Explore
Explore Validate
Validate Learn
Learn Western blot
Western blot Immunocytochemistry
ImmunocytochemistryAntibody data
- Antibody Data
- Antigen structure
- References [28]
- Comments [0]
- Validations
- Immunocytochemistry [2]
- Immunohistochemistry [1]
- Other assay [19]
Submit
Validation data
Reference
Comment
Report error
- Product number
- PA5-16291 - Provider product page

- Provider
- Invitrogen Antibodies
- Product name
- GFAP Polyclonal Antibody
- Antibody type
- Polyclonal
- Antigen
- Other
- Description
- PA5-16291 targets Glial Fibrillary Acidic Protein in IHC (P) applications and shows reactivity with Bovine, Chicken, Guinea Pig, Hamster, Human, mouse, Non-human primate, Ovine, and Rat samples. The PA5-16291 immunogen is gFAP isolated from cow spinal cord.
- Reactivity
- Human, Mouse, Rat, Bovine, Chicken/Avian, Guinea Pig, Hamster
- Host
- Rabbit
- Isotype
- IgG
- Vial size
- 500 μL
- Storage
- 4°C
Submitted references An atlas of endogenous DNA double-strand breaks arising during human neural cell fate determination.
Multichannel optogenetics combined with laminar recordings for ultra-controlled neuronal interrogation.
Chlamydia pneumoniae can infect the central nervous system via the olfactory and trigeminal nerves and contributes to Alzheimer's disease risk.
Ependymal and Neural Stem Cells of Adult Molly Fish (Poecilia sphenops, Valenciennes, 1846) Brain: Histomorphometry, Immunohistochemical, and Ultrastructural Studies.
Extracellular Vesicles From 3xTg-AD Mouse and Alzheimer's Disease Patient Astrocytes Impair Neuroglial and Vascular Components.
Mesenchymal stem cell-derived extracellular vesicles ameliorate Alzheimer's disease-like phenotypes in a preclinical mouse model.
METTL3-dependent RNA m(6)A dysregulation contributes to neurodegeneration in Alzheimer's disease through aberrant cell cycle events.
VPS35 D620N knockin mice recapitulate cardinal features of Parkinson's disease.
Mfn2 Overexpression Attenuates MPTP Neurotoxicity In Vivo.
Delayed microglial depletion after spinal cord injury reduces chronic inflammation and neurodegeneration in the brain and improves neurological recovery in male mice.
Anti-aging effects of Ribes meyeri anthocyanins on neural stem cells and aging mice.
Recombinant Adiponectin Peptide Ameliorates Brain Injury Following Intracerebral Hemorrhage by Suppressing Astrocyte-Derived Inflammation via the Inhibition of Drp1-Mediated Mitochondrial Fission.
Androgen Deprivation Induces Reprogramming of Prostate Cancer Cells to Stem-Like Cells.
CPSF1 mutations are associated with early-onset high myopia and involved in retinal ganglion cell axon projection.
Modulating Expression of Thioredoxin Interacting Protein (TXNIP) Prevents Secondary Damage and Preserves Visual Function in a Mouse Model of Ischemia/Reperfusion.
Zika Virus Targeting in the Developing Brain.
Chronic treatment with resveratrol, a natural polyphenol found in grapes, alleviates oxidative stress and apoptotic cell death in ovariectomized female rats subjected to chronic cerebral hypoperfusion.
Rac1 plays an essential role in axon growth and guidance and in neuronal survival in the central and peripheral nervous systems.
Immunopathological changes in the brain of immunosuppressed mice experimentally infected with Toxocara canis.
Involvement of plasmalogens in post-natal retinal vascular development.
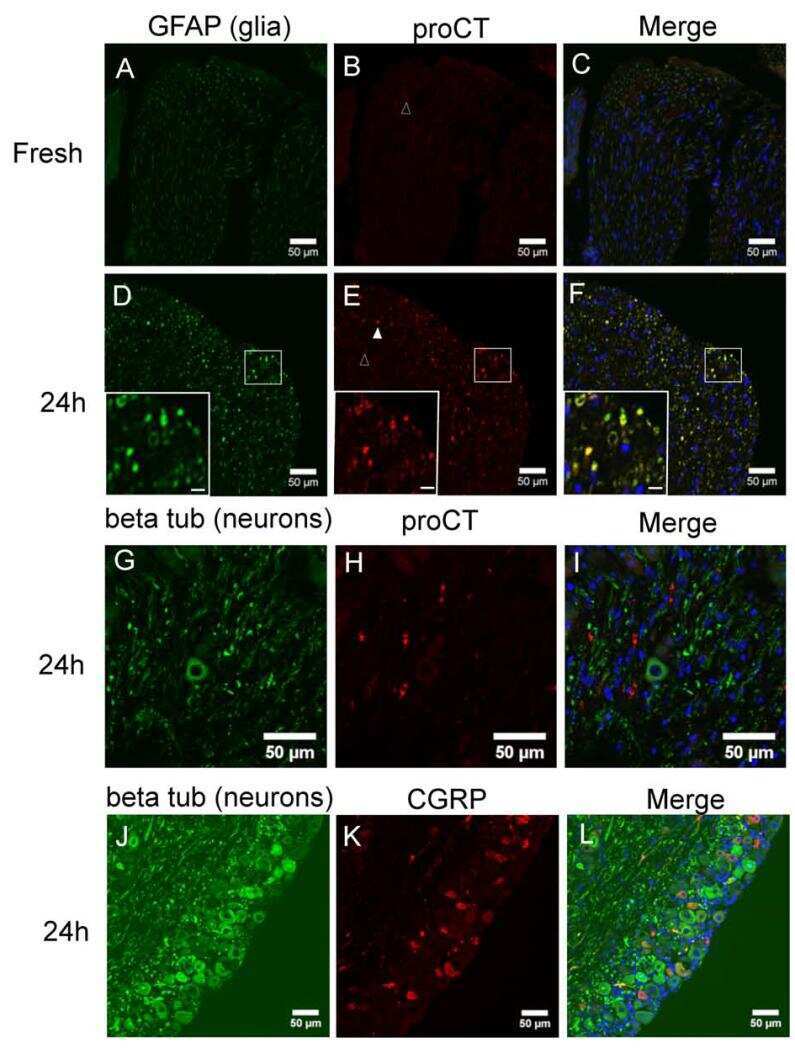
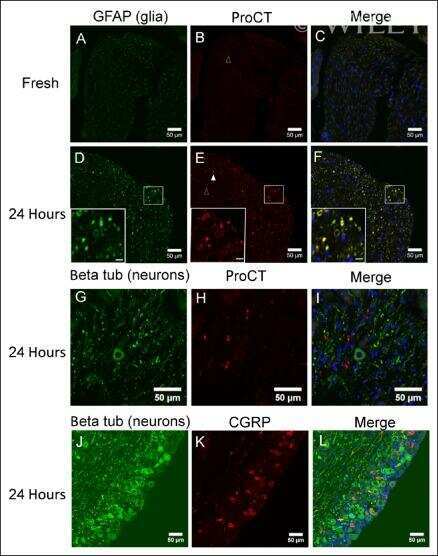
Reactive oxygen species induce procalcitonin expression in trigeminal ganglia glia.
Perinatal inflammation results in decreased oligodendrocyte numbers in adulthood.
Dynamin 2 regulation of integrin endocytosis, but not VEGF signaling, is crucial for developmental angiogenesis.
Romanowsky-Giemsa as a counterstain for immunohistochemistry: optimizing a traditional reagent.
Therapeutic and space radiation exposure of mouse brain causes impaired DNA repair response and premature senescence by chronic oxidant production.
Efficient generation of human iPSCs by a synthetic self-replicative RNA.
Live imaging of mouse endogenous neural progenitors migrating in response to an induced tumor.
DCPIB, a specific inhibitor of volume-regulated anion channels (VRACs), inhibits astrocyte proliferation and cell cycle progression via G1/S arrest.
Ballarino R, Bouwman BAM, Agostini F, Harbers L, Diekmann C, Wernersson E, Bienko M, Crosetto N
Scientific data 2022 Jul 12;9(1):400
Scientific data 2022 Jul 12;9(1):400
Multichannel optogenetics combined with laminar recordings for ultra-controlled neuronal interrogation.
Eriksson D, Schneider A, Thirumalai A, Alyahyay M, de la Crompe B, Sharma K, Ruther P, Diester I
Nature communications 2022 Feb 21;13(1):985
Nature communications 2022 Feb 21;13(1):985
Chlamydia pneumoniae can infect the central nervous system via the olfactory and trigeminal nerves and contributes to Alzheimer's disease risk.
Chacko A, Delbaz A, Walkden H, Basu S, Armitage CW, Eindorf T, Trim LK, Miller E, West NP, St John JA, Beagley KW, Ekberg JAK
Scientific reports 2022 Feb 17;12(1):2759
Scientific reports 2022 Feb 17;12(1):2759
Ependymal and Neural Stem Cells of Adult Molly Fish (Poecilia sphenops, Valenciennes, 1846) Brain: Histomorphometry, Immunohistochemical, and Ultrastructural Studies.
Mokhtar DM, Sayed RKA, Zaccone G, Albano M, Hussein MT
Cells 2022 Aug 27;11(17)
Cells 2022 Aug 27;11(17)
Extracellular Vesicles From 3xTg-AD Mouse and Alzheimer's Disease Patient Astrocytes Impair Neuroglial and Vascular Components.
González-Molina LA, Villar-Vesga J, Henao-Restrepo J, Villegas A, Lopera F, Cardona-Gómez GP, Posada-Duque R
Frontiers in aging neuroscience 2021;13:593927
Frontiers in aging neuroscience 2021;13:593927
Mesenchymal stem cell-derived extracellular vesicles ameliorate Alzheimer's disease-like phenotypes in a preclinical mouse model.
Cone AS, Yuan X, Sun L, Duke LC, Vreones MP, Carrier AN, Kenyon SM, Carver SR, Benthem SD, Stimmell AC, Moseley SC, Hike D, Grant SC, Wilber AA, Olcese JM, Meckes DG Jr
Theranostics 2021;11(17):8129-8142
Theranostics 2021;11(17):8129-8142
METTL3-dependent RNA m(6)A dysregulation contributes to neurodegeneration in Alzheimer's disease through aberrant cell cycle events.
Zhao F, Xu Y, Gao S, Qin L, Austria Q, Siedlak SL, Pajdzik K, Dai Q, He C, Wang W, O'Donnell JM, Tang B, Zhu X
Molecular neurodegeneration 2021 Sep 30;16(1):70
Molecular neurodegeneration 2021 Sep 30;16(1):70
VPS35 D620N knockin mice recapitulate cardinal features of Parkinson's disease.
Niu M, Zhao F, Bondelid K, Siedlak SL, Torres S, Fujioka H, Wang W, Liu J, Zhu X
Aging cell 2021 May;20(5):e13347
Aging cell 2021 May;20(5):e13347
Mfn2 Overexpression Attenuates MPTP Neurotoxicity In Vivo.
Zhao F, Austria Q, Wang W, Zhu X
International journal of molecular sciences 2021 Jan 9;22(2)
International journal of molecular sciences 2021 Jan 9;22(2)
Delayed microglial depletion after spinal cord injury reduces chronic inflammation and neurodegeneration in the brain and improves neurological recovery in male mice.
Li Y, Ritzel RM, Khan N, Cao T, He J, Lei Z, Matyas JJ, Sabirzhanov B, Liu S, Li H, Stoica BA, Loane DJ, Faden AI, Wu J
Theranostics 2020;10(25):11376-11403
Theranostics 2020;10(25):11376-11403
Anti-aging effects of Ribes meyeri anthocyanins on neural stem cells and aging mice.
Gao J, Wu Y, He D, Zhu X, Li H, Liu H, Liu H
Aging 2020 Sep 12;12(17):17738-17753
Aging 2020 Sep 12;12(17):17738-17753
Recombinant Adiponectin Peptide Ameliorates Brain Injury Following Intracerebral Hemorrhage by Suppressing Astrocyte-Derived Inflammation via the Inhibition of Drp1-Mediated Mitochondrial Fission.
Wu X, Luo J, Liu H, Cui W, Guo K, Zhao L, Bai H, Guo W, Guo H, Feng D, Qu Y
Translational stroke research 2020 Oct;11(5):924-939
Translational stroke research 2020 Oct;11(5):924-939
Androgen Deprivation Induces Reprogramming of Prostate Cancer Cells to Stem-Like Cells.
Sánchez BG, Bort A, Vara-Ciruelos D, Díaz-Laviada I
Cells 2020 Jun 10;9(6)
Cells 2020 Jun 10;9(6)
CPSF1 mutations are associated with early-onset high myopia and involved in retinal ganglion cell axon projection.
Ouyang J, Sun W, Xiao X, Li S, Jia X, Zhou L, Wang P, Zhang Q
Human molecular genetics 2019 Jun 15;28(12):1959-1970
Human molecular genetics 2019 Jun 15;28(12):1959-1970
Modulating Expression of Thioredoxin Interacting Protein (TXNIP) Prevents Secondary Damage and Preserves Visual Function in a Mouse Model of Ischemia/Reperfusion.
Coucha M, Shanab AY, Sayed M, Vazdarjanova A, El-Remessy AB
International journal of molecular sciences 2019 Aug 15;20(16)
International journal of molecular sciences 2019 Aug 15;20(16)
Zika Virus Targeting in the Developing Brain.
van den Pol AN, Mao G, Yang Y, Ornaghi S, Davis JN
The Journal of neuroscience : the official journal of the Society for Neuroscience 2017 Feb 22;37(8):2161-2175
The Journal of neuroscience : the official journal of the Society for Neuroscience 2017 Feb 22;37(8):2161-2175
Chronic treatment with resveratrol, a natural polyphenol found in grapes, alleviates oxidative stress and apoptotic cell death in ovariectomized female rats subjected to chronic cerebral hypoperfusion.
Ozacmak VH, Sayan-Ozacmak H, Barut F
Nutritional neuroscience 2016 May;19(4):176-86
Nutritional neuroscience 2016 May;19(4):176-86
Rac1 plays an essential role in axon growth and guidance and in neuronal survival in the central and peripheral nervous systems.
Hua ZL, Emiliani FE, Nathans J
Neural development 2015 Sep 23;10:21
Neural development 2015 Sep 23;10:21
Immunopathological changes in the brain of immunosuppressed mice experimentally infected with Toxocara canis.
Eid MM, El-Kowrany SI, Othman AA, El Gendy DI, Saied EM
The Korean journal of parasitology 2015 Feb;53(1):51-8
The Korean journal of parasitology 2015 Feb;53(1):51-8
Involvement of plasmalogens in post-natal retinal vascular development.
Saab S, Buteau B, Leclère L, Bron AM, Creuzot-Garcher CP, Bretillon L, Acar N
PloS one 2014;9(6):e101076
PloS one 2014;9(6):e101076
Reactive oxygen species induce procalcitonin expression in trigeminal ganglia glia.
Raddant AC, Russo AF
Headache 2014 Mar;54(3):472-84
Headache 2014 Mar;54(3):472-84
Perinatal inflammation results in decreased oligodendrocyte numbers in adulthood.
Graf AE, Haines KM, Pierson CR, Bolon BN, Houston RH, Velten M, Heyob KM, Rogers LK
Life sciences 2014 Jan 17;94(2):164-71
Life sciences 2014 Jan 17;94(2):164-71
Dynamin 2 regulation of integrin endocytosis, but not VEGF signaling, is crucial for developmental angiogenesis.
Lee MY, Skoura A, Park EJ, Landskroner-Eiger S, Jozsef L, Luciano AK, Murata T, Pasula S, Dong Y, Bouaouina M, Calderwood DA, Ferguson SM, De Camilli P, Sessa WC
Development (Cambridge, England) 2014 Apr;141(7):1465-72
Development (Cambridge, England) 2014 Apr;141(7):1465-72
Romanowsky-Giemsa as a counterstain for immunohistochemistry: optimizing a traditional reagent.
Stefanović D, Stefanović M, Nikin Z
Biotechnic & histochemistry : official publication of the Biological Stain Commission 2013 Aug;88(6):329-35
Biotechnic & histochemistry : official publication of the Biological Stain Commission 2013 Aug;88(6):329-35
Therapeutic and space radiation exposure of mouse brain causes impaired DNA repair response and premature senescence by chronic oxidant production.
Suman S, Rodriguez OC, Winters TA, Fornace AJ Jr, Albanese C, Datta K
Aging 2013 Aug;5(8):607-22
Aging 2013 Aug;5(8):607-22
Efficient generation of human iPSCs by a synthetic self-replicative RNA.
Yoshioka N, Gros E, Li HR, Kumar S, Deacon DC, Maron C, Muotri AR, Chi NC, Fu XD, Yu BD, Dowdy SF
Cell stem cell 2013 Aug 1;13(2):246-54
Cell stem cell 2013 Aug 1;13(2):246-54
Live imaging of mouse endogenous neural progenitors migrating in response to an induced tumor.
Elvira G, García I, Benito M, Gallo J, Desco M, Penadés S, Garcia-Sanz JA, Silva A
PloS one 2012;7(9):e44466
PloS one 2012;7(9):e44466
DCPIB, a specific inhibitor of volume-regulated anion channels (VRACs), inhibits astrocyte proliferation and cell cycle progression via G1/S arrest.
He D, Luo X, Wei W, Xie M, Wang W, Yu Z
Journal of molecular neuroscience : MN 2012 Feb;46(2):249-57
Journal of molecular neuroscience : MN 2012 Feb;46(2):249-57
No comments: Submit comment
Supportive validation
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
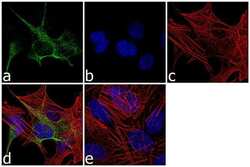
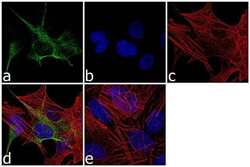
- Experimental details
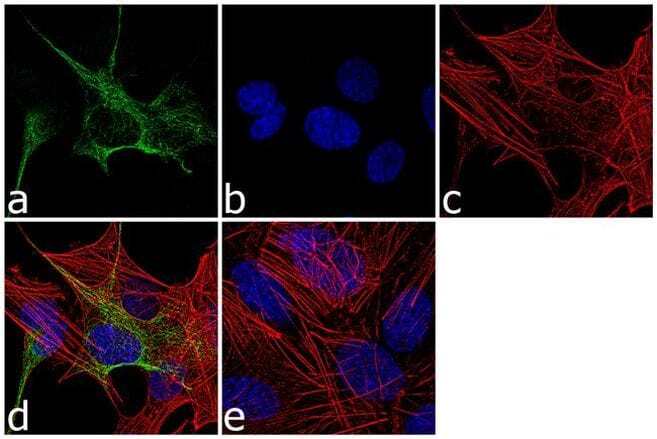
- Immunofluorescence analysis of GFAP was performed using 70% confluent log phase SH-SY5Y cells. The cells were fixed with 4% paraformaldehyde for 10 minutes, permeabilized with 0.1% Triton™ X-100 for 10 minutes, and blocked with 1% BSA for 1 hour at room temperature. The cells were labeled with GFAP Rabbit Polyclonal Antibody (Product # PA5-16291) at 1:250 dilution in 0.1% BSA and incubated for 3 hours at room temperature and then labeled with Goat anti-Rabbit IgG (H+L) Superclonal™ Secondary Antibody, Alexa Fluor® 488 conjugate (Product # A27034) at a dilution of 1:2000 for 45 minutes at room temperature (Panel a: green). Nuclei (Panel b: blue) were stained with SlowFade® Gold Antifade Mountant with DAPI (Product # S36938). F-actin (Panel c: red) was stained with Alexa Fluor® 555 Rhodamine Phalloidin (Product # R415, 1:300). Panel d represents the merged image showing cytoplasmic localization. Panel e shows the no primary antibody control. The images were captured at 60X magnification.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
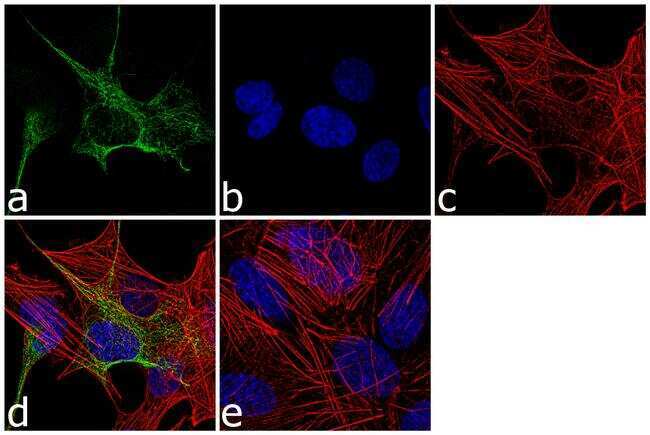
- Experimental details
- Immunofluorescence analysis of GFAP was performed using 70% confluent log phase SH-SY5Y cells. The cells were fixed with 4% paraformaldehyde for 10 minutes, permeabilized with 0.1% Triton™ X-100 for 10 minutes, and blocked with 1% BSA for 1 hour at room temperature. The cells were labeled with GFAP Rabbit Polyclonal Antibody (Product # PA5-16291) at 1:250 dilution in 0.1% BSA and incubated for 3 hours at room temperature and then labeled with Goat anti-Rabbit IgG (Heavy Chain) Superclonal™ Secondary Antibody, Alexa Fluor® 488 conjugate (Product # A27034) at a dilution of 1:2000 for 45 minutes at room temperature (Panel a: green). Nuclei (Panel b: blue) were stained with SlowFade® Gold Antifade Mountant with DAPI (Product # S36938). F-actin (Panel c: red) was stained with Alexa Fluor® 555 Rhodamine Phalloidin (Product # R415, 1:300). Panel d represents the merged image showing cytoplasmic localization. Panel e shows the no primary antibody control. The images were captured at 60X magnification.
Supportive validation
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
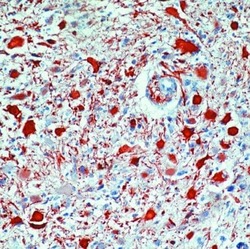
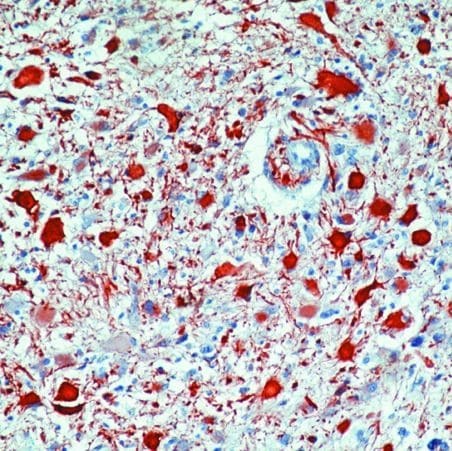
- Formalin-fixed, paraffin-embedded human astrocytoma stained with GFAP antibody using peroxidase-conjugate and AEC chromogen. Note cytoplasmic staining of tumor cells.
Supportive validation
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
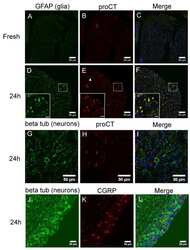
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
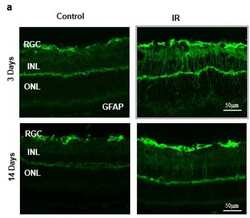
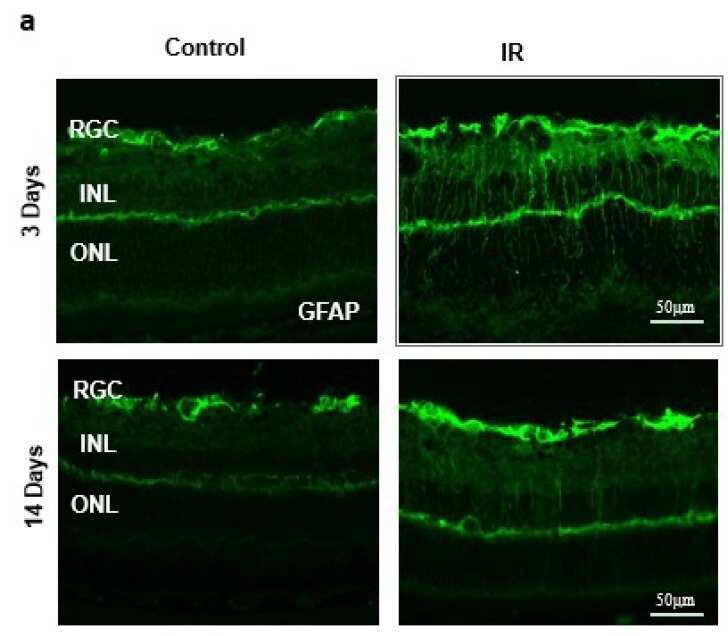
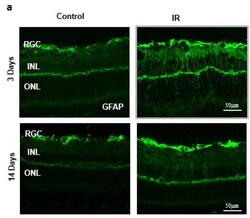
- Figure 2 ( a ) Ischemia/Reperfusion (IR) induced strong activation of Muller cells, the main glia in the retina, assessed by GFAP radial staining after 3-days and persisted for 14-days after ischemia. ( b ) Immunostaining studies using anti-TXNIP (green), anti-glutamine synthetase (GS, red) showed prominent colocalization (yellow) of TXNIP within Muller cells in response to IR when compared to shams.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
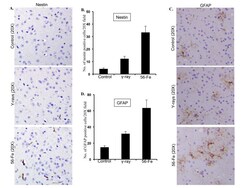
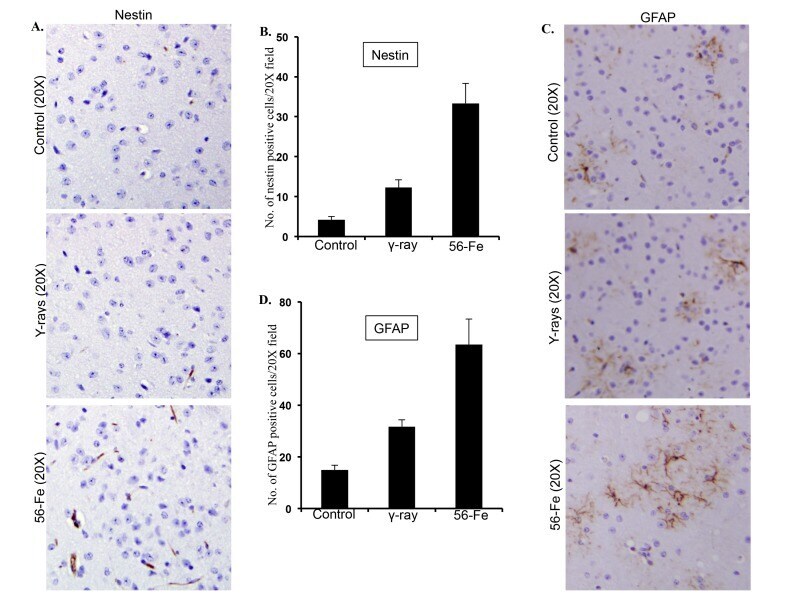
- Figure 5 Assessing reactive gliosis twelve months after radiation exposure. ( A ) Comparing immunohistochemical staining of nestin in cerebral cortex after radiation. ( B ) Quantification of nestin staining in cerebral cortex presented as mean +- SEM. ( C ) Comparing immunohistochemical staining of GFAP in cerebral cortex after radiation. ( D ) Quantification of GFAP staining in cerebral cortex presented as mean +- SEM.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
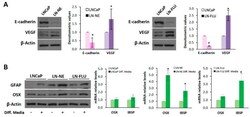
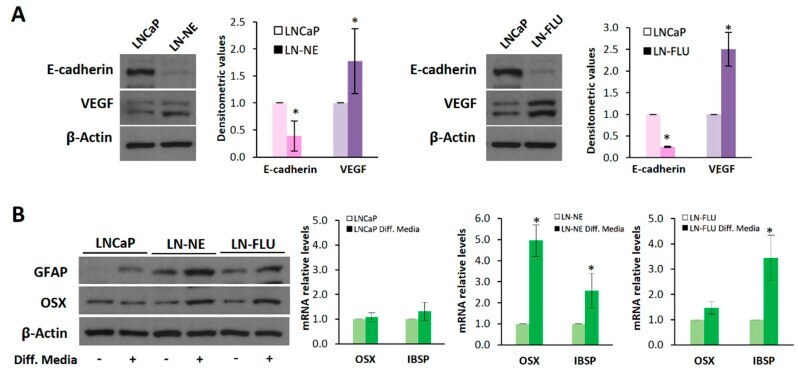
- Figure 2 The prostate cell lines LN-NE and LN-FLU display phenotypic features of stem cells. ( A ) Levels of the epithelial marker E-cadherin and the proangiogenic factor VEGF in androgen-depleted cell lines determined by Western blot and limited to actin as a load control. A representative image of three experiments is shown. Densitometric analysis (n = 3) is shown on the right. ( B ) LNCaP, LN-NE, and LN-FLU cells were incubated in differentiation media for 15 days, and levels of the glial marker GFAP or the osteoblast marker Osx were determined by Western blot. Levels of expression of the osteoblast markers OSX and IBSP determined by qPCR. The data show the relative mRNA expression to GAPDH, which was used as a housekeeping gene. Data represent the mean +- SD of two independent experiments. * p < 0.05 significant difference between LNCaP and LN-NE cells or LNCaP and LN-FLU cells by two-way ANOVA and Sidak's multiple comparisons test.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
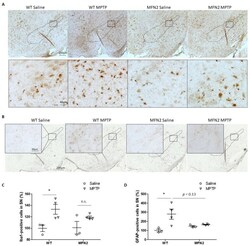
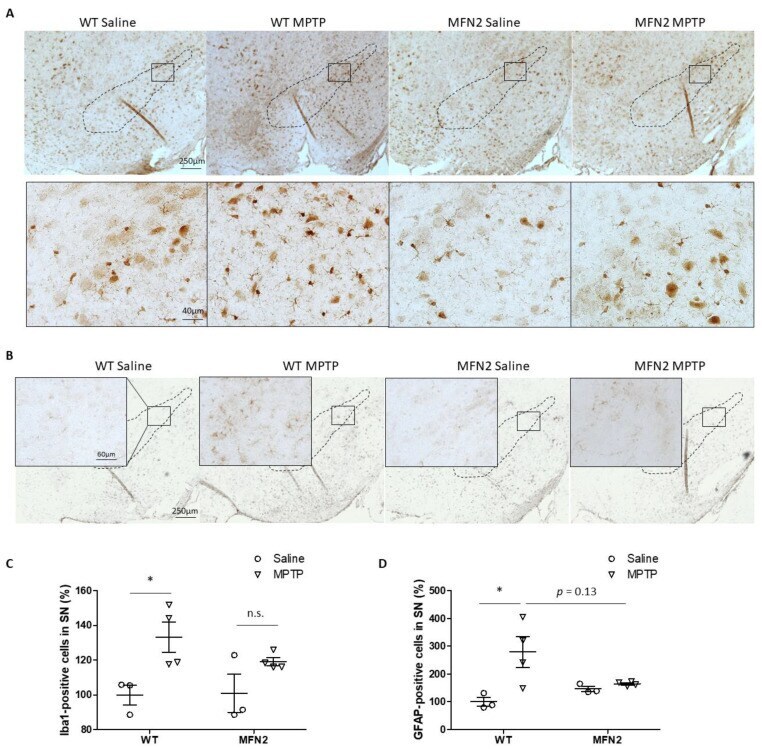
- Figure 4 Mfn2 overexpression inhibited MPTP-induced astrocytosis and microgliosis in the SN. Fixed brains of MPTP- or saline-treated mice were immunostained for iba1 and GFAP. Representative pictures of microglia ( A ) and astrocytes ( B ) are shown. Quantification of the relative number of microglia ( C ) and astrocytes ( D ) in the SN (dashed line) is shown ( n = 4; * p < 0.05 compared to the control. n.s., not significant).
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
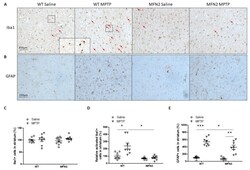
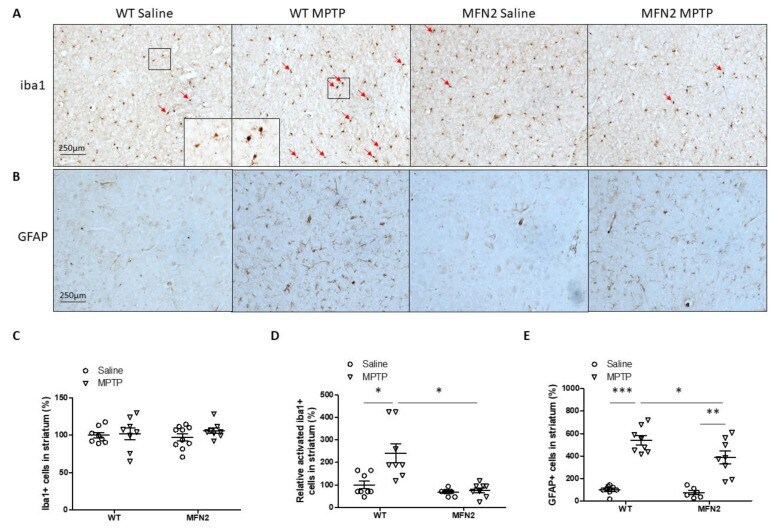
- Figure 7 Mfn2 overexpression inhibited MPTP-induced inflammation in the striatum. Representative pictures of iba1 ( A ) staining in the striatum and quantification of total ( C ) or relative activated ( D ) microglia in the striatum are shown (Red arrows indicate activated microglia). Representative pictures of GFAP staining in the striatum ( B ) and quantification of astrocytes ( E ) in the striatum are shown ( n = 4; * p < 0.05, ** p < 0.01, and *** p < 0.001 compared to the control).
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
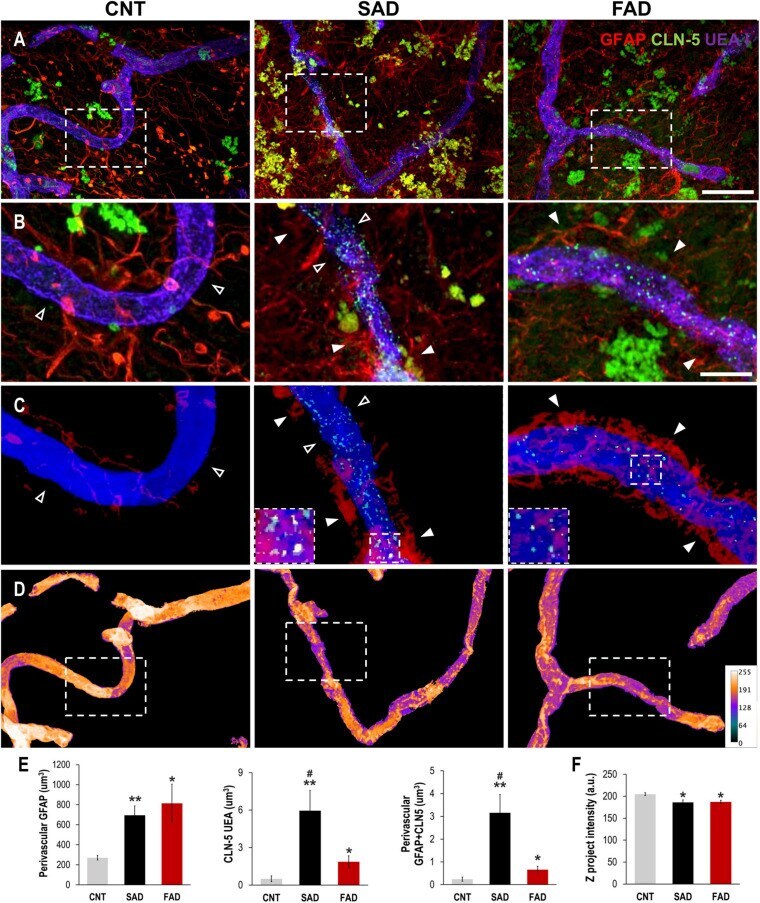
- FIGURE 8 Perivascular GFAP processes are related to altered CLN-5 expression and structural deterioration of the brain vessels in human AD. (A) Representative images of triple immunolabeling of GFAP, CLN-5 and the endothelial marker UEA I showing increased astrocytic reactivity near the vessels, and augmented CLN-5 expression in SAD and FAD. (B) 2X magnifications of the insets in panel (A) highlighting the augment of both perivascular GFAP processes and vascular CLN-5 in AD. (C) Segmented images of GFAP processes present between 0 and 2 mum away from the vessel surface showing the proximity of CLN-5 with vascular areas covered by GFAP. White arrowheads indicate regions where high GFAP vascular coverage coincides with high CLN-5 staining, while empty arrowheads point to areas where both GFAP and CLN-5 are scarce. The insets are 2X magnifications of double positive GFAP-CLN-5 areas. (D) Heat map representation of superposed UEA I signals indicating structural alteration as intensity values go down. Note that the boxes in panel (D) that are enclosing vascular regions with low intensity values, correspond to the images shown in panel (C) which display increased GFAP and CLN-5. (E) Quantification of the perivascular volume of GFAP, vascular CLN-5, and perivascular double positive GFAP-CLN-5. * Comparison with CNT, levels of significance were set to * p < 0.05 and ** p < 0.01. # Comparison SAD with FAD, levels of significance were set to # p < 0.05. (F) Quantification of the Z proj
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
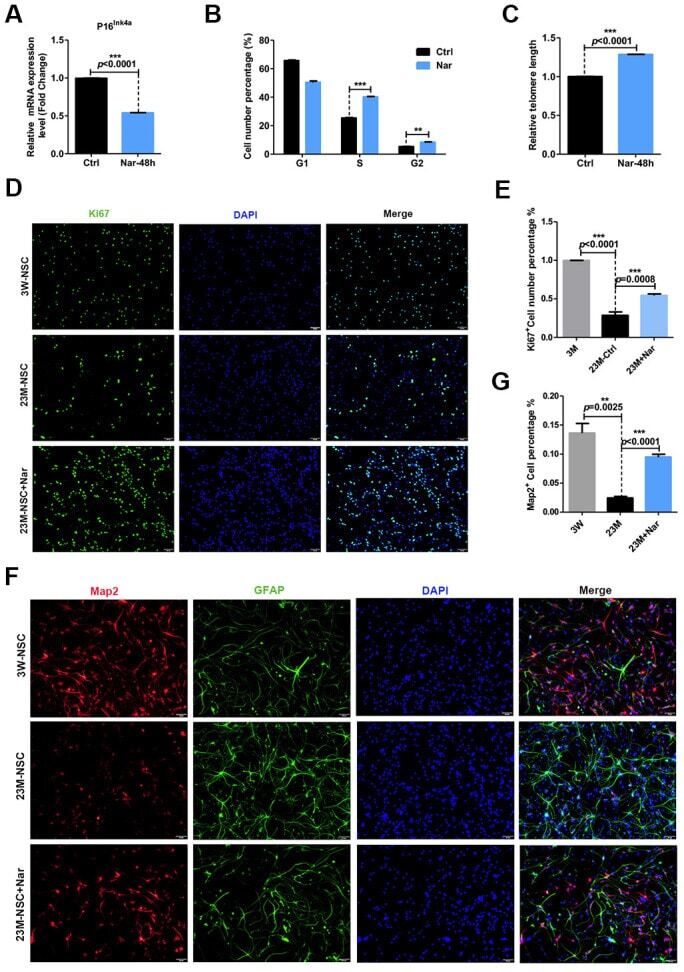
- Figure 3 Effect of naringenin (Nar) on senescence of mouse neural stem cells (mNSCs). ( A ) p16 ink4a mRNA expression was measured by qRT-PCR in 23M-NSCs with and without treatment of 6.8 mug/mL Nar for 48 h. ( B ) Cell cycle phase distributions of 23M-NSCs treated with Nar for 48 h and control cells. ( C ) The relative telomere length of 23M-NSCs increased significantly with Nar treatment. ( D ) Immunofluorescence Ki67 staining of mNSCs treated with Nar for 48 h, with DAPI nuclear labeling. ( E ) Quantification of ( D ). ( F ) Representative fields of MAP2 and GFAP immunofluorescence staining of cultured mNSCs after control and Nar treatment. ( G ) Quantification of ( F ). Data are presented as the mean +- SD of three independent experiments. * P < 0.05, ** P < 0.01, and *** P < 0.0001 compared with untreated cells.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
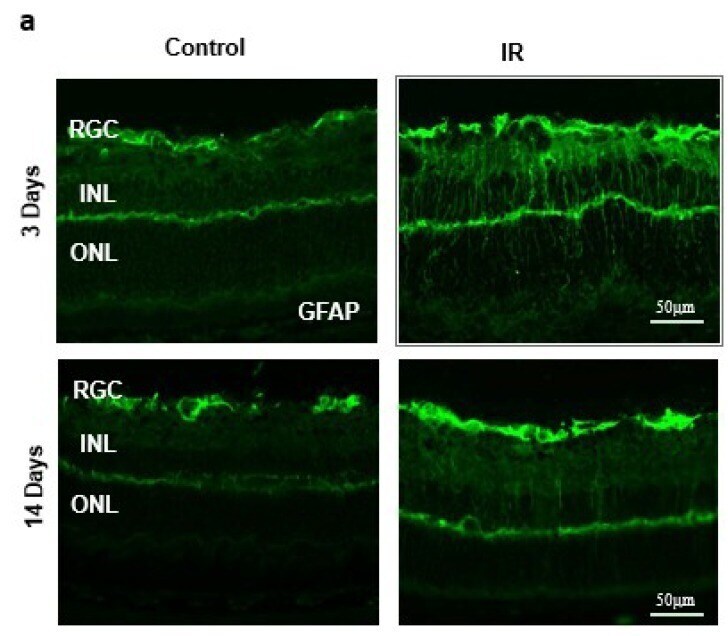
- Figure 2 ( a ) Ischemia/Reperfusion (IR) induced strong activation of Muller cells, the main glia in the retina, assessed by GFAP radial staining after 3-days and persisted for 14-days after ischemia. ( b ) Immunostaining studies using anti-TXNIP (green), anti-glutamine synthetase (GS, red) showed prominent colocalization (yellow) of TXNIP within Muller cells in response to IR when compared to shams.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
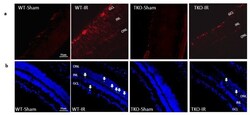
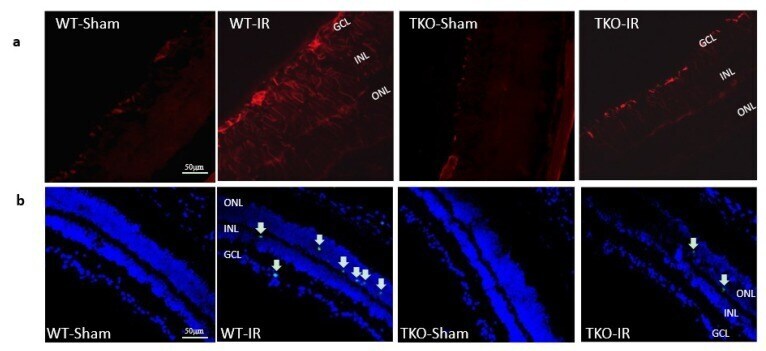
- Figure 5 ( a ) Ischemia/Reperfusion (IR) induced strong activation of Muller cells, the main glia in the retina, assessed by GFAP radial staining after 3-days in WT-IR but not in TKO-IR when compared to shams. ( b ) Ischemia/Reperfusion (IR) sustained retinal cell death indicated by TUNEL-positive cells (arrows) after 3-days in WT-IR, but not in TKO-IR when compared to shams.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
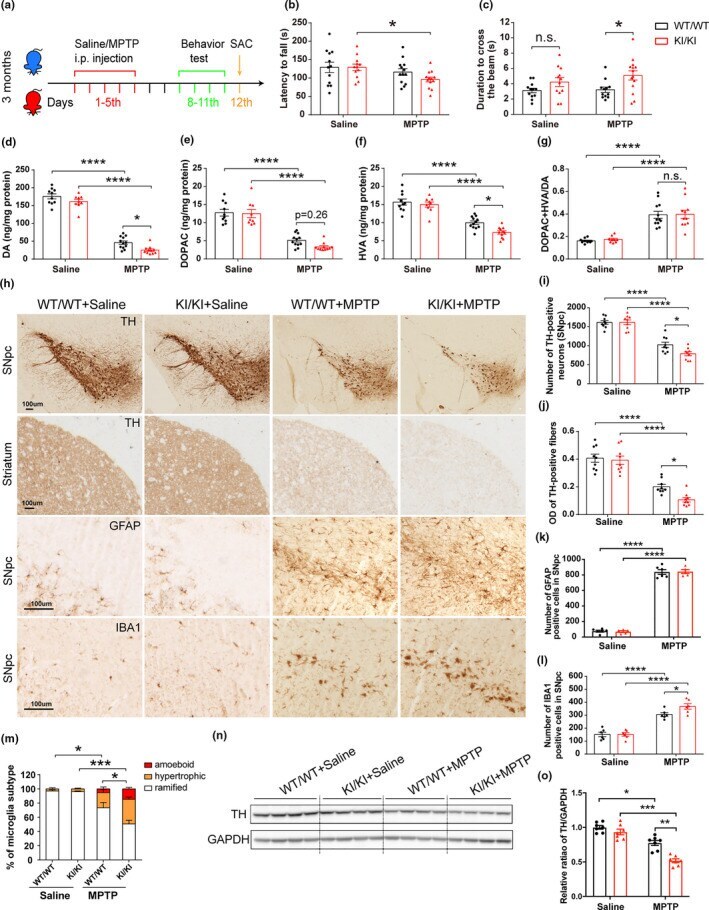
- FIGURE 6 Increased MPTP vulnerability of VPS35 D620N KI mice. (a) A schematic diagram depicts the experimental design. (b-c) Exacerbated motor function deficits in VPS35 D620N / D620N mice as measured by the latency to fall in rotarod test (b) and duration to cross the narrow beam (9 mm) in beam walking test (c) ( n = 12-14/group). (d-g) Concentrations of DA (d), DOPAC (e), HVA (f), and DA turnover rate (DOPAC+HVA/DA) (g) in the STR of VPS35 D620N / D620N and WT controls injected with MPTP/Saline as indicated ( n = 10-12/group). (h) Representative images of TH, GFAP, and IBA1 immunostaining in SNpc and STR from VPS35 D620N / D620N and WT mice injected with MPTP/Saline as indicated. Scale bar, 100 mum. Quantification of TH-positive neurons in SNpc (i), OD of TH-positive fibers in STR (j), GFAP-positive cells (k) and IBA1-positive cells (l,m) in SNpc ( n = 9/group). Representative Western blots (n) and quantification (o) of TH in VM extracts from VPS35 D620N / D620N mice and WT controls injected with MPTP/Saline as indicated ( n = 7/group). Two-way ANOVA with Tukey's post hoc test; mean +- SEM ; * p < 0.05, ** p < 0.01, *** p < 0.001, n.s., not significant
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
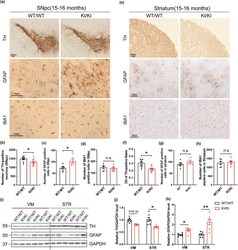
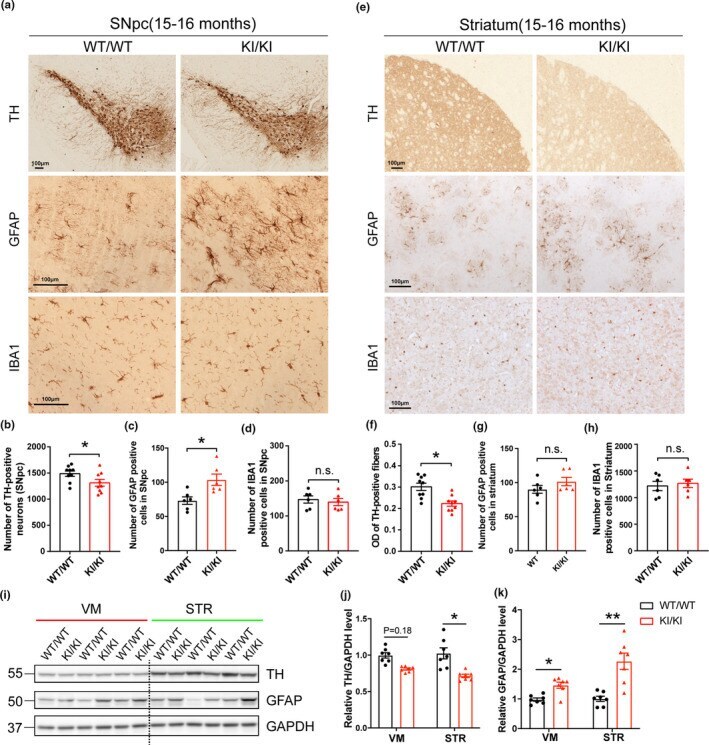
- FIGURE 2 Neuropathological changes in VPS35 D620N KI mice. (a,e) Representative images of TH, GFAP and IBA1 immunostaining in SNpc (a) and STR (e) of 15- to 16-month-old VPS35 D620N / D620N and WT mice ( n = 6-9/group). Scale bar, 100 mum. Quantification of TH-positive neurons (b), GFAP-positive cells (c) and IBA1-positive cells (d) in SNpc. Quantification of optical density (OD) of TH-positive fibers (f), GFAP-positive cells (g), and IBA1-positive cells (h) in STR. (i-k) Representative Western blots (i) and quantification of TH (j) and GFAP (k) in VM and STR extracts from 15- to 16-month-old VPS35 D620N / D620N mice and WT controls ( n = 7/group). Student's t test, unpaired, two-tailed; data are shown as mean +- SEM ; * p < 0.05; ** p < 0.01; n.s., not significant
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
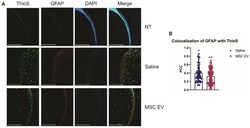
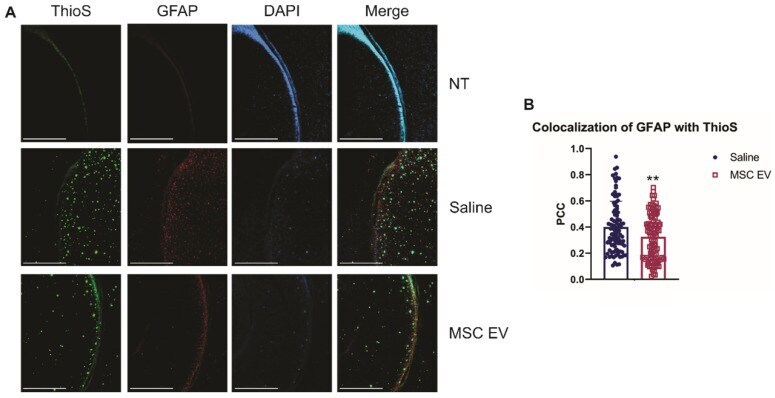
- Figure 6 hMSC-EV treatment decreases colocalization between reactive astrocytes and amyloid plaques. (A) Representative immunohistochemistry images of brain slices stained with ThioS, GFAP, and DAPI. Scale bar = 1 mm. (B) We took 25 one mm 2 images throughout the brain, and Pearson's correlation coefficient (PCC) was obtained through ImageJ analysis to determine colocalization of GFAP and ThioS. **P < 0.01.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
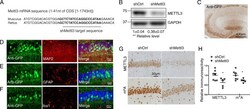
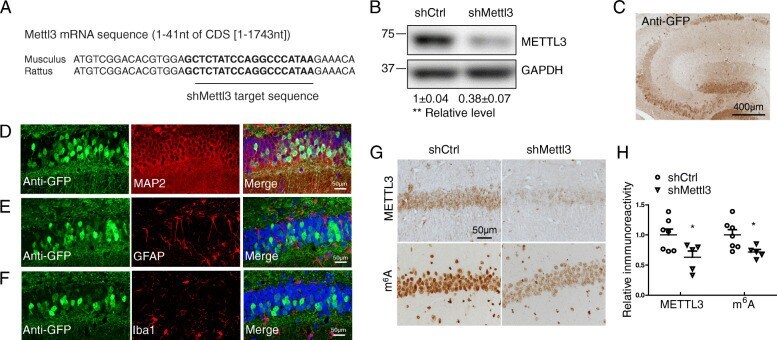
- Fig. 3 Decreased neuronal METTL3 and m 6 A in the hippocampus by AAV-shMettl3. (A) Highly conserved gene sequences of shRNA targeting region in mouse and rat Mettl3 mRNA. (B) Knockdown effect of shMettl3 was confirmed in mouse N2a cells 3 days after transient transfection (Data are means+-SEM from 3 independent experiments). (C) AAV-mediated gene delivery in the hippocampus was confirmed by immunostaining for GFP. WT C57BL/6 mice receiving bilaterally stereotaxic injections of AAV encoding for eGFP plus anti-Mettl3 shRNA (shMettl3) or scrambled control shRNA (shCtrl) into the hippocampus at 2 months of age were analyzed 1 month later. (D-F) Representative images of the AAV-infected areas (GFP-positive) in hippocampus for different cell markers by immunocytochemistry after fixation: MAP 2 for neurons (D), GFAP for astrocytes (E), and Iba1 for microglia (F). (G, H) Representative image of immunoreactivity of METTL3 or m 6 A (G) in AAV-shRNA-injected mice and quantification analysis (H) confirmed that METTL3 protein and m 6 A modification level were reduced in mice injected with shMettl3 virus (n = 5-7 mice per group). GFP staining has been performed in adjacent brain slices at the same time and immunoreactivities of m 6 A and METTL3 were measured in GFP-positive regions. (Data are means+-SEM, * p < 0.05, **p < 0.01, (B, H) unpaired student's t-test)
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
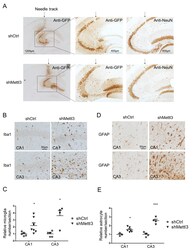
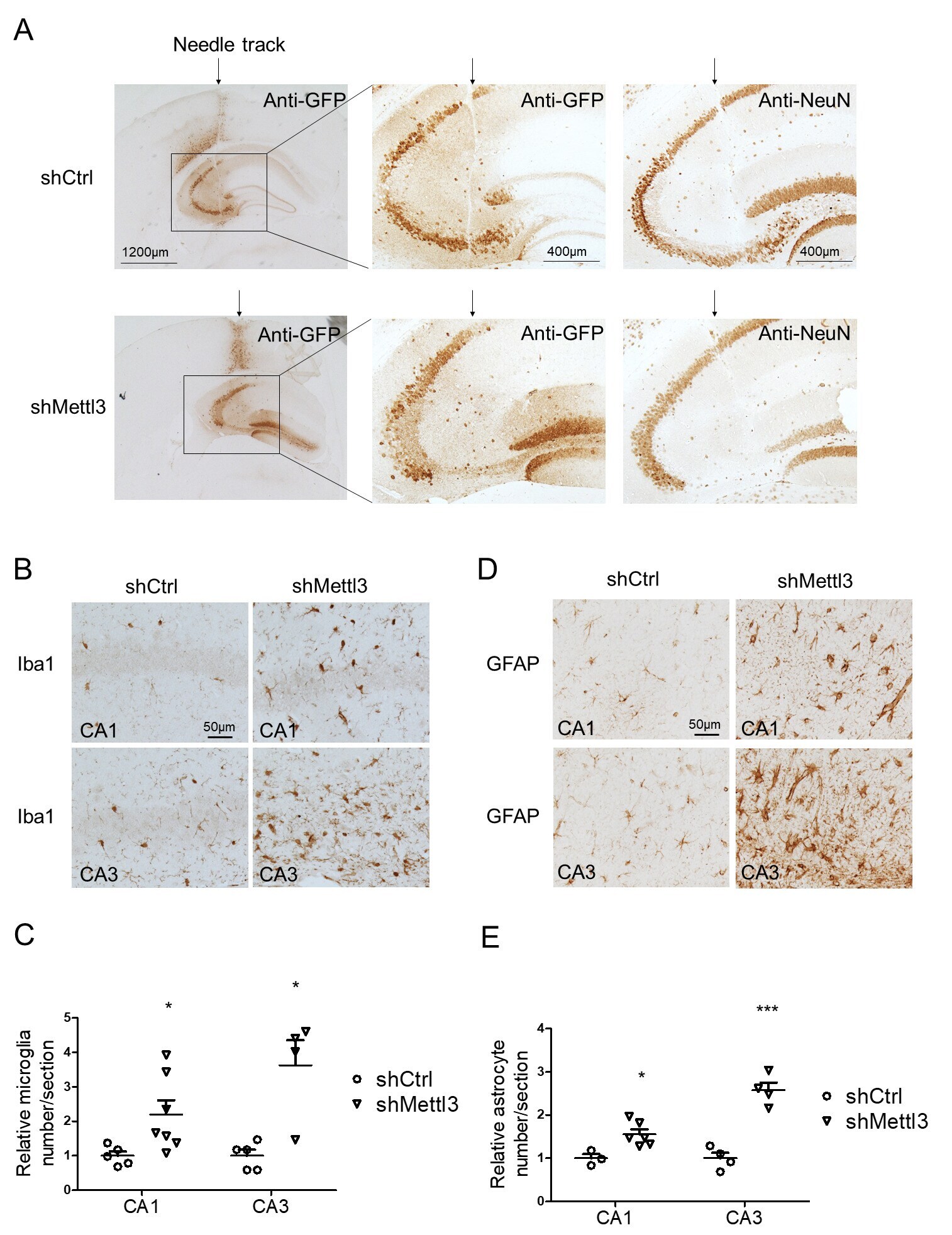
- Additional file 3: Supplementary Fig. 3. Validation of intracranial injection into hippocampus by needle track and induced neuroinflammation by METTL3 depletion in mouse hippocampus. (A) A representative image of needle track (arrow) of AAV-injected mice was shown. GFP immunoreactivity was detected in area adjacent to the needle tack. NeuN staining revealed severe neuronal loss around injected areas only in AAV-shMettl3 injected mice but not AAV-shCtrl injected mice. (B-E) Representative images of immunohistochemistry for Iba1 (B) and GFAP (Thermofisher, D) in hippocampal CA1/2 or CA3 areas in shRNA-injected mice and their quantification (C for astrocyte and E for microglia) analysis showed that METTL3 knockdown caused neuroinflammation in mouse hippocampus. (n = 4-7, *p < 0.5, **p < 0.01; C, E, unpaired student's t-test).
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
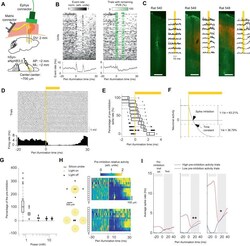
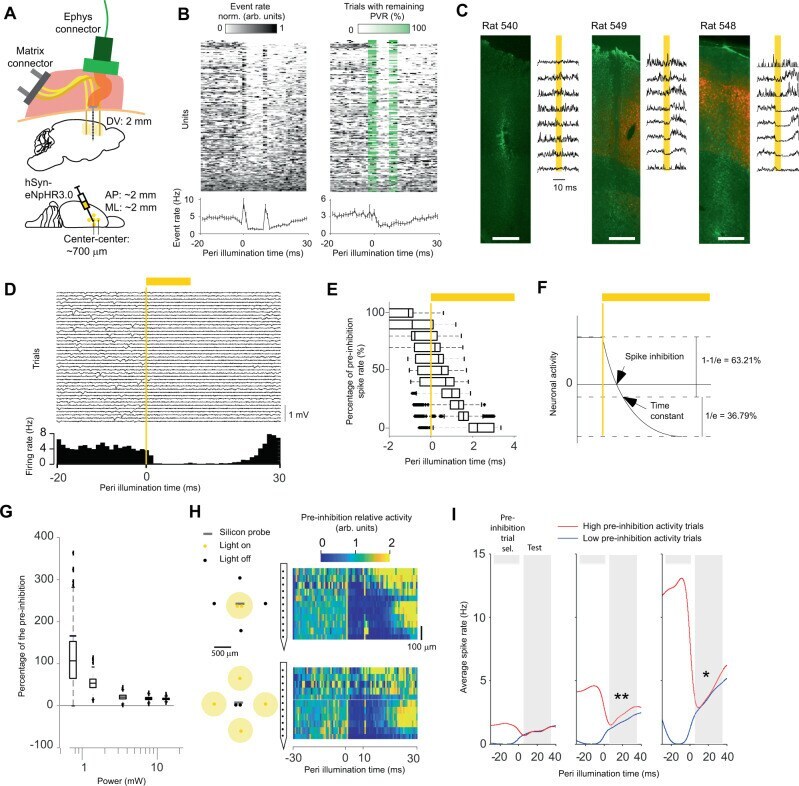
- Ultrafast optogenetic inhibition in freely moving animals. A Injection, stimulation, and recordings in M1. B Activity of sorted units without (left) and with (middle) PVR management. The PVR management algorithm first subtracts an estimate of the PVR and then estimates in which trials the subtraction was incomplete. This remaining PVR trial percentage (green) is then used to weight the average across trials for each time point (lower left and right panel). The data shown are from animals 540, 548, and 549. C Relationship between opsin expression (red) and inhibition strength. To quantify the overall extracellular response, spikes were detected at a threshold of -30 uV on a per-channel basis without PVR management (PVR removed for visibility). GFAP immunostaining (green) was used to identify the BE probe location. Scale bar: 500 um. D Single-trial extracellular traces for 40 inhibitions from one electrode channel (upper panel). Histogram of spikes detected with a threshold of -30 uV (lower panel). E Latency at different percentages of the pre-inhibition spike rate bootstrapped across all sorted units. n = 212 sorted units over two animals. Boxplots: central mark indicates the median, bottom and top edges refer to the 25th and 75th percentiles of the bootstrapped data. F Illustration showing that the spike inhibition latency can be shorter than the time constant of the hyperpolarizing function. G Percentage of pre-inhibition spike rate as a function of the total light power tha
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
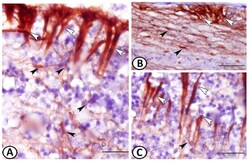
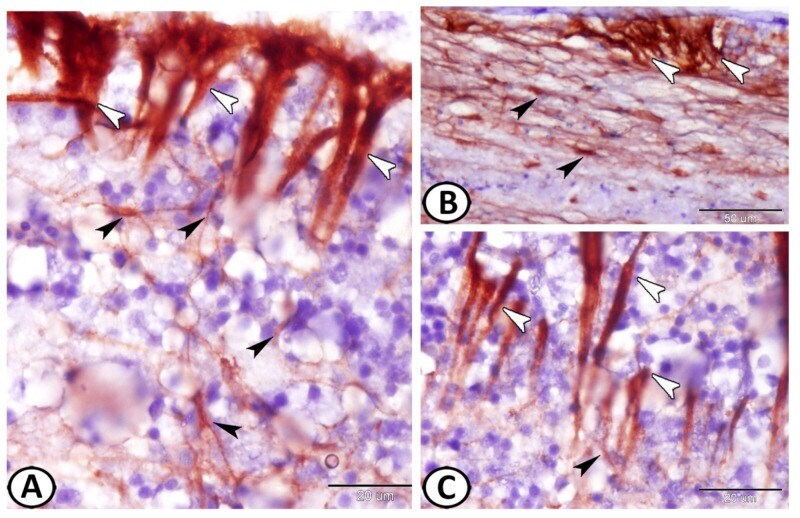
- Figure 3 GFAP immunohistochemistry. ( A - C ) Two types of GFAP immunoreactive cells could be identified: ependymal cells (white arrowheads) and astrocytes (black arrowheads). Note the intense reactions of ECs and the processes of astrocytes that were in direct contact with the processes of the ECs.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
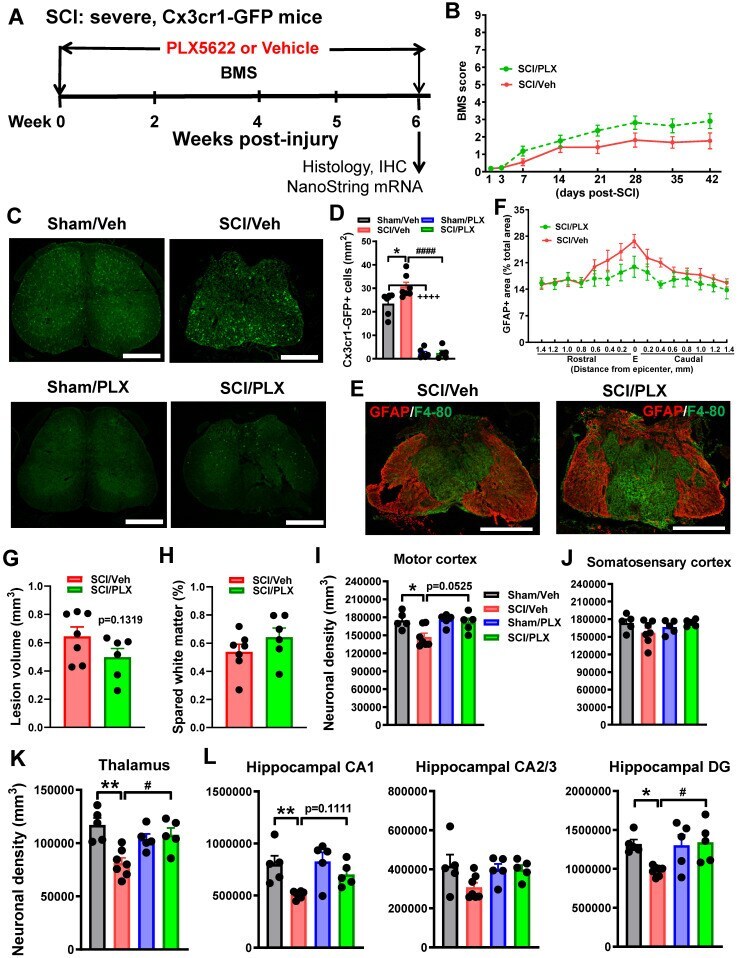
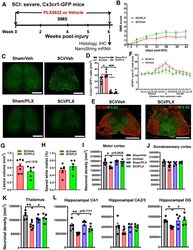
- Figure 5 Depletion of microglia after SCI reduces neurodegeneration in the brain sub-regions. ( A ) Experimental timeline: adult male CX3CR1-GFP mice underwent severe contusion injury at T10. Right after the surgery, PLX5622 (1200 ppm) or Vehicle chow was fed to the mice for the remainder of the study. BMS was recorded on days 1, 3, and weekly up to 6 weeks post-injury. At the 6 weeks post-injury, spinal cord tissue and brain samples were collected for IHC and histological analysis (Stereology). A subset of mice was sacrificed at 6 weeks post-injury and injured spinal cord samples were collected for Nanostring analysis. ( B ) BMS. No significant differences were found in hindlimb locomotor function on the BMS. n = 11 mice/group (2-way ANOVA with repeated measurements following Bonferroni's multiple comparisons test). ( C ) IHC representative images for GFP + cells at 0.4 mm caudal to the epicenter. Scale bar = 500 um. ( D ) IHC GFP + cell counts [n = 6 (Sham/Veh), 7 (SCI/Veh), 6 (Sham/PLX), and 5 (SCI/PLX)]. * p < 0.05, ++++ p < 0.0001, vs. Sham/Veh group. #### p < 0.0001, vs. SCI/Veh group. 2-way ANOVA following Tukey's multiple comparisons test. ( E,F ) Representative images (E) of the lesion epicenter from mice on Vehicle diet and PLX5622 diet at 6 weeks immunostained for GFAP (red) and F4/80 (blue). Scale bar =500 um. Quantification of GFAP+ area (% of total section area) from 1.4 mm rostral or caudal to the lesion epicenter (E). N = 5 mice/group. 2-way ANOVA following Si