Explore
Explore Validate
Validate Learn
Learn Western blot
Western blot ELISA
ELISA Immunocytochemistry
ImmunocytochemistryAntibody data
- Antibody Data
- Antigen structure
- References [190]
- Comments [0]
- Validations
- Immunocytochemistry [2]
- Immunohistochemistry [2]
- Other assay [93]
Submit
Validation data
Reference
Comment
Report error
- Product number
- 34-1700 - Provider product page

- Provider
- Invitrogen Antibodies
- Product name
- Claudin 3 Polyclonal Antibody
- Antibody type
- Polyclonal
- Antigen
- Synthetic peptide
- Description
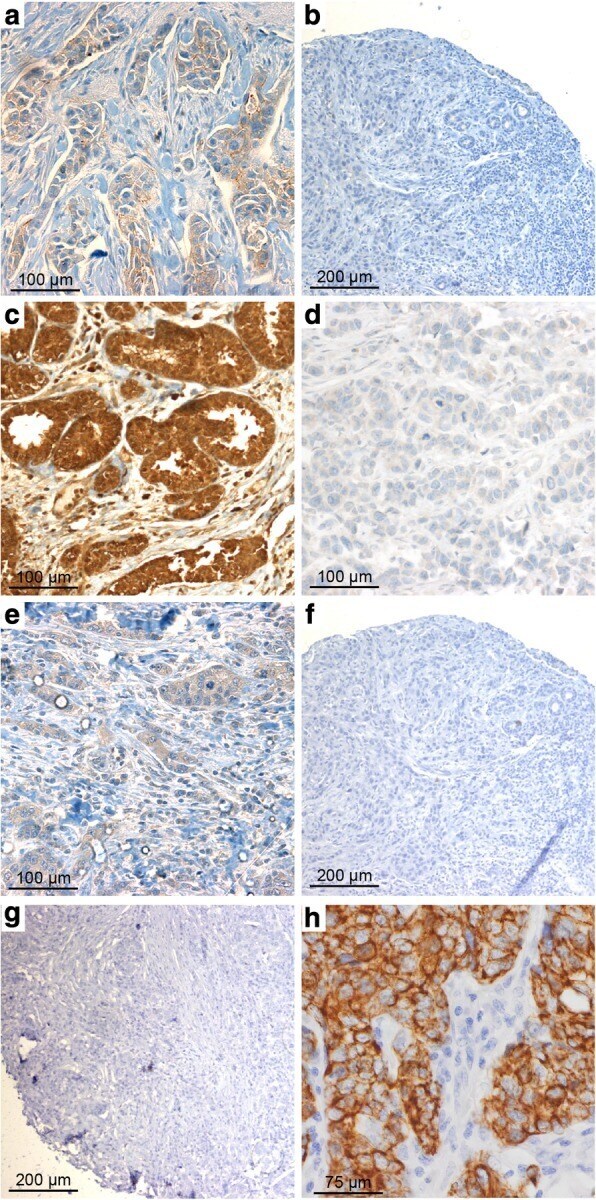
- This antibody is specific for the 22 kDa Claudin-3 protein. Reactivity has been confirmed with mouse liver and lung homogenates, canine MDCK and human MCF-7 cell lysates by western blotting, and with formalin-fixed, paraffin-embedded (FFPE) human normal colon and colon cancer tissues by immunohistochemistry. For best results in immunohistochemistry with formalin-fixed, paraffin-embedded (FFPE) tissues, heat induced epitope retrieval (HIER) with citrate buffer, pH 6.0, is required prior to staining.
- Reactivity
- Human, Mouse, Canine
- Host
- Rabbit
- Isotype
- IgG
- Vial size
- 100 μg
- Concentration
- 0.25 mg/mL
- Storage
- -20°C
Submitted references Cellular Distribution Pattern of tjp1 (ZO-1) in Xenopus laevis Oocytes Heterologously Expressing Claudins.
IL-22 regulates endometrial regeneration by enhancing tight junctions and orchestrating extracellular matrix.
Investigating mammary glands of lactating goats for the presence of tertiary lymphoid organs.
cyc-DEP: Cyclic immunofluorescence profiling of particles collected using dielectrophoresis.
Differential expression and localization of tight junction proteins in the goat epididymis.
MarvelD3 Is Upregulated in Ulcerative Colitis and Has Attenuating Effects during Colitis Indirectly Stabilizing the Intestinal Barrier.
Glycine represses endoplasmic reticulum stress-related apoptosis and improves intestinal barrier by activating mammalian target of rapamycin complex 1 signaling.
SARS-CoV-2-Induced Pathology-Relevance to COVID-19 Pathophysiology.
Organ-specific extracellular matrix directs trans-differentiation of mesenchymal stem cells and formation of salivary gland-like organoids in vivo.
High-Fat Diet Promotes Colorectal Tumorigenesis Through Modulating Gut Microbiota and Metabolites.
Nanoscale segregation of channel and barrier claudins enables paracellular ion flux.
Centella asiatica-Derived Endothelial Paracrine Restores Epithelial Barrier Dysfunction in Radiation-Induced Enteritis.
Intestinal organoid-based 2D monolayers mimic physiological and pathophysiological properties of the pig intestine.
Development of Anti-inflammatory Probiotic Limosilactobacillus reuteri EFEL6901 as Kimchi Starter: in vitro and In vivo Evidence.
Effects of 1,2-Dimethylhydrazine on Barrier Properties of Rat Large Intestine and IPEC-J2 Cells.
Treatment with Edoxaban Attenuates Acute Stroke Severity in Mice by Reducing Blood-Brain Barrier Damage and Inflammation.
Matrix Metalloproteinase MMP-12 Promotes Macrophage Transmigration Across Intestinal Epithelial Tight Junctions and Increases Severity of Experimental Colitis.
Discovery of anti-inflammatory physiological peptides that promote tissue repair by reinforcing epithelial barrier formation.
Ablation of Red Stable Transfected Claudin Expressing Canine Prostate Adenocarcinoma and Transitional Cell Carcinoma Cell Lines by C-CPE Gold-Nanoparticle-Mediated Laser Intervention.
Butyrate Alleviates Cytokine-Induced Barrier Dysfunction by Modifying Claudin-2 Levels.
Targeting PVT1 Exon 9 Re-Expresses Claudin 4 Protein and Inhibits Migration by Claudin-Low Triple Negative Breast Cancer Cells.
A human surfactant B deficiency air-liquid interface cell culture model suitable for gene therapy applications.
Luminal polyethylene glycol solution delays the onset of preservation injury in the human intestine.
Microbiota regulate innate immune signaling and protective immunity against cancer.
Hypoxia and heat stress affect epithelial integrity in a Caco-2/HT-29 co-culture.
Isolation and development of bovine primary respiratory cells as model to study influenza D virus infection.
Tight junctions in the blood-brain barrier promote edema formation and infarct size in stroke - Ambivalent effects of sealing proteins.
Quercetin Alleviates Oxidative Damage by Activating Nuclear Factor Erythroid 2-Related Factor 2 Signaling in Porcine Enterocytes.
Citrus limon Peel Powder Reduces Intestinal Barrier Defects and Inflammation in a Colitic Murine Experimental Model.
JAK-STAT Pathway Inhibition Partially Restores Intestinal Homeostasis in Hdac1- and Hdac2-Intestinal Epithelial Cell-Deficient Mice.
STIM-Orai1 signaling regulates fluidity of cytoplasm during membrane blebbing.
Culture surface protein coatings affect the barrier properties and calcium signalling of hESC-RPE.
Tumor Necrosis Factor Alpha Effects on the Porcine Intestinal Epithelial Barrier Include Enhanced Expression of TNF Receptor 1.
Vitamin D receptor is overexpressed in the duodenum of patients with irritable bowel syndrome.
Occludin and tricellulin facilitate formation of anastomosing tight-junction strand network to improve barrier function.
Blood-Brain Barrier Protein Claudin-5 Expressed in Paired Xenopus laevis Oocytes Mediates Cell-Cell Interaction.
N‑glycosylation and receptor tyrosine kinase signaling affect claudin‑3 levels in colorectal cancer cells.
Metformin and Inhibition of Transforming Growth Factor-Beta Stimulate In Vitro Transport in Primary Renal Tubule Cells.
Combinatorial expression of claudins in the proximal renal tubule and its functional consequences.
Nicotine directly affects milk production in lactating mammary epithelial cells concurrently with inactivation of STAT5 and glucocorticoid receptor in vitro.
NOTCH1 signaling establishes the medullary thymic epithelial cell progenitor pool during mouse fetal development.
Circulating Ouabain Modulates Expression of Claudins in Rat Intestine and Cerebral Blood Vessels.
Maltodextrin-induced intestinal injury in a neonatal mouse model.
Adverse Effects of Coumestrol and Genistein on Mammary Morphogenesis and Future Milk Production Ability of Mammary Epithelial Cells.
Effects of Claudin-1 on the Action of Clostridium perfringens Enterotoxin in Caco-2 Cells.
Claudins and JAM-A coordinately regulate tight junction formation and epithelial polarity.
In vitro and in vivo effects of a mycotoxin, deoxynivalenol, and a trace metal, cadmium, alone or in a mixture on the intestinal barrier.
Fusarium Mycotoxins Disrupt the Barrier and Induce IL-6 Release in a Human Placental Epithelium Cell Line.
Newly synthesized claudins but not occludin are added to the basal side of the tight junction.
Effect of Manitoba-Grown Red-Osier Dogwood Extracts on Recovering Caco-2 Cells from H(2)O(2)-Induced Oxidative Damage.
Association of Cytokeratin 5 and Claudin 3 expression with BRCA1 and BRCA2 germline mutations in women with early breast cancer.
The kidney anion exchanger 1 affects tight junction properties via claudin-4.
Morphologic determinant of tight junctions revealed by claudin-3 structures.
Electrical Stimulation of the Mesencephalic Locomotor Region Has No Impact on Blood-Brain Barrier Alterations after Cerebral Photothrombosis in Rats.
p120ctn-Mediated Organ Patterning Precedes and Determines Pancreatic Progenitor Fate.
Vitamin D Receptor Deletion Leads to the Destruction of Tight and Adherens Junctions in Lungs.
Differential Claudin 3 and EGFR Expression Predicts BRCA1 Mutation in Triple-Negative Breast Cancer.
Loss of connexin43 in murine Sertoli cells and its effect on blood-testis barrier formation and dynamics.
Glial Cells in the Fish Retinal Nerve Fiber Layer Form Tight Junctions, Separating and Surrounding Axons.
Functionalization of gold-nanoparticles by the Clostridium perfringens enterotoxin C-terminus for tumor cell ablation using the gold nanoparticle-mediated laser perforation technique.
Salivary glands regenerate after radiation injury through SOX2-mediated secretory cell replacement.
Claudin-3 Loss Causes Leakage of Sweat from the Sweat Gland to Contribute to the Pathogenesis of Atopic Dermatitis.
Immunohistochemistry in Investigative and Toxicologic Pathology.
Adherens junctions influence tight junction formation via changes in membrane lipid composition.
Claudin Loss-of-Function Disrupts Tight Junctions and Impairs Amelogenesis.
Altered Cytokine Expression and Barrier Properties after In Vitro Infection of Porcine Epithelial Cells with Enterotoxigenic Escherichia coli and Probiotic Enterococcus faecium.
The role of human fibronectin- or placenta basement membrane extract-based gels in favouring the formation of polarized salivary acinar-like structures.
Talin regulates integrin β1-dependent and -independent cell functions in ureteric bud development.
Establishment, maintenance and functional integrity of the blood-testis barrier in organotypic cultures of fresh and frozen/thawed prepubertal mouse testes.
Obesity-induces Organ and Tissue Specific Tight Junction Restructuring and Barrier Deregulation by Claudin Switching.
Downregulation of lipolysis-stimulated lipoprotein receptor promotes cell invasion via claudin-1-mediated matrix metalloproteinases in human endometrial cancer.
Analysis of Epithelial-Mesenchymal Transition Induced by Transforming Growth Factor β.
Highly Conserved Testicular Localization of Claudin-11 in Normal and Impaired Spermatogenesis.
Effects of Osmolality on Paracellular Transport in MDCK II Cells.
Liver kinase B1 regulates hepatocellular tight junction distribution and function in vivo.
Alix-mediated assembly of the actomyosin-tight junction polarity complex preserves epithelial polarity and epithelial barrier.
Modeling immune functions of the mouse blood-cerebrospinal fluid barrier in vitro: primary rather than immortalized mouse choroid plexus epithelial cells are suited to study immune cell migration across this brain barrier.
miR-200 promotes the mesenchymal to epithelial transition by suppressing multiple members of the Zeb2 and Snail1 transcriptional repressor complexes.
Prolactin and glucocorticoid signaling induces lactation-specific tight junctions concurrent with β-casein expression in mammary epithelial cells.
Connexins, E-cadherin, Claudin-7 and β-catenin transiently form junctional nexuses during the post-natal mammary gland development.
Claudin-2 knockout by TALEN-mediated gene targeting in MDCK cells: claudin-2 independently determines the leaky property of tight junctions in MDCK cells.
Effects of Hydrostatic Pressure on Carcinogenic Properties of Epithelia.
Heterogeneity between triple negative breast cancer cells due to differential activation of Wnt and PI3K/AKT pathways.
Mode of action of claudin peptidomimetics in the transient opening of cellular tight junction barriers.
A non-tight junction function of claudin-7-Interaction with integrin signaling in suppressing lung cancer cell proliferation and detachment.
Claudin-4 is required for modulation of paracellular permeability by muscarinic acetylcholine receptor in epithelial cells.
Galacto-oligosaccharides Protect the Intestinal Barrier by Maintaining the Tight Junction Network and Modulating the Inflammatory Responses after a Challenge with the Mycotoxin Deoxynivalenol in Human Caco-2 Cell Monolayers and B6C3F1 Mice.
Overexpression and delocalization of claudin-3 protein in MCF-7 and MDA-MB-415 breast cancer cell lines.
Homoharringtonine increases intestinal epithelial permeability by modulating specific claudin isoforms in Caco-2 cell monolayers.
Expression of tight-junction proteins in human proximal small intestinal mucosa before and after Roux-en-Y gastric bypass surgery.
Intestinal fatty acid binding protein as a marker for intra-abdominal pressure-related complications in patients admitted to the intensive care unit; study protocol for a prospective cohort study (I-Fabulous study).
ZO-1 knockout by TALEN-mediated gene targeting in MDCK cells: involvement of ZO-1 in the regulation of cytoskeleton and cell shape.
Impaired intestinal mucosal barrier upon ischemia-reperfusion: "patching holes in the shield with a simple surgical method".
Autonomous isolation, long-term culture and differentiation potential of adult salivary gland-derived stem/progenitor cells.
Biological significance of FoxN1 gain-of-function mutations during T and B lymphopoiesis in juvenile mice.
Diets high in fermentable protein and fibre alter tight junction protein composition with minor effects on barrier function in piglet colon.
Histopathologic and molecular evaluation of the Organ Procurement and Transplantation Network selection criteria for intestinal graft donation.
Disrupted tight junctions in the small intestine of cystic fibrosis mice.
Activation of Gpr109a, receptor for niacin and the commensal metabolite butyrate, suppresses colonic inflammation and carcinogenesis.
Immunohistological characterization of intercellular junction proteins in rhesus macaque intestine.
Ectopic TBX1 suppresses thymic epithelial cell differentiation and proliferation during thymus organogenesis.
Claudin-3 overexpression increases the malignant potential of colorectal cancer cells: roles of ERK1/2 and PI3K-Akt as modulators of EGFR signaling.
Transmigration of neural stem cells across the blood brain barrier induced by glioma cells.
The tight-junction protein claudin-6 induces epithelial differentiation from mouse F9 and embryonic stem cells.
Guanylate cyclase C limits systemic dissemination of a murine enteric pathogen.
Build them up and break them down: Tight junctions of cell lines expressing typical hepatocyte polarity with a varied repertoire of claudins.
Role of the p63-FoxN1 regulatory axis in thymic epithelial cell homeostasis during aging.
SOX8 regulates permeability of the blood-testes barrier that affects adult male fertility in the mouse.
A complex containing LPP and α-actinin mediates TGFβ-induced migration and invasion of ErbB2-expressing breast cancer cells.
Rab25 regulates integrin expression in polarized colonic epithelial cells.
Derivation of myoepithelial progenitor cells from bipotent mammary stem/progenitor cells.
EpCAM contributes to formation of functional tight junction in the intestinal epithelium by recruiting claudin proteins.
Differential effects of flavonoids on barrier integrity in human intestinal Caco-2 cells.
Dynamic distribution of claudin proteins in pancreatic epithelia undergoing morphogenesis or neoplastic transformation.
Age-Related Disruption of Steady-State Thymic Medulla Provokes Autoimmune Phenotype via Perturbing Negative Selection.
Effects of proinflammatory cytokines on the claudin-19 rich tight junctions of human retinal pigment epithelium.
Two strikingly different signaling pathways are induced by meningococcal type IV pili on endothelial and epithelial cells.
Functional characterization and localization of a gill-specific claudin isoform in Atlantic salmon.
Zonula occludens-1 and -2 regulate apical cell structure and the zonula adherens cytoskeleton in polarized epithelia.
Probiotic bacteria induce maturation of intestinal claudin 3 expression and barrier function.
Complexity and developmental changes in the expression pattern of claudins at the blood-CSF barrier.
Distinct domains of paracingulin are involved in its targeting to the actin cytoskeleton and regulation of apical junction assembly.
Enhanced immunohistochemical resolution of claudin proteins in glycolmethacrylate-embedded tissue biopsies.
Claudin-1 induced sealing of blood-brain barrier tight junctions ameliorates chronic experimental autoimmune encephalomyelitis.
Matrigel improves functional properties of primary human salivary gland cells.
Morphogenesis and maintenance of the 3D thymic medulla and prevention of nude skin phenotype require FoxN1 in pre- and post-natal K14 epithelium.
Identification of a claudin-4 and E-cadherin score to predict prognosis in breast cancer.
The Hedgehog pathway promotes blood-brain barrier integrity and CNS immune quiescence.
Hypoxia increases transepithelial electrical conductance and reduces occludin at the plasma membrane in alveolar epithelial cells via PKC-ζ and PP2A pathway.
Ovarian hormones control the changing expression of claudins and occludin in rat uterine epithelial cells during early pregnancy.
Effects of long-term progesterone on developmental and functional aspects of porcine uterine epithelia and vasculature: progesterone alone does not support development of uterine glands comparable to that of pregnancy.
Astroglial structures in the zebrafish brain.
beta-catenin expression and claudin expression pattern as prognostic factors of prostatic cancer progression.
Minimal effects of VEGF and anti-VEGF drugs on the permeability or selectivity of RPE tight junctions.
Non-invasive markers for early diagnosis and determination of the severity of necrotizing enterocolitis.
Differential expression of claudin tight junction proteins in the human cortical nephron.
Claudin-1 has tumor suppressive activity and is a direct target of RUNX3 in gastric epithelial cells.
Postnatal tissue-specific disruption of transcription factor FoxN1 triggers acute thymic atrophy.
Allogeneic human mesenchymal stem cells restore epithelial protein permeability in cultured human alveolar type II cells by secretion of angiopoietin-1.
Differential distribution of tight junction proteins suggests a role for tanycytes in blood-hypothalamus barrier regulation in the adult mouse brain.
Tight junction proteins in human Schwann cell autotypic junctions.
Claudin-1, -2, -3, -4, -7, -8, and -10 protein expression in biliary tract cancers.
Intestinal cytoskeleton degradation precedes tight junction loss following hemorrhagic shock.
A SNAIL1-SMAD3/4 transcriptional repressor complex promotes TGF-beta mediated epithelial-mesenchymal transition.
New Insight in Loss of Gut Barrier during Major Non-Abdominal Surgery.
Blood-testis barrier dynamics are regulated by testosterone and cytokines via their differential effects on the kinetics of protein endocytosis and recycling in Sertoli cells.
Regulation of testicular tight junctions by gonadotrophins in the adult Djungarian hamster in vivo.
Inducible overexpression of cingulin in stably transfected MDCK cells does not affect tight junction organization and gene expression.
Distribution of tight junction proteins in adult human salivary glands.
Expression of Claudin-1, Claudin-3 and Claudin-5 in human blood-brain barrier mimicking cell line ECV304 is inducible by glioma-conditioned media.
Distinct molecular composition of blood and lymphatic vascular endothelial cell junctions establishes specific functional barriers within the peripheral lymph node.
Structural alterations of epididymal epithelial cells in cathepsin A-deficient mice affect the blood-epididymal barrier and lead to altered sperm motility.
Cadmium causes delayed effects on renal function in the offspring of cadmium-contaminated pregnant female rats.
Coxsackievirus and adenovirus receptor is up-regulated in migratory germ cells during passage of the blood-testis barrier.
Medullary thymic epithelial cells expressing Aire represent a unique lineage derived from cells expressing claudin.
Differences in claudin synthesis in primary cultures of acinar cells from rat salivary gland are correlated with the specific three-dimensional organization of the cells.
Tight and adherens junctions in the ovine uterus: differential regulation by pregnancy and progesterone.
Loss of claudins-1 and -7 and expression of claudins-3 and -4 correlate with prognostic variables in prostatic adenocarcinomas.
Claudin 1 differentiates endometrioid and serous papillary endometrial adenocarcinoma.
Endothelia of term human placentae display diminished expression of tight junction proteins during preeclampsia.
Tight junction proteins and perineurial cells in neurofibromas.
Zinc protects renal function during cadmium intoxication in the rat.
Cingulin regulates claudin-2 expression and cell proliferation through the small GTPase RhoA.
Changes of cell adhesion and extracellular matrix (ECM) components in cervical intraepithelial neoplasia.
Inflammatory processes have differential effects on claudins 2, 3 and 4 in colonic epithelial cells.
Inducible expression of Snail selectively increases paracellular ion permeability and differentially modulates tight junction proteins.
Polyamines are necessary for synthesis and stability of occludin protein in intestinal epithelial cells.
Knockdown of occludin expression leads to diverse phenotypic alterations in epithelial cells.
Leukocyte diapedesis in vivo induces transient loss of tight junction protein at the blood-retina barrier.
Phosphorylation of claudin-3 at threonine 192 by cAMP-dependent protein kinase regulates tight junction barrier function in ovarian cancer cells.
Extracellular signal-regulated kinases 1/2 control claudin-2 expression in Madin-Darby canine kidney strain I and II cells.
Chronic inflammatory pain leads to increased blood-brain barrier permeability and tight junction protein alterations.
Claudin-3 and claudin-4 expression in ovarian epithelial cells enhances invasion and is associated with increased matrix metalloproteinase-2 activity.
Cultured monolayers of the dog jejunum with the structural and functional properties resembling the normal epithelium.
Paracellular Cl- permeability is regulated by WNK4 kinase: insight into normal physiology and hypertension.
The rotavirus surface protein VP8 modulates the gate and fence function of tight junctions in epithelial cells.
Epidermal growth factor receptor activation differentially regulates claudin expression and enhances transepithelial resistance in Madin-Darby canine kidney cells.
Mechanisms of diarrhea in the interleukin-2-deficient mouse model of colonic inflammation.
Expression and function of tight junctions in the crypt epithelium of human palatine tonsils.
Tight junction proteins ZO-1, occludin, and claudins in developing and adult human perineurium.
Role of claudin interactions in airway tight junctional permeability.
Claudin-8 expression in Madin-Darby canine kidney cells augments the paracellular barrier to cation permeation.
Localization of claudin-3 in tight junctions of the blood-brain barrier is selectively lost during experimental autoimmune encephalomyelitis and human glioblastoma multiforme.
Tight junction proteins claudin-3 and claudin-4 are frequently overexpressed in ovarian cancer but not in ovarian cystadenomas.
Effects of hypoxia-reoxygenation on rat blood-brain barrier permeability and tight junctional protein expression.
Bryostatin-1 enhances barrier function in T84 epithelia through PKC-dependent regulation of tight junction proteins.
Claudin-2 expression induces cation-selective channels in tight junctions of epithelial cells.
Claudin-2 expression induces cation-selective channels in tight junctions of epithelial cells.
Mechanisms of diarrhea in collagenous colitis.
Mechanisms of diarrhea in collagenous colitis.
Brunner N, Stein L, Amasheh S
The Journal of membrane biology 2023 Feb;256(1):51-61
The Journal of membrane biology 2023 Feb;256(1):51-61
IL-22 regulates endometrial regeneration by enhancing tight junctions and orchestrating extracellular matrix.
Ganieva U, Schneiderman S, Bu P, Beaman K, Dambaeva S
Frontiers in immunology 2022;13:955576
Frontiers in immunology 2022;13:955576
Investigating mammary glands of lactating goats for the presence of tertiary lymphoid organs.
Tsugami Y, Nakayama S, Suzuki N, Nii T, Isobe N
Frontiers in immunology 2022;13:941333
Frontiers in immunology 2022;13:941333
cyc-DEP: Cyclic immunofluorescence profiling of particles collected using dielectrophoresis.
Gustafson KT, Sayar Z, Le H, Gustafson SL, Gower A, Modestino A, Ibsen S, Heller MJ, Esener S, Eksi SE
Electrophoresis 2022 Sep;43(16-17):1784-1798
Electrophoresis 2022 Sep;43(16-17):1784-1798
Differential expression and localization of tight junction proteins in the goat epididymis.
Kim SW, Jeong YD, Lee GY, Lee J, Lee JY, Kim CL, Ko YG, Lee SS, Kim B
Journal of animal science and technology 2022 May;64(3):500-514
Journal of animal science and technology 2022 May;64(3):500-514
MarvelD3 Is Upregulated in Ulcerative Colitis and Has Attenuating Effects during Colitis Indirectly Stabilizing the Intestinal Barrier.
Weiß F, Czichos C, Knobe L, Voges L, Bojarski C, Michel G, Fromm M, Krug SM
Cells 2022 May 4;11(9)
Cells 2022 May 4;11(9)
Glycine represses endoplasmic reticulum stress-related apoptosis and improves intestinal barrier by activating mammalian target of rapamycin complex 1 signaling.
Yang Y, Fan X, Ji Y, Li J, Dai Z, Wu Z
Animal nutrition (Zhongguo xu mu shou yi xue hui) 2022 Mar;8(1):1-9
Animal nutrition (Zhongguo xu mu shou yi xue hui) 2022 Mar;8(1):1-9
SARS-CoV-2-Induced Pathology-Relevance to COVID-19 Pathophysiology.
Zinserling VA, Semenova NY, Bikmurzina AE, Kruglova NM, Rybalchenko OV, Markov AG
Pathophysiology : the official journal of the International Society for Pathophysiology 2022 Jun 10;29(2):281-297
Pathophysiology : the official journal of the International Society for Pathophysiology 2022 Jun 10;29(2):281-297
Organ-specific extracellular matrix directs trans-differentiation of mesenchymal stem cells and formation of salivary gland-like organoids in vivo.
Tran ON, Wang H, Li S, Malakhov A, Sun Y, Abdul Azees PA, Gonzalez AO, Cao B, Marinkovic M, Singh BB, Dean DD, Yeh CK, Chen XD
Stem cell research & therapy 2022 Jul 15;13(1):306
Stem cell research & therapy 2022 Jul 15;13(1):306
High-Fat Diet Promotes Colorectal Tumorigenesis Through Modulating Gut Microbiota and Metabolites.
Yang J, Wei H, Zhou Y, Szeto CH, Li C, Lin Y, Coker OO, Lau HCH, Chan AWH, Sung JJY, Yu J
Gastroenterology 2022 Jan;162(1):135-149.e2
Gastroenterology 2022 Jan;162(1):135-149.e2
Nanoscale segregation of channel and barrier claudins enables paracellular ion flux.
Gonschior H, Schmied C, Van der Veen RE, Eichhorst J, Himmerkus N, Piontek J, Günzel D, Bleich M, Furuse M, Haucke V, Lehmann M
Nature communications 2022 Aug 25;13(1):4985
Nature communications 2022 Aug 25;13(1):4985
Centella asiatica-Derived Endothelial Paracrine Restores Epithelial Barrier Dysfunction in Radiation-Induced Enteritis.
Kwak SY, Jang WI, Lee SB, Kim MJ, Park S, Cho SS, Kim H, Lee SJ, Shim S, Jang H
Cells 2022 Aug 16;11(16)
Cells 2022 Aug 16;11(16)
Intestinal organoid-based 2D monolayers mimic physiological and pathophysiological properties of the pig intestine.
Hoffmann P, Schnepel N, Langeheine M, Künnemann K, Grassl GA, Brehm R, Seeger B, Mazzuoli-Weber G, Breves G
PloS one 2021;16(8):e0256143
PloS one 2021;16(8):e0256143
Development of Anti-inflammatory Probiotic Limosilactobacillus reuteri EFEL6901 as Kimchi Starter: in vitro and In vivo Evidence.
Seo H, Seong H, Kim GY, Jo YM, Cheon SW, Song Y, Ryu BH, Kang H, Han NS
Frontiers in microbiology 2021;12:760476
Frontiers in microbiology 2021;12:760476
Effects of 1,2-Dimethylhydrazine on Barrier Properties of Rat Large Intestine and IPEC-J2 Cells.
Bekusova V, Droessler L, Amasheh S, Markov AG
International journal of molecular sciences 2021 Sep 24;22(19)
International journal of molecular sciences 2021 Sep 24;22(19)
Treatment with Edoxaban Attenuates Acute Stroke Severity in Mice by Reducing Blood-Brain Barrier Damage and Inflammation.
Bieber M, Foerster KI, Haefeli WE, Pham M, Schuhmann MK, Kraft P
International journal of molecular sciences 2021 Sep 13;22(18)
International journal of molecular sciences 2021 Sep 13;22(18)
Matrix Metalloproteinase MMP-12 Promotes Macrophage Transmigration Across Intestinal Epithelial Tight Junctions and Increases Severity of Experimental Colitis.
Nighot M, Ganapathy AS, Saha K, Suchanec E, Castillo EF, Gregory A, Shapiro S, Ma T, Nighot P
Journal of Crohn's & colitis 2021 Oct 7;15(10):1751-1765
Journal of Crohn's & colitis 2021 Oct 7;15(10):1751-1765
Discovery of anti-inflammatory physiological peptides that promote tissue repair by reinforcing epithelial barrier formation.
Oda Y, Takahashi C, Harada S, Nakamura S, Sun D, Kiso K, Urata Y, Miyachi H, Fujiyoshi Y, Honigmann A, Uchida S, Ishihama Y, Toyoshima F
Science advances 2021 Nov 19;7(47):eabj6895
Science advances 2021 Nov 19;7(47):eabj6895
Ablation of Red Stable Transfected Claudin Expressing Canine Prostate Adenocarcinoma and Transitional Cell Carcinoma Cell Lines by C-CPE Gold-Nanoparticle-Mediated Laser Intervention.
Alnajjar S, Nolte I, Becker A, Schille JT, Trakooljul N, Frank M, Ngezahayo A, Murua Escobar H
International journal of molecular sciences 2021 Nov 13;22(22)
International journal of molecular sciences 2021 Nov 13;22(22)
Butyrate Alleviates Cytokine-Induced Barrier Dysfunction by Modifying Claudin-2 Levels.
Huang X, Oshima T, Tomita T, Fukui H, Miwa H
Biology 2021 Mar 9;10(3)
Biology 2021 Mar 9;10(3)
Targeting PVT1 Exon 9 Re-Expresses Claudin 4 Protein and Inhibits Migration by Claudin-Low Triple Negative Breast Cancer Cells.
Levine F, Ogunwobi OO
Cancers 2021 Mar 2;13(5)
Cancers 2021 Mar 2;13(5)
A human surfactant B deficiency air-liquid interface cell culture model suitable for gene therapy applications.
Munis AM, Hyde SC, Gill DR
Molecular therapy. Methods & clinical development 2021 Mar 12;20:237-246
Molecular therapy. Methods & clinical development 2021 Mar 12;20:237-246
Luminal polyethylene glycol solution delays the onset of preservation injury in the human intestine.
Søfteland JM, Bagge J, Padma AM, Casselbrant A, Zhu C, Wang Y, Hellström M, Olausson M, Oltean M
American journal of transplantation : official journal of the American Society of Transplantation and the American Society of Transplant Surgeons 2021 Jun;21(6):2220-2230
American journal of transplantation : official journal of the American Society of Transplantation and the American Society of Transplant Surgeons 2021 Jun;21(6):2220-2230
Microbiota regulate innate immune signaling and protective immunity against cancer.
Xing C, Wang M, Ajibade AA, Tan P, Fu C, Chen L, Zhu M, Hao ZZ, Chu J, Yu X, Yin B, Zhu J, Shen WJ, Duan T, Wang HY, Wang RF
Cell host & microbe 2021 Jun 9;29(6):959-974.e7
Cell host & microbe 2021 Jun 9;29(6):959-974.e7
Hypoxia and heat stress affect epithelial integrity in a Caco-2/HT-29 co-culture.
Lian P, Braber S, Varasteh S, Wichers HJ, Folkerts G
Scientific reports 2021 Jun 23;11(1):13186
Scientific reports 2021 Jun 23;11(1):13186
Isolation and development of bovine primary respiratory cells as model to study influenza D virus infection.
Uprety T, Sreenivasan CC, Bhattarai S, Wang D, Kaushik RS, Li F
Virology 2021 Jul;559:89-99
Virology 2021 Jul;559:89-99
Tight junctions in the blood-brain barrier promote edema formation and infarct size in stroke - Ambivalent effects of sealing proteins.
Winkler L, Blasig R, Breitkreuz-Korff O, Berndt P, Dithmer S, Helms HC, Puchkov D, Devraj K, Kaya M, Qin Z, Liebner S, Wolburg H, Andjelkovic AV, Rex A, Blasig IE, Haseloff RF
Journal of cerebral blood flow and metabolism : official journal of the International Society of Cerebral Blood Flow and Metabolism 2021 Jan;41(1):132-145
Journal of cerebral blood flow and metabolism : official journal of the International Society of Cerebral Blood Flow and Metabolism 2021 Jan;41(1):132-145
Quercetin Alleviates Oxidative Damage by Activating Nuclear Factor Erythroid 2-Related Factor 2 Signaling in Porcine Enterocytes.
Jia H, Zhang Y, Si X, Jin Y, Jiang D, Dai Z, Wu Z
Nutrients 2021 Jan 26;13(2)
Nutrients 2021 Jan 26;13(2)
Citrus limon Peel Powder Reduces Intestinal Barrier Defects and Inflammation in a Colitic Murine Experimental Model.
Tinh NTT, Sitolo GC, Yamamoto Y, Suzuki T
Foods (Basel, Switzerland) 2021 Jan 25;10(2)
Foods (Basel, Switzerland) 2021 Jan 25;10(2)
JAK-STAT Pathway Inhibition Partially Restores Intestinal Homeostasis in Hdac1- and Hdac2-Intestinal Epithelial Cell-Deficient Mice.
Gonneaud A, Turgeon N, Boisvert FM, Boudreau F, Asselin C
Cells 2021 Jan 23;10(2)
Cells 2021 Jan 23;10(2)
STIM-Orai1 signaling regulates fluidity of cytoplasm during membrane blebbing.
Aoki K, Harada S, Kawaji K, Matsuzawa K, Uchida S, Ikenouchi J
Nature communications 2021 Jan 20;12(1):480
Nature communications 2021 Jan 20;12(1):480
Culture surface protein coatings affect the barrier properties and calcium signalling of hESC-RPE.
Viheriälä T, Sorvari J, Ihalainen TO, Mörö A, Grönroos P, Schlie-Wolter S, Chichkov B, Skottman H, Nymark S, Ilmarinen T
Scientific reports 2021 Jan 13;11(1):933
Scientific reports 2021 Jan 13;11(1):933
Tumor Necrosis Factor Alpha Effects on the Porcine Intestinal Epithelial Barrier Include Enhanced Expression of TNF Receptor 1.
Droessler L, Cornelius V, Markov AG, Amasheh S
International journal of molecular sciences 2021 Aug 14;22(16)
International journal of molecular sciences 2021 Aug 14;22(16)
Vitamin D receptor is overexpressed in the duodenum of patients with irritable bowel syndrome.
Miura K, Oshima T, Ito C, Horikawa T, Yamada M, Tomita T, Fukui H, Miwa H
Journal of gastroenterology and hepatology 2021 Apr;36(4):951-958
Journal of gastroenterology and hepatology 2021 Apr;36(4):951-958
Occludin and tricellulin facilitate formation of anastomosing tight-junction strand network to improve barrier function.
Saito AC, Higashi T, Fukazawa Y, Otani T, Tauchi M, Higashi AY, Furuse M, Chiba H
Molecular biology of the cell 2021 Apr 15;32(8):722-738
Molecular biology of the cell 2021 Apr 15;32(8):722-738
Blood-Brain Barrier Protein Claudin-5 Expressed in Paired Xenopus laevis Oocytes Mediates Cell-Cell Interaction.
Brunner N, Stein L, Cornelius V, Knittel R, Fallier-Becker P, Amasheh S
Frontiers in physiology 2020;11:857
Frontiers in physiology 2020;11:857
N‑glycosylation and receptor tyrosine kinase signaling affect claudin‑3 levels in colorectal cancer cells.
Pérez AG, Andrade-Da-Costa J, De Souza WF, De Souza Ferreira M, Boroni M, De Oliveira IM, Freire-Neto CA, Fernandes PV, De Lanna CA, Souza-Santos PT, Morgado-Díaz JA, De-Freitas-Junior JCM
Oncology reports 2020 Oct;44(4):1649-1661
Oncology reports 2020 Oct;44(4):1649-1661
Metformin and Inhibition of Transforming Growth Factor-Beta Stimulate In Vitro Transport in Primary Renal Tubule Cells.
Love H, Evans R, Humes HD, Roy S, Zent R, Harris R, Wilson M, Fissell WH
Tissue engineering. Part A 2020 Oct;26(19-20):1091-1098
Tissue engineering. Part A 2020 Oct;26(19-20):1091-1098
Combinatorial expression of claudins in the proximal renal tubule and its functional consequences.
Curry JN, Tokuda S, McAnulty P, Yu ASL
American journal of physiology. Renal physiology 2020 May 1;318(5):F1138-F1146
American journal of physiology. Renal physiology 2020 May 1;318(5):F1138-F1146
Nicotine directly affects milk production in lactating mammary epithelial cells concurrently with inactivation of STAT5 and glucocorticoid receptor in vitro.
Kobayashi K, Tsugami Y, Suzuki N, Suzuki T, Nishimura T
Toxicology in vitro : an international journal published in association with BIBRA 2020 Mar;63:104741
Toxicology in vitro : an international journal published in association with BIBRA 2020 Mar;63:104741
NOTCH1 signaling establishes the medullary thymic epithelial cell progenitor pool during mouse fetal development.
Li J, Gordon J, Chen ELY, Xiao S, Wu L, Zúñiga-Pflücker JC, Manley NR
Development (Cambridge, England) 2020 Jun 22;147(12)
Development (Cambridge, England) 2020 Jun 22;147(12)
Circulating Ouabain Modulates Expression of Claudins in Rat Intestine and Cerebral Blood Vessels.
Markov AG, Fedorova AA, Kravtsova VV, Bikmurzina AE, Okorokova LS, Matchkov VV, Cornelius V, Amasheh S, Krivoi II
International journal of molecular sciences 2020 Jul 17;21(14)
International journal of molecular sciences 2020 Jul 17;21(14)
Maltodextrin-induced intestinal injury in a neonatal mouse model.
Singh P, Sanchez-Fernandez LL, Ramiro-Cortijo D, Ochoa-Allemant P, Perides G, Liu Y, Medina-Morales E, Yakah W, Freedman SD, Martin CR
Disease models & mechanisms 2020 Aug 27;13(8)
Disease models & mechanisms 2020 Aug 27;13(8)
Adverse Effects of Coumestrol and Genistein on Mammary Morphogenesis and Future Milk Production Ability of Mammary Epithelial Cells.
Kumai A, Tsugami Y, Wakasa H, Suzuki N, Suzuki T, Nishimura T, Kobayashi K
Advanced biosystems 2020 Apr;4(4):e1900187
Advanced biosystems 2020 Apr;4(4):e1900187
Effects of Claudin-1 on the Action of Clostridium perfringens Enterotoxin in Caco-2 Cells.
Mehdizadeh Gohari I, Li J, Navarro M, Uzal F, McClane B
Toxins 2019 Oct 9;11(10)
Toxins 2019 Oct 9;11(10)
Claudins and JAM-A coordinately regulate tight junction formation and epithelial polarity.
Otani T, Nguyen TP, Tokuda S, Sugihara K, Sugawara T, Furuse K, Miura T, Ebnet K, Furuse M
The Journal of cell biology 2019 Oct 7;218(10):3372-3396
The Journal of cell biology 2019 Oct 7;218(10):3372-3396
In vitro and in vivo effects of a mycotoxin, deoxynivalenol, and a trace metal, cadmium, alone or in a mixture on the intestinal barrier.
Luo S, Terciolo C, Bracarense APFL, Payros D, Pinton P, Oswald IP
Environment international 2019 Nov;132:105082
Environment international 2019 Nov;132:105082
Fusarium Mycotoxins Disrupt the Barrier and Induce IL-6 Release in a Human Placental Epithelium Cell Line.
Seyed Toutounchi N, Hogenkamp A, Varasteh S, Van't Land B, Garssen J, Kraneveld AD, Folkerts G, Braber S
Toxins 2019 Nov 14;11(11)
Toxins 2019 Nov 14;11(11)
Newly synthesized claudins but not occludin are added to the basal side of the tight junction.
Van Itallie CM, Lidman KF, Tietgens AJ, Anderson JM
Molecular biology of the cell 2019 Jun 1;30(12):1406-1424
Molecular biology of the cell 2019 Jun 1;30(12):1406-1424
Effect of Manitoba-Grown Red-Osier Dogwood Extracts on Recovering Caco-2 Cells from H(2)O(2)-Induced Oxidative Damage.
Yang R, Hui Q, Jiang Q, Liu S, Zhang H, Wu J, Lin F, O K, Yang C
Antioxidants (Basel, Switzerland) 2019 Jul 28;8(8)
Antioxidants (Basel, Switzerland) 2019 Jul 28;8(8)
Association of Cytokeratin 5 and Claudin 3 expression with BRCA1 and BRCA2 germline mutations in women with early breast cancer.
Danzinger S, Tan YY, Rudas M, Kastner MT, Weingartshofer S, Muhr D, Singer CF
BMC cancer 2019 Jul 15;19(1):695
BMC cancer 2019 Jul 15;19(1):695
The kidney anion exchanger 1 affects tight junction properties via claudin-4.
Lashhab R, Rumley AC, Arutyunov D, Rizvi M, You C, Dimke H, Touret N, Zimmermann R, Jung M, Chen XZ, Alexander T, Cordat E
Scientific reports 2019 Feb 28;9(1):3099
Scientific reports 2019 Feb 28;9(1):3099
Morphologic determinant of tight junctions revealed by claudin-3 structures.
Nakamura S, Irie K, Tanaka H, Nishikawa K, Suzuki H, Saitoh Y, Tamura A, Tsukita S, Fujiyoshi Y
Nature communications 2019 Feb 18;10(1):816
Nature communications 2019 Feb 18;10(1):816
Electrical Stimulation of the Mesencephalic Locomotor Region Has No Impact on Blood-Brain Barrier Alterations after Cerebral Photothrombosis in Rats.
Schuhmann MK, Stoll G, Papp L, Bohr A, Volkmann J, Fluri F
International journal of molecular sciences 2019 Aug 19;20(16)
International journal of molecular sciences 2019 Aug 19;20(16)
p120ctn-Mediated Organ Patterning Precedes and Determines Pancreatic Progenitor Fate.
Nyeng P, Heilmann S, Löf-Öhlin ZM, Pettersson NF, Hermann FM, Reynolds AB, Semb H
Developmental cell 2019 Apr 8;49(1):31-47.e9
Developmental cell 2019 Apr 8;49(1):31-47.e9
Vitamin D Receptor Deletion Leads to the Destruction of Tight and Adherens Junctions in Lungs.
Chen H, Lu R, Zhang YG, Sun J
Tissue barriers 2018;6(4):1-13
Tissue barriers 2018;6(4):1-13
Differential Claudin 3 and EGFR Expression Predicts BRCA1 Mutation in Triple-Negative Breast Cancer.
Danzinger S, Tan YY, Rudas M, Kastner MT, Weingartshofer S, Muhr D, Singer CF, kConFab Investigators
Cancer investigation 2018;36(7):378-388
Cancer investigation 2018;36(7):378-388
Loss of connexin43 in murine Sertoli cells and its effect on blood-testis barrier formation and dynamics.
Hollenbach J, Jung K, Noelke J, Gasse H, Pfarrer C, Koy M, Brehm R
PloS one 2018;13(6):e0198100
PloS one 2018;13(6):e0198100
Glial Cells in the Fish Retinal Nerve Fiber Layer Form Tight Junctions, Separating and Surrounding Axons.
Garcia-Pradas L, Gleiser C, Wizenmann A, Wolburg H, Mack AF
Frontiers in molecular neuroscience 2018;11:367
Frontiers in molecular neuroscience 2018;11:367
Functionalization of gold-nanoparticles by the Clostridium perfringens enterotoxin C-terminus for tumor cell ablation using the gold nanoparticle-mediated laser perforation technique.
Becker A, Leskau M, Schlingmann-Molina BL, Hohmeier SC, Alnajjar S, Murua Escobar H, Ngezahayo A
Scientific reports 2018 Oct 8;8(1):14963
Scientific reports 2018 Oct 8;8(1):14963
Salivary glands regenerate after radiation injury through SOX2-mediated secretory cell replacement.
Emmerson E, May AJ, Berthoin L, Cruz-Pacheco N, Nathan S, Mattingly AJ, Chang JL, Ryan WR, Tward AD, Knox SM
EMBO molecular medicine 2018 Mar;10(3)
EMBO molecular medicine 2018 Mar;10(3)
Claudin-3 Loss Causes Leakage of Sweat from the Sweat Gland to Contribute to the Pathogenesis of Atopic Dermatitis.
Yamaga K, Murota H, Tamura A, Miyata H, Ohmi M, Kikuta J, Ishii M, Tsukita S, Katayama I
The Journal of investigative dermatology 2018 Jun;138(6):1279-1287
The Journal of investigative dermatology 2018 Jun;138(6):1279-1287
Immunohistochemistry in Investigative and Toxicologic Pathology.
Janardhan KS, Jensen H, Clayton NP, Herbert RA
Toxicologic pathology 2018 Jul;46(5):488-510
Toxicologic pathology 2018 Jul;46(5):488-510
Adherens junctions influence tight junction formation via changes in membrane lipid composition.
Shigetomi K, Ono Y, Inai T, Ikenouchi J
The Journal of cell biology 2018 Jul 2;217(7):2373-2381
The Journal of cell biology 2018 Jul 2;217(7):2373-2381
Claudin Loss-of-Function Disrupts Tight Junctions and Impairs Amelogenesis.
Bardet C, Ribes S, Wu Y, Diallo MT, Salmon B, Breiderhoff T, Houillier P, Müller D, Chaussain C
Frontiers in physiology 2017;8:326
Frontiers in physiology 2017;8:326
Altered Cytokine Expression and Barrier Properties after In Vitro Infection of Porcine Epithelial Cells with Enterotoxigenic Escherichia coli and Probiotic Enterococcus faecium.
Kern M, Günzel D, Aschenbach JR, Tedin K, Bondzio A, Lodemann U
Mediators of inflammation 2017;2017:2748192
Mediators of inflammation 2017;2017:2748192
The role of human fibronectin- or placenta basement membrane extract-based gels in favouring the formation of polarized salivary acinar-like structures.
Maria OM, Liu Y, El-Hakim M, Zeitouni A, Tran SD
Journal of tissue engineering and regenerative medicine 2017 Sep;11(9):2643-2657
Journal of tissue engineering and regenerative medicine 2017 Sep;11(9):2643-2657
Talin regulates integrin β1-dependent and -independent cell functions in ureteric bud development.
Mathew S, Palamuttam RJ, Mernaugh G, Ramalingam H, Lu Z, Zhang MZ, Ishibe S, Critchley DR, Fässler R, Pozzi A, Sanders CR, Carroll TJ, Zent R
Development (Cambridge, England) 2017 Nov 15;144(22):4148-4158
Development (Cambridge, England) 2017 Nov 15;144(22):4148-4158
Establishment, maintenance and functional integrity of the blood-testis barrier in organotypic cultures of fresh and frozen/thawed prepubertal mouse testes.
Rondanino C, Maouche A, Dumont L, Oblette A, Rives N
Molecular human reproduction 2017 May 1;23(5):304-320
Molecular human reproduction 2017 May 1;23(5):304-320
Obesity-induces Organ and Tissue Specific Tight Junction Restructuring and Barrier Deregulation by Claudin Switching.
Ahmad R, Rah B, Bastola D, Dhawan P, Singh AB
Scientific reports 2017 Jul 11;7(1):5125
Scientific reports 2017 Jul 11;7(1):5125
Downregulation of lipolysis-stimulated lipoprotein receptor promotes cell invasion via claudin-1-mediated matrix metalloproteinases in human endometrial cancer.
Shimada H, Satohisa S, Kohno T, Konno T, Takano KI, Takahashi S, Hatakeyama T, Arimoto C, Saito T, Kojima T
Oncology letters 2017 Dec;14(6):6776-6782
Oncology letters 2017 Dec;14(6):6776-6782
Analysis of Epithelial-Mesenchymal Transition Induced by Transforming Growth Factor β.
Valcourt U, Carthy J, Okita Y, Alcaraz L, Kato M, Thuault S, Bartholin L, Moustakas A
Methods in molecular biology (Clifton, N.J.) 2016;1344:147-81
Methods in molecular biology (Clifton, N.J.) 2016;1344:147-81
Highly Conserved Testicular Localization of Claudin-11 in Normal and Impaired Spermatogenesis.
Stammler A, Lüftner BU, Kliesch S, Weidner W, Bergmann M, Middendorff R, Konrad L
PloS one 2016;11(8):e0160349
PloS one 2016;11(8):e0160349
Effects of Osmolality on Paracellular Transport in MDCK II Cells.
Tokuda S, Hirai T, Furuse M
PloS one 2016;11(11):e0166904
PloS one 2016;11(11):e0166904
Liver kinase B1 regulates hepatocellular tight junction distribution and function in vivo.
Porat-Shliom N, Tietgens AJ, Van Itallie CM, Vitale-Cross L, Jarnik M, Harding OJ, Anderson JM, Gutkind JS, Weigert R, Arias IM
Hepatology (Baltimore, Md.) 2016 Oct;64(4):1317-29
Hepatology (Baltimore, Md.) 2016 Oct;64(4):1317-29
Alix-mediated assembly of the actomyosin-tight junction polarity complex preserves epithelial polarity and epithelial barrier.
Campos Y, Qiu X, Gomero E, Wakefield R, Horner L, Brutkowski W, Han YG, Solecki D, Frase S, Bongiovanni A, d'Azzo A
Nature communications 2016 Jun 23;7:11876
Nature communications 2016 Jun 23;7:11876
Modeling immune functions of the mouse blood-cerebrospinal fluid barrier in vitro: primary rather than immortalized mouse choroid plexus epithelial cells are suited to study immune cell migration across this brain barrier.
Lazarevic I, Engelhardt B
Fluids and barriers of the CNS 2016 Jan 29;13:2
Fluids and barriers of the CNS 2016 Jan 29;13:2
miR-200 promotes the mesenchymal to epithelial transition by suppressing multiple members of the Zeb2 and Snail1 transcriptional repressor complexes.
Perdigão-Henriques R, Petrocca F, Altschuler G, Thomas MP, Le MT, Tan SM, Hide W, Lieberman J
Oncogene 2016 Jan 14;35(2):158-72
Oncogene 2016 Jan 14;35(2):158-72
Prolactin and glucocorticoid signaling induces lactation-specific tight junctions concurrent with β-casein expression in mammary epithelial cells.
Kobayashi K, Tsugami Y, Matsunaga K, Oyama S, Kuki C, Kumura H
Biochimica et biophysica acta 2016 Aug;1863(8):2006-16
Biochimica et biophysica acta 2016 Aug;1863(8):2006-16
Connexins, E-cadherin, Claudin-7 and β-catenin transiently form junctional nexuses during the post-natal mammary gland development.
Dianati E, Poiraud J, Weber-Ouellette A, Plante I
Developmental biology 2016 Aug 1;416(1):52-68
Developmental biology 2016 Aug 1;416(1):52-68
Claudin-2 knockout by TALEN-mediated gene targeting in MDCK cells: claudin-2 independently determines the leaky property of tight junctions in MDCK cells.
Tokuda S, Furuse M
PloS one 2015;10(3):e0119869
PloS one 2015;10(3):e0119869
Effects of Hydrostatic Pressure on Carcinogenic Properties of Epithelia.
Tokuda S, Kim YH, Matsumoto H, Muro S, Hirai T, Mishima M, Furuse M
PloS one 2015;10(12):e0145522
PloS one 2015;10(12):e0145522
Heterogeneity between triple negative breast cancer cells due to differential activation of Wnt and PI3K/AKT pathways.
Martínez-Revollar G, Garay E, Martin-Tapia D, Nava P, Huerta M, Lopez-Bayghen E, Meraz-Cruz N, Segovia J, González-Mariscal L
Experimental cell research 2015 Nov 15;339(1):67-80
Experimental cell research 2015 Nov 15;339(1):67-80
Mode of action of claudin peptidomimetics in the transient opening of cellular tight junction barriers.
Staat C, Coisne C, Dabrowski S, Stamatovic SM, Andjelkovic AV, Wolburg H, Engelhardt B, Blasig IE
Biomaterials 2015 Jun;54:9-20
Biomaterials 2015 Jun;54:9-20
A non-tight junction function of claudin-7-Interaction with integrin signaling in suppressing lung cancer cell proliferation and detachment.
Lu Z, Kim DH, Fan J, Lu Q, Verbanac K, Ding L, Renegar R, Chen YH
Molecular cancer 2015 Jun 17;14:120
Molecular cancer 2015 Jun 17;14:120
Claudin-4 is required for modulation of paracellular permeability by muscarinic acetylcholine receptor in epithelial cells.
Cong X, Zhang Y, Li J, Mei M, Ding C, Xiang RL, Zhang LW, Wang Y, Wu LL, Yu GY
Journal of cell science 2015 Jun 15;128(12):2271-86
Journal of cell science 2015 Jun 15;128(12):2271-86
Galacto-oligosaccharides Protect the Intestinal Barrier by Maintaining the Tight Junction Network and Modulating the Inflammatory Responses after a Challenge with the Mycotoxin Deoxynivalenol in Human Caco-2 Cell Monolayers and B6C3F1 Mice.
Akbari P, Braber S, Alizadeh A, Verheijden KA, Schoterman MH, Kraneveld AD, Garssen J, Fink-Gremmels J
The Journal of nutrition 2015 Jul;145(7):1604-13
The Journal of nutrition 2015 Jul;145(7):1604-13
Overexpression and delocalization of claudin-3 protein in MCF-7 and MDA-MB-415 breast cancer cell lines.
Todd MC, Petty HM, King JM, Piana Marshall BN, Sheller RA, Cuevas ME
Oncology letters 2015 Jul;10(1):156-162
Oncology letters 2015 Jul;10(1):156-162
Homoharringtonine increases intestinal epithelial permeability by modulating specific claudin isoforms in Caco-2 cell monolayers.
Watari A, Hashegawa M, Yagi K, Kondoh M
European journal of pharmaceutics and biopharmaceutics : official journal of Arbeitsgemeinschaft fur Pharmazeutische Verfahrenstechnik e.V 2015 Jan;89:232-8
European journal of pharmaceutics and biopharmaceutics : official journal of Arbeitsgemeinschaft fur Pharmazeutische Verfahrenstechnik e.V 2015 Jan;89:232-8
Expression of tight-junction proteins in human proximal small intestinal mucosa before and after Roux-en-Y gastric bypass surgery.
Casselbrant A, Elias E, Fändriks L, Wallenius V
Surgery for obesity and related diseases : official journal of the American Society for Bariatric Surgery 2015 Jan-Feb;11(1):45-53
Surgery for obesity and related diseases : official journal of the American Society for Bariatric Surgery 2015 Jan-Feb;11(1):45-53
Intestinal fatty acid binding protein as a marker for intra-abdominal pressure-related complications in patients admitted to the intensive care unit; study protocol for a prospective cohort study (I-Fabulous study).
Strang SG, Van Waes OJ, Van der Hoven B, Ali S, Verhofstad MH, Pickkers P, Van Lieshout EM
Scandinavian journal of trauma, resuscitation and emergency medicine 2015 Jan 16;23:6
Scandinavian journal of trauma, resuscitation and emergency medicine 2015 Jan 16;23:6
ZO-1 knockout by TALEN-mediated gene targeting in MDCK cells: involvement of ZO-1 in the regulation of cytoskeleton and cell shape.
Tokuda S, Higashi T, Furuse M
PloS one 2014;9(8):e104994
PloS one 2014;9(8):e104994
Impaired intestinal mucosal barrier upon ischemia-reperfusion: "patching holes in the shield with a simple surgical method".
Rosero O, Ónody P, Kovács T, Molnár D, Lotz G, Tóth S, Turóczi Z, Fülöp A, Garbaisz D, Harsányi L, Szijártó A
BioMed research international 2014;2014:210901
BioMed research international 2014;2014:210901
Autonomous isolation, long-term culture and differentiation potential of adult salivary gland-derived stem/progenitor cells.
Baek H, Noh YH, Lee JH, Yeon SI, Jeong J, Kwon H
Journal of tissue engineering and regenerative medicine 2014 Sep;8(9):717-27
Journal of tissue engineering and regenerative medicine 2014 Sep;8(9):717-27
Biological significance of FoxN1 gain-of-function mutations during T and B lymphopoiesis in juvenile mice.
Ruan L, Zhang Z, Mu L, Burnley P, Wang L, Coder B, Zhuge Q, Su DM
Cell death & disease 2014 Oct 9;5(10):e1457
Cell death & disease 2014 Oct 9;5(10):e1457
Diets high in fermentable protein and fibre alter tight junction protein composition with minor effects on barrier function in piglet colon.
Richter JF, Pieper R, Zakrzewski SS, Günzel D, Schulzke JD, Van Kessel AG
The British journal of nutrition 2014 Mar 28;111(6):1040-9
The British journal of nutrition 2014 Mar 28;111(6):1040-9
Histopathologic and molecular evaluation of the Organ Procurement and Transplantation Network selection criteria for intestinal graft donation.
Roskott AM, van Haaften WT, Leuvenink HG, Ploeg RJ, van Goor H, Blokzijl T, Ottens PJ, Dijkstra G, Nieuwenhuijs VB
The Journal of surgical research 2014 Jun 1;189(1):143-51
The Journal of surgical research 2014 Jun 1;189(1):143-51
Disrupted tight junctions in the small intestine of cystic fibrosis mice.
De Lisle RC
Cell and tissue research 2014 Jan;355(1):131-42
Cell and tissue research 2014 Jan;355(1):131-42
Activation of Gpr109a, receptor for niacin and the commensal metabolite butyrate, suppresses colonic inflammation and carcinogenesis.
Singh N, Gurav A, Sivaprakasam S, Brady E, Padia R, Shi H, Thangaraju M, Prasad PD, Manicassamy S, Munn DH, Lee JR, Offermanns S, Ganapathy V
Immunity 2014 Jan 16;40(1):128-39
Immunity 2014 Jan 16;40(1):128-39
Immunohistological characterization of intercellular junction proteins in rhesus macaque intestine.
Gumber S, Nusrat A, Villinger F
Experimental and toxicologic pathology : official journal of the Gesellschaft fur Toxikologische Pathologie 2014 Dec;66(9-10):437-44
Experimental and toxicologic pathology : official journal of the Gesellschaft fur Toxikologische Pathologie 2014 Dec;66(9-10):437-44
Ectopic TBX1 suppresses thymic epithelial cell differentiation and proliferation during thymus organogenesis.
Reeh KA, Cardenas KT, Bain VE, Liu Z, Laurent M, Manley NR, Richie ER
Development (Cambridge, England) 2014 Aug;141(15):2950-8
Development (Cambridge, England) 2014 Aug;141(15):2950-8
Claudin-3 overexpression increases the malignant potential of colorectal cancer cells: roles of ERK1/2 and PI3K-Akt as modulators of EGFR signaling.
de Souza WF, Fortunato-Miranda N, Robbs BK, de Araujo WM, de-Freitas-Junior JC, Bastos LG, Viola JP, Morgado-Díaz JA
PloS one 2013;8(9):e74994
PloS one 2013;8(9):e74994
Transmigration of neural stem cells across the blood brain barrier induced by glioma cells.
Díaz-Coránguez M, Segovia J, López-Ornelas A, Puerta-Guardo H, Ludert J, Chávez B, Meraz-Cruz N, González-Mariscal L
PloS one 2013;8(4):e60655
PloS one 2013;8(4):e60655
The tight-junction protein claudin-6 induces epithelial differentiation from mouse F9 and embryonic stem cells.
Sugimoto K, Ichikawa-Tomikawa N, Satohisa S, Akashi Y, Kanai R, Saito T, Sawada N, Chiba H
PloS one 2013;8(10):e75106
PloS one 2013;8(10):e75106
Guanylate cyclase C limits systemic dissemination of a murine enteric pathogen.
Mann EA, Harmel-Laws E, Cohen MB, Steinbrecher KA
BMC gastroenterology 2013 Sep 2;13:135
BMC gastroenterology 2013 Sep 2;13:135
Build them up and break them down: Tight junctions of cell lines expressing typical hepatocyte polarity with a varied repertoire of claudins.
Grosse B, Degrouard J, Jaillard D, Cassio D
Tissue barriers 2013 Oct 1;1(4):e25210
Tissue barriers 2013 Oct 1;1(4):e25210
Role of the p63-FoxN1 regulatory axis in thymic epithelial cell homeostasis during aging.
Burnley P, Rahman M, Wang H, Zhang Z, Sun X, Zhuge Q, Su DM
Cell death & disease 2013 Nov 21;4(11):e932
Cell death & disease 2013 Nov 21;4(11):e932
SOX8 regulates permeability of the blood-testes barrier that affects adult male fertility in the mouse.
Singh AP, Cummings CA, Mishina Y, Archer TK
Biology of reproduction 2013 May;88(5):133
Biology of reproduction 2013 May;88(5):133
A complex containing LPP and α-actinin mediates TGFβ-induced migration and invasion of ErbB2-expressing breast cancer cells.
Ngan E, Northey JJ, Brown CM, Ursini-Siegel J, Siegel PM
Journal of cell science 2013 May 1;126(Pt 9):1981-91
Journal of cell science 2013 May 1;126(Pt 9):1981-91
Rab25 regulates integrin expression in polarized colonic epithelial cells.
Krishnan M, Lapierre LA, Knowles BC, Goldenring JR
Molecular biology of the cell 2013 Mar;24(6):818-31
Molecular biology of the cell 2013 Mar;24(6):818-31
Derivation of myoepithelial progenitor cells from bipotent mammary stem/progenitor cells.
Zhao X, Malhotra GK, Band H, Band V
PloS one 2012;7(4):e35338
PloS one 2012;7(4):e35338
EpCAM contributes to formation of functional tight junction in the intestinal epithelium by recruiting claudin proteins.
Lei Z, Maeda T, Tamura A, Nakamura T, Yamazaki Y, Shiratori H, Yashiro K, Tsukita S, Hamada H
Developmental biology 2012 Nov 15;371(2):136-45
Developmental biology 2012 Nov 15;371(2):136-45
Differential effects of flavonoids on barrier integrity in human intestinal Caco-2 cells.
Noda S, Tanabe S, Suzuki T
Journal of agricultural and food chemistry 2012 May 9;60(18):4628-33
Journal of agricultural and food chemistry 2012 May 9;60(18):4628-33
Dynamic distribution of claudin proteins in pancreatic epithelia undergoing morphogenesis or neoplastic transformation.
Westmoreland JJ, Drosos Y, Kelly J, Ye J, Means AL, Washington MK, Sosa-Pineda B
Developmental dynamics : an official publication of the American Association of Anatomists 2012 Mar;241(3):583-94
Developmental dynamics : an official publication of the American Association of Anatomists 2012 Mar;241(3):583-94
Age-Related Disruption of Steady-State Thymic Medulla Provokes Autoimmune Phenotype via Perturbing Negative Selection.
Xia J, Wang H, Guo J, Zhang Z, Coder B, Su DM
Aging and disease 2012 Jun;3(3):248-59
Aging and disease 2012 Jun;3(3):248-59
Effects of proinflammatory cytokines on the claudin-19 rich tight junctions of human retinal pigment epithelium.
Peng S, Gan G, Rao VS, Adelman RA, Rizzolo LJ
Investigative ophthalmology & visual science 2012 Jul 27;53(8):5016-28
Investigative ophthalmology & visual science 2012 Jul 27;53(8):5016-28
Two strikingly different signaling pathways are induced by meningococcal type IV pili on endothelial and epithelial cells.
Lécuyer H, Nassif X, Coureuil M
Infection and immunity 2012 Jan;80(1):175-86
Infection and immunity 2012 Jan;80(1):175-86
Functional characterization and localization of a gill-specific claudin isoform in Atlantic salmon.
Engelund MB, Yu AS, Li J, Madsen SS, Færgeman NJ, Tipsmark CK
American journal of physiology. Regulatory, integrative and comparative physiology 2012 Jan 15;302(2):R300-11
American journal of physiology. Regulatory, integrative and comparative physiology 2012 Jan 15;302(2):R300-11
Zonula occludens-1 and -2 regulate apical cell structure and the zonula adherens cytoskeleton in polarized epithelia.
Fanning AS, Van Itallie CM, Anderson JM
Molecular biology of the cell 2012 Feb;23(4):577-90
Molecular biology of the cell 2012 Feb;23(4):577-90
Probiotic bacteria induce maturation of intestinal claudin 3 expression and barrier function.
Patel RM, Myers LS, Kurundkar AR, Maheshwari A, Nusrat A, Lin PW
The American journal of pathology 2012 Feb;180(2):626-35
The American journal of pathology 2012 Feb;180(2):626-35
Complexity and developmental changes in the expression pattern of claudins at the blood-CSF barrier.
Kratzer I, Vasiljevic A, Rey C, Fevre-Montange M, Saunders N, Strazielle N, Ghersi-Egea JF
Histochemistry and cell biology 2012 Dec;138(6):861-79
Histochemistry and cell biology 2012 Dec;138(6):861-79
Distinct domains of paracingulin are involved in its targeting to the actin cytoskeleton and regulation of apical junction assembly.
Paschoud S, Guillemot L, Citi S
The Journal of biological chemistry 2012 Apr 13;287(16):13159-69
The Journal of biological chemistry 2012 Apr 13;287(16):13159-69
Enhanced immunohistochemical resolution of claudin proteins in glycolmethacrylate-embedded tissue biopsies.
Collins JE, Kirk A, Campbell SK, Mason J, Wilson SJ
Methods in molecular biology (Clifton, N.J.) 2011;762:371-82
Methods in molecular biology (Clifton, N.J.) 2011;762:371-82
Claudin-1 induced sealing of blood-brain barrier tight junctions ameliorates chronic experimental autoimmune encephalomyelitis.
Pfeiffer F, Schäfer J, Lyck R, Makrides V, Brunner S, Schaeren-Wiemers N, Deutsch U, Engelhardt B
Acta neuropathologica 2011 Nov;122(5):601-14
Acta neuropathologica 2011 Nov;122(5):601-14
Matrigel improves functional properties of primary human salivary gland cells.
Maria OM, Zeitouni A, Gologan O, Tran SD
Tissue engineering. Part A 2011 May;17(9-10):1229-38
Tissue engineering. Part A 2011 May;17(9-10):1229-38
Morphogenesis and maintenance of the 3D thymic medulla and prevention of nude skin phenotype require FoxN1 in pre- and post-natal K14 epithelium.
Guo J, Rahman M, Cheng L, Zhang S, Tvinnereim A, Su DM
Journal of molecular medicine (Berlin, Germany) 2011 Mar;89(3):263-77
Journal of molecular medicine (Berlin, Germany) 2011 Mar;89(3):263-77
Identification of a claudin-4 and E-cadherin score to predict prognosis in breast cancer.
Szasz AM, Nemeth Z, Gyorffy B, Micsinai M, Krenacs T, Baranyai Z, Harsanyi L, Kiss A, Schaff Z, Tokes AM, Kulka J
Cancer science 2011 Dec;102(12):2248-54
Cancer science 2011 Dec;102(12):2248-54
The Hedgehog pathway promotes blood-brain barrier integrity and CNS immune quiescence.
Alvarez JI, Dodelet-Devillers A, Kebir H, Ifergan I, Fabre PJ, Terouz S, Sabbagh M, Wosik K, Bourbonnière L, Bernard M, van Horssen J, de Vries HE, Charron F, Prat A
Science (New York, N.Y.) 2011 Dec 23;334(6063):1727-31
Science (New York, N.Y.) 2011 Dec 23;334(6063):1727-31
Hypoxia increases transepithelial electrical conductance and reduces occludin at the plasma membrane in alveolar epithelial cells via PKC-ζ and PP2A pathway.
Caraballo JC, Yshii C, Butti ML, Westphal W, Borcherding JA, Allamargot C, Comellas AP
American journal of physiology. Lung cellular and molecular physiology 2011 Apr;300(4):L569-78
American journal of physiology. Lung cellular and molecular physiology 2011 Apr;300(4):L569-78
Ovarian hormones control the changing expression of claudins and occludin in rat uterine epithelial cells during early pregnancy.
Nicholson MD, Lindsay LA, Murphy CR
Acta histochemica 2010;112(1):42-52
Acta histochemica 2010;112(1):42-52
Effects of long-term progesterone on developmental and functional aspects of porcine uterine epithelia and vasculature: progesterone alone does not support development of uterine glands comparable to that of pregnancy.
Bailey DW, Dunlap KA, Frank JW, Erikson DW, White BG, Bazer FW, Burghardt RC, Johnson GA
Reproduction (Cambridge, England) 2010 Oct;140(4):583-94
Reproduction (Cambridge, England) 2010 Oct;140(4):583-94
Astroglial structures in the zebrafish brain.
Grupp L, Wolburg H, Mack AF
The Journal of comparative neurology 2010 Nov 1;518(21):4277-87
The Journal of comparative neurology 2010 Nov 1;518(21):4277-87
beta-catenin expression and claudin expression pattern as prognostic factors of prostatic cancer progression.
Szász AM, Nyirády P, Majoros A, Szendrõi A, Szûcs M, Székely E, Tõkés AM, Romics I, Kulka J
BJU international 2010 Mar;105(5):716-22
BJU international 2010 Mar;105(5):716-22
Minimal effects of VEGF and anti-VEGF drugs on the permeability or selectivity of RPE tight junctions.
Peng S, Adelman RA, Rizzolo LJ
Investigative ophthalmology & visual science 2010 Jun;51(6):3216-25
Investigative ophthalmology & visual science 2010 Jun;51(6):3216-25
Non-invasive markers for early diagnosis and determination of the severity of necrotizing enterocolitis.
Thuijls G, Derikx JP, van Wijck K, Zimmermann LJ, Degraeuwe PL, Mulder TL, Van der Zee DC, Brouwers HA, Verhoeven BH, van Heurn LW, Kramer BW, Buurman WA, Heineman E
Annals of surgery 2010 Jun;251(6):1174-80
Annals of surgery 2010 Jun;251(6):1174-80
Differential expression of claudin tight junction proteins in the human cortical nephron.
Kirk A, Campbell S, Bass P, Mason J, Collins J
Nephrology, dialysis, transplantation : official publication of the European Dialysis and Transplant Association - European Renal Association 2010 Jul;25(7):2107-19
Nephrology, dialysis, transplantation : official publication of the European Dialysis and Transplant Association - European Renal Association 2010 Jul;25(7):2107-19
Claudin-1 has tumor suppressive activity and is a direct target of RUNX3 in gastric epithelial cells.
Chang TL, Ito K, Ko TK, Liu Q, Salto-Tellez M, Yeoh KG, Fukamachi H, Ito Y
Gastroenterology 2010 Jan;138(1):255-65.e1-3
Gastroenterology 2010 Jan;138(1):255-65.e1-3
Postnatal tissue-specific disruption of transcription factor FoxN1 triggers acute thymic atrophy.
Cheng L, Guo J, Sun L, Fu J, Barnes PF, Metzger D, Chambon P, Oshima RG, Amagai T, Su DM
The Journal of biological chemistry 2010 Feb 19;285(8):5836-47
The Journal of biological chemistry 2010 Feb 19;285(8):5836-47
Allogeneic human mesenchymal stem cells restore epithelial protein permeability in cultured human alveolar type II cells by secretion of angiopoietin-1.
Fang X, Neyrinck AP, Matthay MA, Lee JW
The Journal of biological chemistry 2010 Aug 20;285(34):26211-22
The Journal of biological chemistry 2010 Aug 20;285(34):26211-22
Differential distribution of tight junction proteins suggests a role for tanycytes in blood-hypothalamus barrier regulation in the adult mouse brain.
Mullier A, Bouret SG, Prevot V, Dehouck B
The Journal of comparative neurology 2010 Apr 1;518(7):943-62
The Journal of comparative neurology 2010 Apr 1;518(7):943-62
Tight junction proteins in human Schwann cell autotypic junctions.
Alanne MH, Pummi K, Heape AM, Grènman R, Peltonen J, Peltonen S
The journal of histochemistry and cytochemistry : official journal of the Histochemistry Society 2009 Jun;57(6):523-9
The journal of histochemistry and cytochemistry : official journal of the Histochemistry Society 2009 Jun;57(6):523-9
Claudin-1, -2, -3, -4, -7, -8, and -10 protein expression in biliary tract cancers.
Németh Z, Szász AM, Tátrai P, Németh J, Gyorffy H, Somorácz A, Szíjártó A, Kupcsulik P, Kiss A, Schaff Z
The journal of histochemistry and cytochemistry : official journal of the Histochemistry Society 2009 Feb;57(2):113-21
The journal of histochemistry and cytochemistry : official journal of the Histochemistry Society 2009 Feb;57(2):113-21
Intestinal cytoskeleton degradation precedes tight junction loss following hemorrhagic shock.
Thuijls G, de Haan JJ, Derikx JP, Daissormont I, Hadfoune M, Heineman E, Buurman WA
Shock (Augusta, Ga.) 2009 Feb;31(2):164-9
Shock (Augusta, Ga.) 2009 Feb;31(2):164-9
A SNAIL1-SMAD3/4 transcriptional repressor complex promotes TGF-beta mediated epithelial-mesenchymal transition.
Vincent T, Neve EP, Johnson JR, Kukalev A, Rojo F, Albanell J, Pietras K, Virtanen I, Philipson L, Leopold PL, Crystal RG, de Herreros AG, Moustakas A, Pettersson RF, Fuxe J
Nature cell biology 2009 Aug;11(8):943-50
Nature cell biology 2009 Aug;11(8):943-50
New Insight in Loss of Gut Barrier during Major Non-Abdominal Surgery.
Derikx JP, van Waardenburg DA, Thuijls G, Willigers HM, Koenraads M, van Bijnen AA, Heineman E, Poeze M, Ambergen T, van Ooij A, van Rhijn LW, Buurman WA
PloS one 2008;3(12):e3954
PloS one 2008;3(12):e3954
Blood-testis barrier dynamics are regulated by testosterone and cytokines via their differential effects on the kinetics of protein endocytosis and recycling in Sertoli cells.
Yan HH, Mruk DD, Lee WM, Cheng CY
FASEB journal : official publication of the Federation of American Societies for Experimental Biology 2008 Jun;22(6):1945-59
FASEB journal : official publication of the Federation of American Societies for Experimental Biology 2008 Jun;22(6):1945-59
Regulation of testicular tight junctions by gonadotrophins in the adult Djungarian hamster in vivo.
Tarulli GA, Meachem SJ, Schlatt S, Stanton PG
Reproduction (Cambridge, England) 2008 Jun;135(6):867-77
Reproduction (Cambridge, England) 2008 Jun;135(6):867-77
Inducible overexpression of cingulin in stably transfected MDCK cells does not affect tight junction organization and gene expression.
Paschoud S, Citi S
Molecular membrane biology 2008 Jan;25(1):1-13
Molecular membrane biology 2008 Jan;25(1):1-13
Distribution of tight junction proteins in adult human salivary glands.
Maria OM, Kim JW, Gerstenhaber JA, Baum BJ, Tran SD
The journal of histochemistry and cytochemistry : official journal of the Histochemistry Society 2008 Dec;56(12):1093-8
The journal of histochemistry and cytochemistry : official journal of the Histochemistry Society 2008 Dec;56(12):1093-8
Expression of Claudin-1, Claudin-3 and Claudin-5 in human blood-brain barrier mimicking cell line ECV304 is inducible by glioma-conditioned media.
Neuhaus W, Wirth M, Plattner VE, Germann B, Gabor F, Noe CR
Neuroscience letters 2008 Dec 3;446(2-3):59-64
Neuroscience letters 2008 Dec 3;446(2-3):59-64
Distinct molecular composition of blood and lymphatic vascular endothelial cell junctions establishes specific functional barriers within the peripheral lymph node.
Pfeiffer F, Kumar V, Butz S, Vestweber D, Imhof BA, Stein JV, Engelhardt B
European journal of immunology 2008 Aug;38(8):2142-55
European journal of immunology 2008 Aug;38(8):2142-55
Structural alterations of epididymal epithelial cells in cathepsin A-deficient mice affect the blood-epididymal barrier and lead to altered sperm motility.
Hermo L, Korah N, Gregory M, Liu LY, Cyr DG, D'Azzo A, Smith CE
Journal of andrology 2007 Sep-Oct;28(5):784-97
Journal of andrology 2007 Sep-Oct;28(5):784-97
Cadmium causes delayed effects on renal function in the offspring of cadmium-contaminated pregnant female rats.
Jacquillet G, Barbier O, Rubera I, Tauc M, Borderie A, Namorado MC, Martin D, Sierra G, Reyes JL, Poujeol P, Cougnon M
American journal of physiology. Renal physiology 2007 Nov;293(5):F1450-60
American journal of physiology. Renal physiology 2007 Nov;293(5):F1450-60
Coxsackievirus and adenovirus receptor is up-regulated in migratory germ cells during passage of the blood-testis barrier.
Mirza M, Petersen C, Nordqvist K, Sollerbrant K
Endocrinology 2007 Nov;148(11):5459-69
Endocrinology 2007 Nov;148(11):5459-69
Medullary thymic epithelial cells expressing Aire represent a unique lineage derived from cells expressing claudin.
Hamazaki Y, Fujita H, Kobayashi T, Choi Y, Scott HS, Matsumoto M, Minato N
Nature immunology 2007 Mar;8(3):304-11
Nature immunology 2007 Mar;8(3):304-11
Differences in claudin synthesis in primary cultures of acinar cells from rat salivary gland are correlated with the specific three-dimensional organization of the cells.
Qi B, Fujita-Yoshigaki J, Michikawa H, Satoh K, Katsumata O, Sugiya H
Cell and tissue research 2007 Jul;329(1):59-70
Cell and tissue research 2007 Jul;329(1):59-70
Tight and adherens junctions in the ovine uterus: differential regulation by pregnancy and progesterone.
Satterfield MC, Dunlap KA, Hayashi K, Burghardt RC, Spencer TE, Bazer FW
Endocrinology 2007 Aug;148(8):3922-31
Endocrinology 2007 Aug;148(8):3922-31
Loss of claudins-1 and -7 and expression of claudins-3 and -4 correlate with prognostic variables in prostatic adenocarcinomas.
Sheehan GM, Kallakury BV, Sheehan CE, Fisher HA, Kaufman RP Jr, Ross JS
Human pathology 2007 Apr;38(4):564-9
Human pathology 2007 Apr;38(4):564-9
Claudin 1 differentiates endometrioid and serous papillary endometrial adenocarcinoma.
Sobel G, Németh J, Kiss A, Lotz G, Szabó I, Udvarhelyi N, Schaff Z, Páska C
Gynecologic oncology 2006 Nov;103(2):591-8
Gynecologic oncology 2006 Nov;103(2):591-8
Endothelia of term human placentae display diminished expression of tight junction proteins during preeclampsia.
Liévano S, Alarcón L, Chávez-Munguía B, González-Mariscal L
Cell and tissue research 2006 Jun;324(3):433-48
Cell and tissue research 2006 Jun;324(3):433-48
Tight junction proteins and perineurial cells in neurofibromas.
Pummi KP, Aho HJ, Laato MK, Peltonen JT, Peltonen SA
The journal of histochemistry and cytochemistry : official journal of the Histochemistry Society 2006 Jan;54(1):53-61
The journal of histochemistry and cytochemistry : official journal of the Histochemistry Society 2006 Jan;54(1):53-61
Zinc protects renal function during cadmium intoxication in the rat.
Jacquillet G, Barbier O, Cougnon M, Tauc M, Namorado MC, Martin D, Reyes JL, Poujeol P
American journal of physiology. Renal physiology 2006 Jan;290(1):F127-37
American journal of physiology. Renal physiology 2006 Jan;290(1):F127-37
Cingulin regulates claudin-2 expression and cell proliferation through the small GTPase RhoA.
Guillemot L, Citi S
Molecular biology of the cell 2006 Aug;17(8):3569-77
Molecular biology of the cell 2006 Aug;17(8):3569-77
Changes of cell adhesion and extracellular matrix (ECM) components in cervical intraepithelial neoplasia.
Sobel G, Szabó I, Páska C, Kiss A, Kovalszky I, Kádár A, Paulin F, Schaff Z
Pathology oncology research : POR 2005;11(1):26-31
Pathology oncology research : POR 2005;11(1):26-31
Inflammatory processes have differential effects on claudins 2, 3 and 4 in colonic epithelial cells.
Prasad S, Mingrino R, Kaukinen K, Hayes KL, Powell RM, MacDonald TT, Collins JE
Laboratory investigation; a journal of technical methods and pathology 2005 Sep;85(9):1139-62
Laboratory investigation; a journal of technical methods and pathology 2005 Sep;85(9):1139-62
Inducible expression of Snail selectively increases paracellular ion permeability and differentially modulates tight junction proteins.
Carrozzino F, Soulié P, Huber D, Mensi N, Orci L, Cano A, Féraille E, Montesano R
American journal of physiology. Cell physiology 2005 Oct;289(4):C1002-14
American journal of physiology. Cell physiology 2005 Oct;289(4):C1002-14
Polyamines are necessary for synthesis and stability of occludin protein in intestinal epithelial cells.
Guo X, Rao JN, Liu L, Zou T, Keledjian KM, Boneva D, Marasa BS, Wang JY
American journal of physiology. Gastrointestinal and liver physiology 2005 Jun;288(6):G1159-69
American journal of physiology. Gastrointestinal and liver physiology 2005 Jun;288(6):G1159-69
Knockdown of occludin expression leads to diverse phenotypic alterations in epithelial cells.
Yu AS, McCarthy KM, Francis SA, McCormack JM, Lai J, Rogers RA, Lynch RD, Schneeberger EE
American journal of physiology. Cell physiology 2005 Jun;288(6):C1231-41
American journal of physiology. Cell physiology 2005 Jun;288(6):C1231-41
Leukocyte diapedesis in vivo induces transient loss of tight junction protein at the blood-retina barrier.
Xu H, Dawson R, Crane IJ, Liversidge J
Investigative ophthalmology & visual science 2005 Jul;46(7):2487-94
Investigative ophthalmology & visual science 2005 Jul;46(7):2487-94
Phosphorylation of claudin-3 at threonine 192 by cAMP-dependent protein kinase regulates tight junction barrier function in ovarian cancer cells.
D'Souza T, Agarwal R, Morin PJ
The Journal of biological chemistry 2005 Jul 15;280(28):26233-40
The Journal of biological chemistry 2005 Jul 15;280(28):26233-40
Extracellular signal-regulated kinases 1/2 control claudin-2 expression in Madin-Darby canine kidney strain I and II cells.
Lipschutz JH, Li S, Arisco A, Balkovetz DF
The Journal of biological chemistry 2005 Feb 4;280(5):3780-8
The Journal of biological chemistry 2005 Feb 4;280(5):3780-8
Chronic inflammatory pain leads to increased blood-brain barrier permeability and tight junction protein alterations.
Brooks TA, Hawkins BT, Huber JD, Egleton RD, Davis TP
American journal of physiology. Heart and circulatory physiology 2005 Aug;289(2):H738-43
American journal of physiology. Heart and circulatory physiology 2005 Aug;289(2):H738-43
Claudin-3 and claudin-4 expression in ovarian epithelial cells enhances invasion and is associated with increased matrix metalloproteinase-2 activity.
Agarwal R, D'Souza T, Morin PJ
Cancer research 2005 Aug 15;65(16):7378-85
Cancer research 2005 Aug 15;65(16):7378-85
Cultured monolayers of the dog jejunum with the structural and functional properties resembling the normal epithelium.
Weng XH, Beyenbach KW, Quaroni A
American journal of physiology. Gastrointestinal and liver physiology 2005 Apr;288(4):G705-17
American journal of physiology. Gastrointestinal and liver physiology 2005 Apr;288(4):G705-17
Paracellular Cl- permeability is regulated by WNK4 kinase: insight into normal physiology and hypertension.
Kahle KT, Macgregor GG, Wilson FH, Van Hoek AN, Brown D, Ardito T, Kashgarian M, Giebisch G, Hebert SC, Boulpaep EL, Lifton RP
Proceedings of the National Academy of Sciences of the United States of America 2004 Oct 12;101(41):14877-82
Proceedings of the National Academy of Sciences of the United States of America 2004 Oct 12;101(41):14877-82
The rotavirus surface protein VP8 modulates the gate and fence function of tight junctions in epithelial cells.
Nava P, López S, Arias CF, Islas S, González-Mariscal L
Journal of cell science 2004 Nov 1;117(Pt 23):5509-19
Journal of cell science 2004 Nov 1;117(Pt 23):5509-19
Epidermal growth factor receptor activation differentially regulates claudin expression and enhances transepithelial resistance in Madin-Darby canine kidney cells.
Singh AB, Harris RC
The Journal of biological chemistry 2004 Jan 30;279(5):3543-52
The Journal of biological chemistry 2004 Jan 30;279(5):3543-52
Mechanisms of diarrhea in the interleukin-2-deficient mouse model of colonic inflammation.
Barmeyer C, Harren M, Schmitz H, Heinzel-Pleines U, Mankertz J, Seidler U, Horak I, Wiedenmann B, Fromm M, Schulzke JD
American journal of physiology. Gastrointestinal and liver physiology 2004 Feb;286(2):G244-52
American journal of physiology. Gastrointestinal and liver physiology 2004 Feb;286(2):G244-52
Expression and function of tight junctions in the crypt epithelium of human palatine tonsils.
Go M, Kojima T, Takano K, Murata M, Ichimiya S, Tsubota H, Himi T, Sawada N
The journal of histochemistry and cytochemistry : official journal of the Histochemistry Society 2004 Dec;52(12):1627-38
The journal of histochemistry and cytochemistry : official journal of the Histochemistry Society 2004 Dec;52(12):1627-38
Tight junction proteins ZO-1, occludin, and claudins in developing and adult human perineurium.
Pummi KP, Heape AM, Grénman RA, Peltonen JT, Peltonen SA
The journal of histochemistry and cytochemistry : official journal of the Histochemistry Society 2004 Aug;52(8):1037-46
The journal of histochemistry and cytochemistry : official journal of the Histochemistry Society 2004 Aug;52(8):1037-46
Role of claudin interactions in airway tight junctional permeability.
Coyne CB, Gambling TM, Boucher RC, Carson JL, Johnson LG
American journal of physiology. Lung cellular and molecular physiology 2003 Nov;285(5):L1166-78
American journal of physiology. Lung cellular and molecular physiology 2003 Nov;285(5):L1166-78
Claudin-8 expression in Madin-Darby canine kidney cells augments the paracellular barrier to cation permeation.
Yu AS, Enck AH, Lencer WI, Schneeberger EE
The Journal of biological chemistry 2003 May 9;278(19):17350-9
The Journal of biological chemistry 2003 May 9;278(19):17350-9
Localization of claudin-3 in tight junctions of the blood-brain barrier is selectively lost during experimental autoimmune encephalomyelitis and human glioblastoma multiforme.
Wolburg H, Wolburg-Buchholz K, Kraus J, Rascher-Eggstein G, Liebner S, Hamm S, Duffner F, Grote EH, Risau W, Engelhardt B
Acta neuropathologica 2003 Jun;105(6):586-92
Acta neuropathologica 2003 Jun;105(6):586-92
Tight junction proteins claudin-3 and claudin-4 are frequently overexpressed in ovarian cancer but not in ovarian cystadenomas.
Rangel LB, Agarwal R, D'Souza T, Pizer ES, Alò PL, Lancaster WD, Gregoire L, Schwartz DR, Cho KR, Morin PJ
Clinical cancer research : an official journal of the American Association for Cancer Research 2003 Jul;9(7):2567-75
Clinical cancer research : an official journal of the American Association for Cancer Research 2003 Jul;9(7):2567-75
Effects of hypoxia-reoxygenation on rat blood-brain barrier permeability and tight junctional protein expression.
Witt KA, Mark KS, Hom S, Davis TP
American journal of physiology. Heart and circulatory physiology 2003 Dec;285(6):H2820-31
American journal of physiology. Heart and circulatory physiology 2003 Dec;285(6):H2820-31
Bryostatin-1 enhances barrier function in T84 epithelia through PKC-dependent regulation of tight junction proteins.
Yoo J, Nichols A, Mammen J, Calvo I, Song JC, Worrell RT, Matlin K, Matthews JB
American journal of physiology. Cell physiology 2003 Aug;285(2):C300-9
American journal of physiology. Cell physiology 2003 Aug;285(2):C300-9
Claudin-2 expression induces cation-selective channels in tight junctions of epithelial cells.
Amasheh S, Meiri N, Gitter AH, Schöneberg T, Mankertz J, Schulzke JD, Fromm M
Journal of cell science 2002 Dec 15;115(Pt 24):4969-76
Journal of cell science 2002 Dec 15;115(Pt 24):4969-76
Claudin-2 expression induces cation-selective channels in tight junctions of epithelial cells.
Amasheh S, Meiri N, Gitter AH, Schöneberg T, Mankertz J, Schulzke JD, Fromm M
Journal of cell science 2002 Dec 15;115(Pt 24):4969-76
Journal of cell science 2002 Dec 15;115(Pt 24):4969-76
Mechanisms of diarrhea in collagenous colitis.
Bürgel N, Bojarski C, Mankertz J, Zeitz M, Fromm M, Schulzke JD
Gastroenterology 2002 Aug;123(2):433-43
Gastroenterology 2002 Aug;123(2):433-43
Mechanisms of diarrhea in collagenous colitis.
Bürgel N, Bojarski C, Mankertz J, Zeitz M, Fromm M, Schulzke JD
Gastroenterology 2002 Aug;123(2):433-43
Gastroenterology 2002 Aug;123(2):433-43
No comments: Submit comment
Supportive validation
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
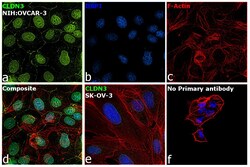
- Experimental details
- Immunofluorescence analysis of Claudin-3 was performed using 70% confluent log phase NIH:OVCAR-3 cells. The cells were fixed with 4% paraformaldehyde for 5 minutes, permeabilized with 0.1% Triton™ X-100 for 10 minutes, and blocked with 2% BSA for 45 minutes at room temperature. The cells were labeled with Claudin 3 Polyclonal Antibody (Product # 34-1700) at 1:100 dilution in 0.1% BSA, incubated at 4 degree celsius overnight and then labeled with Donkey anti-Rabbit IgG (H+L) Highly Cross-Adsorbed Secondary Antibody, Alexa Fluor Plus 488 (Product # A32790), (1:2000 dilution), for 45 minutes at room temperature (Panel a: Green). Nuclei (Panel b:Blue) were stained with ProLong™ Diamond Antifade Mountant with DAPI (Product # P36962). F-actin (Panel c: Red) was stained with Rhodamine Phalloidin (Product # R415, 1:300). Panel d represents the merged image showing cell junctions and nucleus localization. Panel e represents low expressing SK-OV-3 cells. Panel f represents control cells with no primary antibody to assess background. The images were captured at 60X magnification.
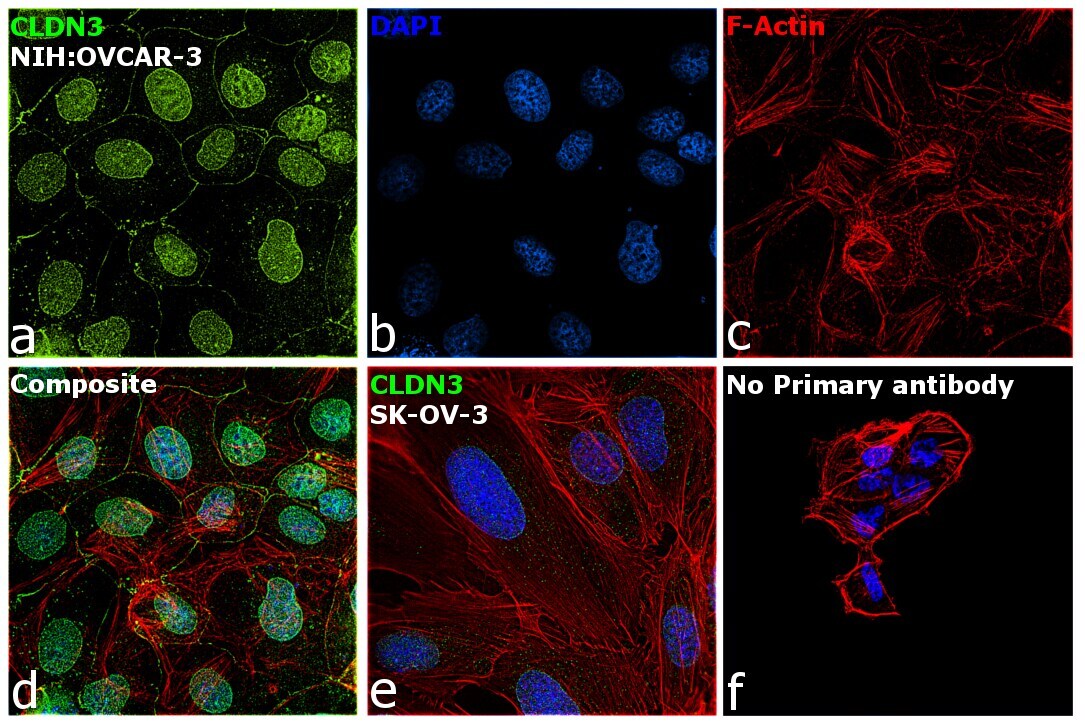
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- Immunofluorescence analysis of Claudin-3 was performed using 70% confluent log phase NIH:OVCAR-3 cells. The cells were fixed with 4% paraformaldehyde for 5 minutes, permeabilized with 0.1% Triton™ X-100 for 10 minutes, and blocked with 2% BSA for 45 minutes at room temperature. The cells were labeled with Claudin 3 Polyclonal Antibody (Product # 34-1700) at 1:100 dilution in 0.1% BSA, incubated at 4 degree celsius overnight and then labeled with Donkey anti-Rabbit IgG (H+L) Highly Cross-Adsorbed Secondary Antibody, Alexa Fluor Plus 488 (Product # A32790), (1:2000 dilution), for 45 minutes at room temperature (Panel a: Green). Nuclei (Panel b:Blue) were stained with ProLong™ Diamond Antifade Mountant with DAPI (Product # P36962). F-actin (Panel c: Red) was stained with Rhodamine Phalloidin (Product # R415, 1:300). Panel d represents the merged image showing cell junctions and nucleus localization. Panel e represents low expressing SK-OV-3 cells. Panel f represents control cells with no primary antibody to assess background. The images were captured at 60X magnification.
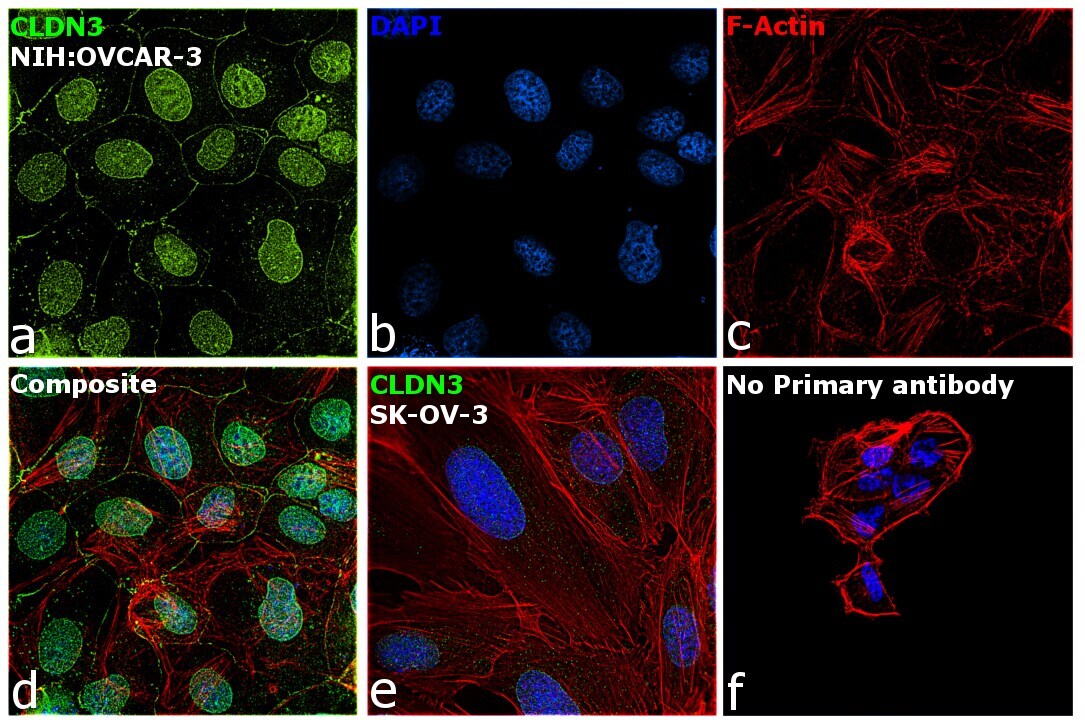
Supportive validation
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
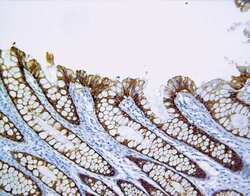
- Experimental details
- Immunohistochemical staining of human colon tissue using Rabbit anti-Claudin-3 polyclonal antibody (PAD: Z23.JM). (Product # 34-1700).
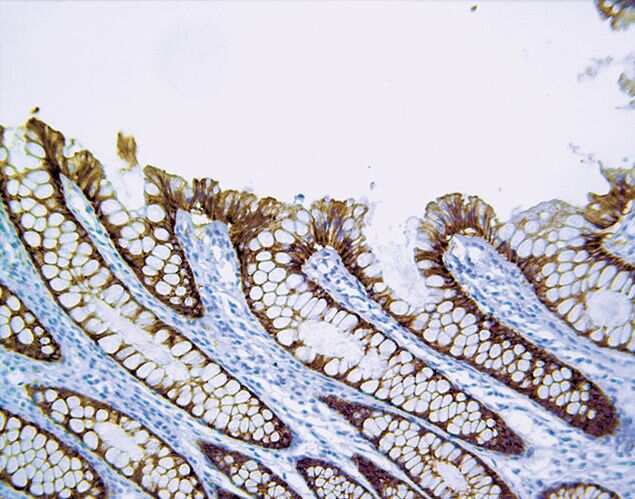
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
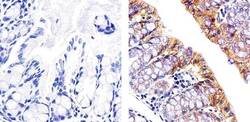
- Experimental details
- Immunohistochemistry analysis of Claudin-3 showing staining in the membrane of paraffin-embedded mouse colon tissue (right) compared to a negative control without primary antibody (left). To expose target proteins, antigen retrieval was performed using 10mM sodium citrate (pH 6.0), microwaved for 8-15 min. Following antigen retrieval, tissues were blocked in 3% H2O2-methanol for 15 min at room temperature, washed with ddH2O and PBS, and then probed with a Claudin-3 polyclonal antibody (Product # 34-1700) diluted in 3% BSA-PBS at a dilution of 1:100 overnight at 4°C in a humidified chamber. Tissues were washed extensively in PBST and detection was performed using an HRP-conjugated secondary antibody followed by colorimetric detection using a DAB kit. Tissues were counterstained with hematoxylin and dehydrated with ethanol and xylene to prep for mounting.
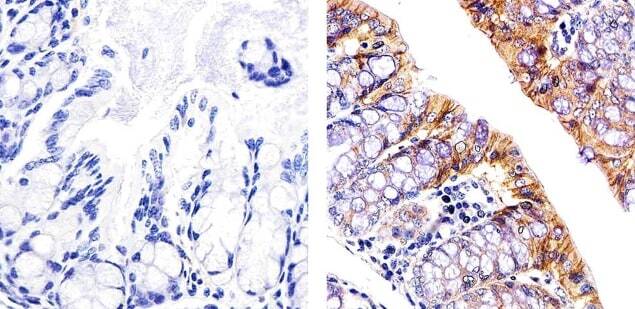
Supportive validation
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
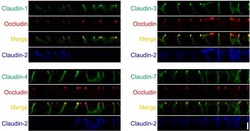
- Experimental details
- Fig 5 Localization of claudins in z-axis plane in control and claudin-2 knockout cells. Immunofluorescence analysis of claudins and occludin in co-culture of control MDCK II cells and claudin-2 knockout clone 1 (KO 1) in z-axis plane. The signals of claudin-1, -3, -4 and -7 were stronger at TJs in claudin-2 knockout cells compared with control cells, and lateral localization of these claudins was similar between control and claudin-2 knockout cells. Scale bar = 5 mum.
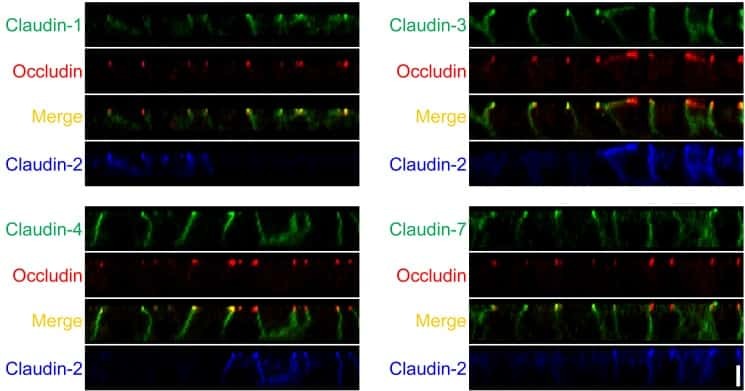
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
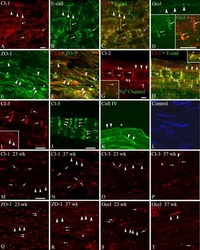
- Experimental details
- NULL
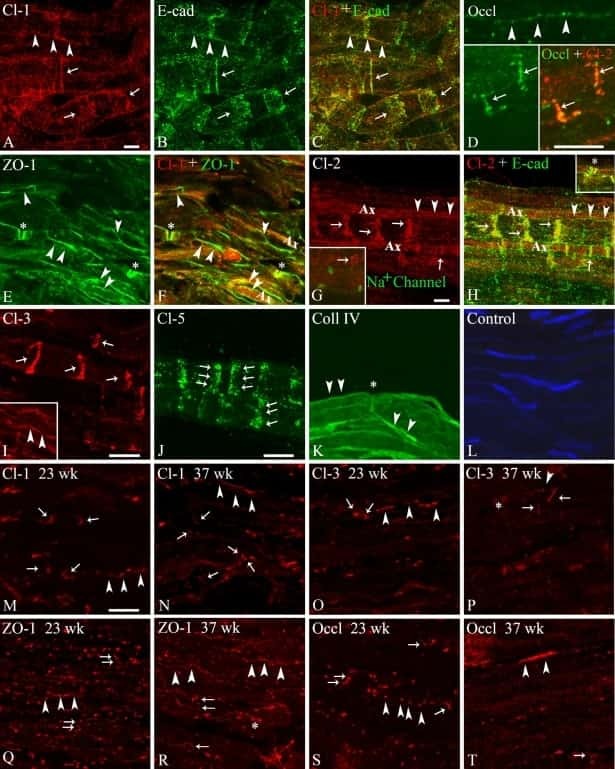
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
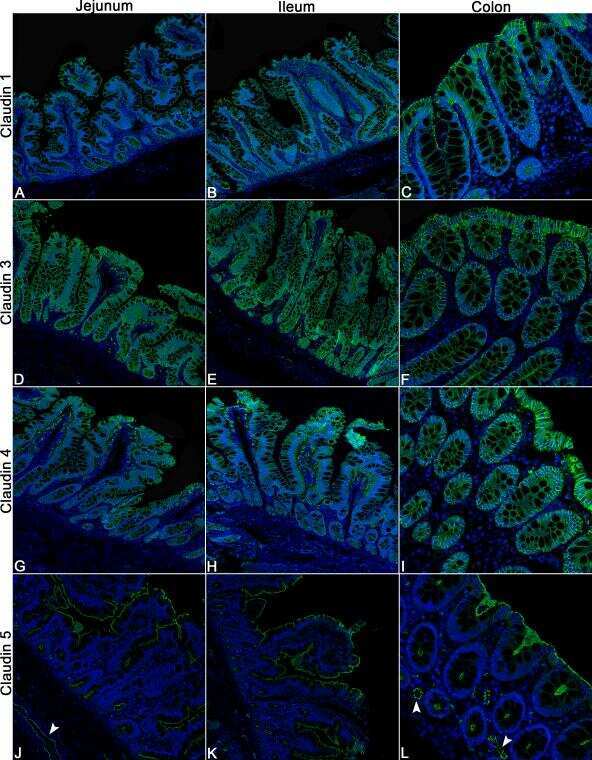
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
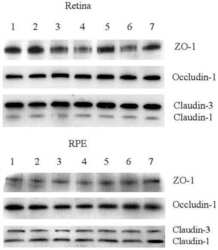
- Experimental details
- NULL
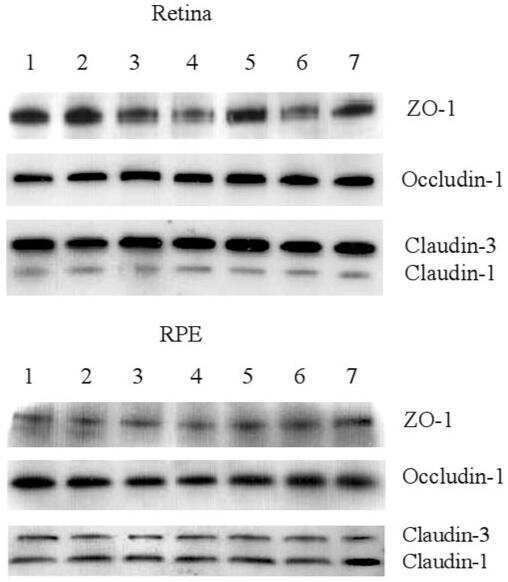
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
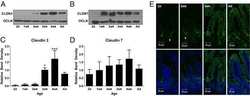
- Experimental details
- NULL
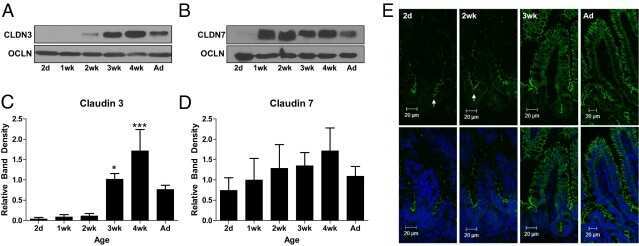
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
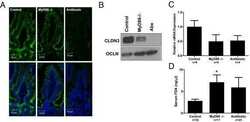
- Experimental details
- NULL
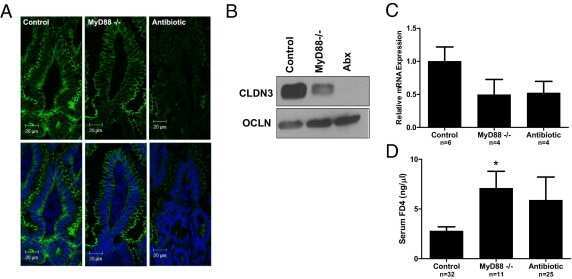
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
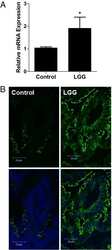
- Experimental details
- NULL
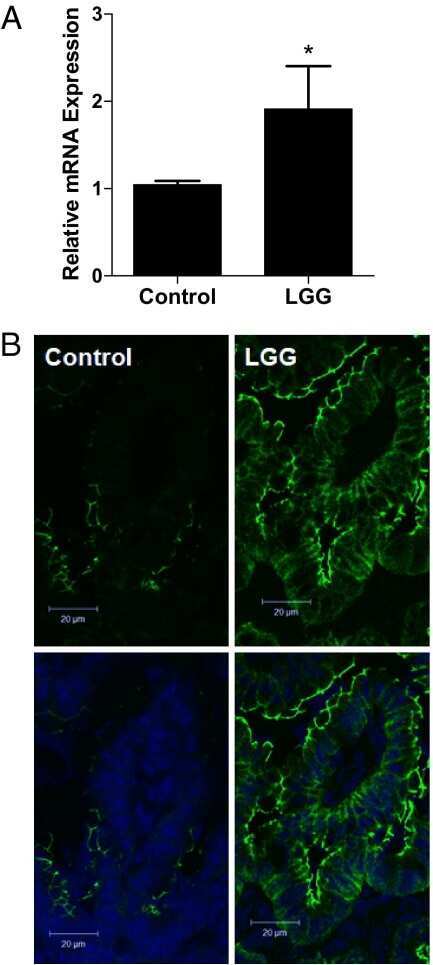
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
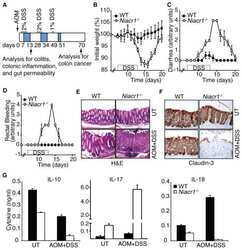
- Experimental details
- NULL
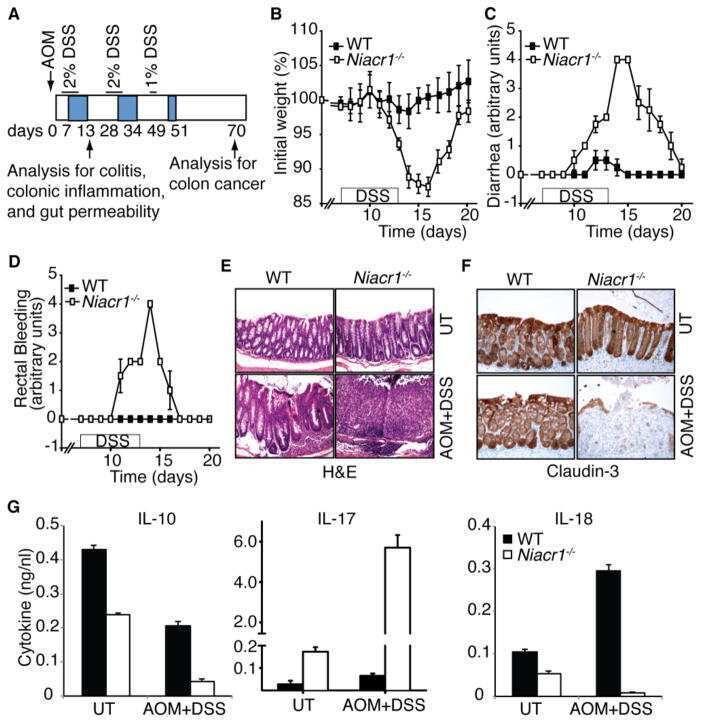
- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
- NULL

- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
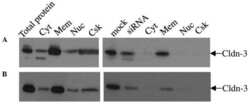
- Experimental details
- NULL
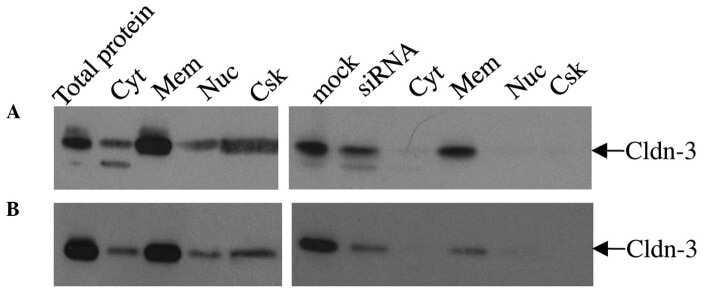
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
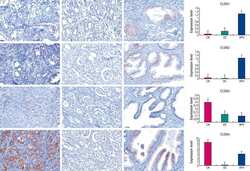
- Experimental details
- NULL
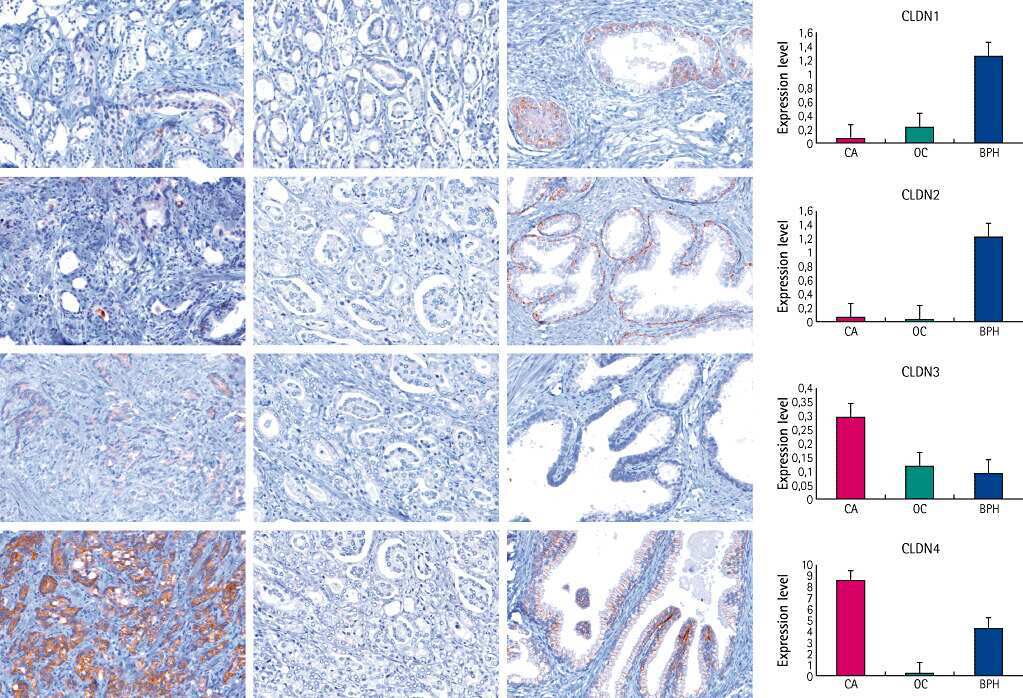
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
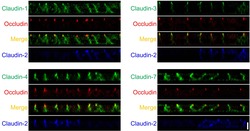
- Experimental details
- NULL
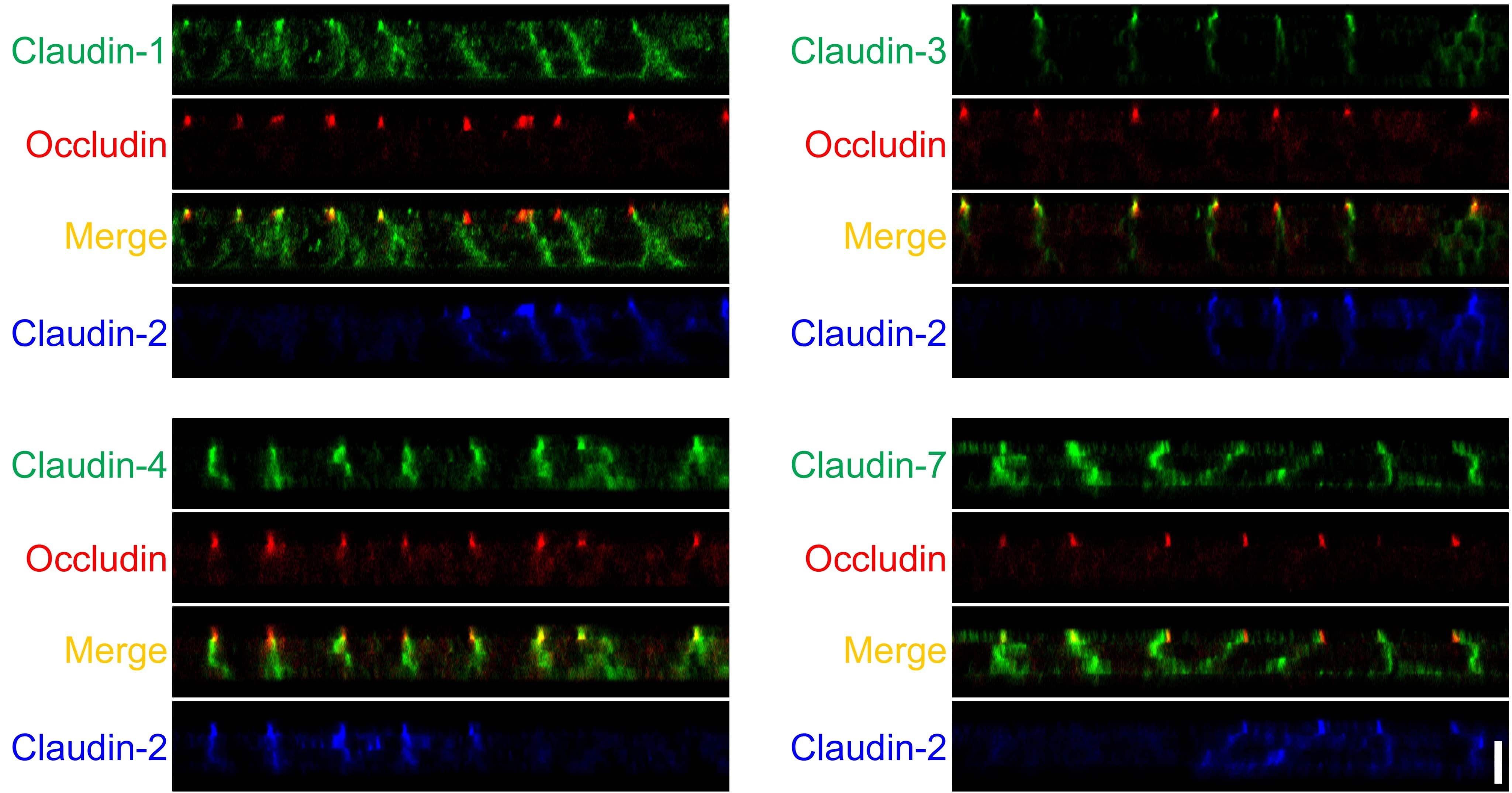
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
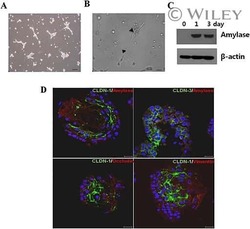
- Experimental details
- NULL
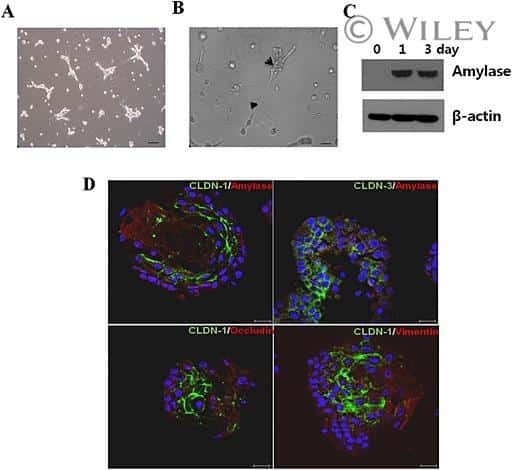
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
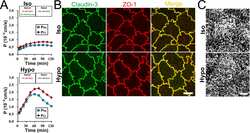
- Experimental details
- NULL
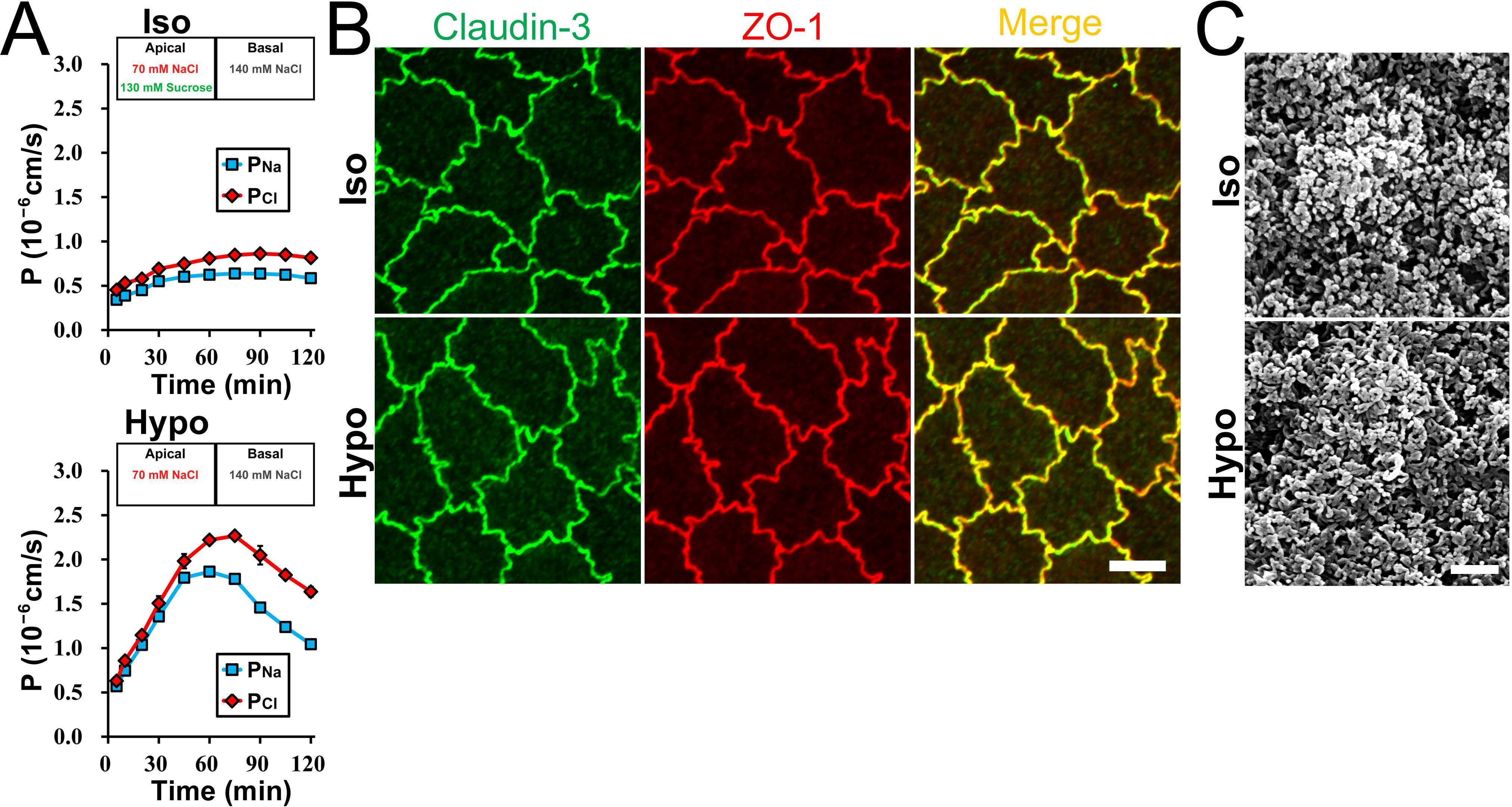
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
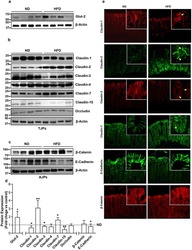
- Experimental details
- NULL
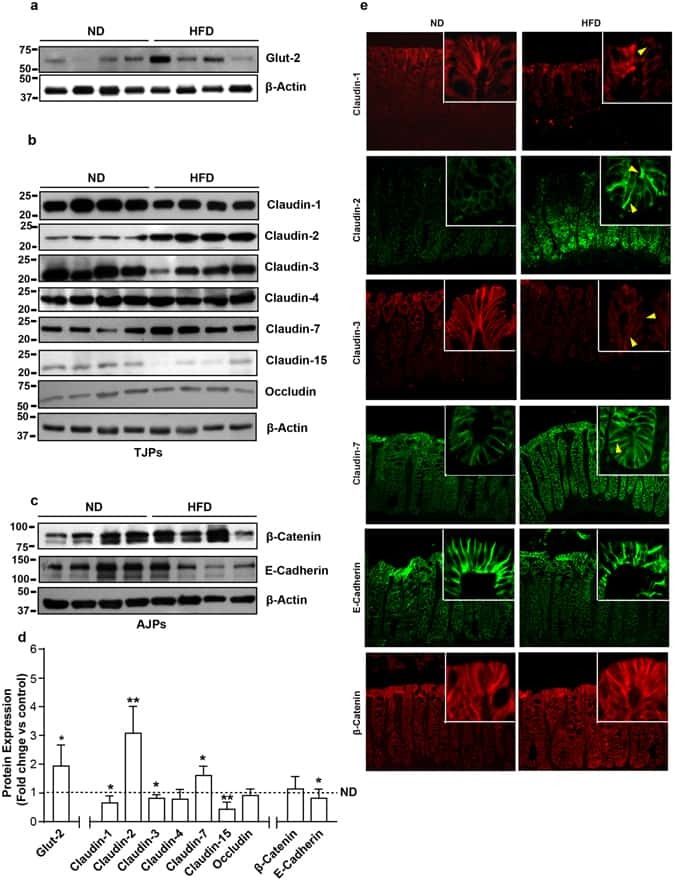
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
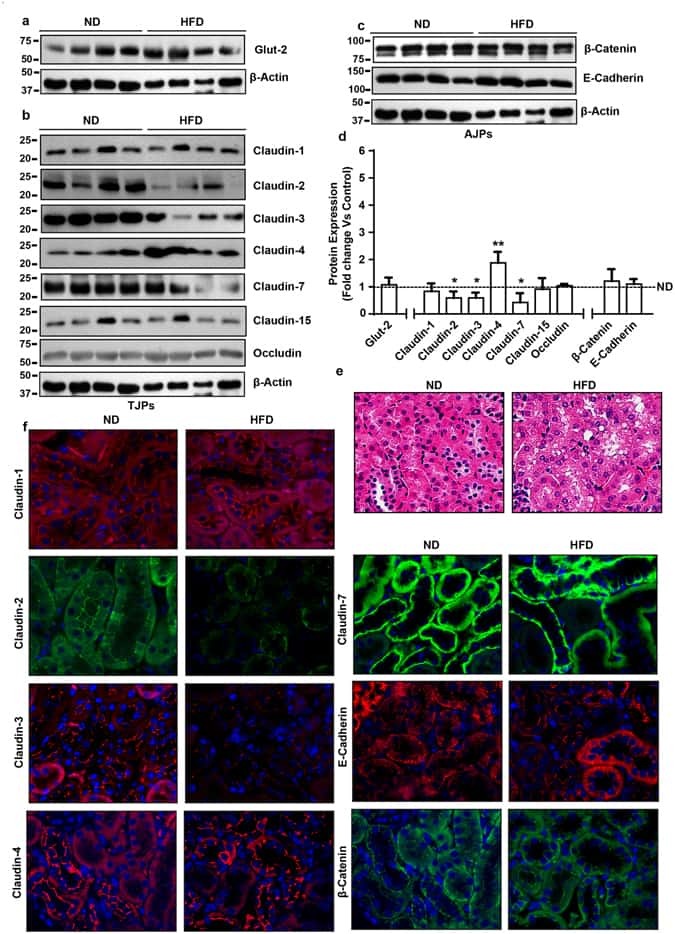
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
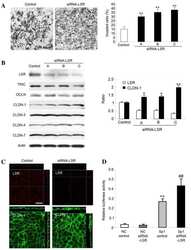
- Experimental details
- NULL
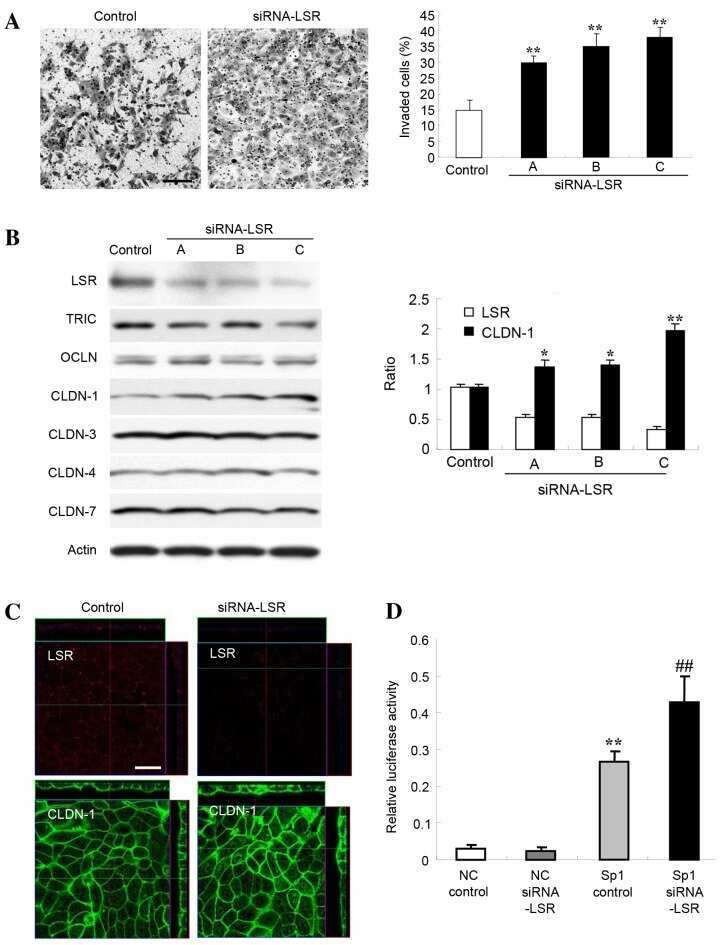
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
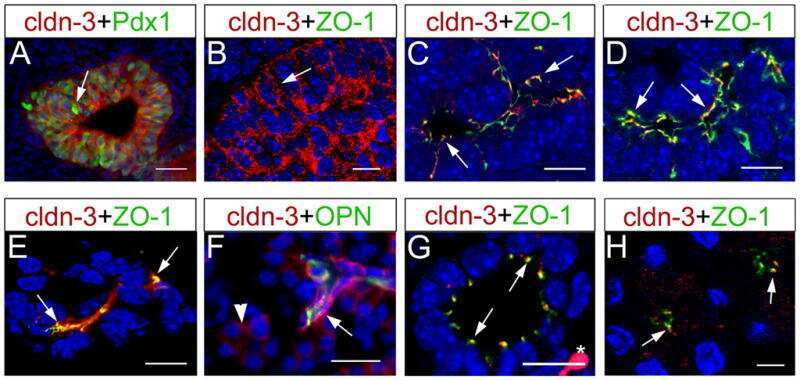
- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
- NULL
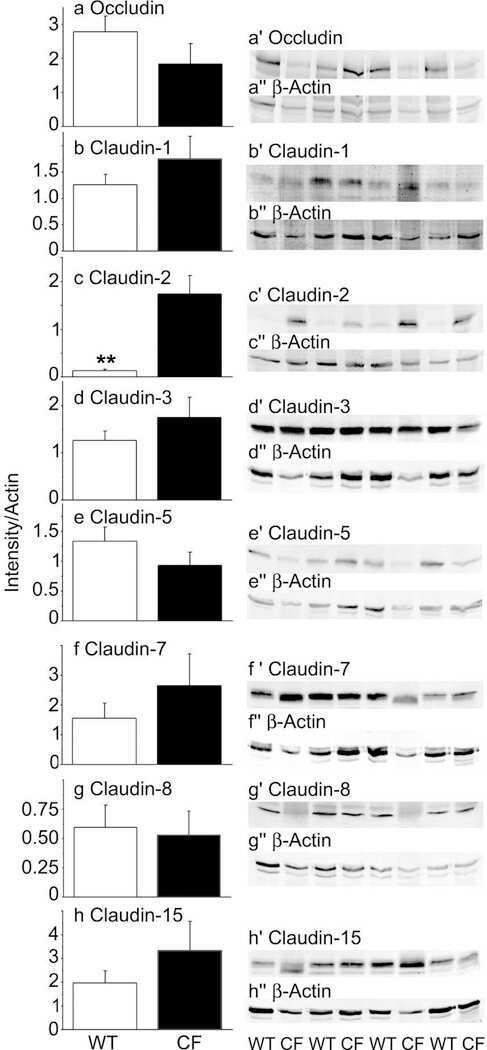
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
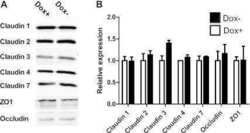
- Experimental details
- NULL
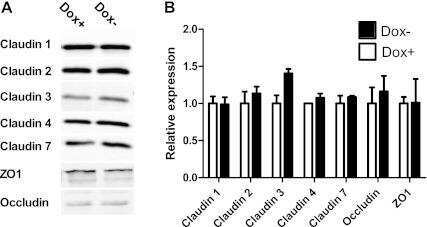
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
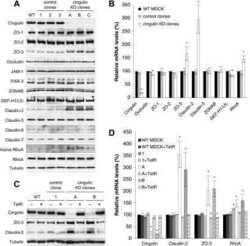
- Experimental details
- NULL
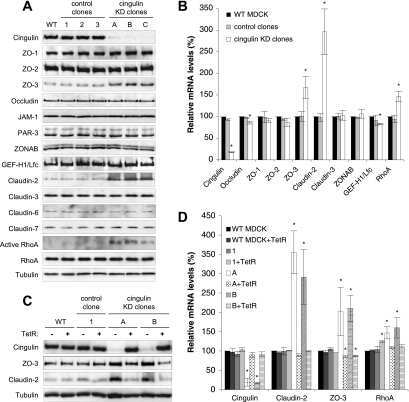
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
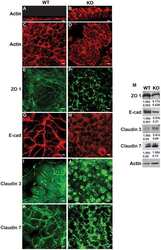
- Experimental details
- NULL
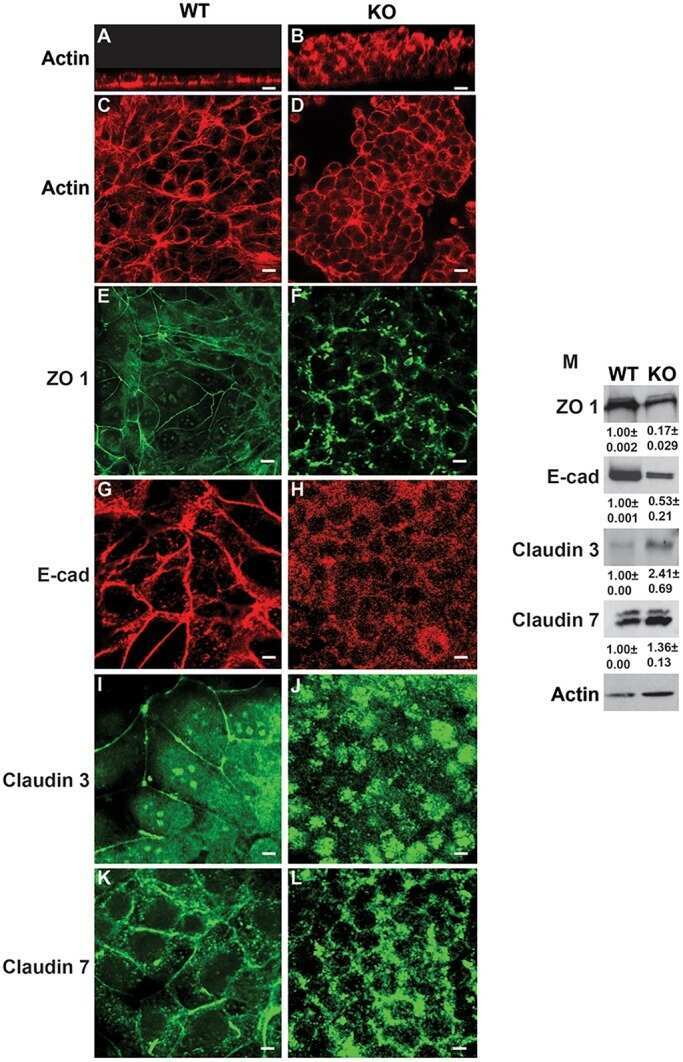
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
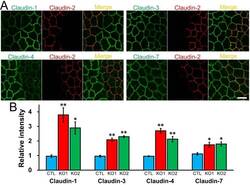
- Experimental details
- Fig 4 Effects of claudin-2 knockout on the localization of other claudins. (A) Immunofluorescence analysis of claudins in co-culture of control MDCK II cells and claudin-2 knockout clone 1 (KO 1). Claudin-1, -3, -4, and -7 showed clearer and stronger signals at TJs in claudin-2 knockout cells than in control cells. Scale bar = 10 mum. (B) Quantification analysis of signal intensity of claudins at TJs in control MDCK II cells and claudin-2 knockout clones. The signal intensity of claudins at TJs in control cells and claudin-2 knockout clones was measured as described in Materials and Methods , and the relative signal intensity of each claudin was calculated as the ratio of the signal intensity in control cells (CTL) and claudin-2 knockout clones (KO 1 and 2) to the signal intensity in control cells. N = 4 for each experiment. * p < 0.05, ** p < 0.01 compared with control.
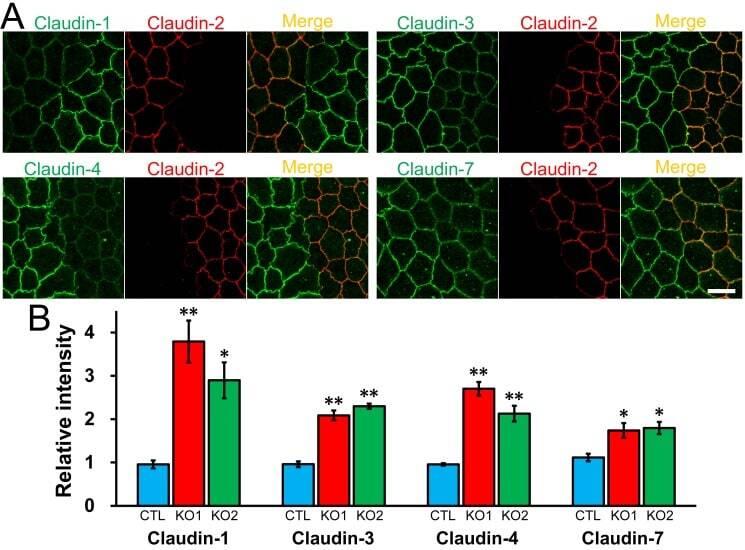
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- Fig 5 Localization of claudins in z-axis plane in control and claudin-2 knockout cells. Immunofluorescence analysis of claudins and occludin in co-culture of control MDCK II cells and claudin-2 knockout clone 1 (KO 1) in z-axis plane. The signals of claudin-1, -3, -4 and -7 were stronger at TJs in claudin-2 knockout cells compared with control cells, and lateral localization of these claudins was similar between control and claudin-2 knockout cells. Scale bar = 5 mum.
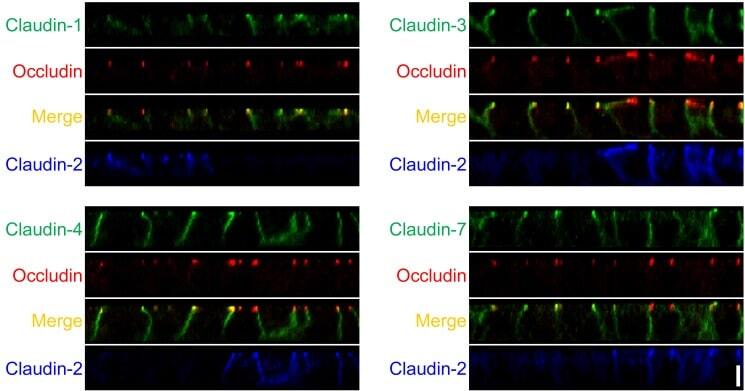
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
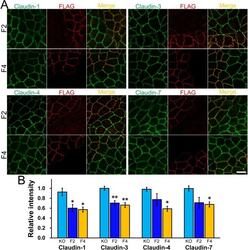
- Experimental details
- Fig 8 Effects of claudin-2 re-expression on the localization of other claudins in claudin-2 knockout cells. (A) Immunofluorescence analysis of claudins and FLAG in co-culture of claudin-2 knockout cells and F2 or F4 clones. Claudin-1, -3, -4, and -7 staining at TJs in F2 and F4 clones was weaker than those in claudin-2 knockout cells. Scale bar = 10 mum. (B) Quantification analysis of the signal intensity of claudins at TJs in F2 and F4 clones. The signal intensity of claudin-1, -3, -4, and -7 at TJs in F2 and F4 clones was compared with that in claudin-2 knockout cells. N = 4 for each experiment. * p < 0.05, ** p < 0.01 compared with claudin-2 knockout cells.
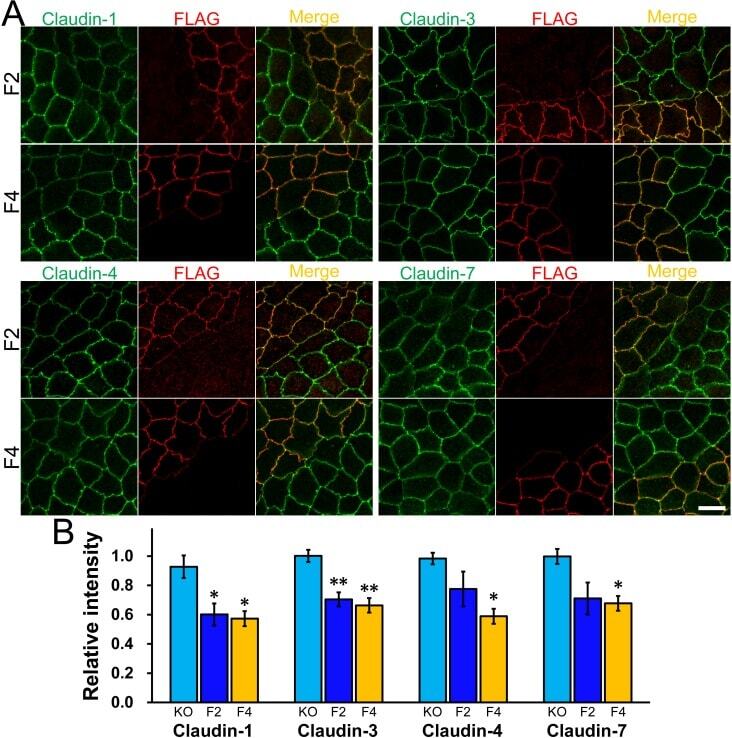
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
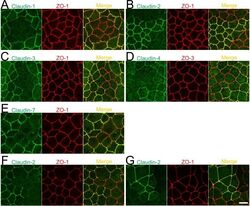
- Experimental details
- Fig 11 Localization of claudins at TJs between control and their respective knockout cells. (A-E) Immunofluorescence analysis of claudins and ZO-1 or ZO-3 in MDCK II cells transfected with the TALEN constructs for claudin-1, -2, -3, -4, or-7 gene knockouts. After transfection, cells were subcultured on filter inserts for 4 days before the analysis of claudin-2 and -3 (B and C), and for 2 days before the analysis of claudin-1, -4, and -7 (A, D, and E). (F) Immunofluorescence analysis of claudin-2 and ZO-1. TALEN constructs for the claudin-2 gene knockout were transfected into MDCK II cells, which were subcultured on filter inserts for 2 days before analysis. (G) Immunofluorescence analysis of claudin-2 and ZO-1 in a co-culture of wild-type and claudin-2 expressing MDCK I cells. Cells were cultured on filter inserts for 4 days before analysis. Scale bar = 10 mum.
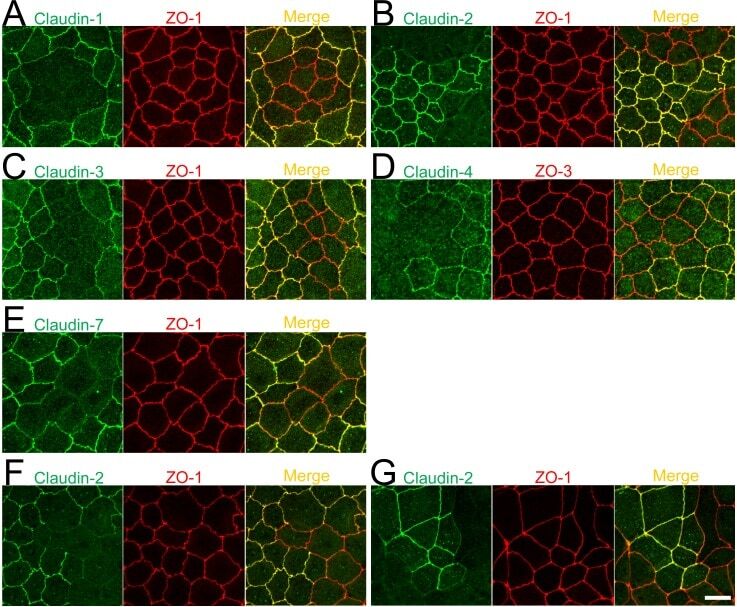
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
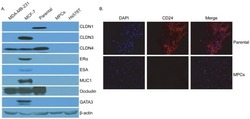
- Experimental details
- Figure 4 Western blot and immunofluorescence analysis of parental bipotent cells and MPCs using various markers from the signature of claudin-low breast cancer subtype. ( A ) Western blot analysis of cell lysates from exponentially proliferating bipotent cells and MPCs were analyzed for expression of indicated markers using specific antibodies. Breast cancer cell lines MDA-MB-231, MCF-7, and Hs578T were used as controls. ( B ) Immunofluorescence staining using CD24 antibody (red), nuclei (blue) represent DAPI staining.
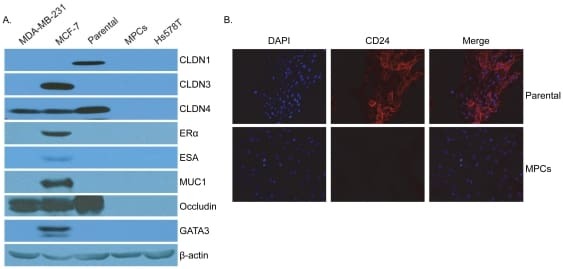
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
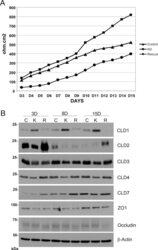
- Experimental details
- FIGURE 6: Rab25 depletion alters the transepithelial resistance and claudin expression. (A) TER was measured in all cell lines from day 3 to day 15 on Transwell filters. During this period, the media were changed every day, and TER was measured in 24-well Transwell inserts. Results are expressed in ohm-cm 2 and are representative of three separate experiments. (B) Equal amounts of total protein lysates (20 mug) from Control, Rab25KD, and Rescue cells were analyzed by Western blots for tight junction proteins. The blots were subsequently stripped and reprobed for beta-actin as a loading control. The results are representative of three separate experiments.
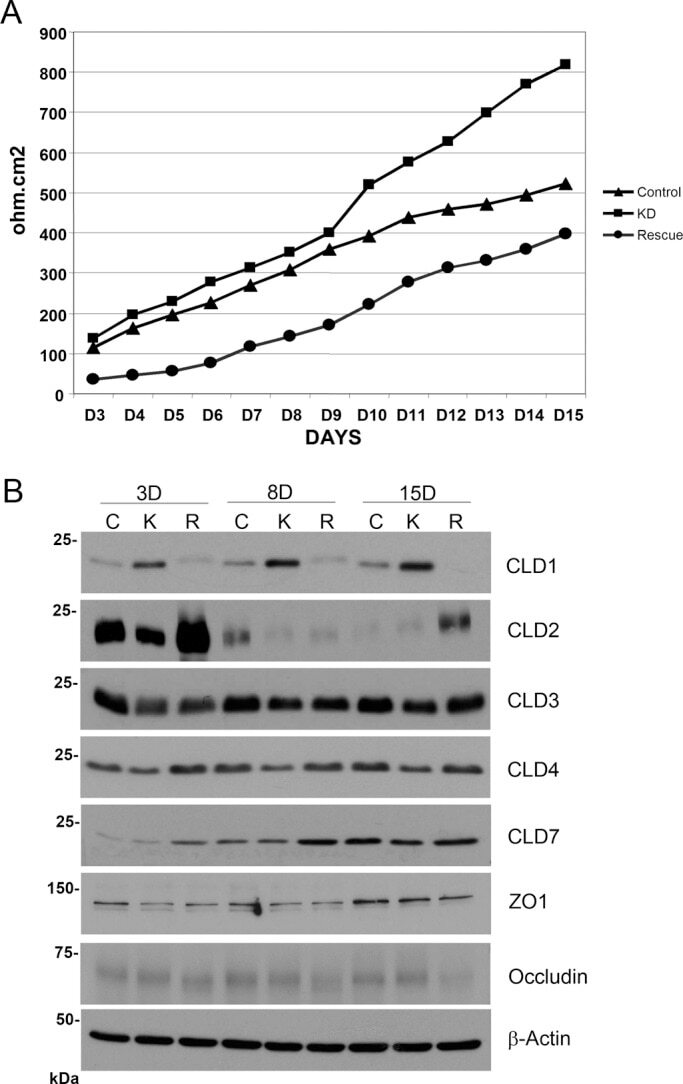
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
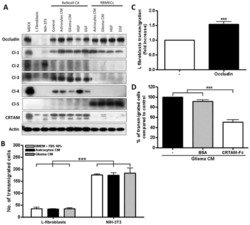
- Experimental details
- Figure 6 Expression of TJ proteins in transmigrating cells favors their passage across RBMECs. A) Analysis by Western blots of the expression of occludin, claudins 1 to 5 and CRTAM in NIH-3T3and L-fibroblasts, ReNcells CX and RBMECs. B) The transmigration of NIH-3T3 and L-fibroblasts acrossRBMECs is independent of the CM present in the basal compartment, but is significantly higher inNIH-3T3 fibroblasts that express claudin-2 than in L-fibroblast that do not express TJ proteins.N = 3, F(1,6) = 232.22, ***P
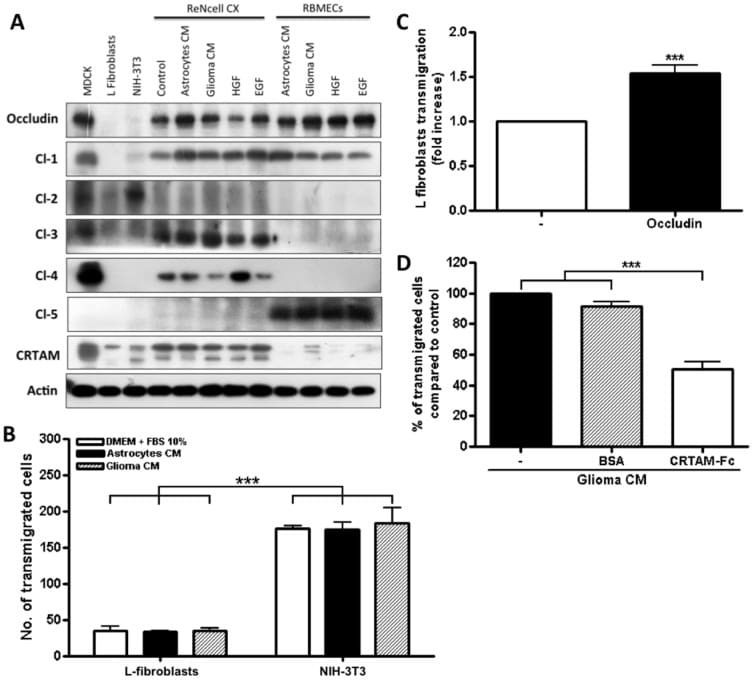
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
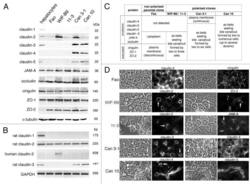
- Experimental details
- Figure 4. Expression and localization of tight junction (TJ) proteins in cell lines expressing typical hepatocyte polarity. Fully polarized cultures were examined. ( A ) western blot. Each lane was loaded with a cell lysate aliquot containing 25 mug total protein. ( B ) RT-PCR. For western blot and RT-PCR, experiments were performed three times with lysates or RNA preparations from two to three independent cultures; one representative experiment is shown. ( C ) Summary of the localization of ten TJ proteins. Experiments were performed at least three times on three independent cultures. ( D ) Images of the localization of some TJ proteins; for each case, the corresponding phase-contrast image is given on the left. Some bile canaliculi (BC) are indicated by arrows (short arrows for BC formed between two cells, long arrows for extended BC). Bar, 10 mum
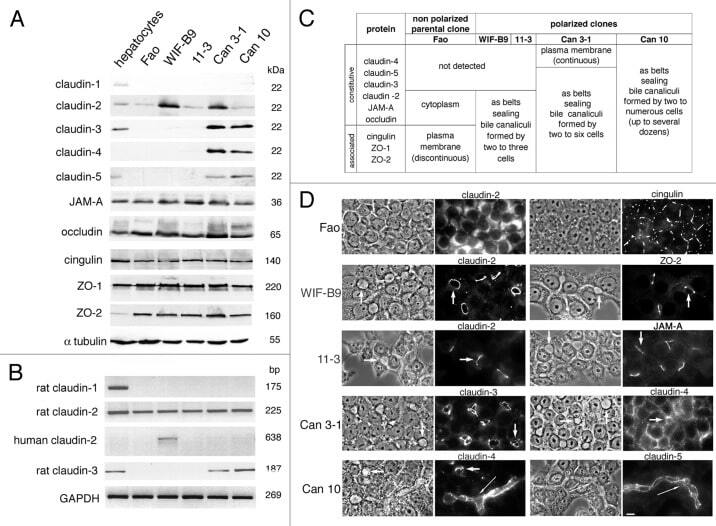
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
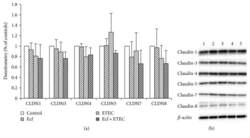
- Experimental details
- Figure 3 Protein expression (Western blot) of claudin-1, claudin-3, claudin-4, claudin-5, claudin-7, and claudin-8 after treatment with bacterial strains (Ecf, ETEC, and Ecf + ETEC). (a) Protein expression relative to respective controls [means +- SEM]. Samples were taken after 8 h. No differences were observed between treatment groups. N = 3 independent experiments per bar. (b) Exemplarily, data of one Western blot is shown. 1 = control (8 h), 2 = Ecf (8 h), 3 = ETEC (8 h), 4 = Ecf (8 h) + ETEC (6 h), and 5 = Ecf (10 h) + ETEC (8 h). Sample 4 was included as a control to rule out effects of longer total bacterial incubation (preincubation) and was not included in the statistical analysis shown in (a).
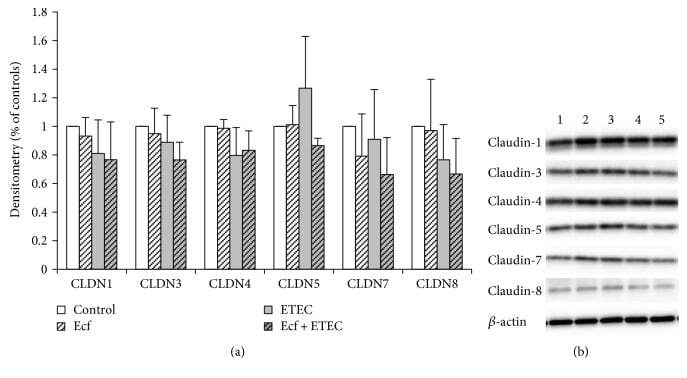
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
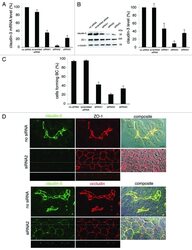
- Experimental details
- Figure 10. Impact of silencing claudin-3 on Can 10 polarity. Cells transfected either by Lipofectamine with no siRNA, scrambled siRNA or three different claudin-3 siRNA were examined 48 h after tranfection. ( A ) RT-qPCR of one representative experiment. Claudin-3 mRNA level is expressed as the percentage of the level of cells treated with no siRNA. ( B ) western blot. An immunoblot corresponding to the experiment shown in A is presented on the left. Densitometric analysis of immunoblots obtained from three independent experiments is on the right. Claudin-3 level, normalized with alpha-tubulin as internal control, was expressed as the percentage of claudin-3 level of cells treated with no siRNA. ( C ) Analysis of cells forming bile canaliculi. Values are means of three independent experiments. ( D ) Co-localization of claudin-3 with ZO-1, and claudin-5 with occludin, studied by confocal microscopy. In each case, the localization of each tight junction protein (compilation of 20-27 xy sections) is shown, followed by a composite image (interferentiel contrast image with the merge image) (right). For cells treated with claudin-3 siRNA2, xz sections taken as indicated (thin white lines) on the corresponding xy compilations are also presented. Bar, 10 mum.
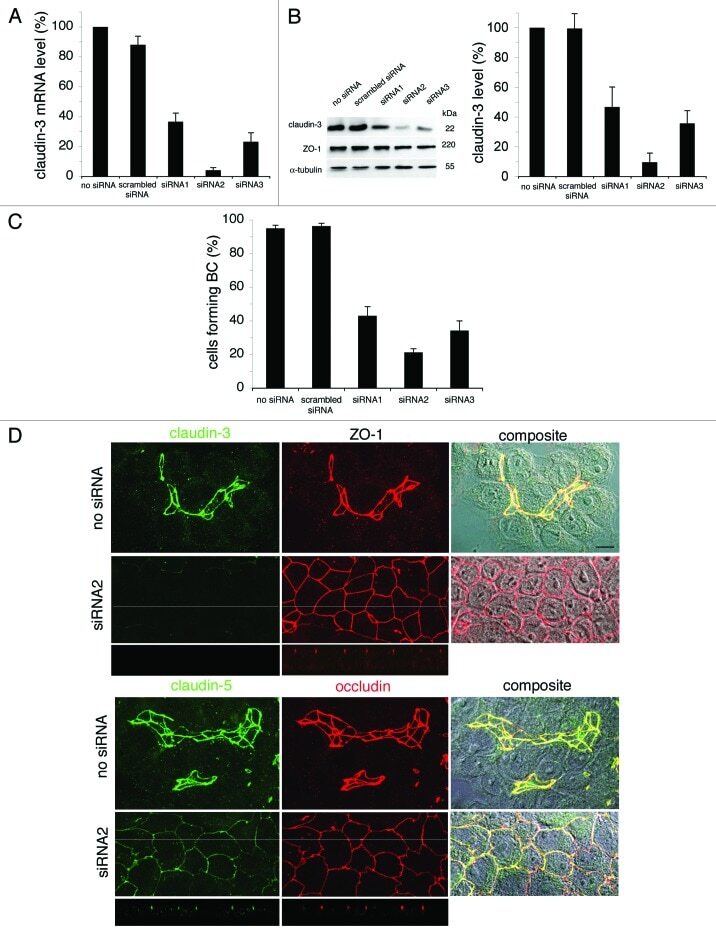
- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
- Figure 2 Time course of urinary claudin-3:creatinine ratio of two representative of the last five children undergoing spinal fusion surgery.
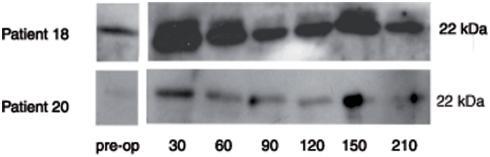
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
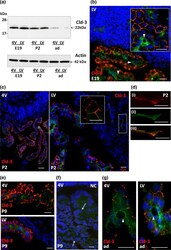
- Experimental details
- Fig. 3 Western blot analysis and cellular distribution of claudin-3 in choroid plexus and cerebral parenchyma during rat brain development. a Representative Western blot of LVCP and 4VCP dissected from E19, P2, and adult animals (10 mug per lane). Cld-3 protein levels decreased in both types of CP during development ( upper panel band at 22 kDa). Actin used as a loading indicator is shown in the lower panel (42 kDa band). b - g Cld-3 immunoreactivity ( red ) in the developing and adult brain. Double immunolabeling with anti-RECA-1 Ab ( green ) allowed visualizing choroidal and parenchymal vessels (e.g. arrowheads in b , g ), and DAPI was used for nuclei staining. During perinatal stages of development, immunodetection revealed a distinct signal associated with the CP epithelial cell membranes as illustrated for LVCP in E19 ( b ), or in P2 animals ( c ). The inserts in b and c highlight the typical honeycomb pattern of choroidal epithelial cell junctions. The area delimited by the rectangle in c , is enlarged in d , which shows Cld-3 in a parenchymal vessel of the developing brain ( i Cld-3 ( red ), ii RECA-1 ( green ) and iii merge staining). Cld-3 immunoreactivity was observed at the epithelial junctions in CPs of P9 animals ( e ), and could still be detected in adult CP ( g ). f A negative control (NC), run in the absence of primary antibody, for a P9 animal ( arrow CP). Scale bars 20 mum. Abbreviations as in Fig. 2
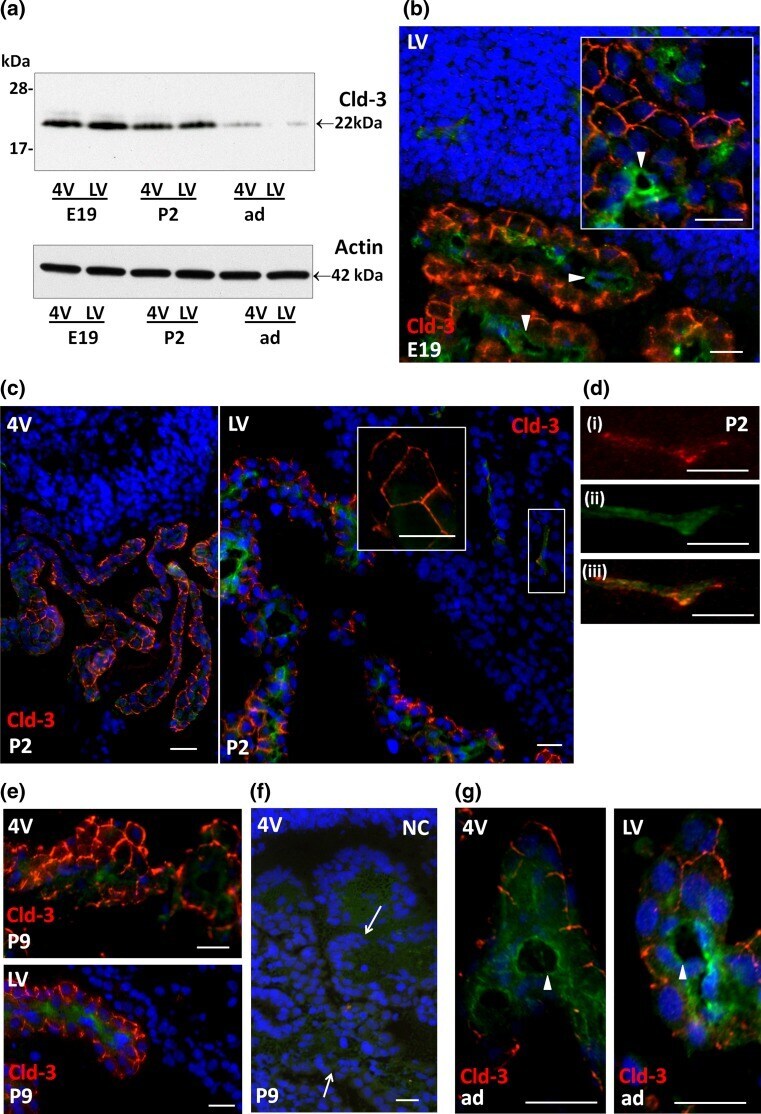
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
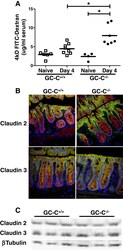
- Experimental details
- Figure 5 GC-C is essential to maintain intestinal barrier function during C. rodentium infection. A : GC-C +/+ and GC-C -/- mice were given a FITC-dextran tracer by oral gavage on day 4 p.i. and serum fluorescence was measured four hours later. Relative to naive mice as well as infected GC-C +/+ animals, GC-C -/- mice have a substantial and significant increase in tracer movement from the gut lumen into the blood indicating a loss of intestinal barrier function. (* P
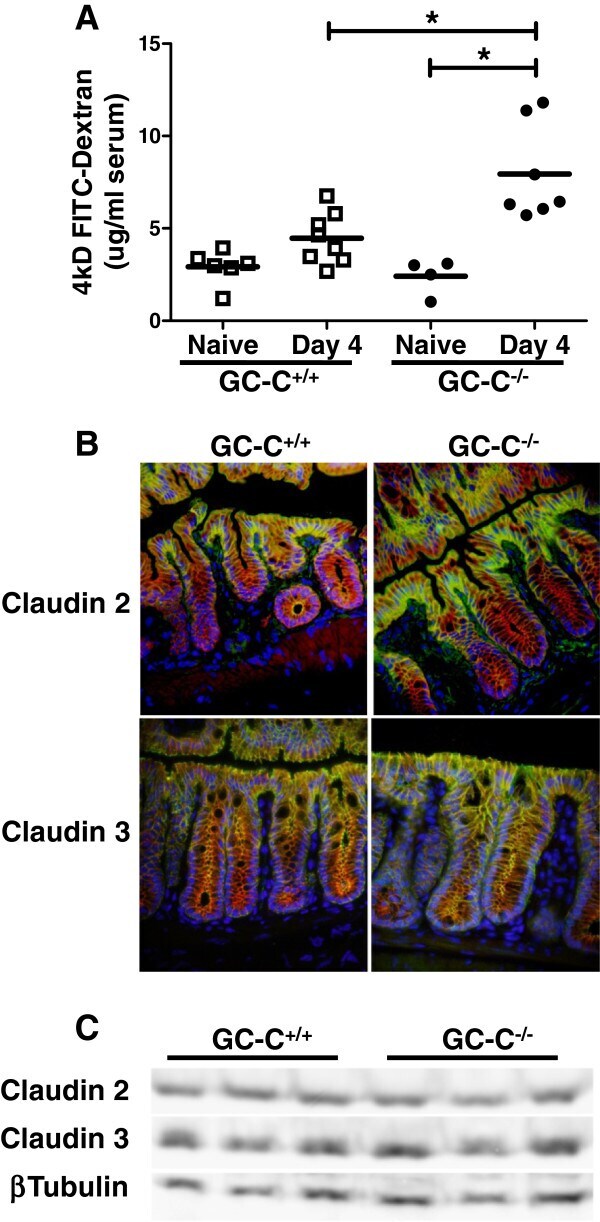
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
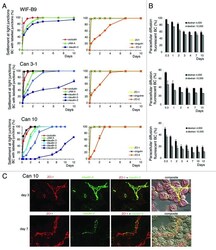
- Experimental details
- Figure 7. Settlement of tight junction (TJ) proteins and paracellular permeability throughout the polarization of cell lines expressing typical hepatocyte polarity. ( A ) Settlement of constitutive (left) and associated- (right) TJ proteins at TJs sealing bile canaliculi (BC), analyzed by immunolocalization. ( B ) Paracellular permeability throughout the settlement process. Values are means of three independent cultures of each cell line ; 11-3 values (not shown) were similar to those shown here for WIF-B9 cells. In Can3-1 cells, claudin-4 was not settled at TJs as the other TJ proteins, and was present at the whole plasma from the first day. ( C ) Localization of claudin-3 with ZO-1 on day 3, and of claudin-2 with ZO-1 on day 7, by confocal microscopy in Can 10 cells. The localization of the TJ proteins (compilation of 21-24 xy sections) is shown, followed by the merge image and the composite image (interferentiel contrast image + merge image), Note that on day 3 claudin-3 and ZO-1 were completely settled at the TJs sealing bile canaliculi, whether bile canaliculi were formed between two cells or more. By contrast, claudin-2 was not completely settled on day 7. It was either absent or partially present at the TJs where ZO-1 localized. Bar 10 mum,
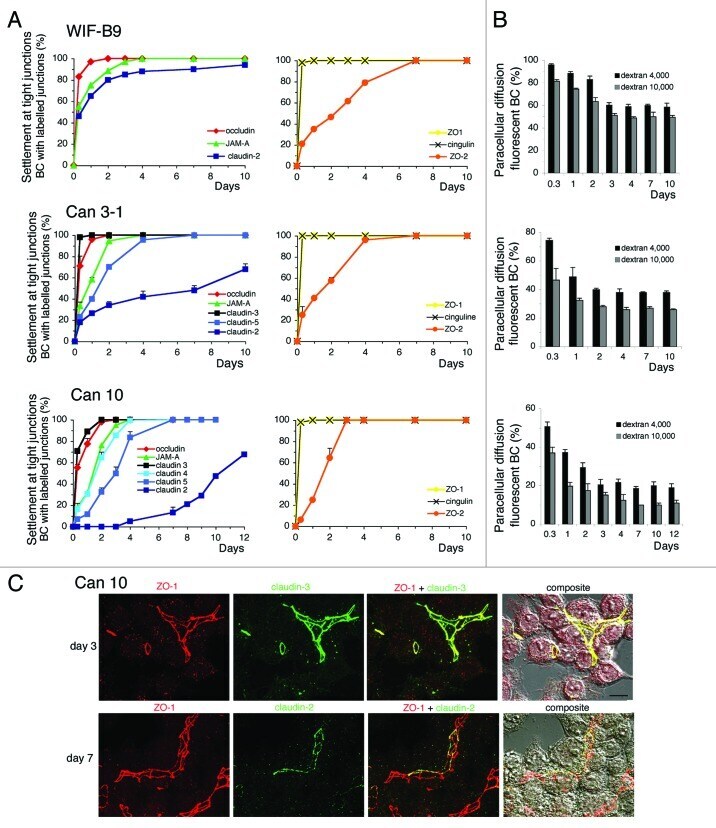
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
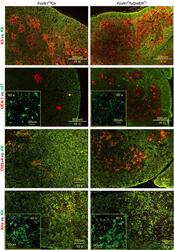
- Experimental details
- Figure 2 Overexpression of FoxN1 mediated by uCreER T+ in juvenile mice induced medullary-biased abnormality of thymic microstructure. Representative immunofluorescence staining shows the thymuses of FoxN1 Tg uCreER T+ juvenile (~20 days after birth) mice (right panels) had infused thymic medullary islets (top panels), an increase in UEA-1 + and Cld3,4 + TECs (panels in middle two rows), an increase in Aire + mTECs (bottom panels), and dimness of 5 beta t + bright dendritic-shaped spots (second row from top), compared to their normal littermate controls (left panels). This experiment was repeated three times with at least three animals in each group producing consistent results
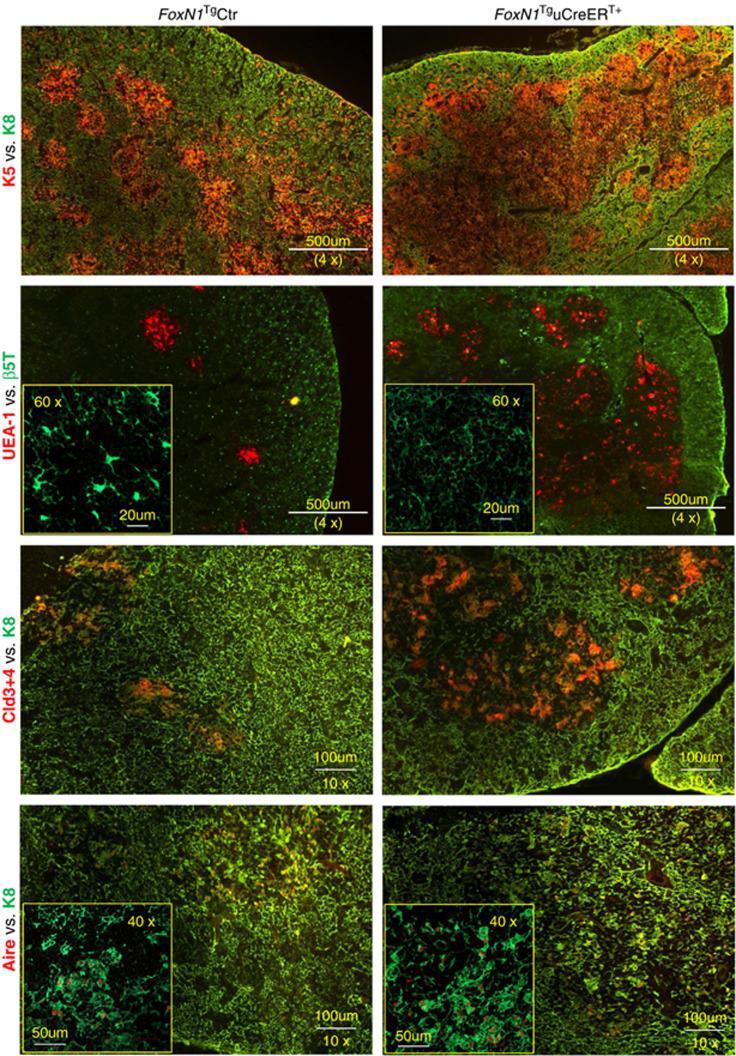
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
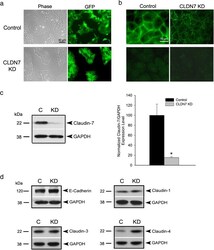
- Experimental details
- Fig. 1 Knockdown of claudin-7 in HCC827 lung cancer cells. a Representative phase and green fluorescence images of live control and claudin-7 KD cells. Both control and claudin-7 shRNA lentivirus constructs contain a GFP expression sequence. b Top panel is the anti-claudin-7 immunofluorescence staining of control and claudin-7 KD cells. Claudin-7 level was dramatically decreased in claudin-7 KD cells. The bottom panel is the anti-GFP immunofluorescence staining. The cells were fixed with 100 % methanol, which lead to GFP proteins leaking out of the cells so that the green fluorescence shown in the top panel was only the claudin-7 signal. c Western blot shows the diminished level of claudin-7 in the KD cells ( left ). The statistical analysis of claudin-7 expression in control and KD cells from five independent experiments is shown on right. * P < 0.05. d Protein expression levels of E-cadherin, claudin-1, -3, and -4 in control and KD cells. Only claudin-4 expression level was increased in KD cells. GAPDH served as a loading control
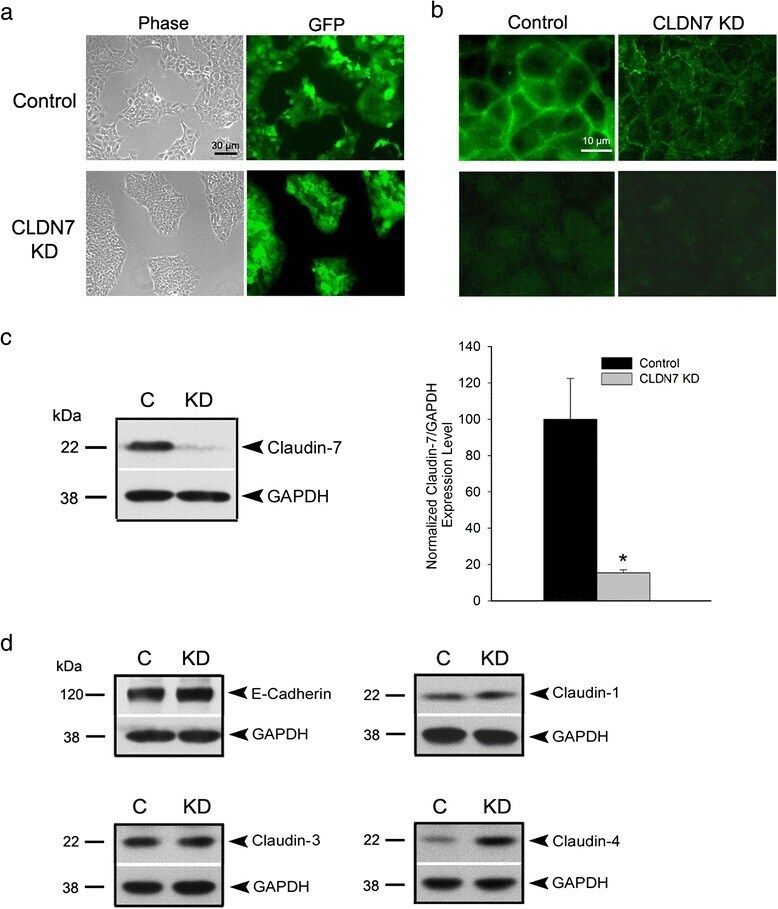
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
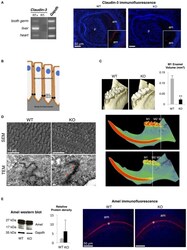
- Experimental details
- Figure 1 Dental phenotype of Claudin-3 KO mice. (A) Claudin-3 mRNA expression was analyzed in mouse tissues by RT-PCR using forward primer 5'-ATG TGG CGC GTT TCG-3' and reverse primer 5'-GCG AGT CGT ACA TTT TGC-3' for Claudin-3 (124 bp), and forward primer 5'-TGT GTC CGT CGT GGA TCT GA-3' and reverse primer 5'-TTG CTG TTG AAG TCG CAG GAG-3' for Gapdh (150 bp). A representative gel image of the selected tissues showed Claudin-3 expression in mouse in post-natal day 3 tooth germ, liver and heart. Identification of claudin-3 in the continuously growing incisor was performed by immunofluorescence using claudin-3 antibody (#34-1700, Invitrogen) at 1/100 dilution and a Goat anti-Rabbit antibody (#R-6394, Invitrogen). Claudin-3 was localized at the distal end of secretory ameloblasts (am). Protein expression was also observed at the basal end of the cells ( n = 3 per group). (p) pulp. (B) Recapitulative schema of claudin TJ proteins expression at the apical end of secretory ameloblasts. Claudin-3, -16, and -19 were shown to be expressed at TJ level of ameloblast secretory ends during enamel formation in the mouse. (C) 3D volume rendering from Micro-CT data showed severe enamel loss on lingual molar cusps in Claudin-3 KO mice. Quantitative analysis confirmed a significant lower enamel volume in Claudin-3 KO mice when compared to WT (0.022 vs. 0.120 mm 3 respectively) ( n = 8 per group). In WT mice, incisor mineralization is observed under the t

- Submitted by
- Invitrogen Antibodies (provider)
- Main image
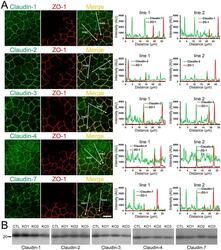
- Experimental details
- Figure 12 Effects of ZO-1 knockout on the localization and expression levels of claudins. (A) Effects of ZO-1 knockout on the localization of claudins. Control and ZO-1 knockout cells of clone 1 were co-cultured on filter inserts. Signal intensity of claudins on lines shown in confocal microscopic images ( arrows ) were analyzed. Claudin-2 fluorescent signal at TJs was increased but claudin-1 and -7 signals at TJs were reduced in ZO-1 knockout cells. Scale bar, 10 um. (B) Immunoblots of claudins in control MDCK II cells and ZO-1 knockout clones. Similar expression levels of claudins were observed in control and knockout cells.
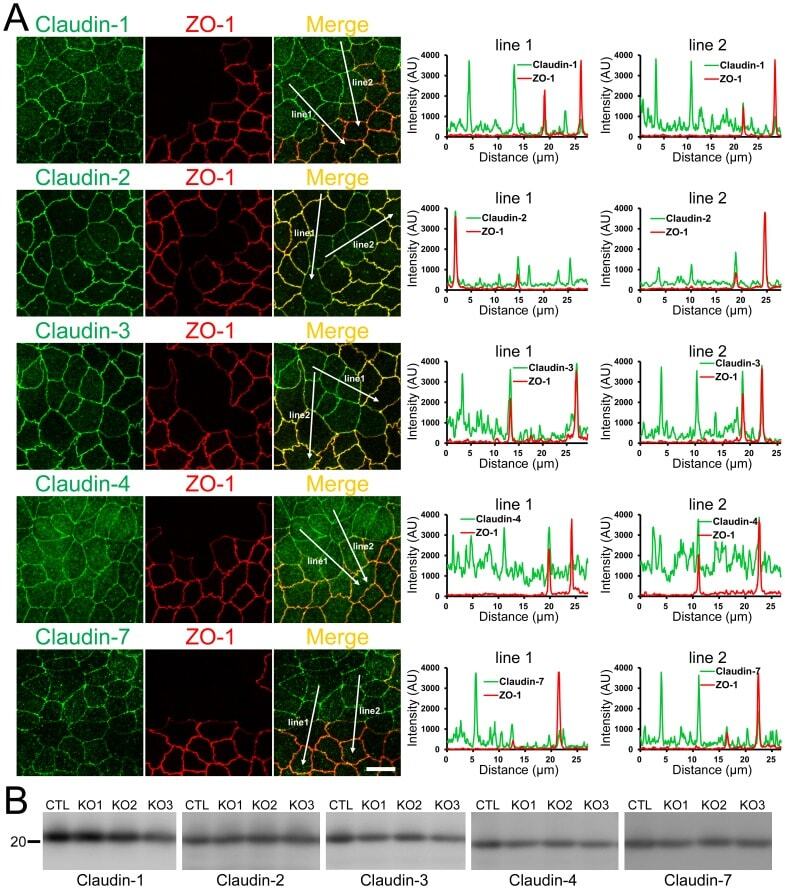
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
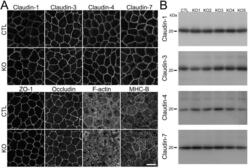
- Experimental details
- Fig 3 Effects of claudin-2 knockout on the localization of other TJ proteins and cytoskeleton. (A) Immunofluorescence analysis of claudin-1, -3, -4, -7, ZO-1, occludin, F-actin, and myosin heavy chain II-B (MHC-B) in control and claudin-2 knockout cells. Claudin-1 and -7 showed a tendency to be more clearly localized at TJs in claudin-2 knockout cells. Scale bar = 10 mum. (B) Immunoblots of claudin-1, -3, -4, and -7 in control MDCK II cells and claudin-2 knockout clones. Similar expression levels of claudin-1, -3, -4, and -7 were observed in control cells and knockout clones.
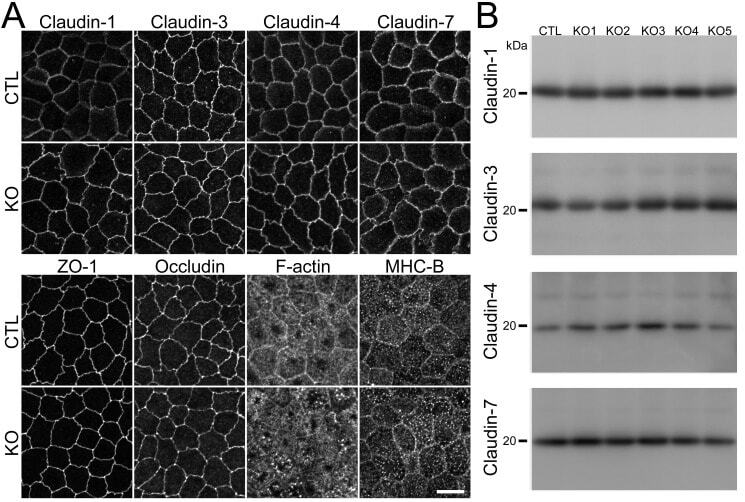
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- Fig. 8 Immunohistochemical detection of claudins in choroid plexuses of developing human brain. Immunoreactivity of Cld-1, -2, and -3 is presented on the left, central, and right panels , respectively, with hematoxylin counterstaining. a Staining of a WD8 fetal brain. Cld-1 labeling, observed in both LVCP and 4VCP, was continuous and localized apically at intercellular junctions of the epithelium ( arrows in inserted high magnification micrograph ). Cld-2 immunoreactivity displayed a similar apical, but patchy pattern in both LVCP and 4VCP. The higher magnification ( insert in LVCP) illustrates the discontinuous labeling ( arrows ). Cld-3 was also detected in the epithelium of both CP ( arrows ). Inserts are higher magnifications highlighting the continuous staining pattern. b Staining of a WD22 fetal brain. Cld-1 immunoreactivity was present at all epithelial junctions (shown in 4VCP). A continuous apical labeling of the epithelium was observed for Cld-2 (LVCP is shown). Cld-3 labeling was faint but continuous in LVCP epithelial cells. c Staining of a WD39 fetal brain. A distinct and continuous labeling of the choroidal epithelium was observed for all Cld-1 (shown for LVCP), -2 (shown for 4VCP), and -3. The continuous signal for Cld-3 was only associated with the epithelium ( arrows ) and not with stromal vessels ( arrowheads in LVCP). The insert shows a similar Cld-3 staining for 4VCP. Abbreviations as in Fig. 2 . Scale bar 50 mum
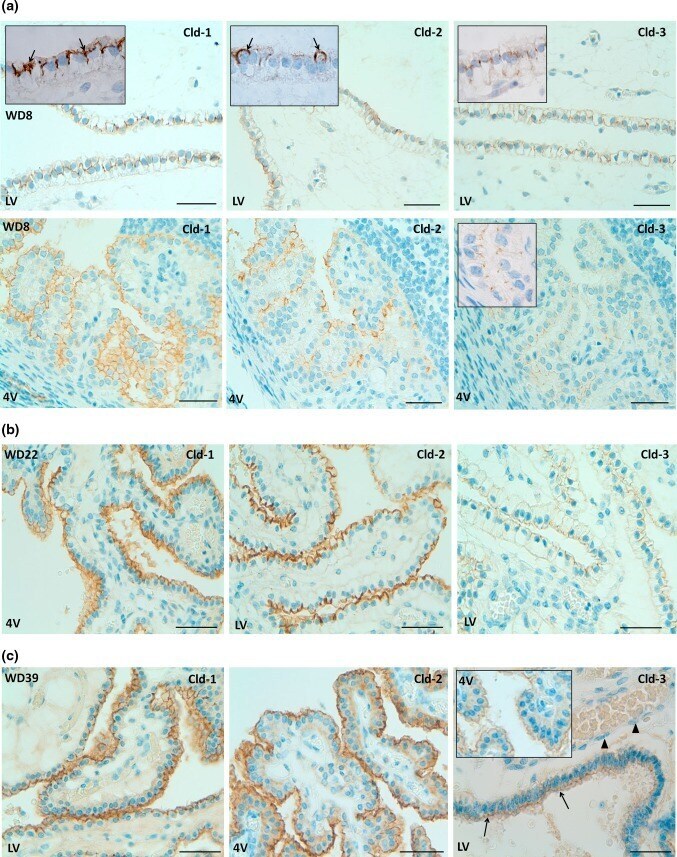
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
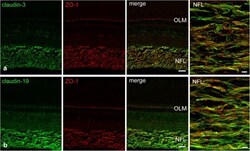
- Experimental details
- FIGURE 6 Double-immunostains for the tight junction proteins (green) claudin-3 (a) and claudin-19 (b) and for the adaptor protein ZO-1 (red, a,b ). The nerve fiber layer (NFL) shown at high magnification in the right images, is strongly positive for both claudin antibodies. Immunoreactivity for ZO-1 was also detected in the NFL, as well as in the outer limiting membrane (OLM). In the NFL, claudin and ZO-1 stains largely overlap with the claudins showing more continuous label strands often but not always running parallel to nerve fibers. Scale bar 20 mum for low power, 5 mum for high power images.
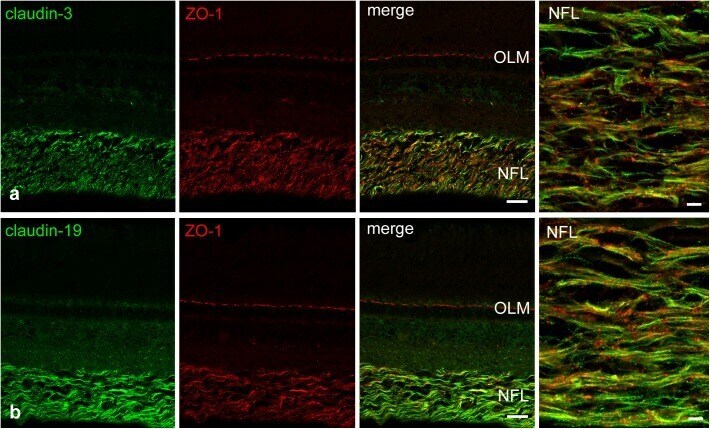
- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
- Figure 3 Representative photographs of claudin-2, claudin-3, claudin-4, and zonula occludens-1 immunostained in longitudinal jejunal sections of the three experimental groups. Claudin-2 immunoreaction in the sham-operated group (a) was low and was mainly found in the cytoplasm of the intestinal epithelial cells. Claudin-2 expression in the groups exposed to ischemia-reperfusion was substantially higher and was principally found in the lateral membrane of the epithelial cells (b, c). The sham-operated group presented strong membrane claudin-3 immunostaining (d), which was remarkably decreased in the ischemia-reperfusion- (IR-) group (e), but not in the postconditioned- (PC-) group (f). Minimal claudin-4 immunostaining was observed in the jejunal samples of the sham-operated animals (g). The two groups challenged with 60 minutes of mesenteric ischemia and 6 hours of reperfusion (h, i) showed high membrane claudin-4 expression, which was limited to the tips of the villi. Zonula occludens-1 was strongly immunostained in the jejunal samples of the sham-operated animals (j). Following small intestinal ischemia-reperfusion, intestinal epithelial cell with zonula occludens-1 membrane immunostaining notably decreased in the IR-group (k); in the PC-group minimal loss of zonula occludens-1 membrane immunostaining was detected (l). Each photograph was taken with the same magnification. Scale bar: 20 mu m.
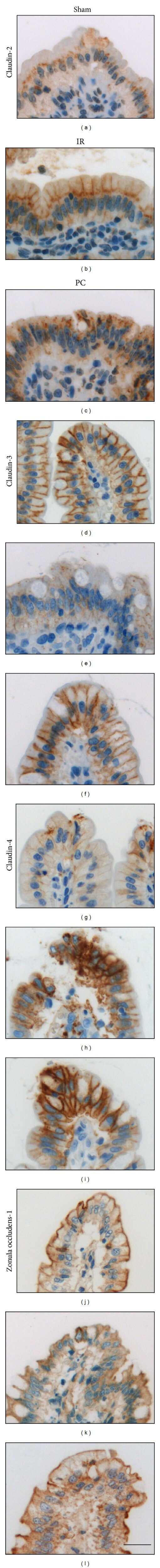
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
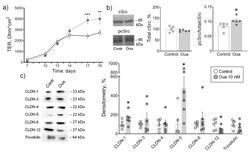
- Experimental details
- Figure 1 Epithelial barrier formation in IPEC-J2 cells grown in control medium and in the presence of 10 nM ouabain (Oua). After 7 days of culture, transepithelial resistance (TER) values were determined every 2-3 days using an EVOM volt-ohmmeter; after 19 days, cells were used for subsequent analysis. ( a ) Transepithelial resistance (TER) ( n = 6 for each group). ( b ) The expression level of total cSrc (central plot, the relative expression of cSrc protein shown as a percentage of average level in control samples taken for 100%) and the cSrc-kinase activation by phosphorylation (right plot, shown as a ratio between immunoblot intensity corresponding to phosphorylated pcSrc over total cSrc, as it is shown in the representative immunoblots in left panel) ( n = 5 for each group). ( c ) Western blot analysis of claudin (CLDN) and tricellulin expression ( n = 6 for each group); left panel shows representative immunoblots. Original images for Western blots using Stain-Free gels as a loading control are shown in Supplemental Materials . The number of symbols corresponds to the number of samples. One-way ANOVA with Dunnett correction: * p < 0.05, ** p < 0.01 and *** p < 0.001 compared with the corresponding control.
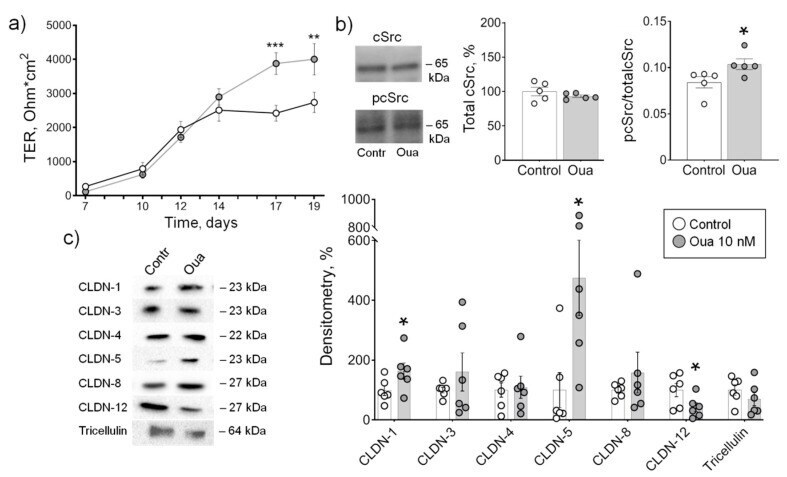
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
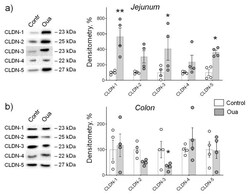
- Experimental details
- Figure 3 Chronic exposure to ouabain (Oua) differently alters claudin expression in rat jejunum ( a ) and colon ( b ). Rats were intraperitoneally injected with ouabain (1 µg/kg) for 4 days. Western blot analysis of claudins (CLDN) expression ( n = 4 for each group); left panel shows representative immunoblots. Original images for Western blots using Stain-Free gels as a loading control are shown in Supplemental Materials . The number of symbols corresponds to the number of samples. One-way ANOVA with Dunnett correction: * p < 0.05 and ** p < 0.01 compared with the corresponding control (vehicle treated group).
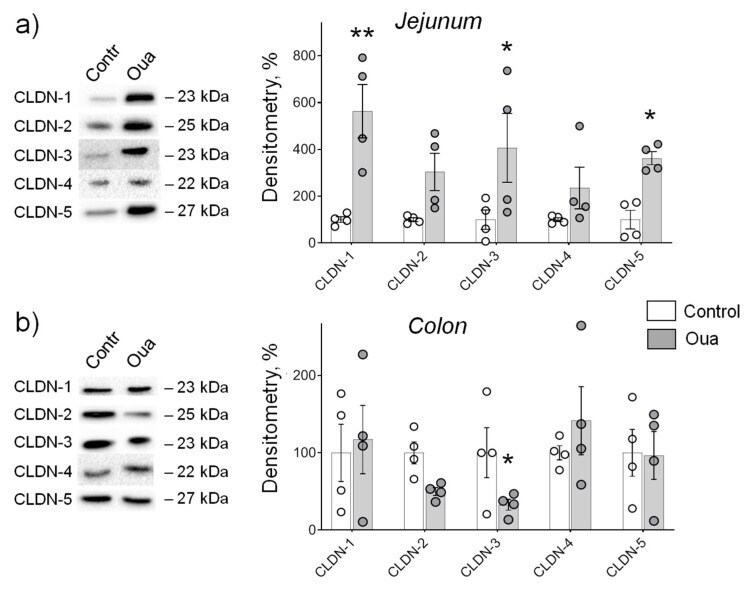
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
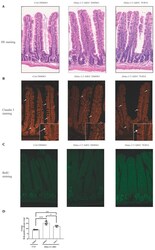
- Experimental details
- Figure 5 Tofacitinib treatment ameliorates intestinal architecture and decreases intestinal epithelial cell proliferation in villin-Cre Hdac1 and Hdac2 deficient mice. Jejunal tissue sections, from wild-type (Ctrl) and villin-Cre Hdac1 -/- ; Hdac2 -/- mice ( Hdac1/2 DeltaIEC) treated without or with Tofacitinib (DMSO or TOFA) for five days, were stained with hematoxylin and eosin (HE) ( A ) (Magnification: 10x) or were stained with an antibody against claudin 3 ( n = 3) (white arrows) ( B ) (Magnification: 20x). ( C ) Wild-type (Ctrl) and villin-Cre Hdac1 -/- ; Hdac2 -/- mice, treated without or with Tofacitinib (DMSO or TOFA) for five days, were injected with BrdU for 2 h. Jejunal tissue sections were stained with an antibody against incorporated BrdU. Magnification: 10x. ( D ) The average number of BrdU-labeled proliferative cells per jejunal crypts was measured ( n = 3, 20-30 crypts each). Results represent the mean +- SEM (* p
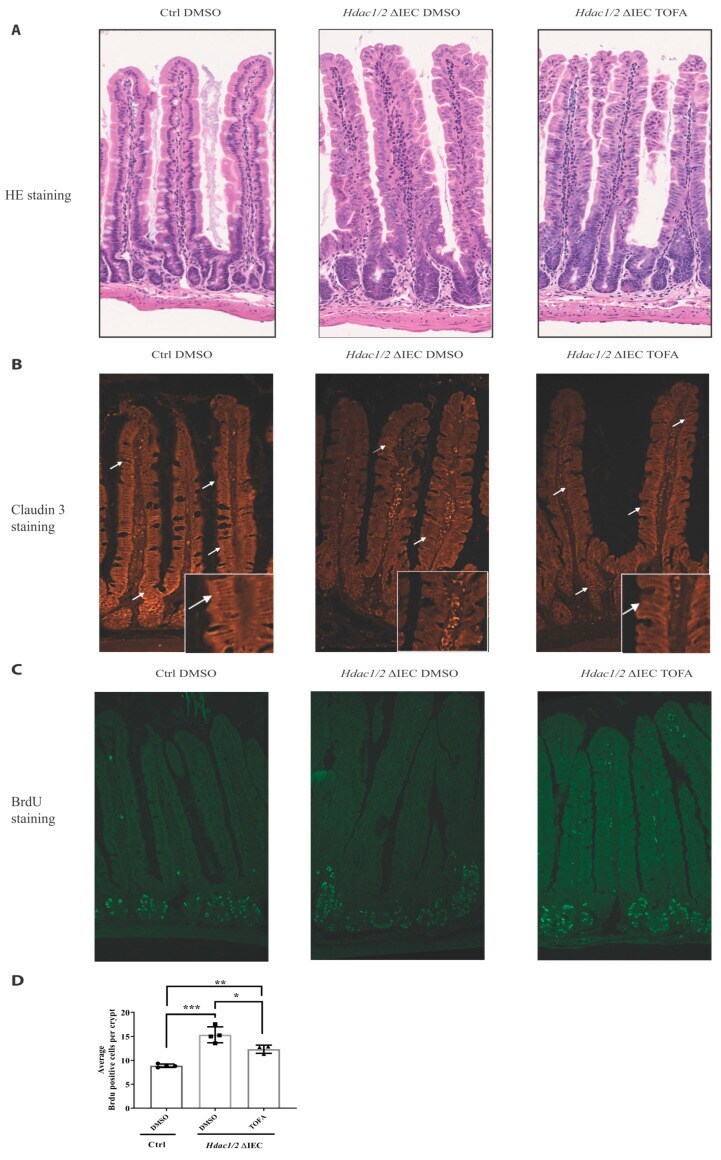
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- Figure 3 Quercetin attenuated diquat-induced ROS accumulation and reduced protein abundance of tight junction proteins in IPEC-1 cells. IPEC-1 cells were treated as described in Figure 2 . ( A ) DCFH-DA-positive populations were detected by flow cytometry analysis. ( B ) Intracellular ROS levels were determined by fluorescence microscope (magnification x200). ( C ) Intracellular GSH levels were measured. ( D , E ) Protein abundance for ZO-1, ZO-2, ZO-3, occludin, claudin-1, claudin-3, and claudin-4 were determined and analyzed by the Western blot assay. beta-actin was used as the loading control. Representative experimental results were repeated in three independent experiments and values are means +- SEMs, n = 3. Means without a common letter are considered as a significant difference, p < 0.05.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- Figure 4 Effects of whole LP powder, methanol (MetOH) extract, and MetOH extraction residue (MetOH residue) on tight junction protein expression in the colon of dextran sodium sulfate (DSS)-induced colitic mice. Protein expression of zonula occludens (ZO)-1, ZO-2, occludin, claudin-3, and claudin-7 in the colon of mice fed diets with and without whole LP powder, MetOH extract, and MetOH residue, with or without DSS administration, as determined by immunoblot analysis. Values are the mean +- SEM ( n = 7). Means without a common letter differ, p < 0.05.
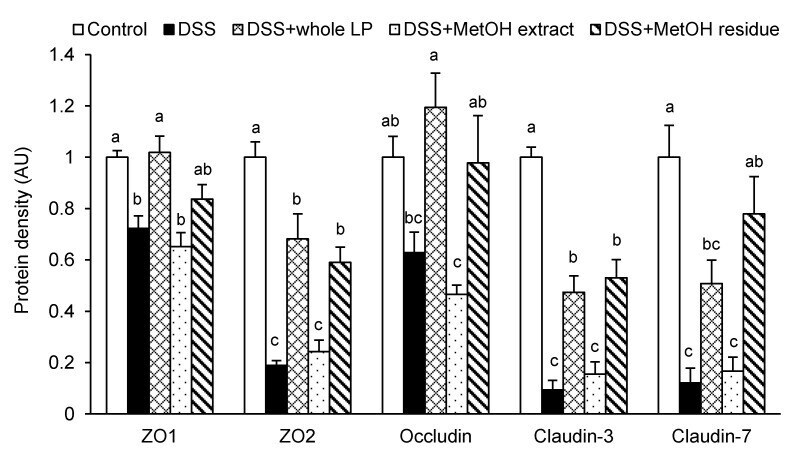
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- Figure 5 Effects of whole LP powder, methanol (MetOH extract), and MetOH extraction residue (MetOH residue) on claudin-3 expression in the colon of dextran sodium sulfate (DSS)-induced colitic mice. Immunolocalization of claudin-3 in the colon of mice fed diets with and without whole LP powder, MetOH extract, and MetOH residue, with or without DSS administration, as analyzed by immunofluorescence microscopy. Representative images of seven mice in each group are shown.
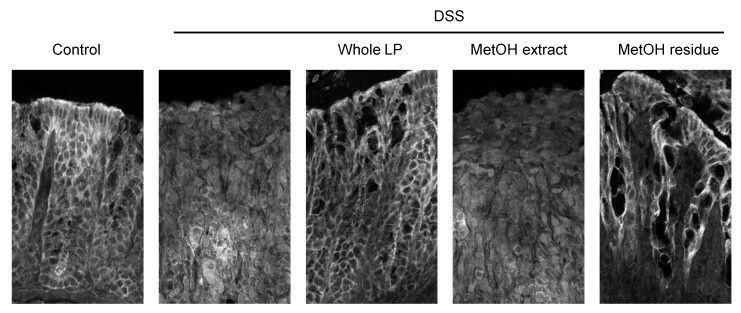
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
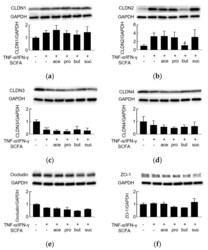
- Experimental details
- Figure 2 The effect of short-chain fatty acids on TNF-alpha/IFN-gamma-induced dysregulation of tight junction proteins. Western blot analyses of ( a ) claudin-1 (CLDN1, 22 kDa), ( b ) claudin-2 (CLDN2, 22 kDa), ( c ) claudin-3 (CLDN3, 22 kDa), ( d ) claudin-4 (CLDN4, 22 kDa), ( e ) occludin (65 kDa), and ( f ) ZO-1 (220 kDa) proteins were performed following stimulation on the basolateral side of the membrane, with the combination of TNF-alpha/IFN-gamma (10 ng/mL), and on the apical side with acetate (ace), propionate (pro), butyrate (but), or succinate (suc) (2 mM). * p < 0.05 compared to medium-treated control; # p < 0.05 compared to TNF-alpha/IFN-gamma group; **** p < 0.0001 compared to medium-treated control.
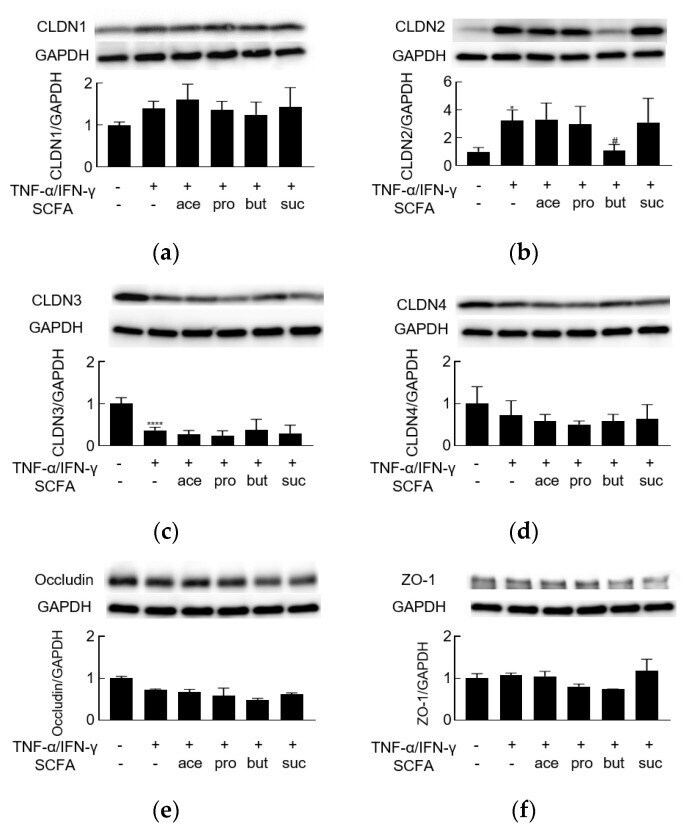
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
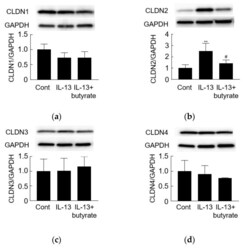
- Experimental details
- Figure 4 The effect of butyrate on IL-13-induced dysregulation of tight junction proteins. Western blot analyses of ( a ) claudin-1 (CLDN1, 22 kDa), ( b ) claudin-2 (CLDN2, 22 kDa), ( c ) claudin-3 (CLDN3, 22 kDa), ( d ) claudin-4 (CLDN4, 22 kDa), ( e ) occludin (65 kDa), and ( f ) ZO-1 (220 kDa) proteins were performed following stimulation on the basolateral side of the membrane with IL-13 (10 ng/mL) and on the apical side with butyrate (but, 2 mM). ** p < 0.01 compared to medium-treated control (Cont); # p < 0.05 compared to IL-13 group.
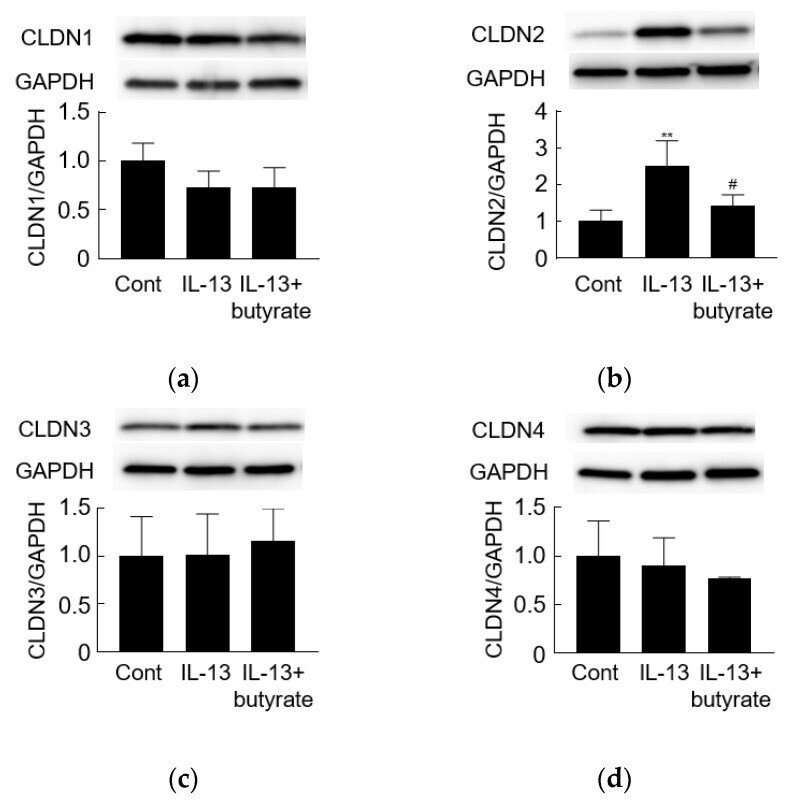
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
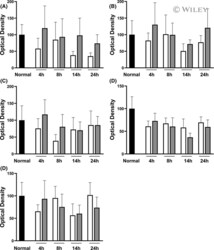
- Experimental details
- 3 FIGURE Immunoblotting using antibodies against zonula occludens (ZO)-1 (A), tricellulin (B), occludin (C), claudin-3 (D), and claudin-4 (E) in intestines undergoing VP only (white bar) and in intestines receiving additional luminal preservation (light gray bar). The black bar denotes the baseline value obtained in the same intestines before cold preservation. Data expressed as mean +- standard error of the mean, * p < .05.
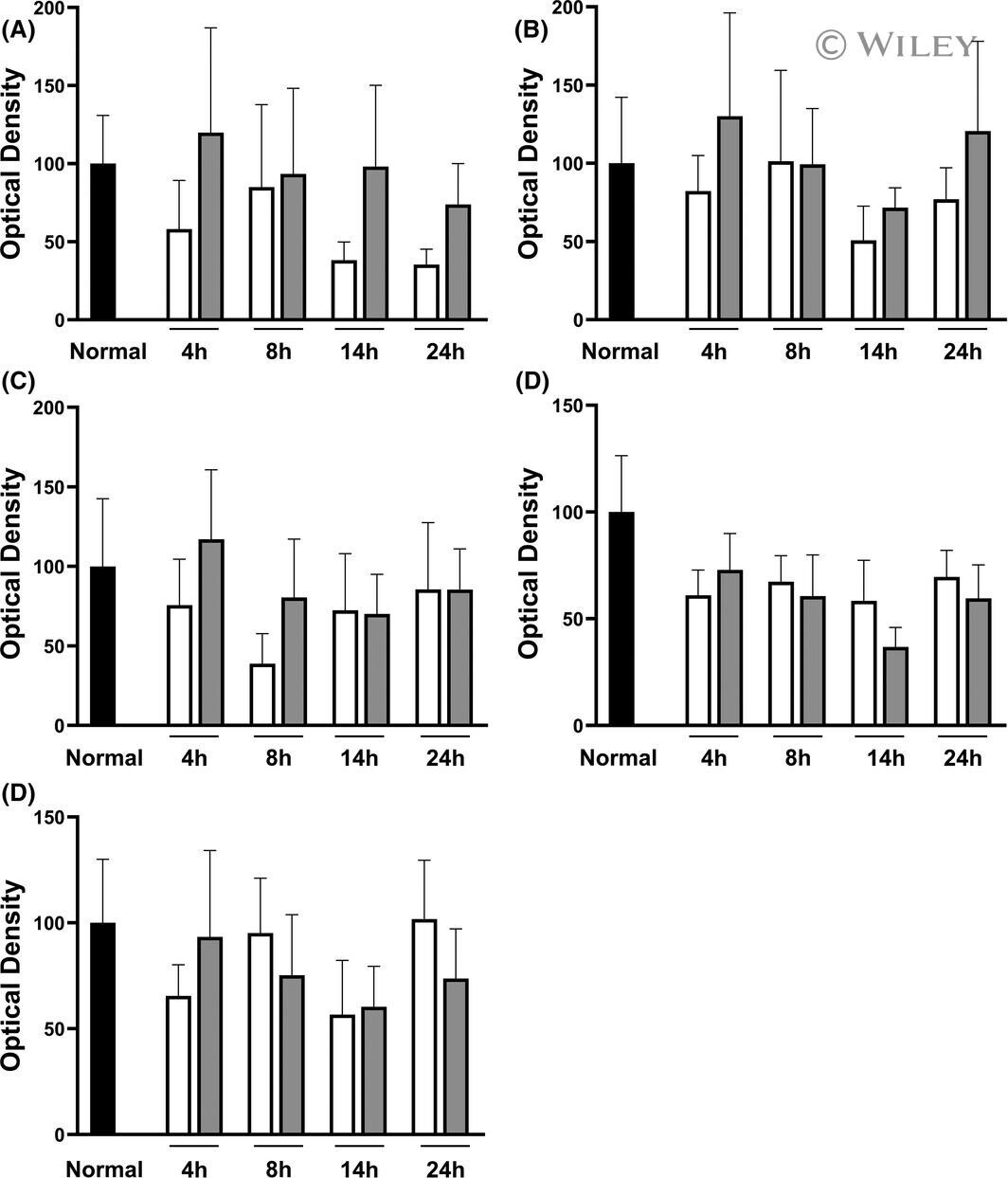
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
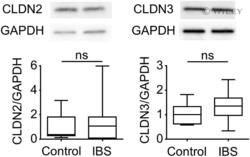
- Experimental details
- 4 FIGURE Expression levels of claudin-2 (CLDN2) and claudin-3 (CLDN3) proteins in duodenal mucosa. CLDN2 (22 kDa), CLDN3 (22 kDa), and GAPDH protein expression were evaluated by western blotting in controls ( n = 13) and patients with IBS ( n = 12). GAPDH, glyceraldehyde 3-phosphate dehydrogenase; ns, not significant.
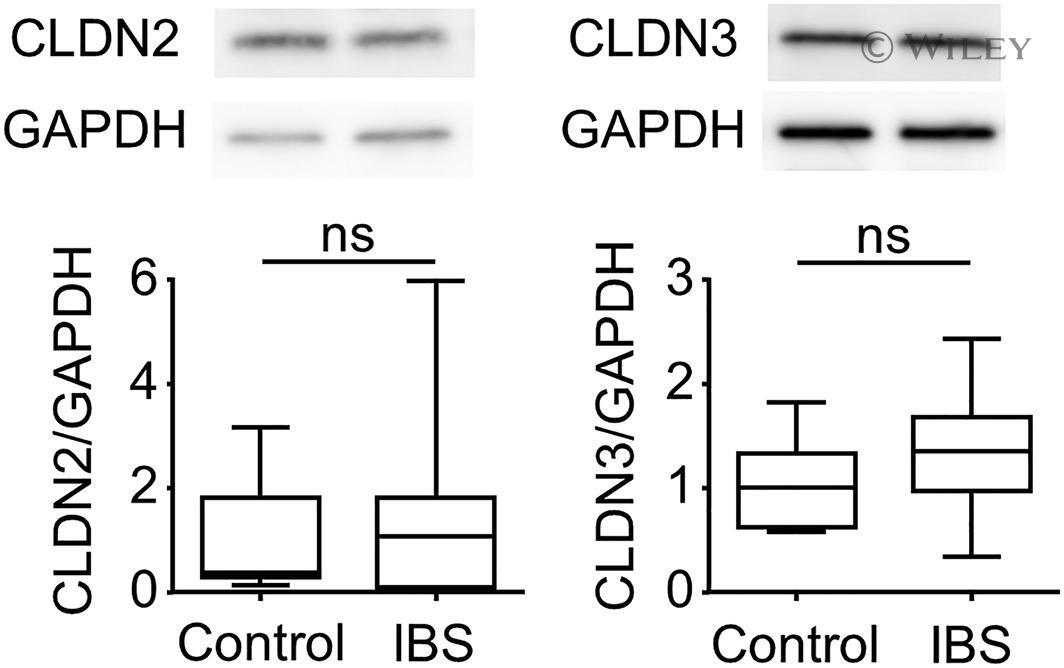
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
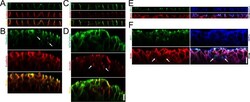
- Experimental details
- Fig 6 Localization of claudin-3, E-cadherin, occludin and Na + /K + ATPase under the 'Apical' and 'Basal' conditions in MDCK I cells and barrier function of TJs around the cavities within the multi-layered epithelia. (A and B) Immunofluorescence microscopy for claudin-3 and E-cadherin in z-axis plane under the 'Apical' (A) and 'Basal' (B) conditions in MDCK I cells. Signals of claudin-3 were observed within the multi-layered MDCK I cells under the 'Basal' condition ( arrows ). (C and D) Immunofluorescence microscopy for occludin and Na + /K + ATPase in z-axis plane under the 'Apical' (C) and 'Basal' (D) conditions in MDCK I cells. Signals of occludin were observed within the multi-layered MDCK I cells under the 'Basal' condition ( arrows ). (E and F) A tracer experiment using a primary amine-reactive biotinylation reagent was performed in MDCK I cells cultured under the 'Apical' (E) and 'Basal' (F) conditions. The biotinylation reagent was administered in the basal side for 10 min, and the bound biotin was detected by streptavidin. The biotinylation reagent appeared to stop at TJs around the cavities within the multi-layered epithelia under the 'Basal' condition ( arrows ). Scale bars = 10 mum.
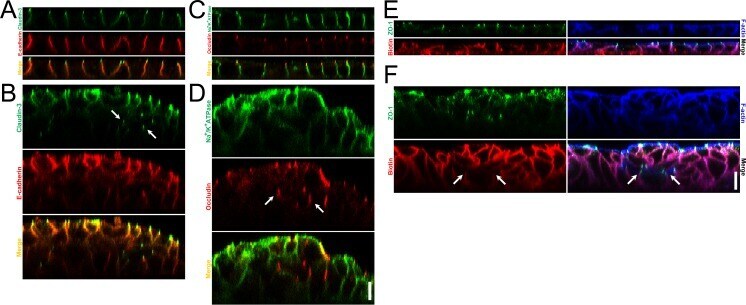
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
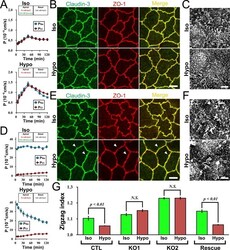
- Experimental details
- Fig 9 Effects of apical hyposmolality in claudin-2 knockout MDCK II cells. (A) Time course of P Na and P Cl in claudin-2 knockout MDCK II cell clone (knockout clone 1 in a previous study [ 22 ]). The values of P Na and P Cl were increased and then decreased during 120 min of incubation, and the charge selectivity showed no remarkable change under the 'apical hyposmotic' condition. N = 4 for each experiment. (B) Immunofluorescence microscopy for claudin-3 and ZO-1. The signals of claudin-3 and ZO-1 under the 'apical hyposmotic' condition were similar to those under the 'apical isosmotic' condition. Scale bar = 5 mum. (C) Scanning electron microscopy of claudin-2 knockout MDCK II cells. Scale bar = 2 mum. (D) Time course of P Na and P Cl in claudin-2 knockout MDCK II cells expressing exogenous claudin-2. The clones were established from the claudin-2 knockout clone 1 as described in a previous study (F4 clone in a previous study [ 22 ]). P Na was decreased and P Cl was increased gradually with time under the 'apical hyposmotic' condition. N = 3 for each experiment. (E) Immunofluorescence microscopy for claudin-3 and ZO-1. The protruded signals of claudin-3 and ZO-1 from the lines of cell-cell contacts were observed under the 'apical hyposmotic' condition ( arrowheads ). Scale bar = 5 mum. (F) Scanning electron microscopy of claudin-2 knockout MDCK II cells expressing exogenous claudin-2. Globular structures were observed around cell-cell contacts under the 'apical hyposmotic' c
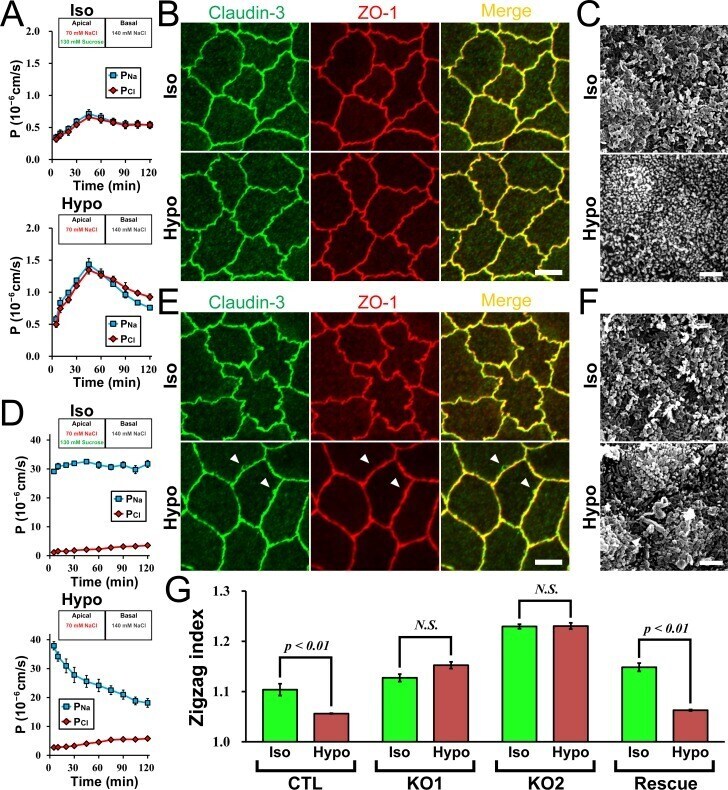
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
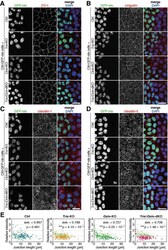
- Experimental details
- FIGURE 2: TJs of Tric -KO, Ocln -KO, and Tric / Ocln -dKO cells appear normal in fluorescence microscopy. (A-D) Ctrl, Tric -KO #1, Ocln -KO #1, or Tric / Ocln -dKO #1 cells were mixed and cocultured with Ctrl-GFP-nls cells (green). Cells were immunostained with rat anti-ZO-1 mAb (A), mouse anti-cingulin mAb (B), mouse anti-claudin-1 mAb (C) or rabbit anti-claudin-3 pAb (D) (red) and stained with DAPI (blue). Scale bars, 20 um. (E) Quantification of fluorescence intensity of claudin-3 at cell-cell junctions. The intensity at each cell-cell junction in the Ctrl (dark blue closed circles), Tric -KO #1 (orange closed circles), Ocln -KO #1 (green closed circles), and Tric / Ocln -dKO #1 cells (magenta closed circles) were normalized to the averaged intensity in the Ctrl-GFP-nls cells (lime-green open circles) and plotted against the length of the cell-cell junctions. n = 158 and 120 (Ctrl), 202 and 134 ( Tric -KO), 160 and 139 ( Ocln -KO), and 209 and 136 ( Tric / Ocln -dKO). *** p < 0.001 in Welch's t test of junction-length-weighted average of the fluorescence intensity.

- Submitted by
- Invitrogen Antibodies (provider)
- Main image
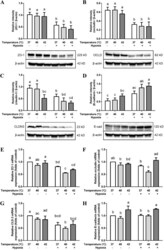
- Experimental details
- Figure 4 Effects of hypoxia and/or heat treatment on TJ/AJ proteins in the co-culture model. Relative protein expression of ZO-1 ( A ), occludin ( B ), claudin-3 ( C ) and E-cadherin ( D ) assessed by Western Blot. All target proteins were normalized to reference protein beta-actin. ZO-1 ( E ), occludin ( F ), claudin-3 ( G ) and E-cadherin ( H ) mRNA levels were assessed by qRT-PCR. The target genes were normalized to housekeeping gene beta-actin. All values were presented as means +- SD (N = 3, n = 3). Statistical differences were analyzed by two-way ANOVA followed by the Bonferroni's multiple comparison test. Means without a common letter differ at p < 0.05. OCLD occludin; CLDN3 claudin-3; E-cad E-cadherin.
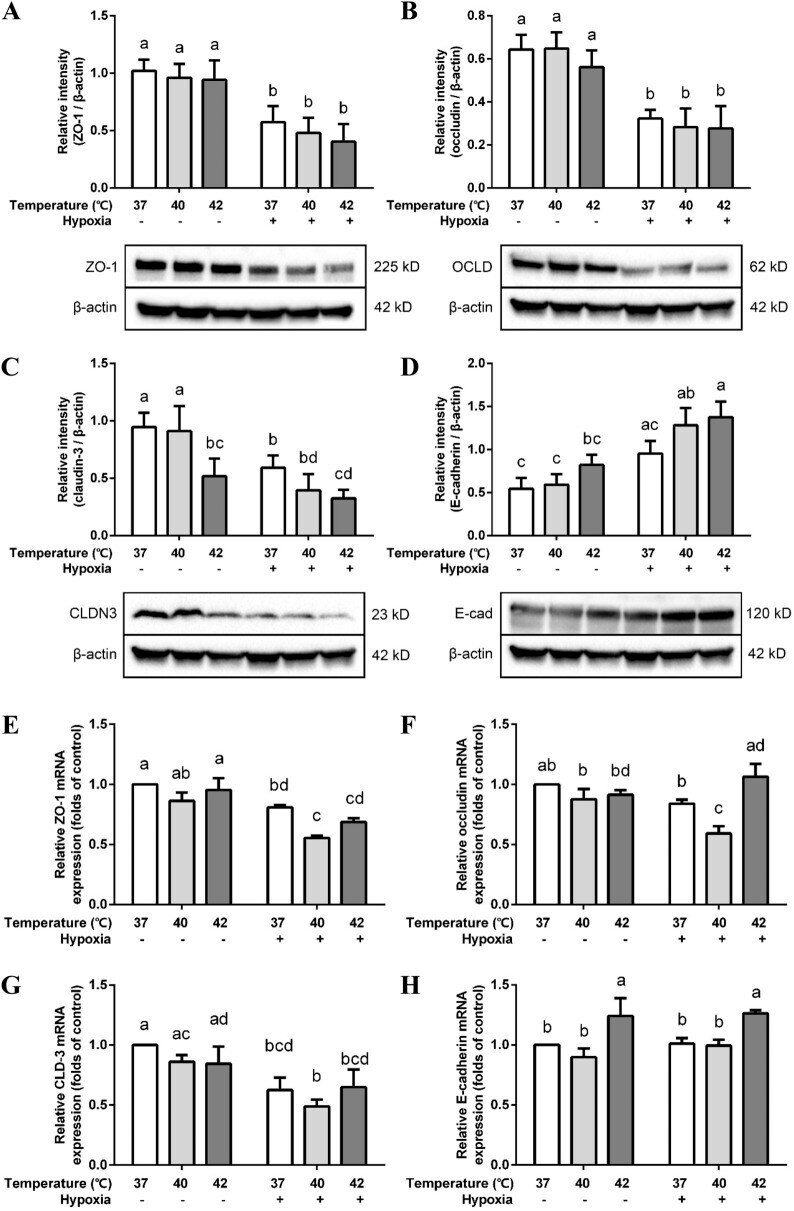
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- 10.1371/journal.pone.0256143.g005 Fig 5 Cellular arrangement (cross sections) of jejunum organoids grown in 2D on Snapwells (r) for 18 days. Immunohistochemical staining (including isotype controls) of Claudin-2 and Claudin-3 was performed. Blue triangles indicate ceklls with positive Claudin-2 signal, orange triangles indicate cells with a positive Claudin-3 signal. Representative figures are shown, chosen from three independent experiments with three technical replicates per staining.
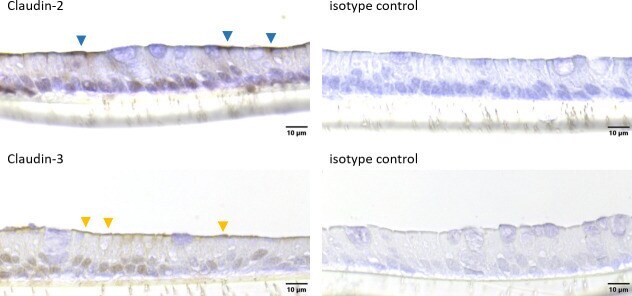
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
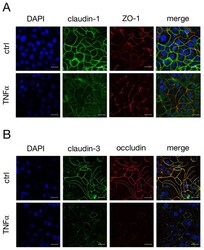
- Experimental details
- Figure 3 Confocal laser scanning immunofluorescence microscopy was performed to analyze the localization of ( A ) claudin-1 (green) and ZO-1 (red), or ( B ) claudin-3 (green) and occludin (red) after incubation with TNFalpha for 48 h. Nuclei were stained in blue (DAPI). Claudin-3 shows a strong colocalization with occludin (merge, yellow signal, B) (scale bar: 20 um; n = 4; representative pictures).
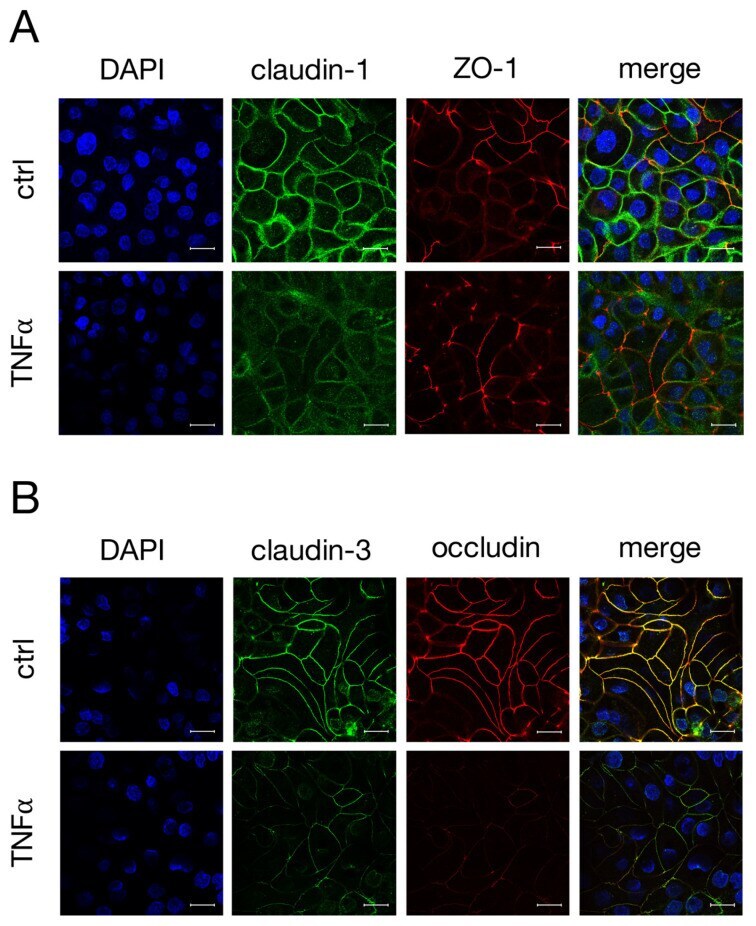
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
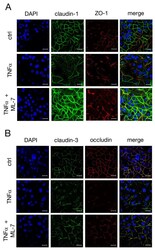
- Experimental details
- Figure 7 Confocal laser scanning immunofluorescence microscopy of ( A ) claudin-1 (green) and ZO-1 (red), as well as ( B ) claudin-3 (green) and occludin (red), nuclei were stained in blue (DAPI). IPEC-J2 monolayers were treated for 48 h with TNFalpha in the presence or absence of the specific MLCK blocker ML-7. ( B ) The yellow signal in the merged pictures represents the colocalization of claudin-3 and occludin (scale bar: 20 um; n = 4; representative images).
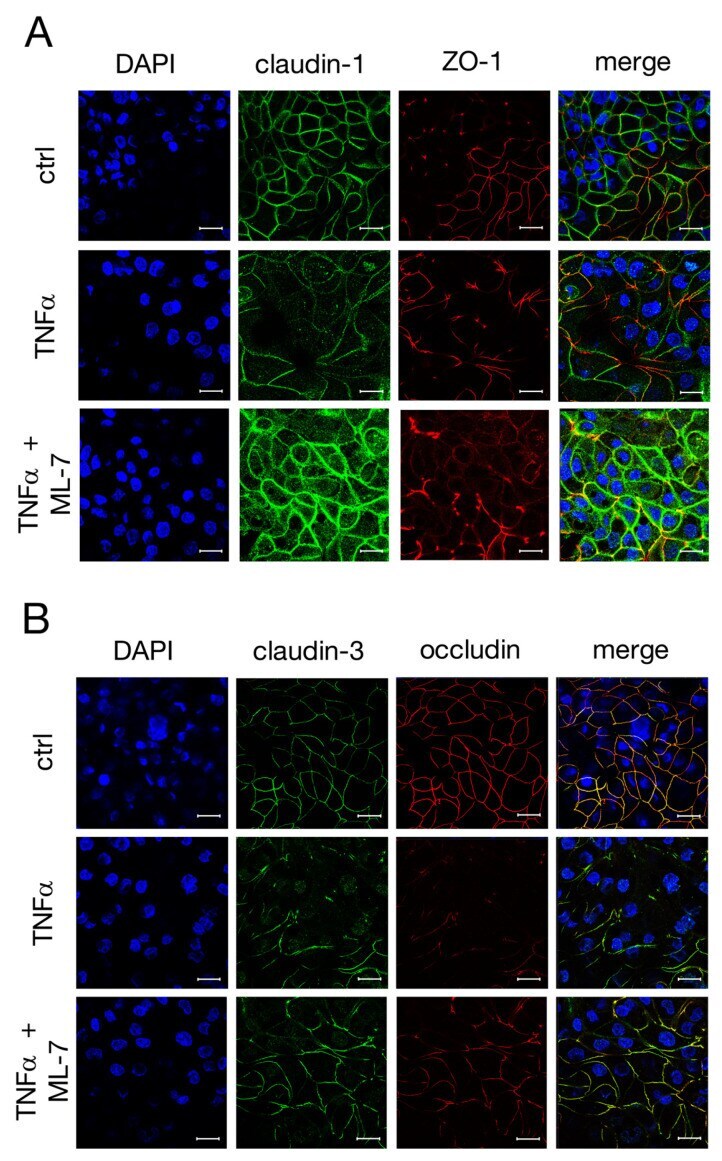
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
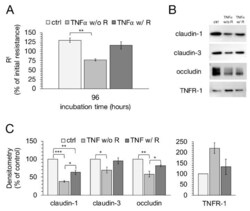
- Experimental details
- Figure 8 After IPEC-J2 cells were treated for 48 h with 1000 U/mL TNFalpha, a medium exchange was carried out and cells were either further incubated with the cytokine (TNFalpha without recovery; w/o R) or incubated with normal medium without TNFalpha (TNFalpha with recovery; w/R) for another 48 h. ( A ) TEER values of groups treated with TNFalpha removal were compared to cells treated without removal of the cytokine, respectively (one-way ANOVA, ** p < 0.01; n = 7-13). ( B ) Representative Western blot bands and ( C ) Densitometry of IPEC-J2 cells with or without recovery after 48 h incubation with TNFalpha revealed an increase in claudin-1 and occludin after removal of the cytokine, while claudin-3 expression did not show significant changes. The expression level of TNFR-1 also did not seem to be affected by TNFalpha removal. The shown data are presented as mean +- SEM (one-way ANOVA, * p < 0.05, ** p < 0.01, *** p < 0.001; n = 3).
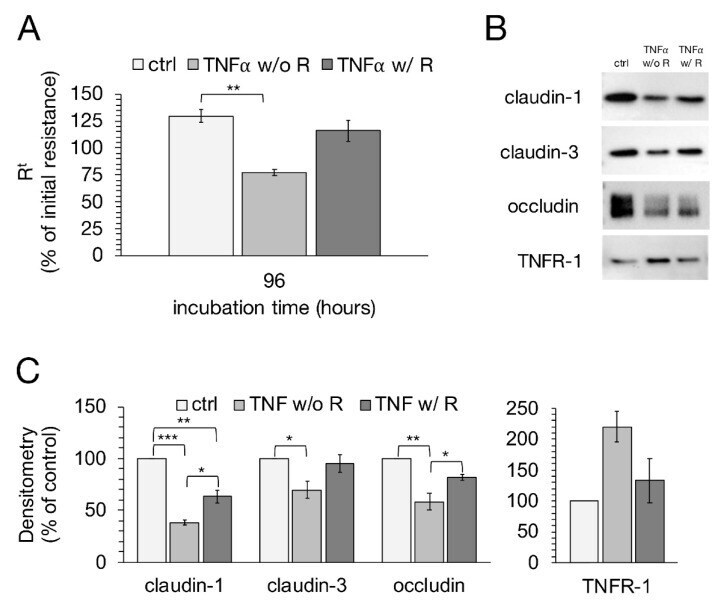
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- Fig. 6 Activated Ezrin inhibits SOCE by directly binding to Orai1 and promotes retraction of membrane blebs. a Representative still images of membrane blebbing in DLD1 cells expressing Lifeact-RFP and GFP-E-Syt1 from five independent experiments. See also Supplementary Movie 6 . b , c DLD1 cells expressing Lifeact-RFP with either GFP-E-Syt1 ( b ) or GCaMP6s ( c ) were treated with the actin polymerization inhibitor Latrunculin B (LatB, 10 uM). Arrowheads show disrupted actin cortex. Results shown are representative of three independent experiments. See also Supplementary Movie 7 . d - f Biochemical analyses of ezrin's role in STIM1-Orai1 complex formation. d Orai1-HA was co-expressed with either GFP or GFP-ezrin. Inputs and HA immunoprecipitates were immunoblotted with antibodies against HA and GFP. e STIM1-FLAG and Orai1-HA were co-expressed with either GFP or GFP-ezrin. Inputs and FLAG immunoprecipitates were immunoblotted with antibodies against FLAG, HA, and GFP. f GFP expression levels (shaded in green) and co-precipitated Orai1-HA (individual measurements) were quantified and normalized to GFP-expressing control. Data presented are means +- SD based on the values from four independent experiments. g , h Proximity ligation assay (PLA) for in situ detection of Orai1-STIM1 interaction. Representative images from five independent experiments are shown in g . h Quantification of PLA signals in expanding (low Lifeact intensity) and retracting (high Lifeact intensity) blebs ba
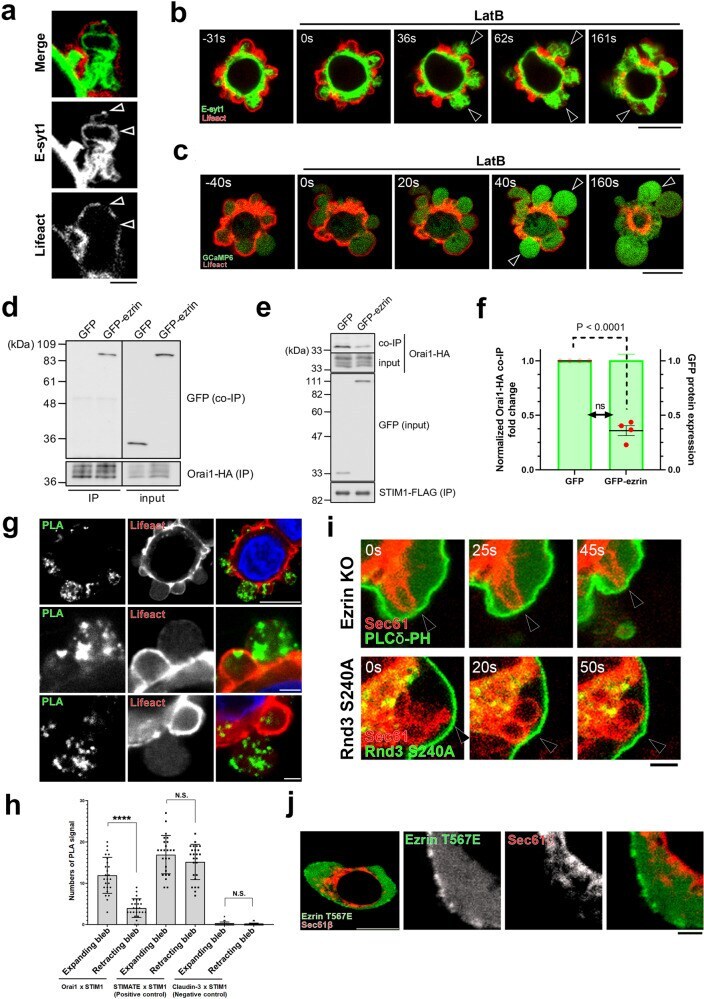
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- Figure 1. Claudin-3 protein levels in CRC samples and survival features of molecular subtypes substratified by CLDN3 expression levels. (A and B) Samples of adjacent normal tissue (N) and tumor tissue (T) were obtained and processed for further analysis of claudin-3 levels by western blotting (n=14). Results of four representative patients are shown. The line graph represents the increase (blue) or decrease (yellow) in the ratio of claudin-3 densitometry units normalized by endogenous protein control (GAPDH) in tumor and normal samples of each patient. Prognostic value of CLDN3 expression (upper and lower tertiles) in (C) unclassified data (n=414) and within (D) CMS1 (n=69), (E) CMS2 (n=250), (F) CMS3 (n=44), and (G) CMS4 (n=52). CRC, colorectal cancer; CLDN3 , gene encoding claudin-3.
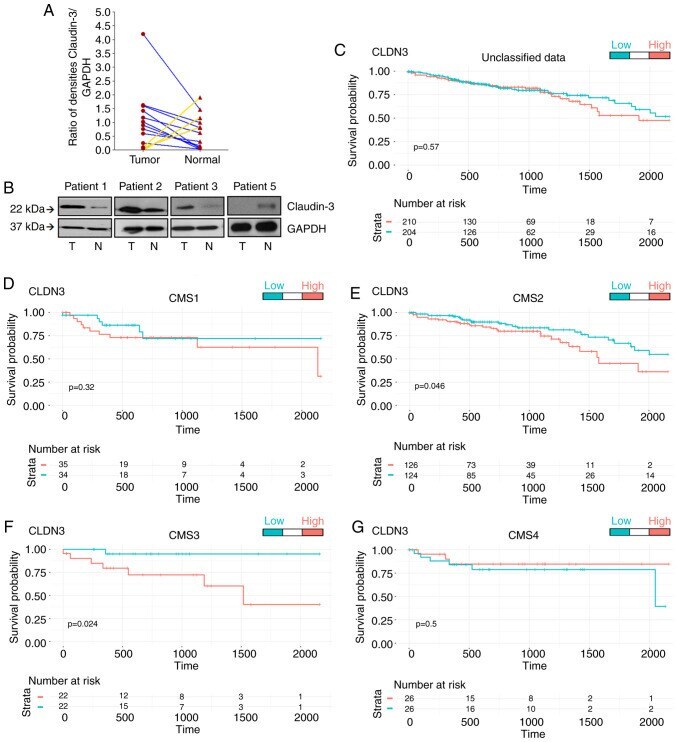
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
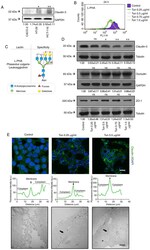
- Experimental details
- Figure 2. Effects of N -glycan biosynthesis inhibition on TJ stability. (A) Cell lysates from Caco-2, HT-29, and HCT-116 cells were analyzed by western blotting for claudin-3. (B) HCT-116 cells were treated with different concentrations of tunicamycin for 24 h. After treatment, the cells were incubated with FITC-conjugated lectin L-PHA and analyzed by flow cytometry. The histograms of fluorescence were generated by the Cell Quest software: control (purple); 0.25 ug/ml (green); 0.5 ug/ml (pink); 0.75 ug/ml (blue); and 1 ug/ml (orange). (C) Lectin L-PHA specificity. (D) Cell lysates were obtained after 24 h treatment with tunicamycin and analyzed by western blotting for claudin-3, occludin, and ZO-1. Tubulin was used as an endogenous protein control. (E, upper panel) Cell monolayers were fixed and stained for claudin-3 (green) and nuclei (blue) (DAPI). Representative images were obtained by confocal microscopy. The graphs represent the fluorescence intensity in cytosolic and membrane regions of neighboring cells. Scale bar, 10 um. (E, lower panel) Cells were cultured in filters of Transwell polycarbonate, and the functionality of TJs was analyzed by transmission electron microscopy (MET) using ruthenium red as a tracer. The images are representative of ultrathin sections of treated and control cells. Black arrows indicate the cell-cell contact. Scale bar, 2 um. The numerical values represent densitometric units +- standard error (n=3). ns, not significant (P>0.05); *P
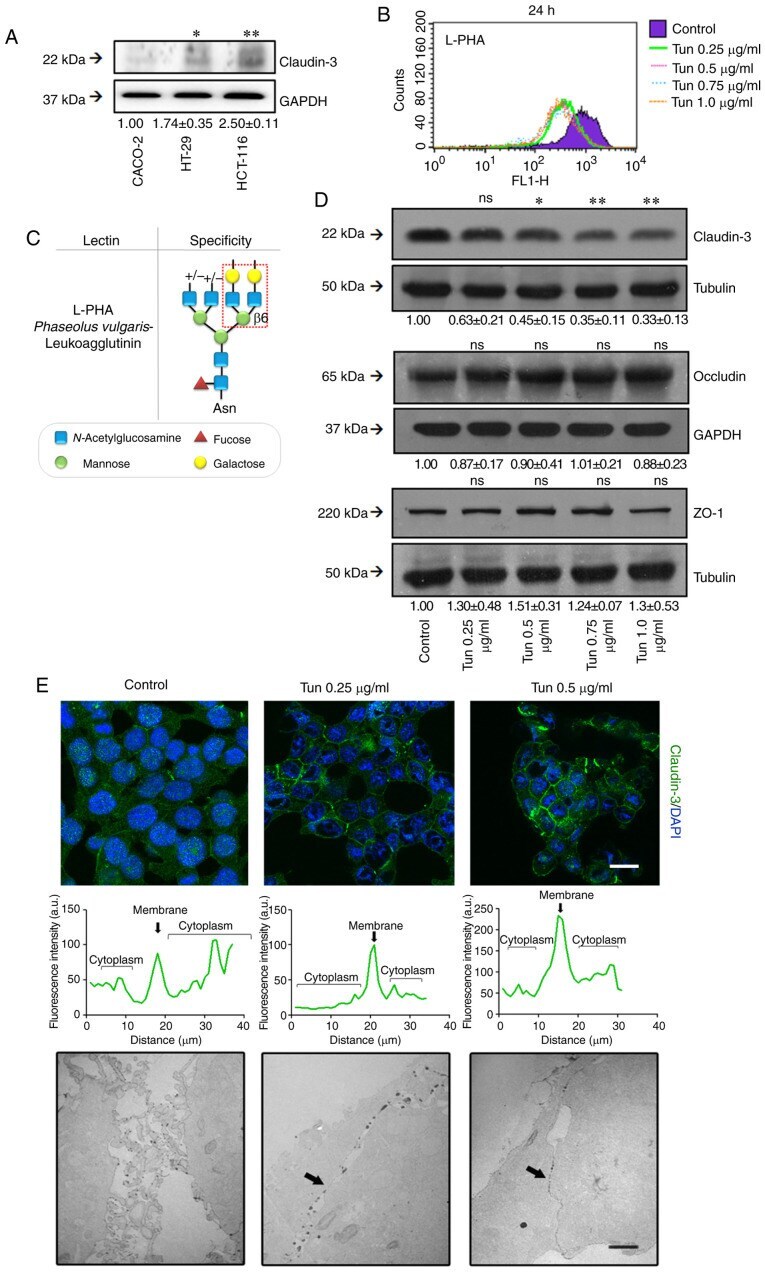
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- Figure 3. Effects of inhibition of N -glycan biosynthesis on RTK functionality. (A) After treatment with tunicamycin, cell lysates were obtained and analyzed by western blotting for p-EGFR, EGFR, p-IGF1R, and IGF1R. (B and C) Cells were treated with different concentrations of PD153035 or OSI906 for 24 h, and then levels of RTK phosphorylation and claudin-3 were assessed by western blotting. (D) Illustration showing RTK-related or non-RTK-related pathways, as well as on the relationship of this regulatory network with the regulation of CLDN3 expression. (E-H) Effects of specific inhibitors on claudin-3 levels. Cells were treated with different concentrations of (E) U73122 (an inhibitor of PLC-dependent processes), (F) PD98059 (a MEK1 inhibitor), (G) Ly294002 (a PI3K inhibitor), and (H) STA-21 (a STAT3 inhibitor) for 24 h, and then levels of claudin-3 were assessed by western blotting. (I) Cells were treated with paired combinations of inhibitors: PD98059 (21 ug/ml) and Ly294002 (6 ug/ml), or STA-21 (7.5 ug/ml) and U73122 (21 ug/ml). (J) Cells were treated with Forskolin or H-89 (a PKA inhibitor) for 24 h, and then levels of claudin-3 were assessed by western blotting. The numerical values represent densitometric units +- standard error (n=3). ns, not significant (P>0.05); *P
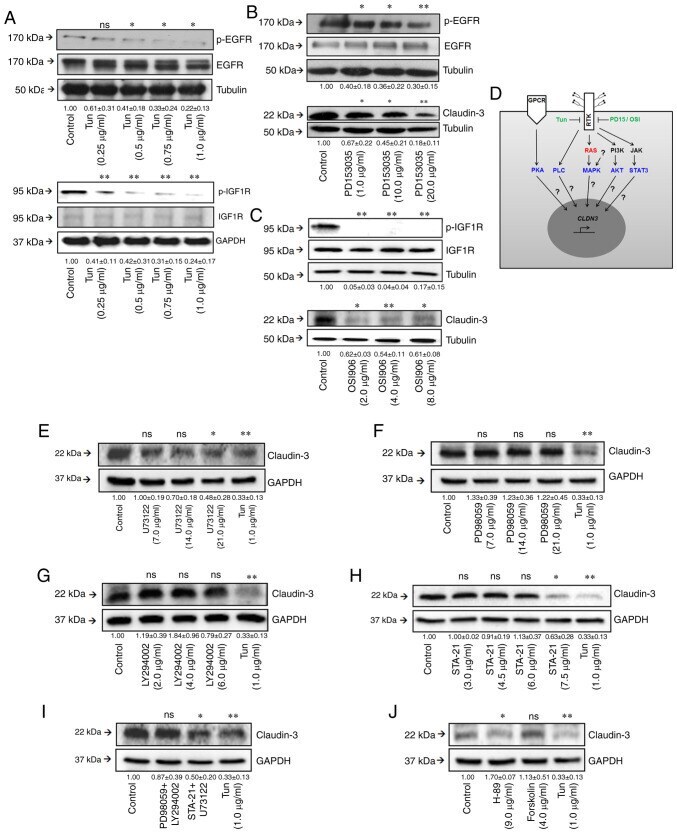
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
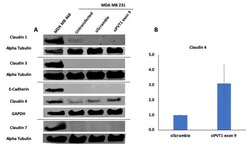
- Experimental details
- Figure 6 SiRNA targeting of PVT1 exon 9 induces claudin 4 protein re-expression in MDA MB 231 CL TNBC cells. ( A ) MDA MB 231 CL TNBC cells were transfected with PVT1 exon 9 specific siRNAs (siPVT1 exon 9) for 24 h. Western blotting was performed using specific antibodies against claudin 1, claudin 3, claudin 4, claudin 7 and E-Cadherin. SiRNA targeting of PVT1 exon 9 induced claudin 4 protein re-expression in MDA MB 231 CL TNBC cells in comparison to MDA MB 231 CL TNBC cells transfected with only control scramble non-targeting siRNA (siScramble). ( B ) Quantification of relative claudin 4 protein expression normalized to GAPDH protein expression; N = 2.
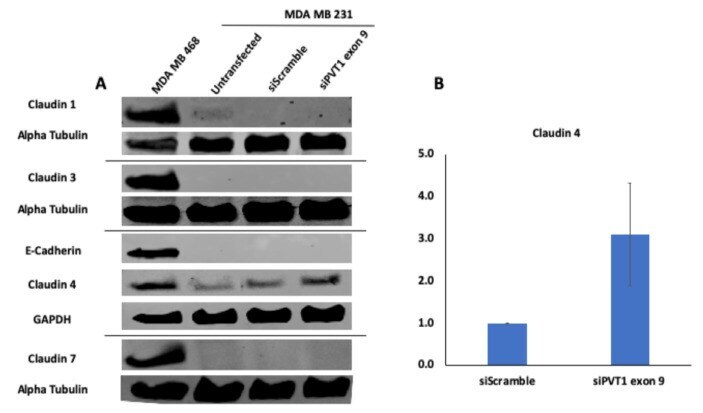
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- Figure 2 Edoxaban treatment stabilizes the blood-brain barrier and leads to anti-edematous effects in ischemic stroke. ( A ) Left : Representative corresponding coronal brain sections of vehicle (veh)-, edoxaban (Edox)- or phenprocoumon (Phen)-treated mice after the injection of the vascular tracer Evans Blue. Brains were analyzed at day 1 after transient middle cerebral artery occlusion (tMCAO) (Scale bar = 10 mm). Vascular leakage was decreased in the cortical and subcortical areas after edoxaban treatment. Right : Tissue concentration of Evans Blue in the ischemic (ipsilateral) and contralateral hemispheres of vehicle-, edoxaban-, or phenprocoumon-treated mice 24 h after tMCAO determined by photometry ( n = 5-8). ( B ) Edema formation as measured by brain water content in the ipsilateral and contralateral hemispheres of vehicle-, edoxaban-, or phenprocoumon-treated mice 24 h after tMCAO ( n = 4-6). ( C ) Top : Claudin (Cldn)-3,-5 and Occludin (Occl) protein expression in the ischemic cortices and basal ganglia of vehicle-, edoxaban-, or phenprocoumon-treated mice at day 1 after tMCAO as determined by immunoblot. Actin was used as loading control. Bottom : Densitometric quantification of Cldn-3, -5 and Occl immunoreactivity in the ipsilateral cortex and basal ganglia of vehicle-, edoxaban-, or phenprocoumon-treated mice ( n = 4). * p < 0.05.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- Figure 2 Localization of claudin-3 ( A --control, B --tumor) and claudin-4 ( C --control, D --tumor) in rat colon by immunofluorescence confocal microscopy (green); bar: 50 mum.
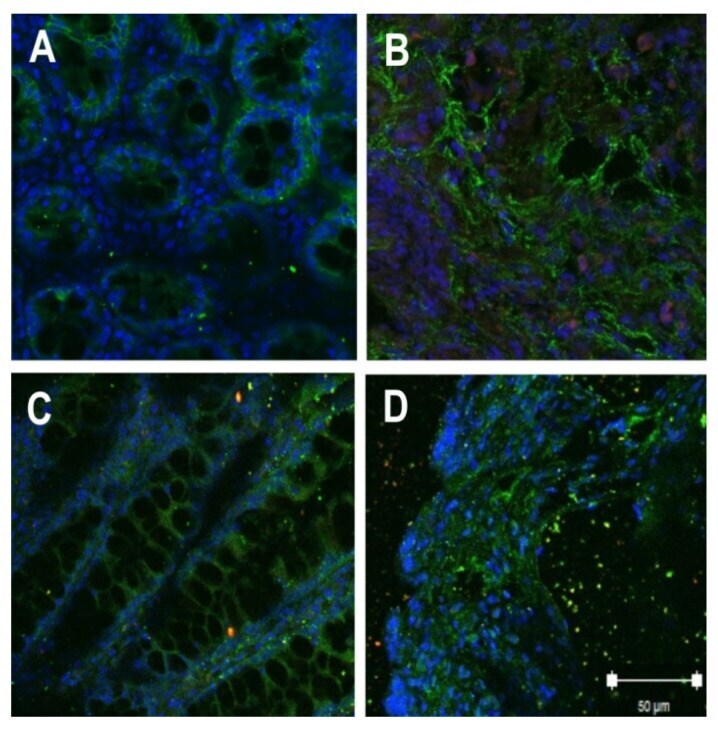
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- Fig. 1. A1AT C-terminal peptides induce TJ assembly. ( A ) Schema of recovery from the DSS-induced colitis model in mice. ( B ) Immunofluorescence of claudin-3 and DAPI in colon sections from mice treated as in (A). Scale bar, 20 mum. ( C ) Schema of the CCM treatment of A431 cells. ( D ) Immunofluorescence of claudin-1 and alpha-catenin in A431 cells treated for 3 hours with CCM from mice treated as in (A). Scale bar, 20 mum. ( E ) Quantification of relative claudin-1 intensity at cell-cell boundaries shown in (D) [ n = 5 images (ctrl and recover), n = 6 images (DSS) from two independent samples]. ( F ) Amino acid sequences of the JIP m35 , JIP m36 , and JIP m40 . ( G ) Western blot of CCM with anti-A1AT C terminus antibodies. Quantitative value of each band was presented. ( H ) Quantification of relative claudin-1 intensity at cell-cell boundaries of A431 cells treated with the indicated peptides (20 muM) or HBSS for 3 hours. See fig. S5B ( n = 5 images from two independent samples). ( I ) Relative barrier permeability of A431 cells ( n = 3 independent samples). ( J ) Immuno freeze-fracture replica electron microscopy of A431 cells treated with JIP m35 (20 muM) or HBSS and immunolabeled with anti-claudin-1 antibodies. NM, nuclear membrane; Cyt, cytoplasm; PF, P face. Scale bar, 500 nm. ( K ) Immunofluorescence of claudin-4 and Epsilon-cadherin in EpH4 cells treated with or without JIP m35 (70 muM) for 3 hours. Scale bars, 10 mum. ( L ) TER measurements of EpH4 cells treated
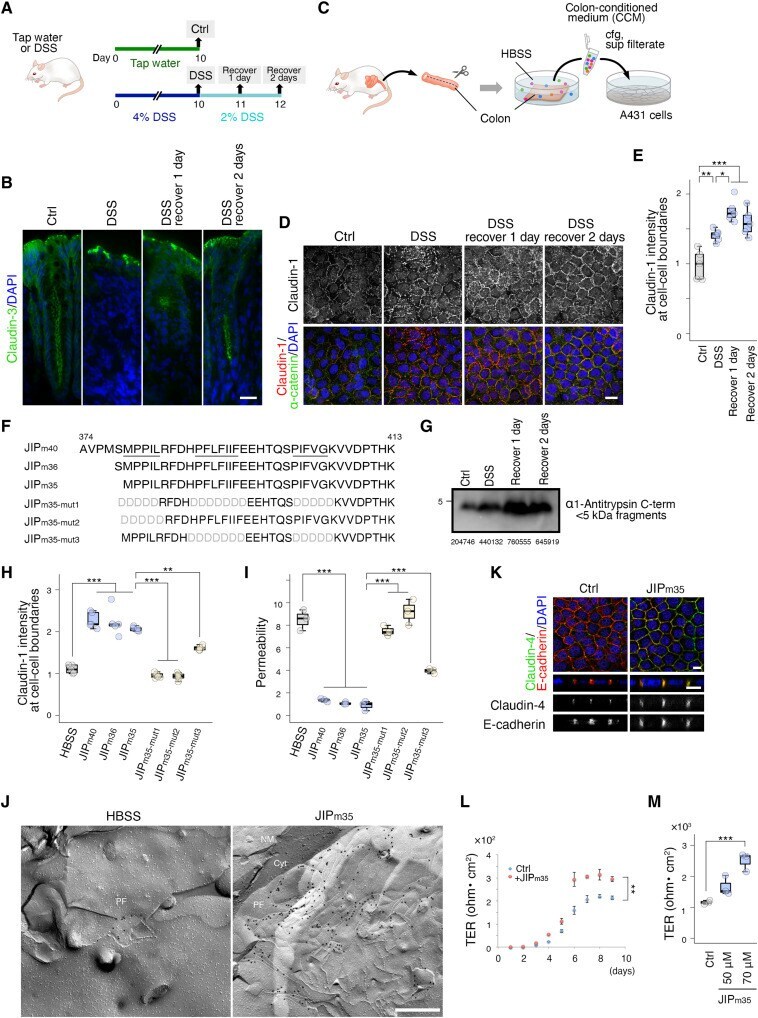
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- Fig. 4. Administration of JIPs restores TJs and ameliorates the symptoms of the DSS-induced colitis model in mice. ( A ) Schema of JIP m35 administration to DSS-induced colitis mice. ( B ) Immunofluorescence of claudin-3 and E-cadherin in colon sections from mice treated with DSS or tap water with 2.3 mg/kg per dose JIP m35 , JIP m35-mut1 , or the vehicle control for 10 days. Scale bar, 20 mum. Similar results were obtained in two independent experiments. ( C ) H&E staining of colon sections on day 10. Scale bar, 50 mum. Similar results were obtained in two independent experiments. ( D ) Relative intestinal permeability measured by plasma leakage of FITC-dextran (4 kDa) (ctrl + vehicle, DSS + JIP m35 ; n = 8 mice, ctrl + JIP m35 ; n = 7 mice, DSS + vehicle; n = 6 mice, DSS + JIP m35-mut1 ; n = 4 mice). ( E ) Immunofluorescence of JIP m35 and E-cadherin in colon sections from mice treated with DSS or tap water with 11.1 mg/kg JIP m35 or the vehicle control for 10 consecutive days. Scale bar, 20 mum. Similar results were obtained in two independent experiments. ( F ) DAI in accordance with assessment of stool consistency and fecal blood ( n = 3 mice). ( G ) Quantification of the numbers of Gr-1-positive cells in colon sections from mice treated as described in Fig. 4A ( n = 3 images from three mice). See fig. S11D. ( H ) Body weight changes in mice treated with DSS or tap water with daily administration of JIP m35 , JIP m35-mut1 , or the vehicle control for up to 25 consecutive
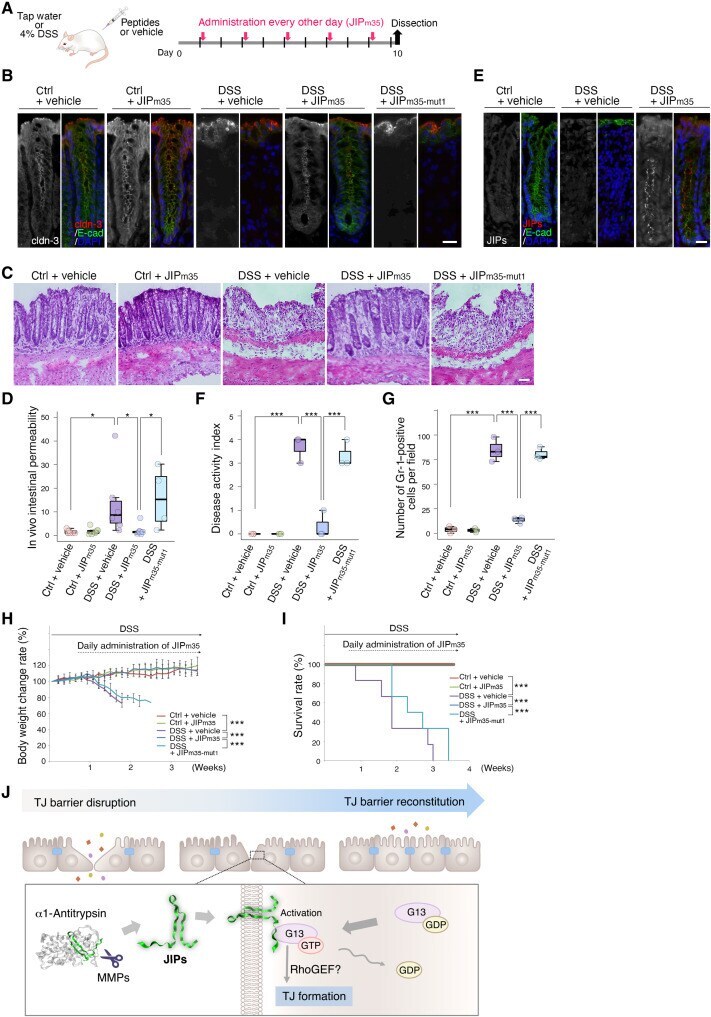
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- Fig. 5. Human A1AT C-terminal peptides induce TJ assembly in A431 cells and DSS-induced colitis mice. ( A ) Amino acid sequences of the human A1AT C terminus, JIP h42 /CAAP48, and mutant forms of JIP h42 /CAAP48 used in this study. ( B ) Immunofluorescence of claudin-1 and alpha-catenin in A431 cells treated with the indicated peptides at 20 muM or control HBSS for 3 hours. Scale bar, 20 mum. ( C ) Quantification of relative claudin-1 intensity at cell-cell boundaries of A431 cells shown in (B) ( n = 5 images from independent two samples). Similar results were obtained in two independent experiments. ( D ) Relative barrier permeability measured by paracellular tracer flux analysis using FITC-dextran (4 kDa) in A431 cells treated as in (B) and (C) ( n = 3 independent samples). ( E ) Measurement of G 13 activation in the presence of JIP h42 , JIP h42-mut1 , JIP h42-mut2, JIP h42-mut3 , and JIP h42-mut4 , which was assayed in GEF buffer with 5 pmol G 13 , 10 muM GTP, and 10 mM DTT. Synthetic peptides were prepared at the indicated concentrations ( n = 3 independent samples). ( F ) Schema of JIP h42 /CAAP48 administration to DSS-induced colitis mice. ( G ) Immunofluorescence of claudin-3 and E-cadherin in colon sections from mice treated with DSS or tap water with 1.5 mg/kg per dose JIP h42 /CAAP48 or vehicle control for 10 days. Scale bar, 20 mum ( H ) DAI in accordance with assessment of stool consistency and fecal blood ( n = 5 mice). ( I ) Immunofluorescence of Gr-1 in colon
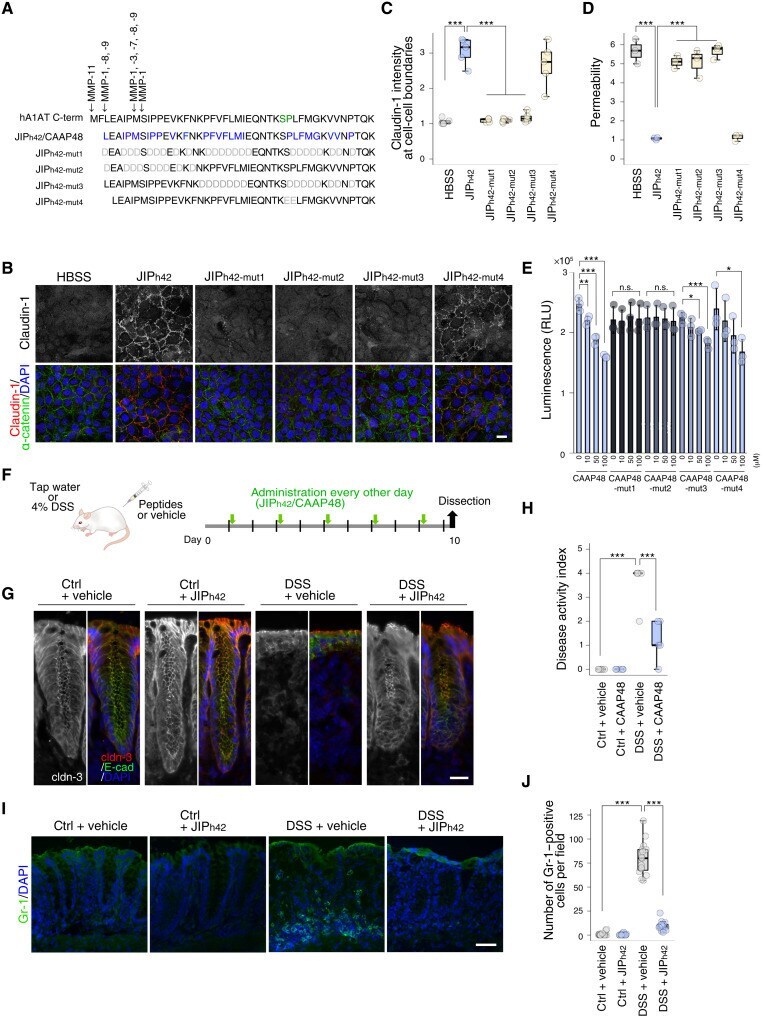
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
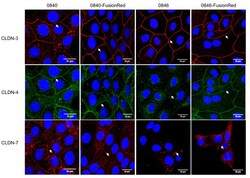
- Experimental details
- Figure 2 Expression of CLDNs protein in native and transfected tumor cell lines. Cells were subjected to immunostaining with corresponding antibodies using fluorescein-conjugated secondary antibodies, showing red signals for CLDN-3 and -7 and green signals for CLDN-4. DAPI was used for blue nuclei staining. Images were observed under confocal microscopy. Arrows indicate CLDN localization on cell-cell contact.
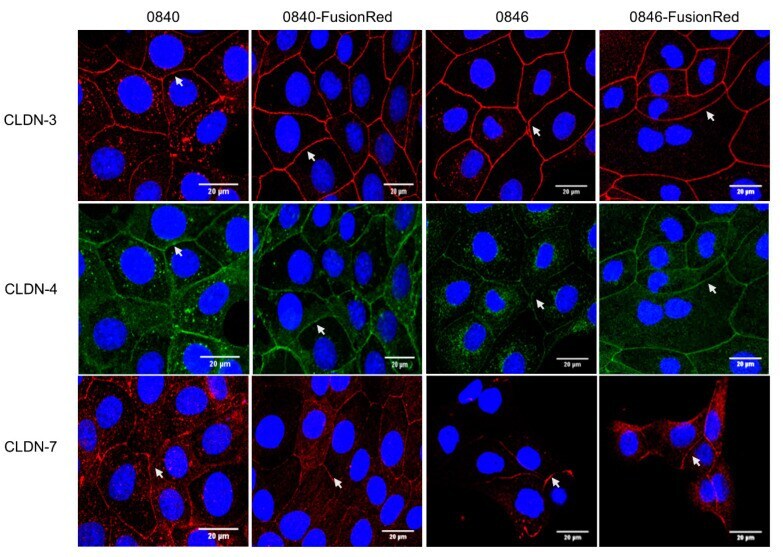
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
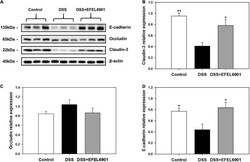
- Experimental details
- FIGURE 5 Effects of Limosilactobacillus reuteri EFEL6901 strain on tight junction protein levels in mouse colon. (A) Representative western blotting of E-cadherin, occludin, claudin-3, and beta-actin (loading control). Quantification of E-cadherin (B) , occludin (C) , and claudin-3 (D) protein expression levels in the colon. Results are expressed as means +- standard error ( n = 7). Significant differences are presented with the DSS group: * p < 0.05; ** p < 0.01.
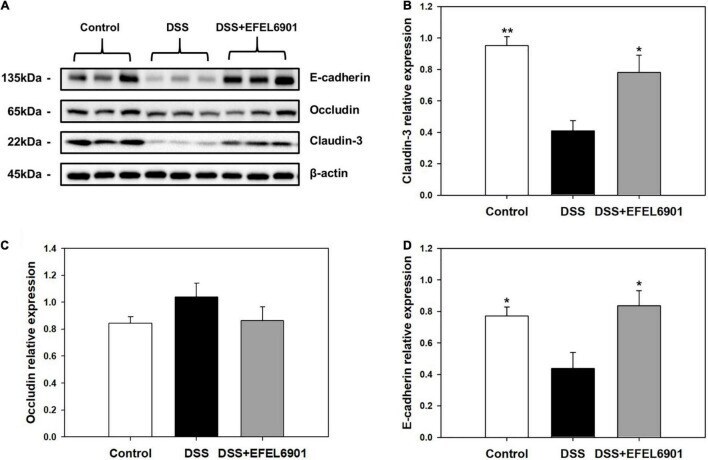
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
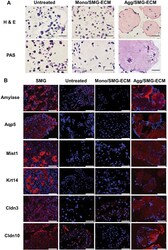
- Experimental details
- Histological analysis of SMG-ECM-treated cultures reveals that aggregates display an expression pattern different from monolayer cells. A Paraffin sections of cells from 14 day cultures (i.e., untreated BM-MSCs [Untreated], SMG-ECM-treated aggregates [Agg/SMG-ECM], and monolayer [Mono/SMG-ECM] cells) were prepared and stained with H&E or PAS. PAS-positive cells are identified with arrows in the figure. Bar: 50 um. B Immunofluorescence staining of cells prepared as in A and rat SMG tissue (positive control) was used to evaluate the presence and localization of amylase, Aqp5, Mist1, Krt14, Cldn3, and Cldn10 protein. Staining with nonspecific isotype antibody was used as a negative control (not shown). Bar: 50 um
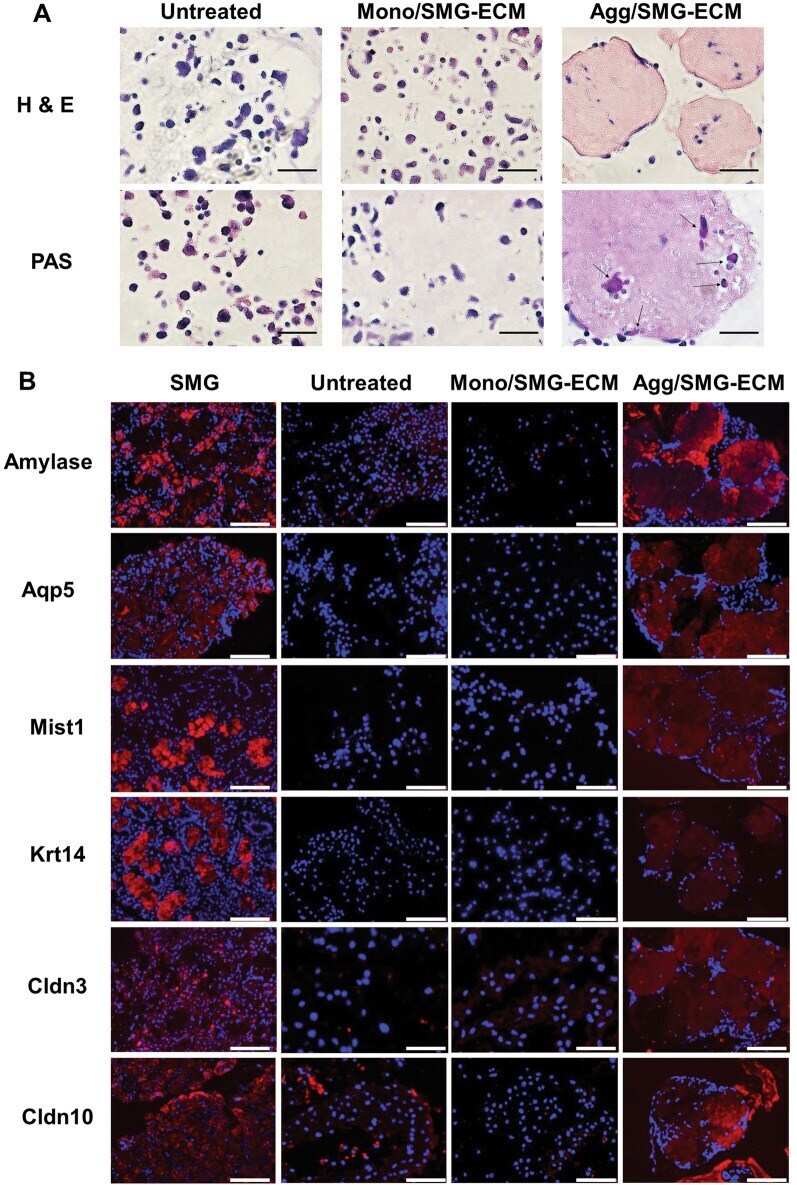
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- 2 FIGURE Validation of fluorophore conjugated primary antibodies on LNCaP cells: LNCaP cells were fixed, permeabilized, incubated with 1:100 of each antibody, and imaged with a confocal microscope. Fluorescent signal from primary antibody is shown in first column, nuclear 4',6-diamidino-2-phenylindole (DAPI) stain in second column and merged image in third column. Arrows point to accurate cellular locations of specific fluorescent signal acquired from each antibody staining. Antibodies conjugated to Alexa Fluor 488 (AF488) and Alexa Fluor 647 (AF647) are shown in green and red respectively, DAPI signal is shown in blue. CD9 stains cell membrane, fatty acid synthase (FASN) is found in cytoplasm, fibrinogen is not expressed by LNCaP cells, phospho-histone H2A.X (gammaH2AX) is visualized as nuclear puncta, phosphoglycerate kinase 1 (PGK1) stains the cytoplasm, and claudin 3 (CLDN3) stain is localized to the membrane (scale bars are 20 um).
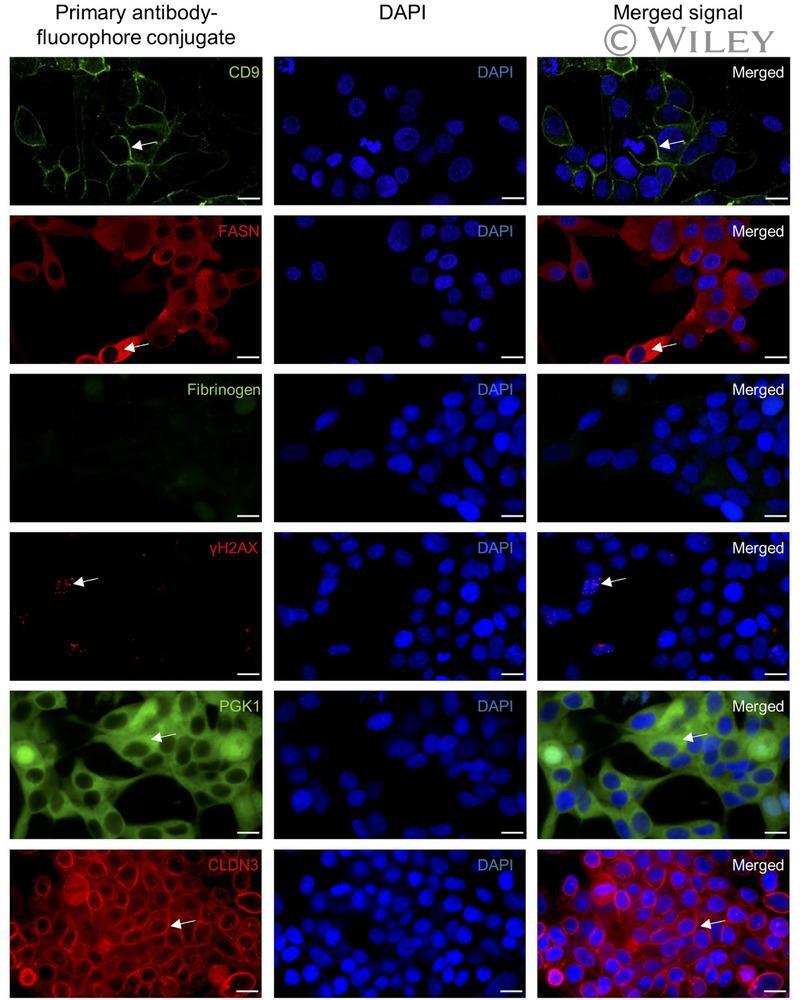
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- Claudins form characteristic copolymers including segregation of channel-forming claudins from Cldn3. a Scheme is illustrating predicted claudin-claudin organization patterns. Two claudins (yellow and magenta) that are compatible with each other tightly colocalize in the TJ (pink). Incompatible claudins separate into different strands or even larger claudin-specific parts. This separation might facilitate permeability of specific ions (cyan) over the TJ meshwork. b Representative STED images of TJ-like meshwork of the observed five organization patterns formed by the indicated co-expressed claudins (SNAP-tagged (yellow; BG-Atto590) and YFP-tagged (magenta; alpha-GFP-NB-Atto647N)) in fixed COS-7 cells are shown. Tight colocalization: intermixing of mesh-forming claudins, integration of non-mesh-forming with a mesh-forming claudin or induction (de novo mesh-forming of co-expressed non-mesh-forming claudins). Separated claudins: segregation and exclusion. c Pearson correlation analysis of Cldn3 co-expressed with barrier-forming (gray) or channel-forming (magenta) claudins. Co-expression of Cldn3 with Cldn3 (yellow) served as positive control. Data represent the mean +- SD. Every n represents the Pearson of one TJ-like meshwork. n (Cldn3 + Cldn3) = 55; n (Cldn3 + Cldn1) = 54; n (Cldn3 + Cldn5) = 20; n (Cldn3 + Cldn6) = 20; n (Cldn3 + Cldn7) = 17; n (Cldn3 + Cldn9) = 15; n (Cldn3 + Cldn14) = 19; n (Cldn3 + Cldn19b) = 25; n (Cldn3 + Cldn2) = 36; n (Cldn3 + Cldn10a) = 30; n (Cldn3 +
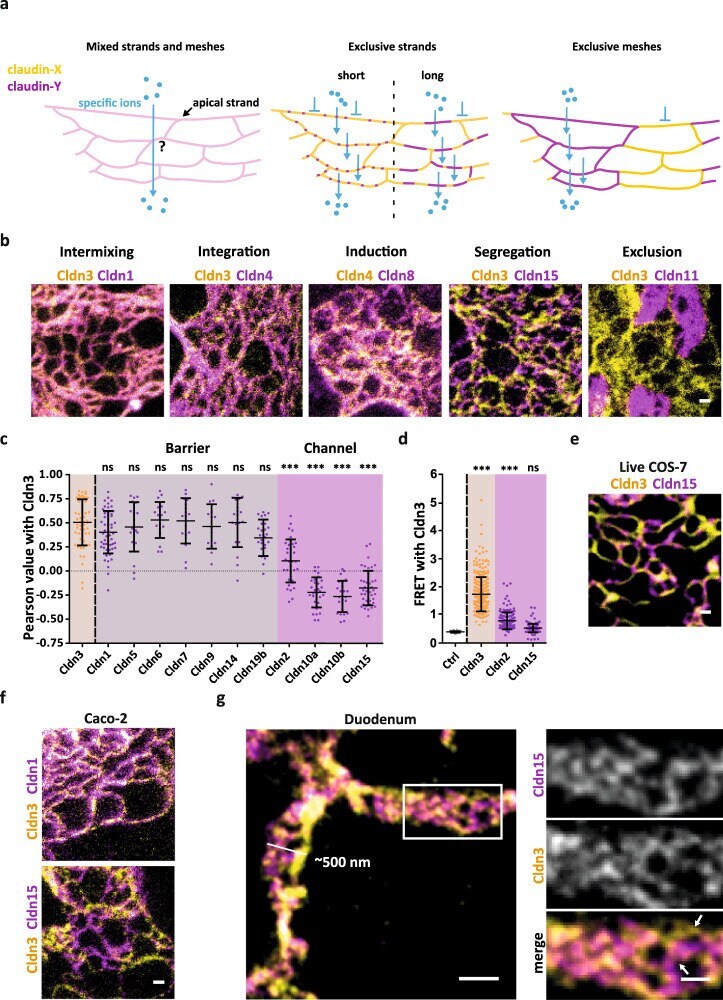
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
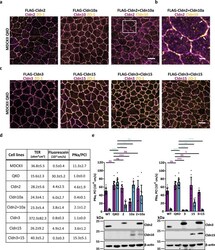
- Experimental details
- Segregation enables paracellular ion flux over the TJ meshwork. a Representative confocal images of MDCKII QKO cells stably expressing (sT) FLAG-Cldn2, FLAG-Cldn10a, and FLAG-Cldn2+FLAG-Cldn10a. Single claudin expressing cells were immunostained with anti-Cldn2 (magenta; 2 nd -Atto647N) or anti-Cldn10 (magenta; 2 nd -Atto647N) and anti-ZO-1 (yellow; 2 nd -AF594) antibody. Double claudin expressing cells were immunostained with anti-Cldn2 (magenta; 2 nd -Atto647N) and anti-Cldn10 (yellow; 2 nd -AF594). The white rectangle indicates the region of interest for the magnification in ( b ). b Magnification of FLAG-Cldn2+FLAG-Cldn10a expressing cells from ( a ). White arrows point out differently sized TJ parts that contain only Cldn10a. c Representative confocal images of MDCKII QKO sT FLAG-Cldn3, FLAG-Cldn15 and FLAG-Cldn3+FLAG-Cldn15. Single and double claudin expressing cells were stained with anti-Cldn3 (magenta; 2 nd -Atto647N) or anti-Cldn15 (magenta; 2 nd -Atto647N) and anti-ZO-1 (yellow; 2 nd -AF594) antibody. d Summary of the TER (ohm*cm 2 ), fluorescein flux (10 -6 cm/s) and PNa/PCl ratio from the measured cell lines: MDCKII, MDCKII QKO and MDCKII QKO sT FLAG-tagged Cldn2, Cldn3, Cldn10a, Cldn15 and MDCKII QKO sT FLAG-tagged Cldn2 + Cldn10a and FLAG-tagged Cldn3 + Cldn15. Data represent the mean +- SD from 3 to 5 independent experiments. Single data points are shown in Supplementary Fig. 12 . e Absolute permeability for sodium (PNa; magenta bars) and chloride (PCl; cyan b
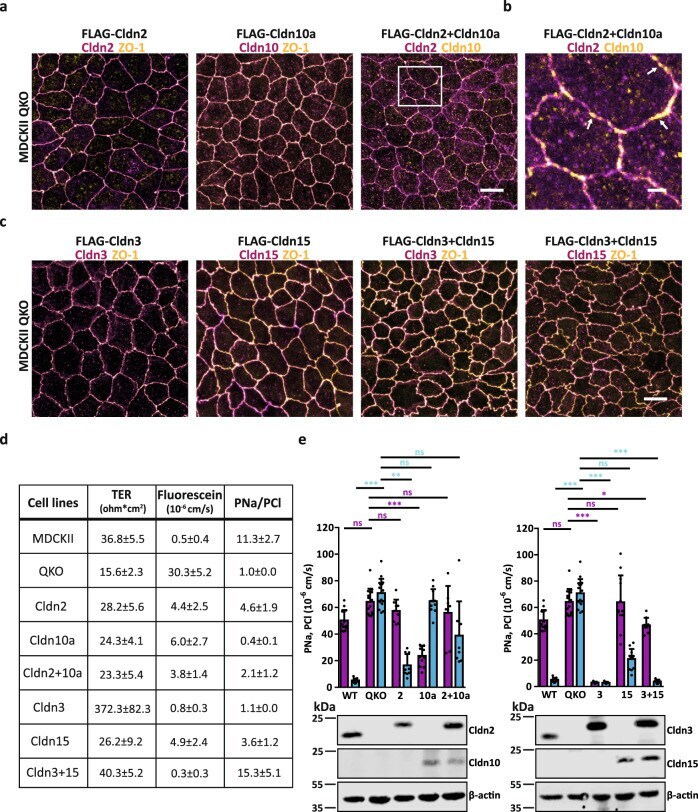
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- Centella asiatica -derived endothelial epidermal growth factor mitigates radiation-induced enteritis with epithelial barrier restoration in mouse model. Mouse groups are as follows: control (Con), irradiated (IR), irradiated mouse administrated with the conditioned media (CM) of irradiated HUVECs (IR + CM), CM of Centella asiatica (CA)-treated irradiated HUVECs (IR + CA-CM), and recombinant EGF (IR + rEGF). ( A ) Hematoxylin and eosin staining of mouse intestinal tissue sections was performed in each group. ( B ) Proliferation was assessed by staining intestinal sections with ki-67. ( C ) The lengths of villi and crypts in the intestinal sections were quantified. Histological scoring was assessed based on the degree of epithelial architecture maintenance, crypt disruption, vascular enlargement, and infiltration of inflammatory cells in the lamina propria of the ileum of Con, IR, IR + CM, IR + CA-CM, and IR + rEGF groups (0 = none, 1 = mild, 2 = moderate, and 3 = high). ( D ) Body weights of Con, IR, IR + CM, IR + CA-CM, and IR + rEGF groups were determined. ( E ) Immunohistochemistry against epithelial barrier-related molecules, e.g., villin, zonula occuludens 1 (Zo1), desmoglein 2 (Dsg2), and claudin 3 (Cldn3), in the Con, IR, IR + CM, IR + CA-CM, and IR + rEGF groups was performed. ( F ) mRNA levels of the epithelial barrier-related molecules in Con, IR, IR + CM, IR + CA-CM, and IR + rEGF groups were assessed by qRT-PCR. Data are presented as the mean +- standard error of t
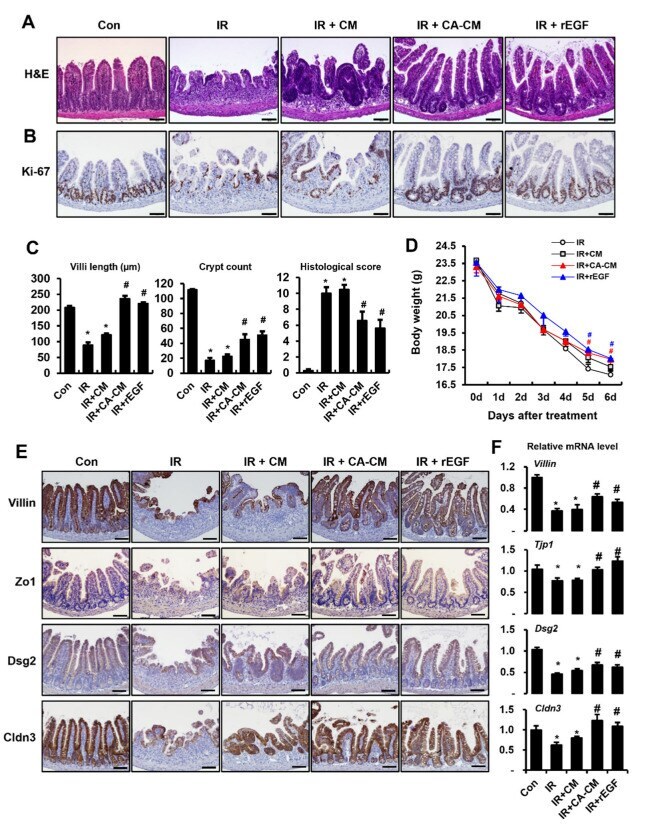
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
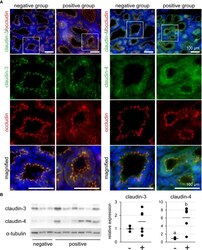
- Experimental details
- Figure 5 (A) Representative images of immunofluorescence against claudin-3 and claudin-4 in tertiary lymphoid organ-positive and -negative goat mammary gland tissues. Occludin is a marker for tight junctions. Scale bar, 100 mum. (B) Bands detected by western blot and the results of densitometry analyses. Alpha-tubulin serves as internal control; (-) lymphocyte aggregation-negative group; (+) lymphocyte aggregation-positive group. Different letters indicate a significant difference between groups (p < 0.05).
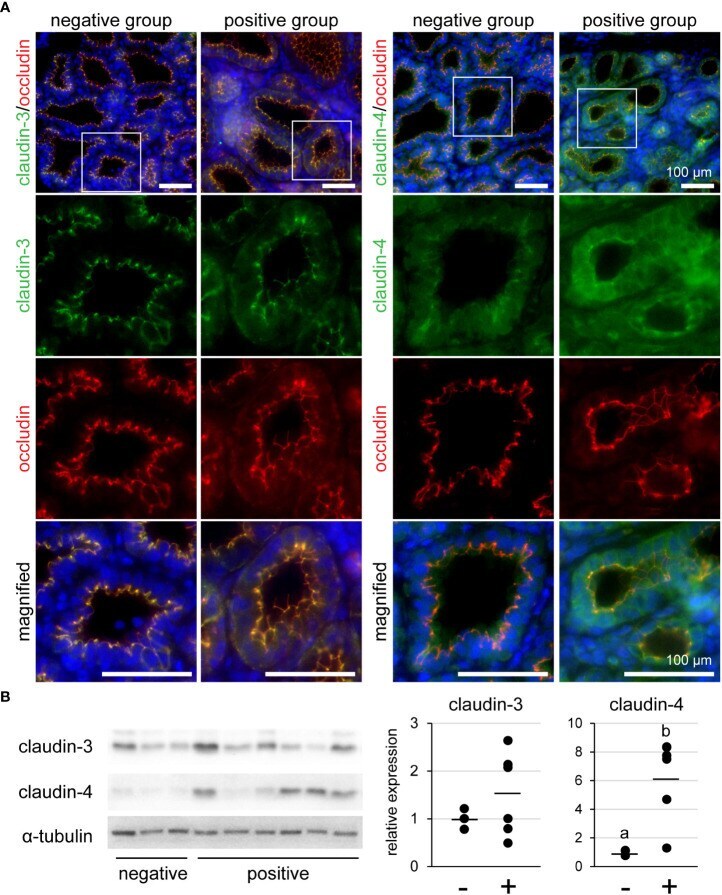
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
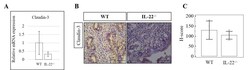
- Experimental details
- Supplementary Figure 4 The expression of claudin-3 in IL-22 -/- and WT mice. (A) IL22 -/- , n=6 and WT, n=5 mice uterine samples were harvested 48 hours post intraperitoneal LPS injection. Total mRNA samples were separated and processed by qRT-PCR to evaluate the expression of claudin-3. The Ct values were normalized against the Ct values for GAPDH from the same preparation. The data are provided as mean +- SEM from 3 independent experiments. (B) IL-22 -/- , n=6 and WT, n=5 mice uterine samples (half of one horn) were harvested 48 hours post intraperitoneal LPS injection, frozen fixed, and processed for IHC analysis. (C) H-score of claudin-3 was calculated to measure whether the difference of the staining between the groups is statistically significant. The data from t-test are provided as mean +- SEM. WT, wild type, IL-22 -/- , IL-22 knock out, H-score, histological score. Original magnification 40x, scale bar 100 um. Click here for additional data file.
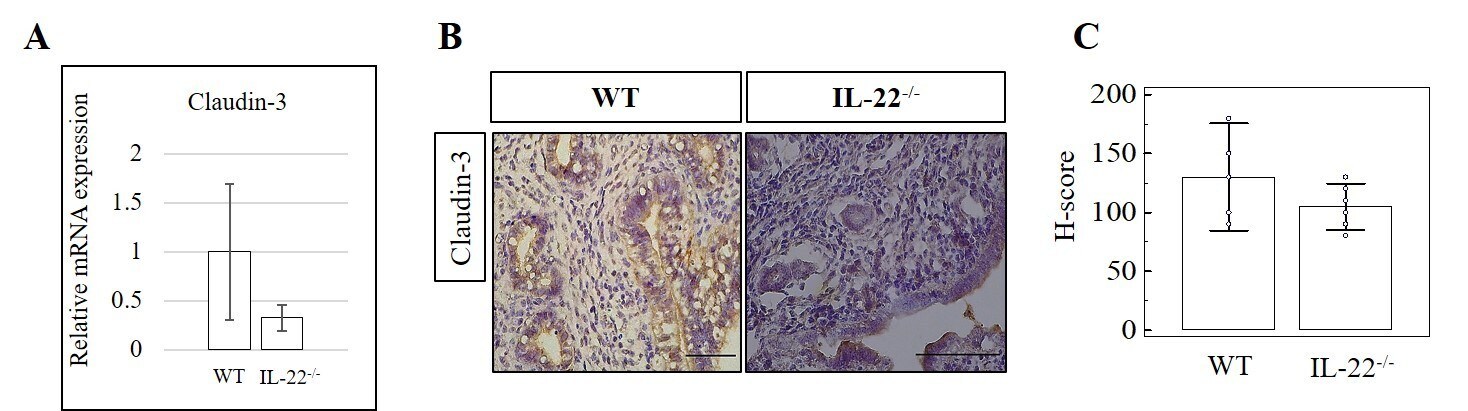
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- FIGURE 2: The localization of some, but not all, TJ proteins to the AJC is attenuated in ZO dKD cell lines. (A-C) A 1-mum-thick, maximum-density projection of the apical domain, en face, acquired at the AJC (see Materials and Methods ). (A) Occludin and cingulin localization at the AJC is reduced, whereas JAM-A and tricellulin appear relatively normal in dKD cells relative to control cells. Claudin-4 staining (green) is included with tricellulin staining (red) to highlight tricellular junctions. (B) Claudin-1 and -2 staining at the AJC is reduced, whereas claudin-3 and -4 appear relatively normal in dKD cells. Note again the dramatic change in cell shape at the AJC in dKD cells. Scale bar: 10 mum.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- Figure 2 Establishment of claudin-1 (CLDN-1) knockout clones in Caco-2 cells. ( A ) Genomic DNA isolated from wild-type Caco-2 cells or CLDN-1 knockout clones was subjected to PCR for amplification of the CRISPR-Cas9 targeting site in the cldn-1 gene. After purifying and annealing the PCR products, the products were incubated with T7 endonuclease I, which recognizes and cleaves mismatched DNA. Two shorter bands of the predicted size were generated in some clones, indicating that CRISPR-Cas9 had successfully introduced indel mutations at the targeted cldn-1 site. This analysis indicated that the Ko-C1 clone contains only WT alleles, but the Ko-T1 and Ko-C12 clones had deletion alleles. ( B ) Immunoblots comparing CLDN-1 and CLDN-4 production by control Caco-2 cells and CLDN-1 knockout clones. These results indicated that the Ko-C12 clone completely lost the ability to produce CLDN-1, so this clone was selected for further experiments. ( C ) Immunoblot analysis comparing production of other tight junction proteins, including claudin-3 (CLDN-3), claudin-4 (CLDN-4), occludin and ZO-1, by CLDN-1 mutant cells vs. wild-type Caco-2 cells. GAPDH and E-cadherin were used as endogenous controls. For these blots, target proteins in cell lysates (30 ug protein/sample) were detected by Western blotting using specific primary antibodies and suitable horseradish peroxidase-labeled secondary antibodies. Results shown are representative of three repetitions.
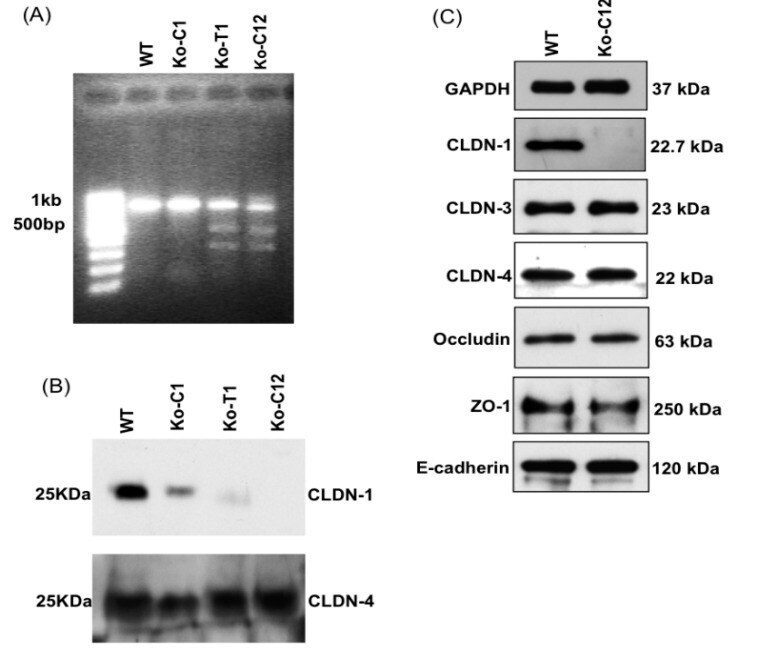
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
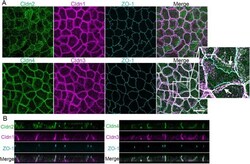
- Experimental details
- FIGURE 2: Endogenous clds are variably distributed between the apical tight junction and lateral membrane in MDCK II cells. (A) Top, Maximum-intensity projections of immunofluorescence confocal colocalization of cldn2, cldn1, ZO-1, and merge (left to right panels) show that cldn1 is more laterally disposed than cldn2 or ZO-1. Bottom, Colocalization of cldn4, cldn3, ZO-1, and merge (left to right panels) reveals cldn4 and cldn3 at the lateral membrane as well as colocalized with ZO-1. Arrows in enlargement show apparent concentrations of lateral cldns likely related to lateral membrane infoldings (see Supplemental Figure S1). (B) z -Axis images as in A show ZO-1 focused at the apical junctional region and cldns colocalized with ZO-1 but also distributed along the lateral membrane.
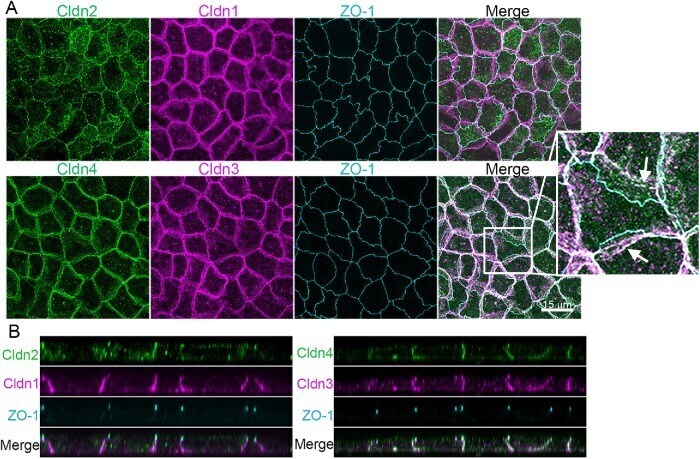
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
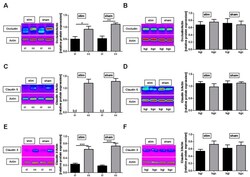
- Experimental details
- Figure 2 Stimulation of the mesencephalic locomotor region does not modulate photothrombosis-driven tight junction protein degradation. ( A , B ), Representative anti-occludin Western blot analysis (cc, cortex contralesional; ci, cortex ipsilesional; bgc, basal ganglia contralesional; bgi, basal ganglia ipsilesional) and densitometric quantification of occludin protein expression in the basal ganglial as well as cortical regions 27 h after photothrombosis of unstimulated (sham) and high-frequency stimulated (stim) rats ( n = 5-7/group). One-way ANOVA. * p < 0.05; ** p < 0.01. ( C , D ), Representative anti-claudin 5 Western blot analysis (cc, cortex contralesional; ci, cortex ipsilesional; bgc, basal ganglia contralesional; bgi, basal ganglia ipsilesional; bd, beyond detection limit) and densitometric quantification of claudin 5 protein expression in the basal ganglial as well as cortical regions 27 h after photothrombosis within the two treatment groups ( n = 5-7/group). One-way ANOVA. ( E , F ), Representative anti-claudin 3 Western blot analysis (cc, cortex contralesional; ci, cortex ipsilesional; bgc, basal ganglia contralesional; bgi, basal ganglia ipsilesional; bd, beyond detection limit) and densitometric quantification of claudin 3 protein expression in the basal ganglial as well as cortical regions 27 h after photothrombosis within the two treatment groups ( n = 5-7/group). One-way ANOVA. *** p < 0.001.
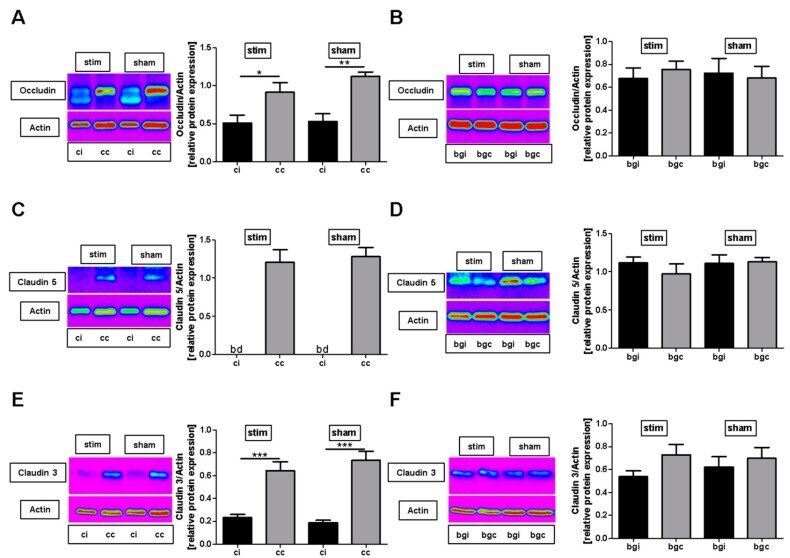
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- Figure 9 Effects of deoxynivalenol (DON), T-2 toxin, and zearalenone (ZEN) exposure on ( A ) occludin (OCLD) and ( B ) claudin (CLDN)-4 localization in BeWo cells, visualized by immunofluorescence staining (400x magnification). Scale bars represent 50 mum.
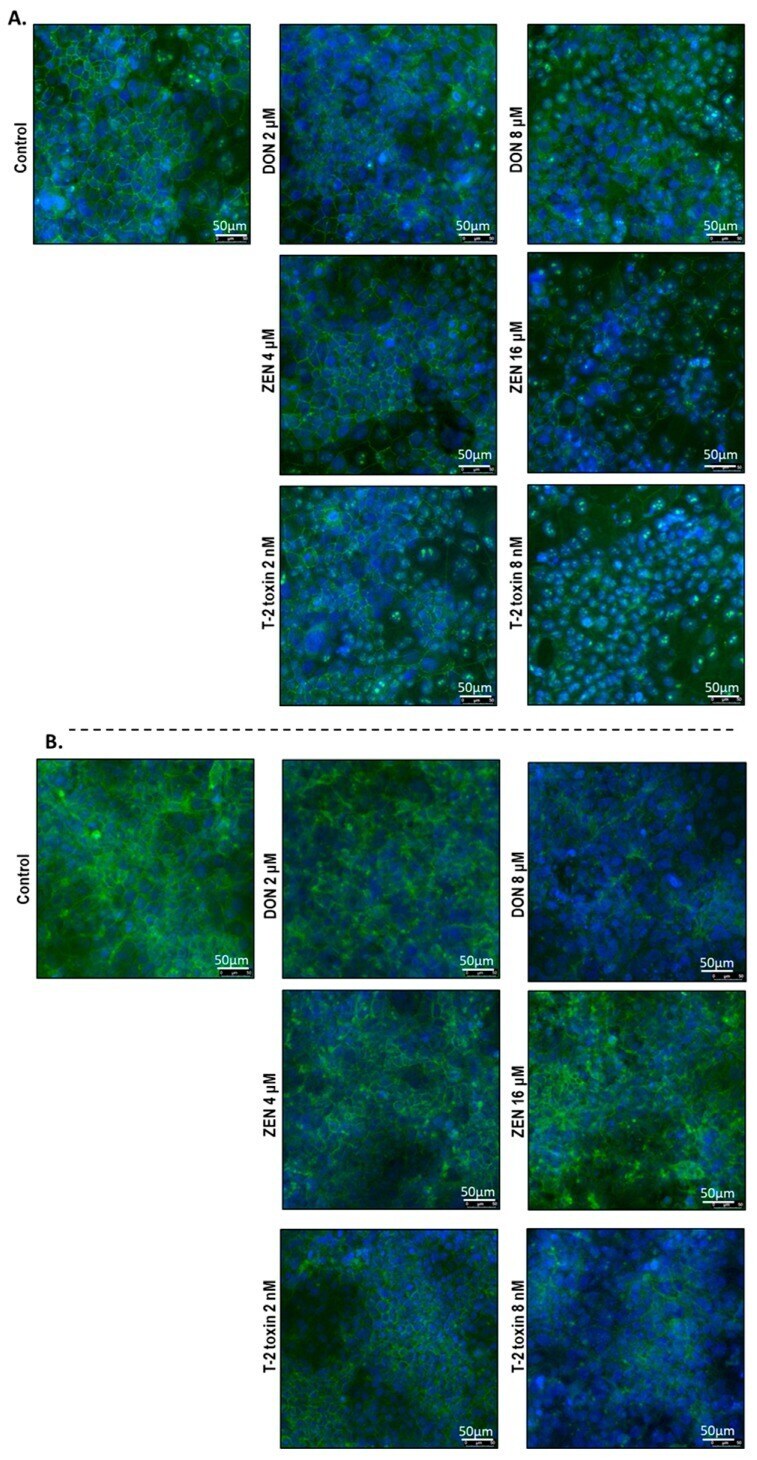
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
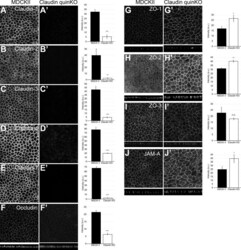
- Experimental details
- Figure 2. Localization of TJ proteins in claudin quinKO cells. (A-J) Immunofluorescence analyses of parental MDCK II cells (A-J) and claudin quinKO cells (A'-J'). (A-E) Claudin-1 (A), claudin-2 (B), claudin-3 (C), claudin-4 (D), and claudin-7 (E) expression was abolished in claudin quinKO cells. (F) Occludin localization to apical junctions was reduced. (G-I) ZO-1 (G) and ZO-2 (H) were more concentrated at apical junctions, while ZO-3 (I) localization was not altered in claudin quinKO cells. (J) JAM-A was more concentrated at apical junctions in claudin quinKO cells. Graphs are quantitation of the fluorescence intensity and represent mean +- SD ( n = 3 each). *, P < 0.05; **, P < 0.005; ***, P < 0.0005, compared by t test. Scale bar: 20 um. n.s., not significant.
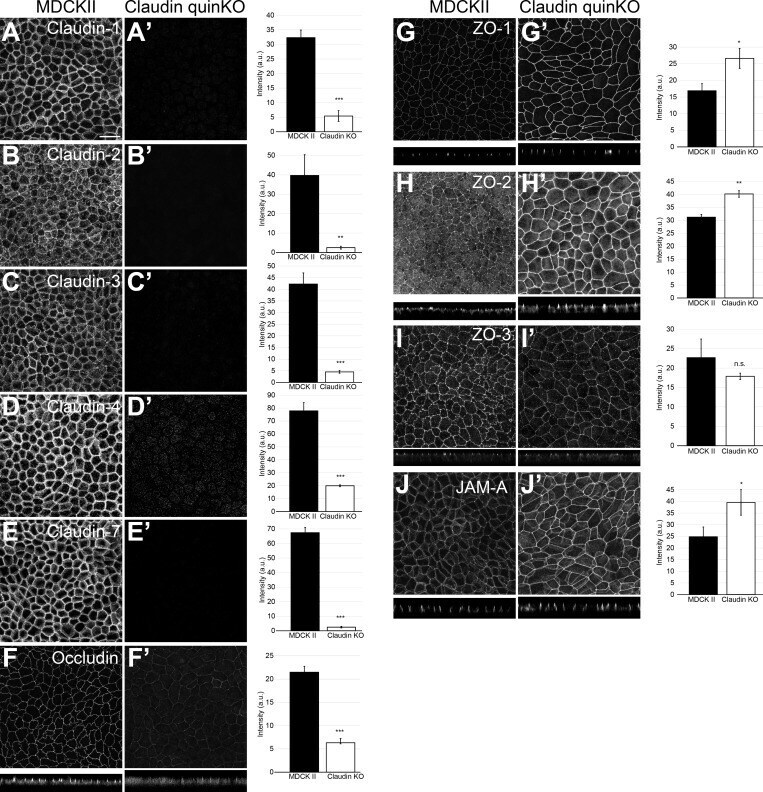
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
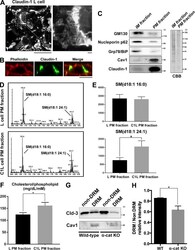
- Experimental details
- Figure 3. Cholesterol is enriched in the TJ-containing PM fraction. (A) C1L cells were fixed and stained with an anti-claudin-1 pAb. (B) C1L cells were fixed and stained with an anti-claudin-1 pAb (green) and phalloidin (red). Bars, 10 um. (C) Immunoblot analysis of the PM and IM fractions of C1L cells. Each membrane fraction (5 ug) was separated by SDS-PAGE, transferred to a nitrocellulose membrane, and probed with antibodies against the indicated marker proteins (left). Coomassie brilliant blue (CBB) staining is shown on the right. (D) Positive ion mass spectra of SM species in the PM fractions of L and C1L cells. The SM molecular species corresponding with each peak are indicated. The x and y axes show the total carbon chain length and the number of carbon-carbon double bonds of individual lipid molecular species, respectively. (E) Quantification of the indicated SM species in the PM fractions of L cells and C1L cells. (F) Quantification of the cholesterol-to-phospholipid ratio in the PM fractions of L and C1L cells. (G) Immunoblot analysis of the DRM and non-DRM fractions of WT and alpha-catenin-KO EpH4 cells using pAbs against the DRM marker proteins claudin-3 and caveolin-1. Results in C, D, and G are representative of three independent experiments. (H) Quantification of the ratio of the claudin-3 level in the DRM fraction to that in the non-DRM fraction in WT and alpha-catenin-KO EpH4 cells. Error bars show SD calculated based on three independent experi
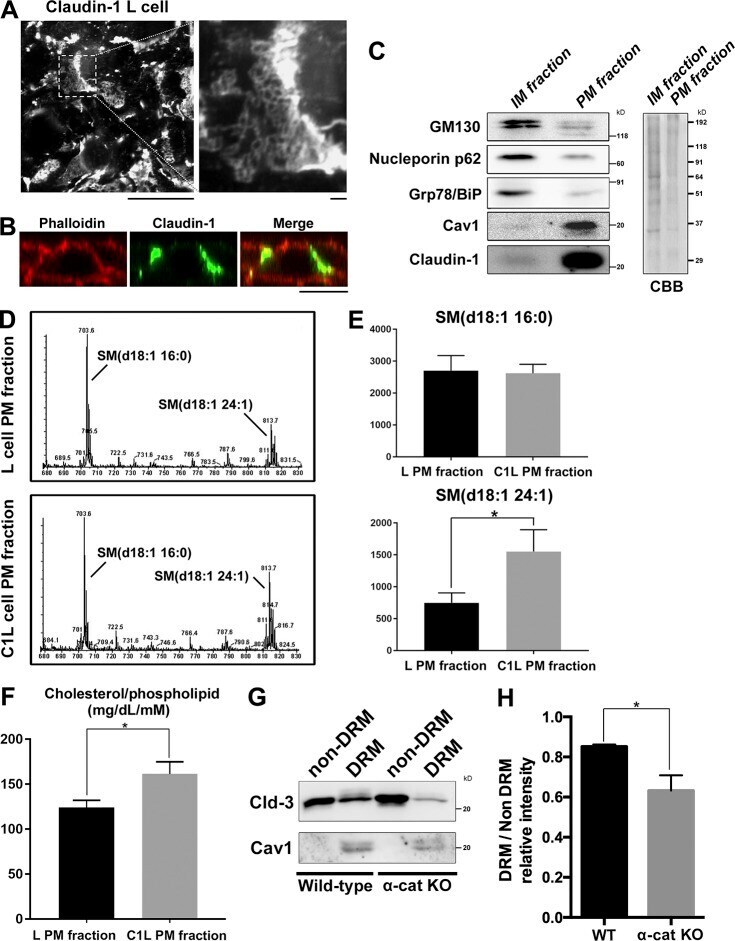
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
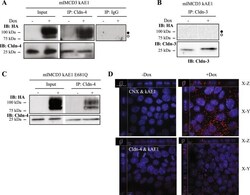
- Experimental details
- Figure 4 kAE1 protein is in close proximity and interacts with claudin-4. ( A ) Confluent mIMCD3 cells stably expressing kAE1 protein in an inducible manner were grown for 10 days in a 10 cm dish, lysed (Input) and claudin-4 was immunoprecipitated using a rabbit anti-claudin-4 antibody (IP lanes). As a control an irrelevant IgG antibody was used (right panel). Eluted proteins were separated on SDS-PAGE gel prior to immunoblotting with a mouse anti-claudin-4 (bottom blot) or mouse anti-HA (top blot) antibody. Dark circle corresponds to kAE1 carrying complex oligosaccharide, white circle indicates kAE1 carrying high mannose oligosaccharide. ( B ) A similar experiment was performed but immunoprecipitating claudin-3. No kAE1 co-immunoprecipitated with claudin-3 (top panel). ( C ) The claudin-4 immunoprecipitation was repeated using mIMCD3 kAE1 E681Q cells and shows that claudin-4 interacts with the mutant. ( D ) Proximity ligation assay was performed on mIMCD3 cells stably expressing kAE1 grown for 10 days on semi-permeable filters. CNX, corresponding to calnexin, was used as a positive control 15 , Cldn-4 corresponds to claudin-4. Cells were examined under a confocal microscope using a 63 X objective and sections (X-Z) or top views (X-Y) are shown. Red signal indicates that the 2 proteins labeled are within 30 to 40 nm of distance from each other. Nuclear staining is shown in blue as DAPI staining.
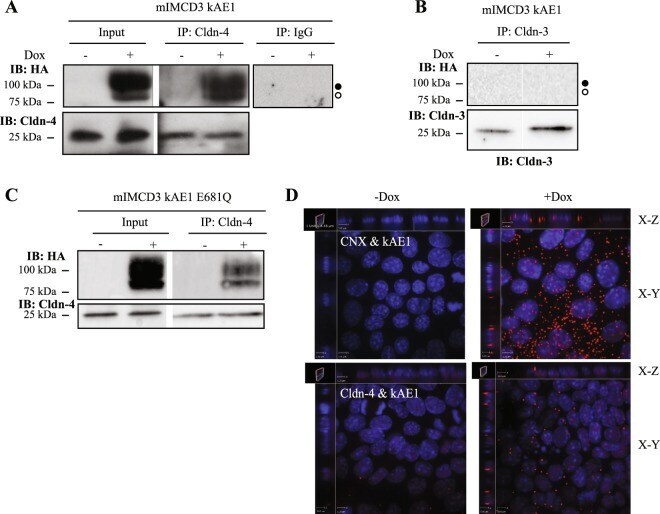
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- Fig. 1 Claudin (CLDN) 3. Positive expression in BRCA1 breast cancer (BC) ( a ) and negative control (isotypic antibody) ( b ). Positive CLDN4 expression in BRCA1 -mutated BC ( c ), negative control of CLDN4 (isotypic antibody) ( d ). Positive CLDN7 in BRCA2 BC ( e ), negative CLDN7 in BRCA1 -related tumor ( f ), and negative control of CLDN7 (isotypic antibody) ( g ). Positive CK5 staining in BRCA1 BC ( h )