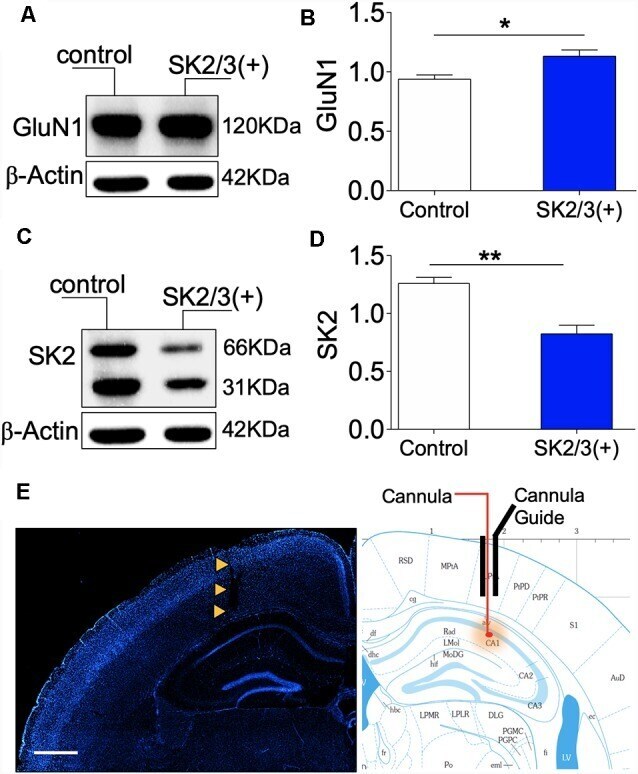
- Figure 7 Activity-dependent expression of SK2 and GluN1. (A,B) Western blot and bar graph demonstrating hippocampal NMDAR-GluN1 expression. (C,D) Representative immunoblot and bar graph illustrating hippocampal SK2 expression. (E) Fluorescence image (DAPI) showing the track of the cannula guide and cannula for intrahippocampal injections. Scale bar = 0.5 mm ( E ; B,D ; * p < 0.05, ** p < 0.01).
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
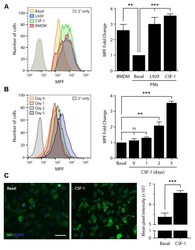
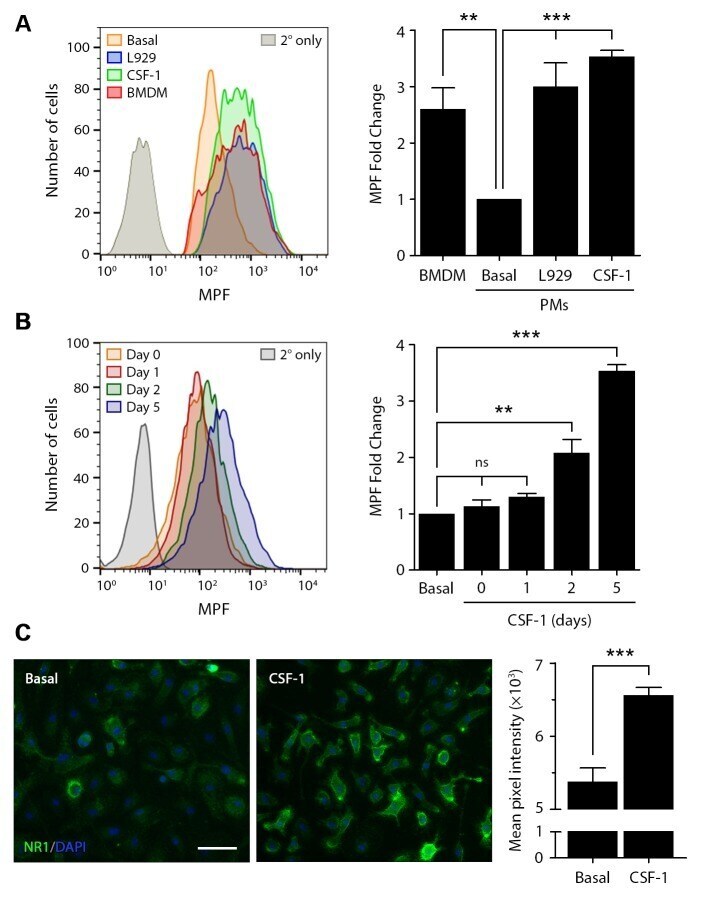
- Fig 5 CSF-1 regulates cell surface NMDA-R in macrophages. Cell surface NMDA-R was labeled in BMDMs and PMs using NR1 subunit-specific primary antibody and Alexa Fluor 647-conjugated secondary antibody. (A) Representative flow cytometry histograms are shown for BMDMs, quiescent PMs (basal), PMs treated with L929 cell-conditioned medium for 5 days (L929), and PMs treated with CSF-1 (10 ng/mL) for 5 days. As a control, PMs were treated with secondary antibody alone (2deg only). The bar graph (right panel) shows the fold increase in the MPF compared with quiescent PMs (no exposure to L929 cell conditioned medium or CSF-1) (mean +- SEM, n = 4; one-way ANOVA with Bonferroni post-hoc test, **p
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
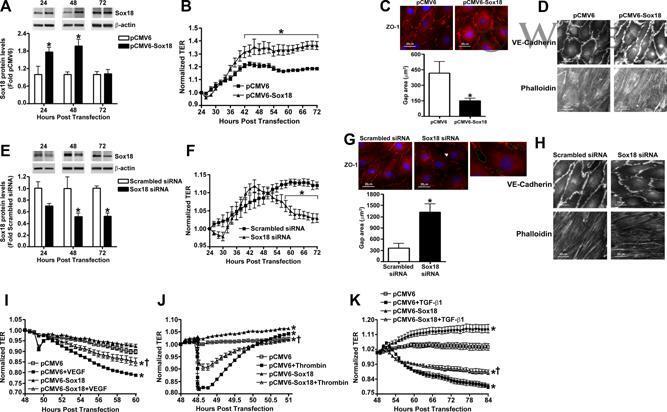
- Sox18 protects endothelial barrier function in pulmonary artery endothelial cells. PAEC were transfected with either pCMV6 or pCMV6-Sox18 for 24-72 h. Immunoblot analysis indicated an increase in Sox18 protein levels after 24 h and 48 h of transfection (A). PAEC monolayers grown on gold microelectrodes (B) or glass coverslips (C,D) were transfected with either pCMV6 or pCMV6-Sox18. After 42 h post transfection, the normalized trans-endothelial resistance (TER) was significantly increased in pCMV6-Sox18 transfected PAEC (B). In addition, immunofluorescent staining of tight junctions was performed using an Alexa Fluor 594 conjugated Zonula occludens-1 (ZO-1) antibody; the nuclei were stained with DAPI. After 48 h, the over-expression of pCMV6-Sox18 decreased inter-cellular gap formation (C). Immunofluorescent analysis of adherens junctions indicated increased VE-cadherin staining but no changes in stress fiber formation (D). In contrast, the siRNA mediated knockdown of Sox18 decreased Sox18 protein levels after 48 and 72 h (E) and the TER after 58 h of transfection (F). Additionally, the immunofluorescent staining of inter-cellular tight junctions demonstrated that, after 48 h, the Sox18 depleted monolayers displayed enhanced gap formation (G). Similarly, these cells displayed a loss of junctional VE-cadherin staining but no alterations in stress fiber formation (H). Further, when the PAEC transfected with pCMV6 were exposed to VEGF (500 ng/mL) (I), thrombin (100 nm) (J), or TG