Explore
Explore Validate
Validate Learn
Learn Western blot
Western blotAntibody data
- Antibody Data
- Antigen structure
- References [24]
- Comments [0]
- Validations
- Western blot [2]
- Immunocytochemistry [1]
- Immunohistochemistry [3]
- Flow cytometry [1]
- Other assay [5]
Submit
Validation data
Reference
Comment
Report error
- Product number
- MA5-13128 - Provider product page

- Provider
- Invitrogen Antibodies
- Product name
- Cyclin B1 Monoclonal Antibody (V152)
- Antibody type
- Monoclonal
- Antigen
- Other
- Description
- MA5-13128 targets Cyclin B1 in FACS, IHC (P), and WB applications and shows reactivity with Hamster and Human samples.
- Antibody clone number
- V152
- Concentration
- 0.1 mg/mL
Submitted references Menadione reduces CDC25B expression and promotes tumor shrinkage in gastric cancer.
The most abundant maternal lncRNA Sirena1 acts post-transcriptionally and impacts mitochondrial distribution.
Targeted inhibition of metastatic melanoma through interference with Pin1-FOXM1 signaling.
Aurora kinase A is not involved in CPEB1 phosphorylation and cyclin B1 mRNA polyadenylation during meiotic maturation of porcine oocytes.
Cellular distributions of molecules with altered expression specific to thyroid proliferative lesions developing in a rat thyroid carcinogenesis model.
Cyclin B1 is commonly expressed in the cytoplasm of primary human acute myelogenous leukemia cells and serves as a leukemia-associated antigen associated with autoantibody response in a subset of patients.
Cdc6 stability is regulated by the Huwe1 ubiquitin ligase after DNA damage.
Antigenic profiling of glioma cells to generate allogeneic vaccines or dendritic cell-based therapeutics.
The cyclin-dependent kinase inhibitor seliciclib (R-roscovitine; CYC202) decreases the expression of mitotic control genes and prevents entry into mitosis.
HPV16 E1--E4 protein is phosphorylated by Cdk2/cyclin A and relocalizes this complex to the cytoplasm.
Prognostic significance of expressions of cell-cycle regulatory proteins in gastrointestinal stromal tumor and the relevance of the risk grade.
In vitro and in vivo pharmacokinetic-pharmacodynamic relationships for the trisubstituted aminopurine cyclin-dependent kinase inhibitors olomoucine, bohemine and CYC202.
Human papillomavirus type 16 E1 E4-induced G2 arrest is associated with cytoplasmic retention of active Cdk1/cyclin B1 complexes.
The Cyclin-dependent kinase inhibitor CYC202 (R-roscovitine) inhibits retinoblastoma protein phosphorylation, causes loss of Cyclin D1, and activates the mitogen-activated protein kinase pathway.
Complex protein-DNA dynamics at the latent origin of DNA replication of Epstein-Barr virus.
Mitotic cell cycle proteins increase in podocytes despite lack of proliferation.
Citron kinase is a cell cycle-dependent, nuclear protein required for G2/M transition of hepatocytes.
Kinetics of endomitosis in primary murine megakaryocytes.
Regulation of cell cycle and cyclins by 16alpha-hydroxyestrone in MCF-7 breast cancer cells.
Expression of the ALK protein by anaplastic large-cell lymphomas correlates with high proliferative activity.
Expression of the ALK protein by anaplastic large-cell lymphomas correlates with high proliferative activity.
Expression of p34(cdc2) and cyclins A and B compared to other proliferative features of non-Hodgkin's lymphomas: a multivariate cluster analysis.
Expression of p34(cdc2) and cyclins A and B compared to other proliferative features of non-Hodgkin's lymphomas: a multivariate cluster analysis.
Retinoblastoma-related p107 and pRb2/p130 proteins in malignant lymphomas: distinct mechanisms of cell growth control.
Bona AB, Calcagno DQ, Ribeiro HF, Muniz JAPC, Pinto GR, Rocha CAM, Lacreta Junior ACC, de Assumpção PP, Herranz JAR, Burbano RR
Therapeutic advances in gastroenterology 2020;13:1756284819895435
Therapeutic advances in gastroenterology 2020;13:1756284819895435
The most abundant maternal lncRNA Sirena1 acts post-transcriptionally and impacts mitochondrial distribution.
Ganesh S, Horvat F, Drutovic D, Efenberkova M, Pinkas D, Jindrova A, Pasulka J, Iyyappan R, Malik R, Susor A, Vlahovicek K, Solc P, Svoboda P
Nucleic acids research 2020 Apr 6;48(6):3211-3227
Nucleic acids research 2020 Apr 6;48(6):3211-3227
Targeted inhibition of metastatic melanoma through interference with Pin1-FOXM1 signaling.
Kruiswijk F, Hasenfuss SC, Sivapatham R, Baar MP, Putavet D, Naipal KA, van den Broek NJ, Kruit W, van der Spek PJ, van Gent DC, Brenkman AB, Campisi J, Burgering BM, Hoeijmakers JH, de Keizer PL
Oncogene 2016 Apr 28;35(17):2166-77
Oncogene 2016 Apr 28;35(17):2166-77
Aurora kinase A is not involved in CPEB1 phosphorylation and cyclin B1 mRNA polyadenylation during meiotic maturation of porcine oocytes.
Komrskova P, Susor A, Malik R, Prochazkova B, Liskova L, Supolikova J, Hladky S, Kubelka M
PloS one 2014;9(7):e101222
PloS one 2014;9(7):e101222
Cellular distributions of molecules with altered expression specific to thyroid proliferative lesions developing in a rat thyroid carcinogenesis model.
Woo GH, Takahashi M, Inoue K, Fujimoto H, Igarashi K, Kanno J, Hirose M, Nishikawa A, Shibutani M
Cancer science 2009 Apr;100(4):617-25
Cancer science 2009 Apr;100(4):617-25
Cyclin B1 is commonly expressed in the cytoplasm of primary human acute myelogenous leukemia cells and serves as a leukemia-associated antigen associated with autoantibody response in a subset of patients.
Ersvaer E, Zhang JY, McCormack E, Olsnes A, Anensen N, Tan EM, Gjertsen BT, Bruserud O
European journal of haematology 2007 Sep;79(3):210-25
European journal of haematology 2007 Sep;79(3):210-25
Cdc6 stability is regulated by the Huwe1 ubiquitin ligase after DNA damage.
Hall JR, Kow E, Nevis KR, Lu CK, Luce KS, Zhong Q, Cook JG
Molecular biology of the cell 2007 Sep;18(9):3340-50
Molecular biology of the cell 2007 Sep;18(9):3340-50
Antigenic profiling of glioma cells to generate allogeneic vaccines or dendritic cell-based therapeutics.
Zhang JG, Eguchi J, Kruse CA, Gomez GG, Fakhrai H, Schroter S, Ma W, Hoa N, Minev B, Delgado C, Wepsic HT, Okada H, Jadus MR
Clinical cancer research : an official journal of the American Association for Cancer Research 2007 Jan 15;13(2 Pt 1):566-575
Clinical cancer research : an official journal of the American Association for Cancer Research 2007 Jan 15;13(2 Pt 1):566-575
The cyclin-dependent kinase inhibitor seliciclib (R-roscovitine; CYC202) decreases the expression of mitotic control genes and prevents entry into mitosis.
Whittaker SR, Te Poele RH, Chan F, Linardopoulos S, Walton MI, Garrett MD, Workman P
Cell cycle (Georgetown, Tex.) 2007 Dec 15;6(24):3114-31
Cell cycle (Georgetown, Tex.) 2007 Dec 15;6(24):3114-31
HPV16 E1--E4 protein is phosphorylated by Cdk2/cyclin A and relocalizes this complex to the cytoplasm.
Davy CE, Ayub M, Jackson DJ, Das P, McIntosh P, Doorbar J
Virology 2006 May 25;349(1):230-44
Virology 2006 May 25;349(1):230-44
Prognostic significance of expressions of cell-cycle regulatory proteins in gastrointestinal stromal tumor and the relevance of the risk grade.
Nakamura N, Yamamoto H, Yao T, Oda Y, Nishiyama K, Imamura M, Yamada T, Nawata H, Tsuneyoshi M
Human pathology 2005 Jul;36(7):828-37
Human pathology 2005 Jul;36(7):828-37
In vitro and in vivo pharmacokinetic-pharmacodynamic relationships for the trisubstituted aminopurine cyclin-dependent kinase inhibitors olomoucine, bohemine and CYC202.
Raynaud FI, Whittaker SR, Fischer PM, McClue S, Walton MI, Barrie SE, Garrett MD, Rogers P, Clarke SJ, Kelland LR, Valenti M, Brunton L, Eccles S, Lane DP, Workman P
Clinical cancer research : an official journal of the American Association for Cancer Research 2005 Jul 1;11(13):4875-87
Clinical cancer research : an official journal of the American Association for Cancer Research 2005 Jul 1;11(13):4875-87
Human papillomavirus type 16 E1 E4-induced G2 arrest is associated with cytoplasmic retention of active Cdk1/cyclin B1 complexes.
Davy CE, Jackson DJ, Raj K, Peh WL, Southern SA, Das P, Sorathia R, Laskey P, Middleton K, Nakahara T, Wang Q, Masterson PJ, Lambert PF, Cuthill S, Millar JB, Doorbar J
Journal of virology 2005 Apr;79(7):3998-4011
Journal of virology 2005 Apr;79(7):3998-4011
The Cyclin-dependent kinase inhibitor CYC202 (R-roscovitine) inhibits retinoblastoma protein phosphorylation, causes loss of Cyclin D1, and activates the mitogen-activated protein kinase pathway.
Whittaker SR, Walton MI, Garrett MD, Workman P
Cancer research 2004 Jan 1;64(1):262-72
Cancer research 2004 Jan 1;64(1):262-72
Complex protein-DNA dynamics at the latent origin of DNA replication of Epstein-Barr virus.
Ritzi M, Tillack K, Gerhardt J, Ott E, Humme S, Kremmer E, Hammerschmidt W, Schepers A
Journal of cell science 2003 Oct 1;116(Pt 19):3971-84
Journal of cell science 2003 Oct 1;116(Pt 19):3971-84
Mitotic cell cycle proteins increase in podocytes despite lack of proliferation.
Petermann AT, Pippin J, Hiromura K, Monkawa T, Durvasula R, Couser WG, Kopp J, Shankland SJ
Kidney international 2003 Jan;63(1):113-22
Kidney international 2003 Jan;63(1):113-22
Citron kinase is a cell cycle-dependent, nuclear protein required for G2/M transition of hepatocytes.
Liu H, Di Cunto F, Imarisio S, Reid LM
The Journal of biological chemistry 2003 Jan 24;278(4):2541-8
The Journal of biological chemistry 2003 Jan 24;278(4):2541-8
Kinetics of endomitosis in primary murine megakaryocytes.
Carow CE, Fox NE, Kaushansky K
Journal of cellular physiology 2001 Sep;188(3):291-303
Journal of cellular physiology 2001 Sep;188(3):291-303
Regulation of cell cycle and cyclins by 16alpha-hydroxyestrone in MCF-7 breast cancer cells.
Lewis JS, Thomas TJ, Klinge CM, Gallo MA, Thomas T
Journal of molecular endocrinology 2001 Dec;27(3):293-307
Journal of molecular endocrinology 2001 Dec;27(3):293-307
Expression of the ALK protein by anaplastic large-cell lymphomas correlates with high proliferative activity.
Leoncini L, Lazzi S, Scano D, Mura A, Onida A, Massarelli G, Tosi P, Barbini P, Cevenini G, Massai MR, Pileri S, Falini B, Giordano A, Kraft R, Laissue JA, Cottier H
International journal of cancer 2000 Jun 15;86(6):777-81
International journal of cancer 2000 Jun 15;86(6):777-81
Expression of the ALK protein by anaplastic large-cell lymphomas correlates with high proliferative activity.
Leoncini L, Lazzi S, Scano D, Mura A, Onida A, Massarelli G, Tosi P, Barbini P, Cevenini G, Massai MR, Pileri S, Falini B, Giordano A, Kraft R, Laissue JA, Cottier H
International journal of cancer 2000 Jun 15;86(6):777-81
International journal of cancer 2000 Jun 15;86(6):777-81
Expression of p34(cdc2) and cyclins A and B compared to other proliferative features of non-Hodgkin's lymphomas: a multivariate cluster analysis.
Leoncini L, Cossu A, Megha T, Bellan C, Lazzi S, Luzi P, Tosi P, Barbini P, Cevenini G, Pileri S, Giordano A, Kraft R, Laissue JA, Cottier H
International journal of cancer 1999 Oct 8;83(2):203-9
International journal of cancer 1999 Oct 8;83(2):203-9
Expression of p34(cdc2) and cyclins A and B compared to other proliferative features of non-Hodgkin's lymphomas: a multivariate cluster analysis.
Leoncini L, Cossu A, Megha T, Bellan C, Lazzi S, Luzi P, Tosi P, Barbini P, Cevenini G, Pileri S, Giordano A, Kraft R, Laissue JA, Cottier H
International journal of cancer 1999 Oct 8;83(2):203-9
International journal of cancer 1999 Oct 8;83(2):203-9
Retinoblastoma-related p107 and pRb2/p130 proteins in malignant lymphomas: distinct mechanisms of cell growth control.
Leoncini L, Bellan C, Cossu A, Claudio PP, Lazzi S, Cinti C, Cevenini G, Megha T, Laurini L, Luzi P, Orcioni GF, Piccioli M, Pileri S, Giardino C, Tosi P, Giordano A
Clinical cancer research : an official journal of the American Association for Cancer Research 1999 Dec;5(12):4065-72
Clinical cancer research : an official journal of the American Association for Cancer Research 1999 Dec;5(12):4065-72
No comments: Submit comment
Supportive validation
- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
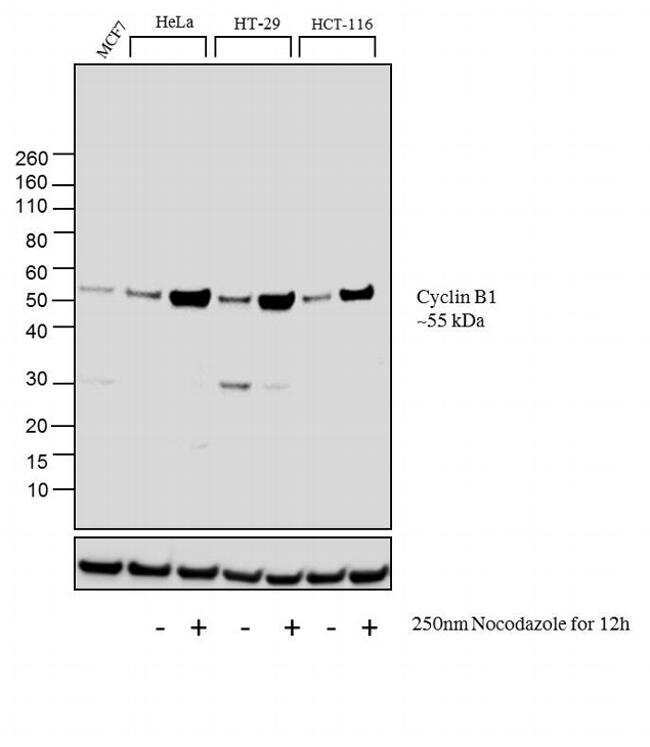
- Western blot analysis was performed on membrane enriched extracts (30 µg lysate) of MCF7 (Lane 1), HeLa (Lane 2), HeLa treated with Nocodazole (250 nm Nocodazole for 12h) (Lane 3), HT-29 (Lane 4), HT-29 treated with Nocodazole (250 nm Nocodazole for 12h) (Lane 5), HCT-116 (Lane6) and HCT-116 treated with Nocodazole (250 nm Nocodazole for 12h) (Lane 7).The blots were probed with Cyclin B1 Mouse monoclonal Antibody (Product # MA5-13128, 2 µg/mL) and detected by chemiluminescence using Goat anti-Mouse IgG (H+L) Superclonal™ Secondary Antibody, HRP conjµgate (Product # A28177, 0.4 µg/mL, 1:2500 dilution). A 55 kDa band corresponding to Cyclin B1 was observed and was enhanced upon treatment in cell lines tested. Known quantity of protein samples were electrophoresed using Novex® NuPAGE® 4-12 % Bis-Tris gel (Product # NP0322BOX), XCell SureLock™ Electrophoresis System (Product # EI0002) and Novex® Sharp Pre-Stained Protein Standard (Product # LC5800). Resolved proteins were then transferred onto a nitrocellulose membrane with iBlot® 2 Dry Blotting System (Product # IB21001). The membrane was probed with the relevant primary and secondary Antibody following blocking with 5% skimmed milk. Chemiluminescent detection was performed using Pierce™ ECL Western Blotting Substrate (Product # 32106).
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
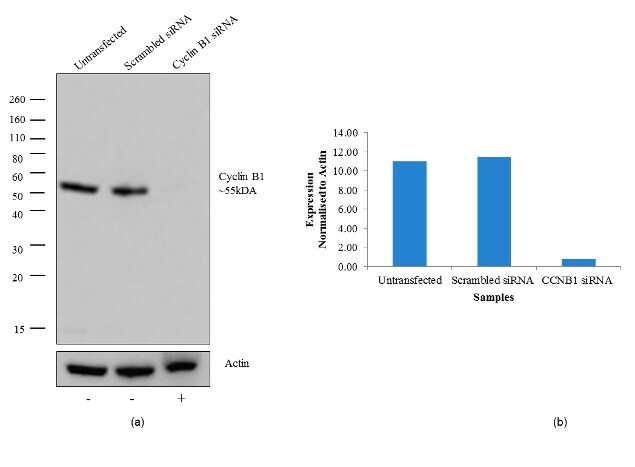
- Knockdown of Cyclin B1 was achieved by transfecting HeLa cells with Cyclin B1 specific validated siRNA (Silencer® select Product # s2516). Western blot analysis (Fig a) was performed using whole cell lysates from the Cyclin B1 knock down cells (lane 3), non-specific scrambled siRNA transfected cells (lane 2) and untransfected cells (lane 1). The blots were probed with Cyclin B1 Mouse monoclonal Antibody (Product # MA5-13128, 2 µg/mL) and Goat anti-Mouse IgG (H+L) Superclonal™ Secondary Antibody, HRP conjugate (Product # A28177, 0.4 µg/mL, 1:2500 dilution). Densitometric analysis of this western blot is shown in histogram(Fig b). Loss of signal upon siRNA mediated knock down confirms that antibody is specific to Cyclin B1.
Supportive validation
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
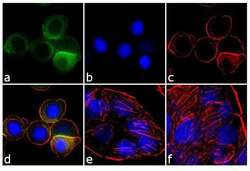
- Experimental details
- Immunofluorescence analysis of Cyclin B1 was performed using 70% confluent log phase HT-29 cells treated 100 ng of Nocodazole for 16 hours. The cells were fixed with 4% paraformaldehyde for 10 minutes, permeabilized with 0.1% Triton™ X-100 for 10 minutes, and blocked with 1% BSA for 1 hour at room temperature. The cells were labeled with Cyclin B1 Mouse (V152) Monoclonal Antibody (Product # MA5-13128) at 2µg/mL in 0.1% BSA and incubated for 3 hours at room temperature and then labeled with Goat anti-Mouse IgG (H+L) Superclonal™ Secondary Antibody, Alexa Fluor® 488 conjµgate (Product # A28175) at a dilution of 1:2000 for 45 minutes at room temperature (Panel a: green). Nuclei (Panel b: blue) were stained with SlowFade® Gold Antifade Mountant with DAPI (Product # S36938). F-actin (Panel c: red) was stained with Alexa Fluor® 555 Rhodamine Phalloidin (Product # R415, 1:300). Panel d represents the merged image showing cytoplasmic localization. Panel e is untreated cell with no signal. Panel f represents control cells with no primary antibody to assess background. The images were captured at 60X magnification.
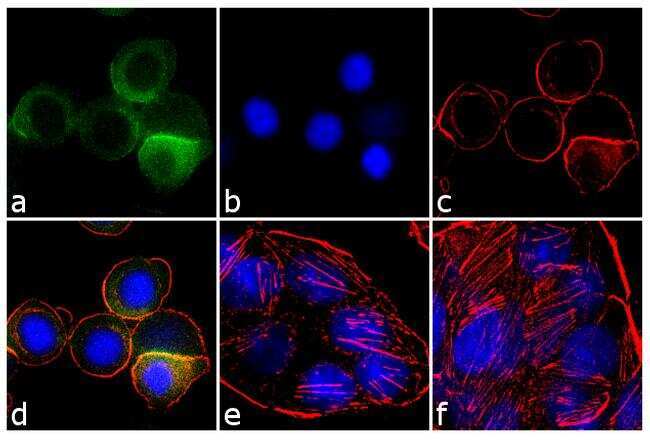
Supportive validation
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
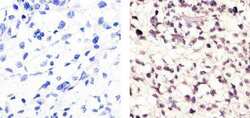
- Experimental details
- Immunohistochemistry analysis of Cyclin B1 (V152) showing staining in the nucleus and weak cytoplasm of paraffin-embedded human lung squamous carcinoma (right) compared to a negative control without primary antibody (left). To expose target proteins, antigen retrieval was performed using 10mM sodium citrate (pH 6.0), microwaved for 8-15 min. Following antigen retrieval, tissues were blocked in 3% H2O2-methanol for 15 min at room temperature, washed with ddH2O and PBS, and then probed with a Cyclin B1 Antibody (V152) Mouse Monoclonal Antibody (Product # MA5-13128) diluted in 3% BSA-PBS at a dilution of 1:100 for 1 hour at 37°C in a humidified chamber. Tissues were washed extensively in PBST and detection was performed using an HRP-conjugated secondary antibody followed by colorimetric detection using a DAB kit. Tissues were counterstained with hematoxylin and dehydrated with ethanol and xylene to prep for mounting.
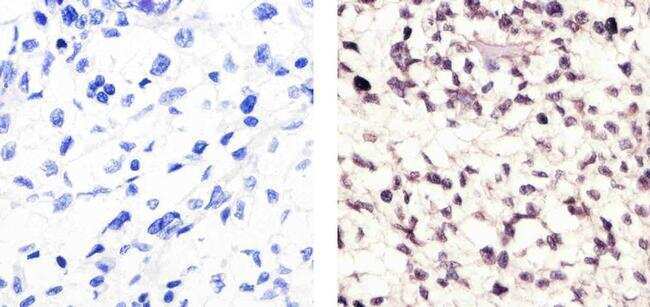
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
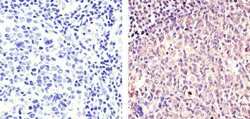
- Experimental details
- Immunohistochemistry analysis of Cyclin B1 (V152) showing staining in the cytoplasm and nucleus of paraffin-embedded human tonsil tissue (right) compared to a negative control without primary antibody (left). To expose target proteins, antigen retrieval was performed using 10mM sodium citrate (pH 6.0), microwaved for 8-15 min. Following antigen retrieval, tissues were blocked in 3% H2O2-methanol for 15 min at room temperature, washed with ddH2O and PBS, and then probed with a Cyclin B1 Antibody (V152) Mouse Monoclonal Antibody (Product # MA5-13128) diluted in 3% BSA-PBS at a dilution of 1:100 for 1 hour at 37°C in a humidified chamber. Tissues were washed extensively in PBST and detection was performed using an HRP-conjugated secondary antibody followed by colorimetric detection using a DAB kit. Tissues were counterstained with hematoxylin and dehydrated with ethanol and xylene to prep for mounting.
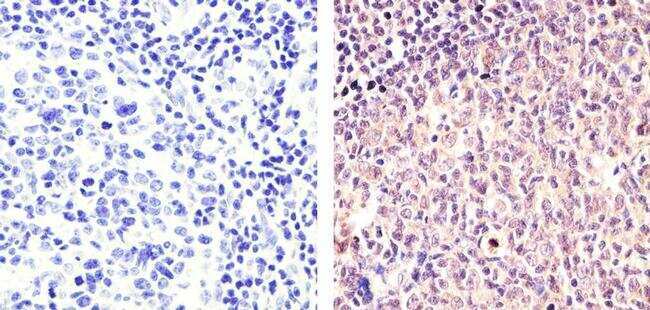
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
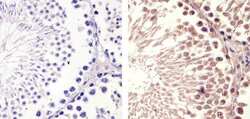
- Experimental details
- Immunohistochemistry analysis of Cyclin B1 (V152) showing staining in the cytoplasm and nucleus of paraffin-embedded mouse testis tissue (right) compared to a negative control without primary antibody (left). To expose target proteins, antigen retrieval was performed using 10mM sodium citrate (pH 6.0), microwaved for 8-15 min. Following antigen retrieval, tissues were blocked in 3% H2O2-methanol for 15 min at room temperature, washed with ddH2O and PBS, and then probed with a Cyclin B1 Antibody (V152) Mouse Monoclonal Antibody (Product # MA5-13128) diluted in 3% BSA-PBS at a dilution of 1:100 for 1 hour at 37°C in a humidified chamber. Tissues were washed extensively in PBST and detection was performed using an HRP-conjugated secondary antibody followed by colorimetric detection using a DAB kit. Tissues were counterstained with hematoxylin and dehydrated with ethanol and xylene to prep for mounting.
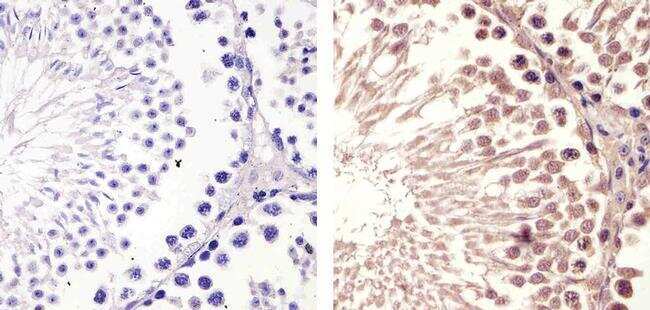
Supportive validation
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
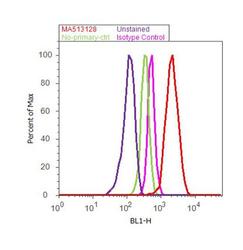
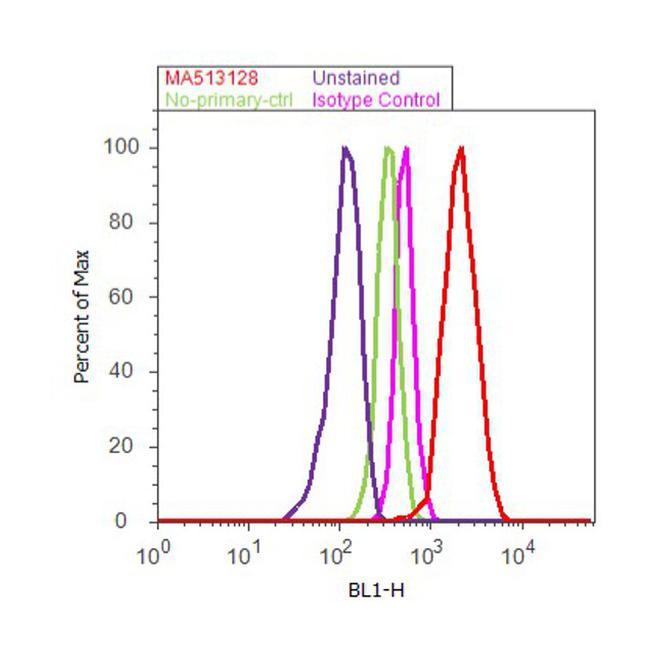
- Experimental details
- Flow cytometry analysis of Cyclin B1 was done on MCF7 cells. Cells were fixed with 70% ethanol for 10 minutes, permeabilized with 0.25% Triton™ X-100 for 20 minutes, and blocked with 5% BSA for 30 minutes at room temperature. Cells were labeled with Cyclin B1 Mouse Monoclonal Antibody (Product # MA5-13128, red histogram) or with mouse isotype control (pink histogram) at 3-5 µg/million cells in 2.5% BSA. After incubation at room temperature for 2 hours, the cells were labeled with Alexa Fluor® 488 Rabbit Anti-Mouse Secondary Antibody (Product # A11059) at a dilution of 1:400 for 30 minutes at room temperature. The representative 10,000 cells were acquired and analyzed for each sample using an Attune® Acoustic Focusing Cytometer. The purple histogram represents unstained control cells and the green histogram represents no-primary-antibody control..
Supportive validation
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
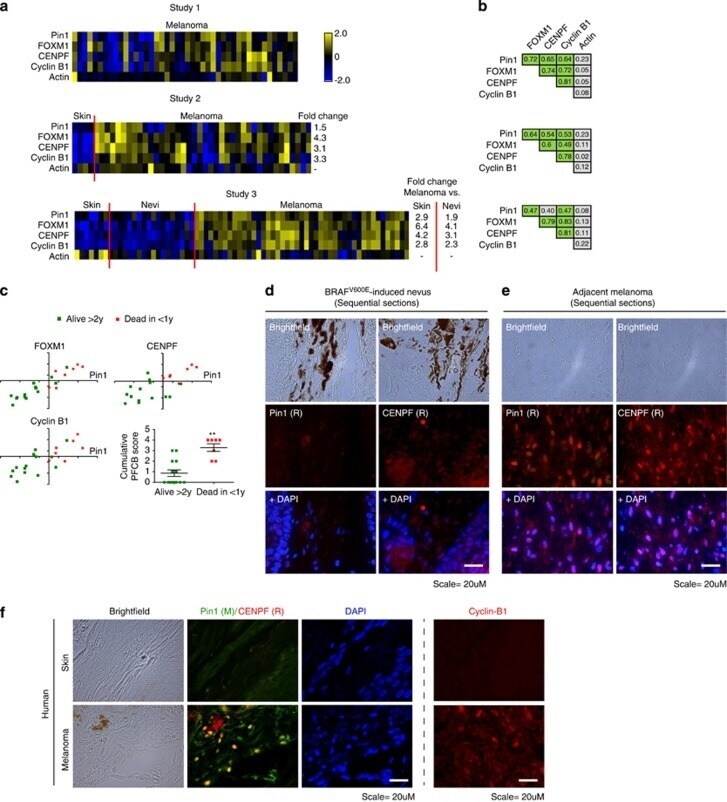
- Figure 2 FOXM1, CENPF, Cyclin B1 and Pin1 mRNA levels correlate in malignant melanoma and associate with poor disease outcome. ( a ) Heatmaps showing relative z -score corrected mRNA expression profiles of Pin1, FOXM1, CENPF, Cyclin B1 and actin in three independent Oncomine data sets. 43 , 44 , 48 Where applicable, the fold difference between normal skin, benign nevi and melanoma are indicated, all of which are significant ( P 0.45; P 2 years or
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
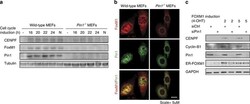
- Figure 3 Pin1 is required for FOXM1 activity. ( a ) Expression of FOXM1 and CENPF is Pin1-dependent. Immunoblot analysis of the indicated proteins from lysates of wild type and Pin1 - /- MEFs synchronized in G0 by serum deprivation or induced to enter cell cycle progression by 10% FCS for the indicated intervals. N =Nocodazole-induced G2/M arrest. ( b ) Pin1 -/- MEFs show reduced FOXM1 expression under basal conditions. Immunocytochemical detection of Pin1 and FOXM1 in proliferating wild type and Pin1 -/- MEFs. The dashed lines indicate the contours of the nuclei. A merged image is shown to compare co-localization. ( c ) The induction of the FOXM1 target genes CENPF and Cylin-B1 by 4-OHT activatable FOXM1-ER is Pin1-dependent. U2OS cells stably expressing FOXM1-ER were transfected with control siRNA (siCtrl) or siRNA against Pin1 (siPin1). After 24 h they were exposed to 2 or 5muM 4-OHT and 24 h later processed for immunoblot analysis of the indicated proteins. Note that the ER-FOXM1 band is not significantly reduced by Pin1 knockdown, indicating stabilization due to the ER fragment.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
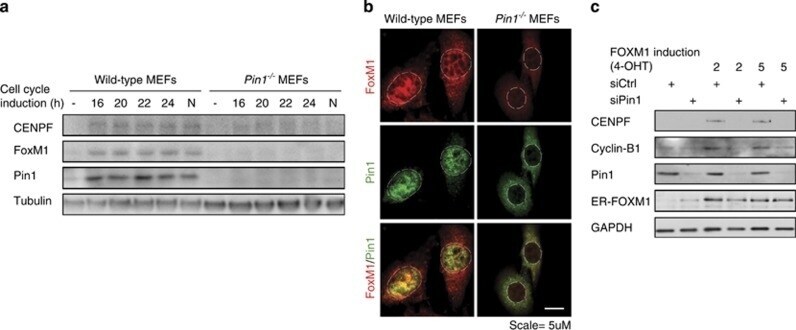
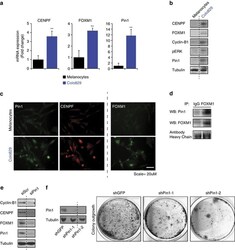
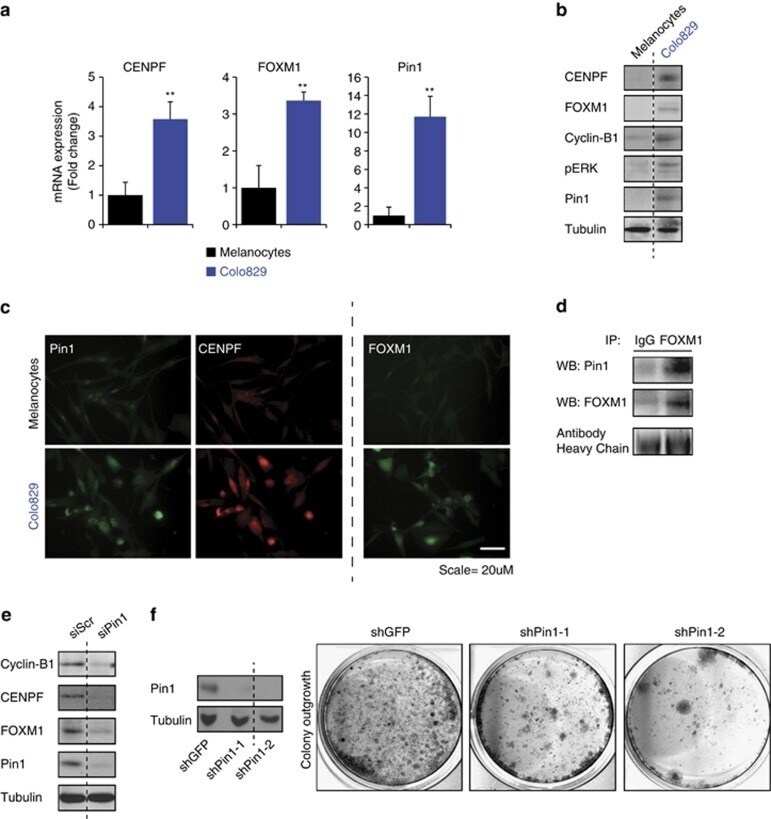
- Figure 5 Pin1 is required for FOXM1 activity and melanoma proliferation in an endogenous BRAF V600E background. ( a - c ) Colo829 melanoma cells express elevated levels of Pin1, FOXM1, CENPF and Cyclin B1. ( a ) QPCR-based detection of the indicated mRNAs in primary neonatal Human Epidermal Melanocytes (HEMn) and Colo829 melanoma cells. ( b ) Immunoblot detection of the indicated proteins corresponding to the mRNAs interrogated in ( a ). ( c ) Detection of the indicated proteins interrogated in b by immunofluorescence. ( d ) Endogenous Pin1 and FOXM1 interact in Colo829 cells. Co-immunoprecipitation (IP) of Colo829 lysates (Input) using an antibody against FOXM1 or control IgG. The antibody heavy chains show equal amounts of FOXM1 and control IgG antibodies were used. ( e ) Pin1 is essential for FOXM1 expression and activity. Colo829 cells were transfected with a control siRNA (siCtrl) or siRNA against Pin1 (siPin1) and processed for immunoblot analysis of the indicated proteins. ( f ) Prolonged Pin1 depletion represses Colo829 outgrowth. Colo829 were stably transduced with two independent lentivirally delivered shRNAs targeting Pin1 (shPin1) or a control shRNA (shGFP) and colony formation was addressed 7 days later. ** P
- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
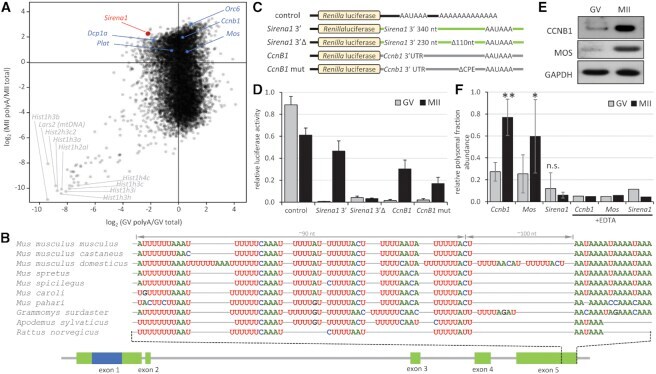
- Figure 5. Cytoplasmic polyadenylation of Sirena1 . ( A ) Sirena1 RNA abundance in RNA-seq data indicates cytoplasmic polyadenylation during meiotic maturation. The plot depicts the ratios of poly(A)-selected RNA-seq data () relative to total RNA-seq data (), positive numbers and negative log 2 values indicate enrichment and depletion of transcripts in poly(A)-selected RNA-seq data, respectively. In blue are labeled dormant mRNAs with a short poly(A) tail in GV oocytes, which undergo polyadenylation during meiotic maturation. Gray labels depict histone and mitochondrial genes underrepresented in poly(A)-selected RNA-seq data. ( B ) Sequence organization and conservation of the 3'end of Sirena1 . Depicted are putative CPE elements ~100 nucleotides from the poly(A) signal in ten rodent genomes. ( C ) Schematic design of reporters for analysis of cytoplasmic polyadenylation. Renilla luciferase reporter (control) was polyadenylated, other reporters were not polyadenylated. ( D ) Relative luciferase activity of microinjected reporters. Reporters were microinjected into GV oocytes, which were either matured for 20 h (MII) or cultured in the presence of milrinone, which prevents resumption of meiosis (GV). Activity was calculated as luciferase activity of each Renilla reporter divided by a co-injected polyadenylated firefly reporter standard. Error bars = SD. ( E ) Western blot analysis of MOS and CCNB1 proteins in fully-grown GV oocytes and MII eggs. ( F ) Relative abundance of Mos
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
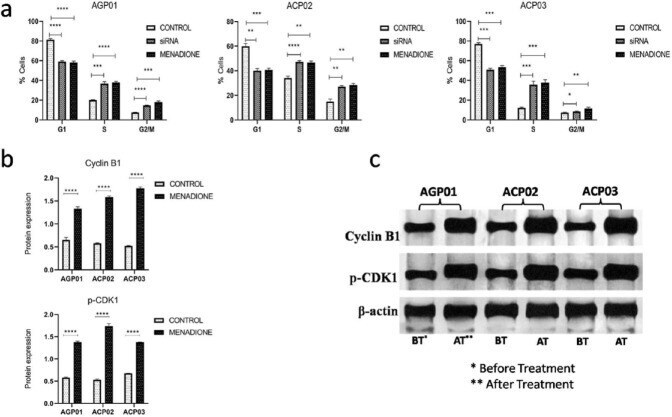
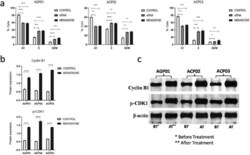
- Figure 2. Menadione induced cell cycle arrest at the G2/M phase. (a) Cell cycle analysis by flow cytometry showing a significant increase in the percentage of cells at G2/M phase after the treatment with menadione in all three GC cell lines. (b) Menadione high significantly increased protein expression of Cyclin B1 and p-CDK1, confirming its potential to arrest cell cycle progression at the G2/M phase. (c) Protein expression of Cyclin B1 and p-CDK1 demonstrated by western blot. The control groups are the respective nontreated cell line.