Explore
Explore Validate
Validate Learn
Learn Western blot
Western blot ELISA
ELISA Immunocytochemistry
ImmunocytochemistryAntibody data
- Antibody Data
- Antigen structure
- References [31]
- Comments [0]
- Validations
- Immunocytochemistry [8]
- Immunohistochemistry [8]
- Other assay [18]
Submit
Validation data
Reference
Comment
Report error
- Product number
- PA1-933 - Provider product page

- Provider
- Invitrogen Antibodies
- Product name
- Parvalbumin Polyclonal Antibody
- Antibody type
- Polyclonal
- Antigen
- Purifed from natural sources
- Description
- PA1-933 detects parvalbumin protein in human and rat samples. PA1-933 has successfully been used in Western blot, ELISA, immunoprecipitation and immunohistochemical procedures. By Western blot, this antibody detects a 12 kDa protein representing parvalbumin from rat cerebellum. Parvalbumin protein is relatively small and, therefore, it is recommended that the electrophoresis be performed using tricine-SDS-PAGE gels and transferred to a nylon membrane. The PA1-933 immunizing protein corresponds to purified parvalbumin from rat skeletal muscle. Reconstitute in 100 µL PBS to create a stock of 1 mg/mL.
- Reactivity
- Human, Rat
- Host
- Rabbit
- Isotype
- IgG
- Vial size
- 100 μg
- Concentration
- 1 mg/mL
- Storage
- -20°C, Avoid Freeze/Thaw Cycles
Submitted references alms1 mutant zebrafish do not show hair cell phenotypes seen in other cilia mutants.
Inhibitory Neural Network's Impairments at Hippocampal CA1 LTP in an Aged Transgenic Mouse Model of Alzheimer's Disease.
Chronic loss of inhibition in piriform cortex following brief, daily optogenetic stimulation.
Diffusible Tumor Necrosis Factor-Alpha (TNF-α) Promotes Noise-Induced Parvalbumin-Positive (PV+) Neuron Loss and Auditory Processing Impairments.
Mechanotransduction Activity Facilitates Hair Cell Toxicity Caused by the Heavy Metal Cadmium.
Resistance to noise-induced gap detection impairment in FVB mice is correlated with reduced neuroinflammatory response and parvalbumin-positive neuron loss.
Spatial Expression Pattern of the Major Ca(2+)-Buffer Proteins in Mouse Retinal Ganglion Cells.
Group II metabotropic glutamate receptor expressing neurons in anterior cingulate cortex become sensitized after inflammatory and neuropathic pain.
Breakdown of spatial coding and interneuron synchronization in epileptic mice.
An Autaptic Culture System for Standardized Analyses of iPSC-Derived Human Neurons.
The role of retrograde intraflagellar transport genes in aminoglycoside-induced hair cell death.
Characterization of Detergent Insoluble Proteome in Chronic Traumatic Encephalopathy.
Dysbindin Deficiency Modifies the Expression of GABA Neuron and Ion Permeation Transcripts in the Developing Hippocampus.
Holothurian Nervous System Diversity Revealed by Neuroanatomical Analysis.
The reemergence of long-term potentiation in aged Alzheimer's disease mouse model.
Changes in parvalbumin immunoreactive retinal ganglion cells and amacrine cells after optic nerve injury.
The effects of postnatal exposure to low-dose bisphenol-A on activity-dependent plasticity in the mouse sensory cortex.
Aging somatosensory cortex displays increased density of WFA-binding perineuronal nets associated with GAD-negative neurons.
Cholinergic neurons excite cortically projecting basal forebrain GABAergic neurons.
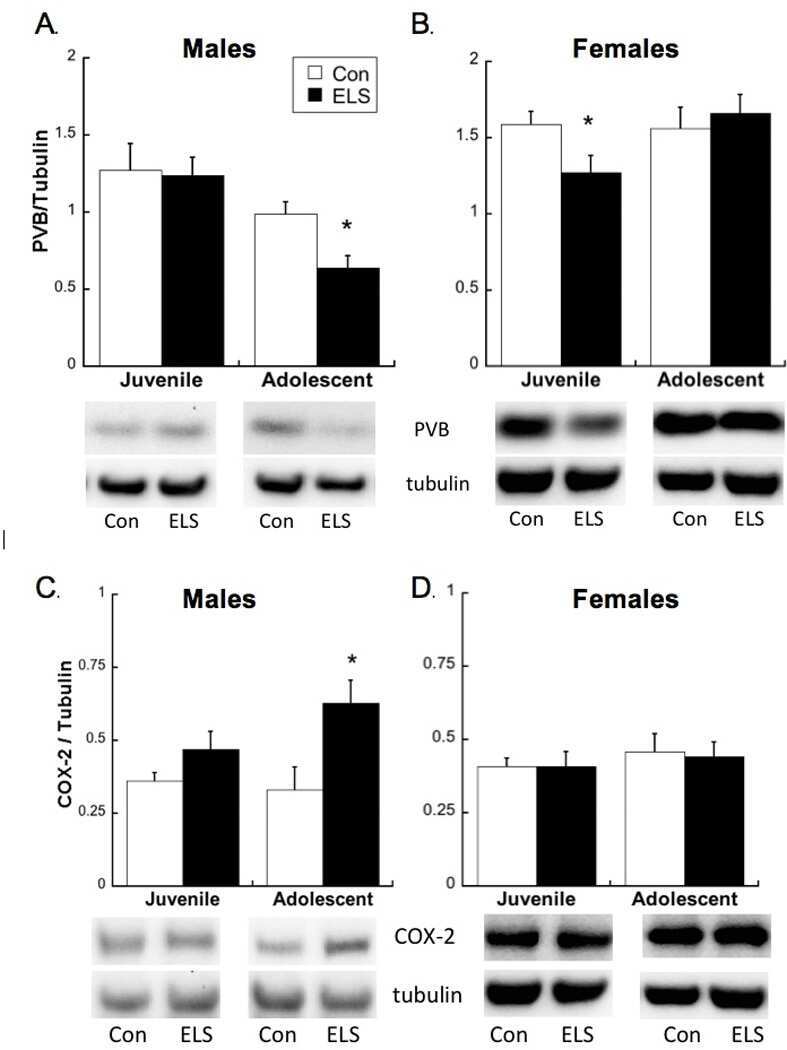
Early life stress disrupts social behavior and prefrontal cortex parvalbumin interneurons at an earlier time-point in females than in males.
Calbindin-D32k is localized to a subpopulation of neurons in the nervous system of the sea cucumber Holothuria glaberrima (Echinodermata).
Parvalbumin gene transfer impairs skeletal muscle contractility in old mice.
Ataxic Guillain-Barré syndrome and acute sensory ataxic neuropathy form a continuous spectrum.
Ageing, but not yet senescent, rats exhibit reduced muscle quality and sarcoplasmic reticulum function.
Organization of glial cells in the adult sea cucumber central nervous system.
Induction of striatal neurogenesis and generation of region-specific functional mature neurons after ischemia by growth factors. Laboratory investigation.
Parvalbumin-containing neurons, perineuronal nets and experience-dependent plasticity in murine barrel cortex.
Endonuclease G expression in thalamic reticular nucleus after global cerebral ischemia.
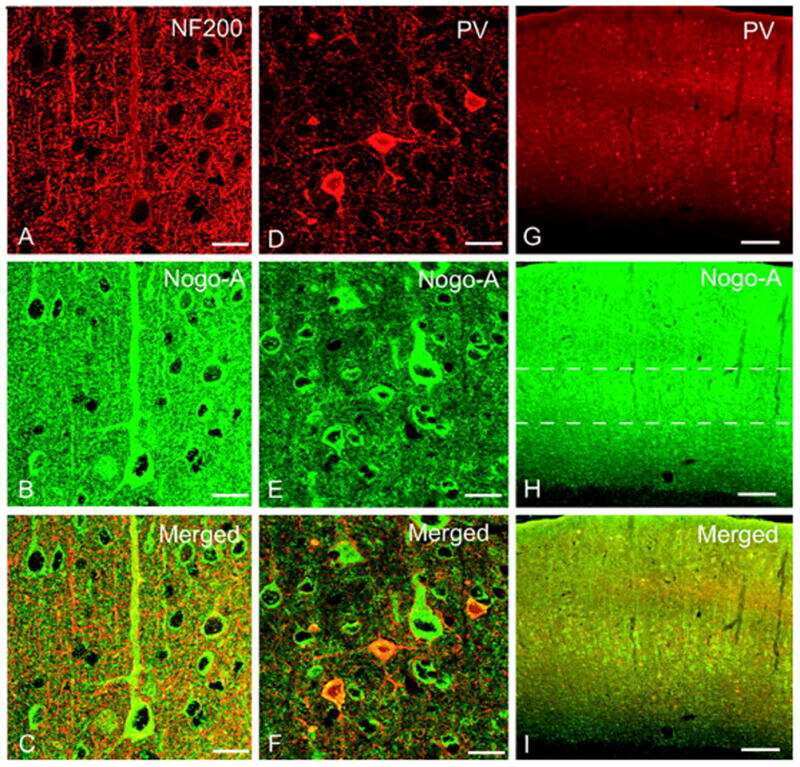
Nogo-A expression after focal ischemic stroke in the adult rat.
Immunoreactive parvalbumin concentrations in parahippocampal gyrus decrease in patients with Alzheimer's disease.
Sensitive immunoassay for rat parvalbumin: tissue distribution and developmental changes.
Parkinson L, Stawicki TM
PloS one 2021;16(4):e0246844
PloS one 2021;16(4):e0246844
Inhibitory Neural Network's Impairments at Hippocampal CA1 LTP in an Aged Transgenic Mouse Model of Alzheimer's Disease.
Seo HJ, Park JE, Choi SM, Kim T, Cho SH, Lee KH, Song WK, Song J, Jeong HS, Kim DH, Kim BC
International journal of molecular sciences 2021 Jan 12;22(2)
International journal of molecular sciences 2021 Jan 12;22(2)
Chronic loss of inhibition in piriform cortex following brief, daily optogenetic stimulation.
Ryu B, Nagappan S, Santos-Valencia F, Lee P, Rodriguez E, Lackie M, Takatoh J, Franks KM
Cell reports 2021 Apr 20;35(3):109001
Cell reports 2021 Apr 20;35(3):109001
Diffusible Tumor Necrosis Factor-Alpha (TNF-α) Promotes Noise-Induced Parvalbumin-Positive (PV+) Neuron Loss and Auditory Processing Impairments.
Deng D, Wang W, Bao S
Frontiers in neuroscience 2020;14:573047
Frontiers in neuroscience 2020;14:573047
Mechanotransduction Activity Facilitates Hair Cell Toxicity Caused by the Heavy Metal Cadmium.
Schmid C, Alampi I, Briggs J, Tarcza K, Stawicki TM
Frontiers in cellular neuroscience 2020;14:37
Frontiers in cellular neuroscience 2020;14:37
Resistance to noise-induced gap detection impairment in FVB mice is correlated with reduced neuroinflammatory response and parvalbumin-positive neuron loss.
Zinsmaier AK, Wang W, Zhang L, Hossainy NN, Bao S
Scientific reports 2020 Nov 24;10(1):20445
Scientific reports 2020 Nov 24;10(1):20445
Spatial Expression Pattern of the Major Ca(2+)-Buffer Proteins in Mouse Retinal Ganglion Cells.
Kovács-Öller T, Szarka G, Tengölics ÁJ, Ganczer A, Balogh B, Szabó-Meleg E, Nyitrai M, Völgyi B
Cells 2020 Mar 25;9(4)
Cells 2020 Mar 25;9(4)
Group II metabotropic glutamate receptor expressing neurons in anterior cingulate cortex become sensitized after inflammatory and neuropathic pain.
Chen S, Kadakia F, Davidson S
Molecular pain 2020 Jan-Dec;16:1744806920915339
Molecular pain 2020 Jan-Dec;16:1744806920915339
Breakdown of spatial coding and interneuron synchronization in epileptic mice.
Shuman T, Aharoni D, Cai DJ, Lee CR, Chavlis S, Page-Harley L, Vetere LM, Feng Y, Yang CY, Mollinedo-Gajate I, Chen L, Pennington ZT, Taxidis J, Flores SE, Cheng K, Javaherian M, Kaba CC, Rao N, La-Vu M, Pandi I, Shtrahman M, Bakhurin KI, Masmanidis SC, Khakh BS, Poirazi P, Silva AJ, Golshani P
Nature neuroscience 2020 Feb;23(2):229-238
Nature neuroscience 2020 Feb;23(2):229-238
An Autaptic Culture System for Standardized Analyses of iPSC-Derived Human Neurons.
Rhee HJ, Shaib AH, Rehbach K, Lee C, Seif P, Thomas C, Gideons E, Guenther A, Krutenko T, Hebisch M, Peitz M, Brose N, Brüstle O, Rhee JS
Cell reports 2019 May 14;27(7):2212-2228.e7
Cell reports 2019 May 14;27(7):2212-2228.e7
The role of retrograde intraflagellar transport genes in aminoglycoside-induced hair cell death.
Stawicki TM, Linbo T, Hernandez L, Parkinson L, Bellefeuille D, Rubel EW, Raible DW
Biology open 2019 Jan 14;8(1)
Biology open 2019 Jan 14;8(1)
Characterization of Detergent Insoluble Proteome in Chronic Traumatic Encephalopathy.
Cherry JD, Zeineddin A, Dammer EB, Webster JA, Duong D, Seyfried NT, Levey AI, Alvarez VE, Huber BR, Stein TD, Kiernan PT, McKee AC, Lah JJ, Hales CM
Journal of neuropathology and experimental neurology 2018 Jan 1;77(1):40-49
Journal of neuropathology and experimental neurology 2018 Jan 1;77(1):40-49
Dysbindin Deficiency Modifies the Expression of GABA Neuron and Ion Permeation Transcripts in the Developing Hippocampus.
Larimore J, Zlatic SA, Arnold M, Singleton KS, Cross R, Rudolph H, Bruegge MV, Sweetman A, Garza C, Whisnant E, Faundez V
Frontiers in genetics 2017;8:28
Frontiers in genetics 2017;8:28
Holothurian Nervous System Diversity Revealed by Neuroanatomical Analysis.
Díaz-Balzac CA, Lázaro-Peña MI, Vázquez-Figueroa LD, Díaz-Balzac RJ, García-Arrarás JE
PloS one 2016;11(3):e0151129
PloS one 2016;11(3):e0151129
The reemergence of long-term potentiation in aged Alzheimer's disease mouse model.
Huh S, Baek SJ, Lee KH, Whitcomb DJ, Jo J, Choi SM, Kim DH, Park MS, Lee KH, Kim BC
Scientific reports 2016 Jul 5;6:29152
Scientific reports 2016 Jul 5;6:29152
Changes in parvalbumin immunoreactive retinal ganglion cells and amacrine cells after optic nerve injury.
Hong CJH, Siddiqui AM, Sabljic TF, Ball AK
Experimental eye research 2016 Apr;145:363-372
Experimental eye research 2016 Apr;145:363-372
The effects of postnatal exposure to low-dose bisphenol-A on activity-dependent plasticity in the mouse sensory cortex.
Kelly EA, Opanashuk LA, Majewska AK
Frontiers in neuroanatomy 2014;8:117
Frontiers in neuroanatomy 2014;8:117
Aging somatosensory cortex displays increased density of WFA-binding perineuronal nets associated with GAD-negative neurons.
Karetko-Sysa M, Skangiel-Kramska J, Nowicka D
Neuroscience 2014 Sep 26;277:734-46
Neuroscience 2014 Sep 26;277:734-46
Cholinergic neurons excite cortically projecting basal forebrain GABAergic neurons.
Yang C, McKenna JT, Zant JC, Winston S, Basheer R, Brown RE
The Journal of neuroscience : the official journal of the Society for Neuroscience 2014 Feb 19;34(8):2832-44
The Journal of neuroscience : the official journal of the Society for Neuroscience 2014 Feb 19;34(8):2832-44
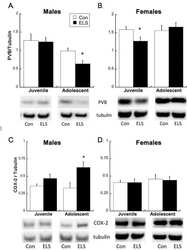
Early life stress disrupts social behavior and prefrontal cortex parvalbumin interneurons at an earlier time-point in females than in males.
Holland FH, Ganguly P, Potter DN, Chartoff EH, Brenhouse HC
Neuroscience letters 2014 Apr 30;566:131-6
Neuroscience letters 2014 Apr 30;566:131-6
Calbindin-D32k is localized to a subpopulation of neurons in the nervous system of the sea cucumber Holothuria glaberrima (Echinodermata).
Díaz-Balzac CA, Lázaro-Peña MI, García-Rivera EM, González CI, García-Arrarás JE
PloS one 2012;7(3):e32689
PloS one 2012;7(3):e32689
Parvalbumin gene transfer impairs skeletal muscle contractility in old mice.
Murphy KT, Ham DJ, Church JE, Naim T, Trieu J, Williams DA, Lynch GS
Human gene therapy 2012 Aug;23(8):824-36
Human gene therapy 2012 Aug;23(8):824-36
Ataxic Guillain-Barré syndrome and acute sensory ataxic neuropathy form a continuous spectrum.
Ito M, Matsuno K, Sakumoto Y, Hirata K, Yuki N
Journal of neurology, neurosurgery, and psychiatry 2011 Mar;82(3):294-9
Journal of neurology, neurosurgery, and psychiatry 2011 Mar;82(3):294-9
Ageing, but not yet senescent, rats exhibit reduced muscle quality and sarcoplasmic reticulum function.
Russ DW, Grandy JS, Toma K, Ward CW
Acta physiologica (Oxford, England) 2011 Mar;201(3):391-403
Acta physiologica (Oxford, England) 2011 Mar;201(3):391-403
Organization of glial cells in the adult sea cucumber central nervous system.
Mashanov VS, Zueva OR, Garcia-Arraras JE
Glia 2010 Oct;58(13):1581-93
Glia 2010 Oct;58(13):1581-93
Induction of striatal neurogenesis and generation of region-specific functional mature neurons after ischemia by growth factors. Laboratory investigation.
Yoshikawa G, Momiyama T, Oya S, Takai K, Tanaka J, Higashiyama S, Saito N, Kirino T, Kawahara N
Journal of neurosurgery 2010 Oct;113(4):835-50
Journal of neurosurgery 2010 Oct;113(4):835-50
Parvalbumin-containing neurons, perineuronal nets and experience-dependent plasticity in murine barrel cortex.
Nowicka D, Soulsby S, Skangiel-Kramska J, Glazewski S
The European journal of neuroscience 2009 Dec 3;30(11):2053-63
The European journal of neuroscience 2009 Dec 3;30(11):2053-63
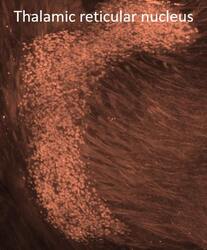
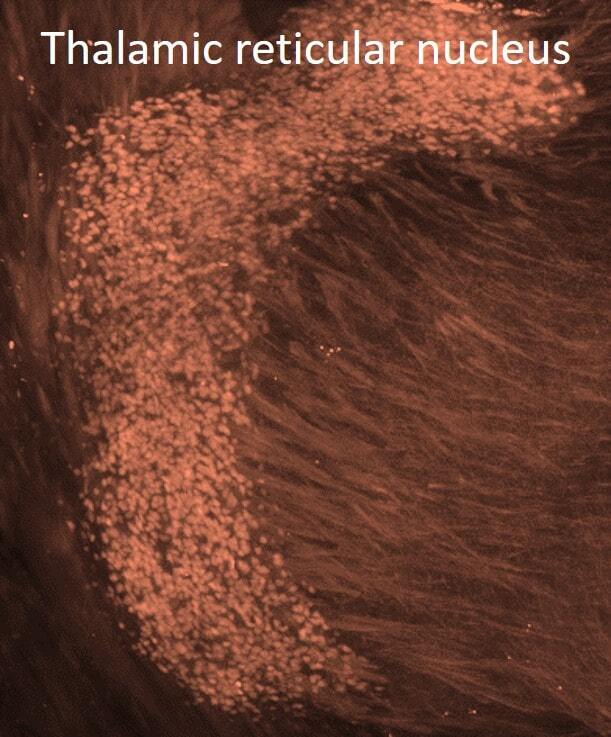
Endonuclease G expression in thalamic reticular nucleus after global cerebral ischemia.
Nielsen M, Zimmer J, Diemer NH
Experimental brain research 2008 Sep;190(1):81-9
Experimental brain research 2008 Sep;190(1):81-9
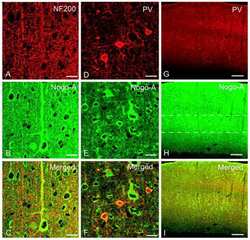
Nogo-A expression after focal ischemic stroke in the adult rat.
Cheatwood JL, Emerick AJ, Schwab ME, Kartje GL
Stroke 2008 Jul;39(7):2091-8
Stroke 2008 Jul;39(7):2091-8
Immunoreactive parvalbumin concentrations in parahippocampal gyrus decrease in patients with Alzheimer's disease.
Inaguma Y, Shinohara H, Inagaki T, Kato K
Journal of the neurological sciences 1992 Jul;110(1-2):57-61
Journal of the neurological sciences 1992 Jul;110(1-2):57-61
Sensitive immunoassay for rat parvalbumin: tissue distribution and developmental changes.
Inaguma Y, Kurobe N, Shinohara H, Kato K
Biochimica et biophysica acta 1991 Sep 2;1075(1):68-74
Biochimica et biophysica acta 1991 Sep 2;1075(1):68-74
No comments: Submit comment
Supportive validation
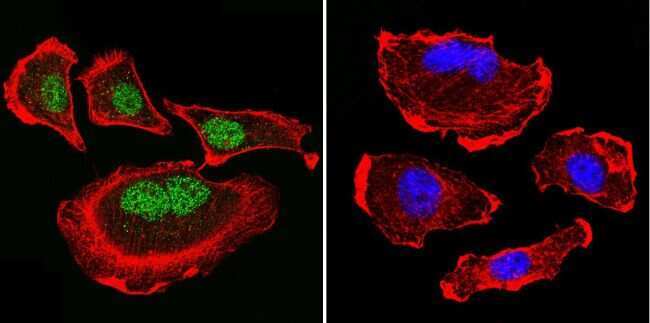
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
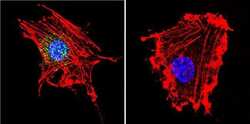
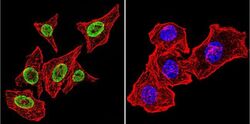
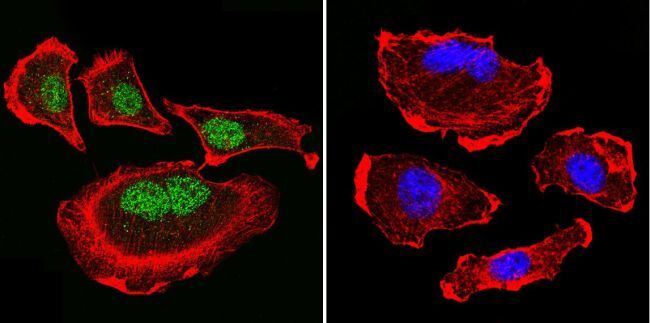
- Immunofluorescent analysis of Parvalbumin using Anti-Parvalbumin Polyclonal Antibody (Product # PA1-933) shows staining in C6 Cells. Parvalbumin staining (green), F-Actin staining with Phalloidin (red) and nuclei with DAPI (blue) is shown. Cells were grown on chamber slides and fixed with formaldehyde prior to staining. Cells were probed without (control) or with or an antibody recognizing Parvalbumin (Product # PA1-933) at a dilution of 1:100 over night at 4°C, washed with PBS and incubated with a DyLight-488 conjugated secondary antibody (Product # 35552, Goat Anti-Rabbit). Images were taken at 60X magnification.
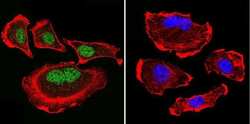
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
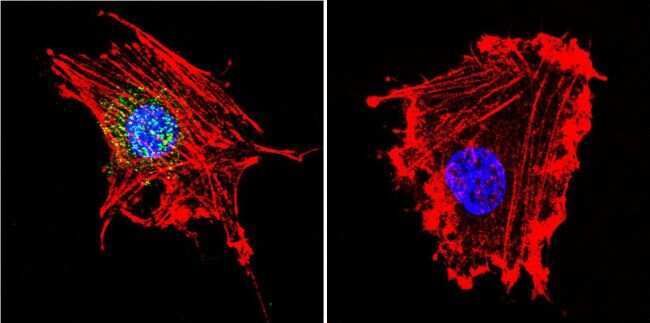
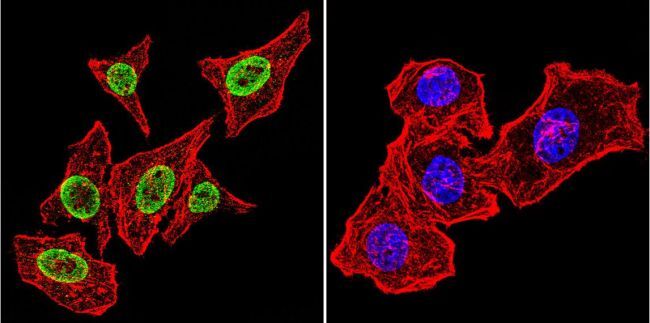
- Immunofluorescent analysis of Parvalbumin using Anti-Parvalbumin Polyclonal Antibody (Product # PA1-933) shows staining in Hela Cells. Parvalbumin staining (green), F-Actin staining with Phalloidin (red) and nuclei with DAPI (blue) is shown. Cells were grown on chamber slides and fixed with formaldehyde prior to staining. Cells were probed without (control) or with or an antibody recognizing Parvalbumin (Product # PA1-933) at a dilution of 1:100 over night at 4°C, washed with PBS and incubated with a DyLight-488 conjugated secondary antibody (Product # 35552, Goat Anti-Rabbit). Images were taken at 60X magnification.
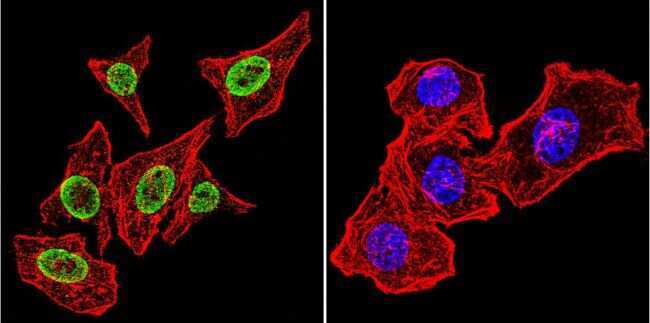
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
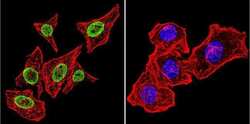
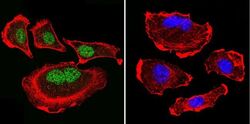
- Immunofluorescent analysis of Parvalbumin using Anti-Parvalbumin Polyclonal Antibody (Product # PA1-933) shows staining in U251 Cells. Parvalbumin staining (green), F-Actin staining with Phalloidin (red) and nuclei with DAPI (blue) is shown. Cells were grown on chamber slides and fixed with formaldehyde prior to staining. Cells were probed without (control) or with or an antibody recognizing Parvalbumin (Product # PA1-933) at a dilution of 1:200 over night at 4°C, washed with PBS and incubated with a DyLight-488 conjugated secondary antibody (Product # 35552, Goat Anti-Rabbit). Images were taken at 60X magnification.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
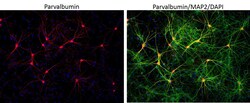
- Experimental details
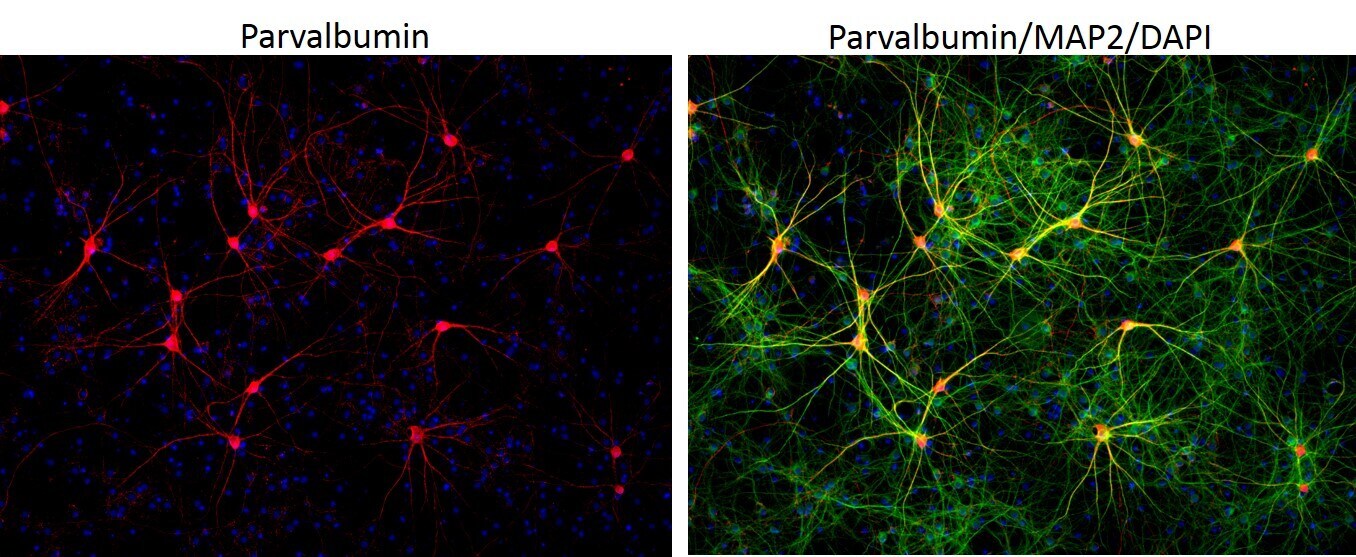
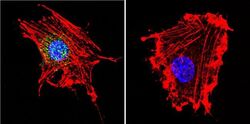
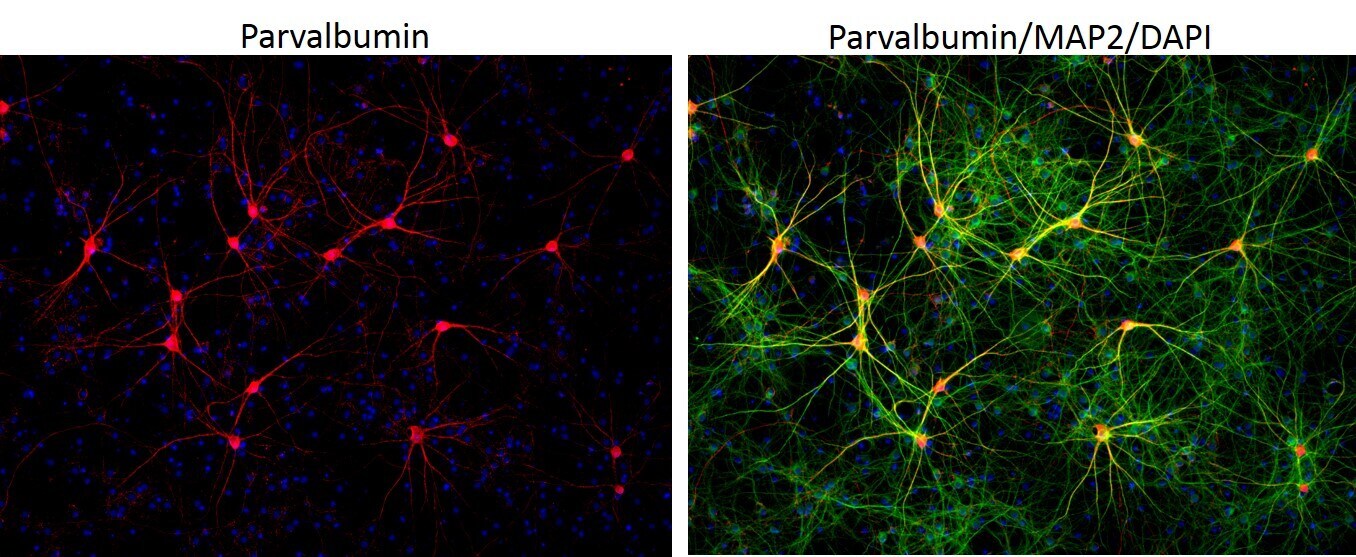
- Immunofluorescent analysis of MAP2 (green) and parvalbumin (red) on rat primary Hippocampal neurons (E18) (Product # A15587) cultured for 28 days in the B-27 Plus Neuronal Culture System (Product # A3653401). At day 28 the cells were fixed with 4% paraformaldehyde for 15 min, permeabilized with 0.1% triton x-100 for 30min, and blocked with 1% BSA for 30 min at room temperature. Cells were stained with anti-parvalbumin antibody (Product # PA1-933) at a dilution of 1:200, and anti-MAP2 (Product # 13-1500) at a dilution of1:4000, in 1% BSA staining buffer, overnight at 4C, and then incubated with Alexa Fluor 488 conjugated donkey anti-rabbit (Product # A-21206) and Alexa Fluor 594 donkey anti-mouse (Product # A-21203) antibodies at a dilution of 1:1000 for 30 min. at room temp. Wash 3 times with DPBS. Stain with DAPI for nucleus.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
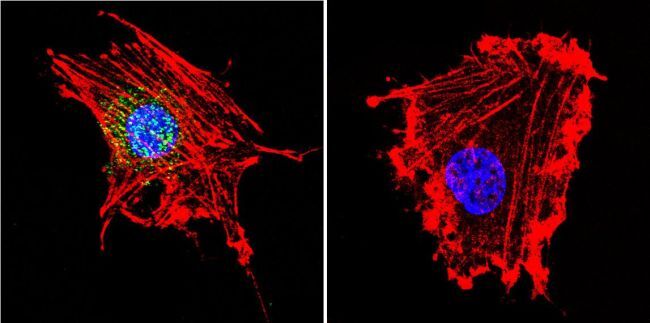
- Immunofluorescent analysis of Parvalbumin using Anti-Parvalbumin Polyclonal Antibody (Product # PA1-933) shows staining in Hela Cells. Parvalbumin staining (green), F-Actin staining with Phalloidin (red) and nuclei with DAPI (blue) is shown. Cells were grown on chamber slides and fixed with formaldehyde prior to staining. Cells were probed without (control) or with or an antibody recognizing Parvalbumin (Product # PA1-933) at a dilution of 1:100 over night at 4°C, washed with PBS and incubated with a DyLight-488 conjugated secondary antibody (Product # 35552, Goat Anti-Rabbit). Images were taken at 60X magnification.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- Immunofluorescent analysis of Parvalbumin using Anti-Parvalbumin Polyclonal Antibody (Product # PA1-933) shows staining in U251 Cells. Parvalbumin staining (green), F-Actin staining with Phalloidin (red) and nuclei with DAPI (blue) is shown. Cells were grown on chamber slides and fixed with formaldehyde prior to staining. Cells were probed without (control) or with or an antibody recognizing Parvalbumin (Product # PA1-933) at a dilution of 1:200 over night at 4°C, washed with PBS and incubated with a DyLight-488 conjugated secondary antibody (Product # 35552, Goat Anti-Rabbit). Images were taken at 60X magnification.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
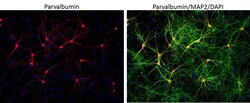
- Immunofluorescent analysis of Parvalbumin using Anti-Parvalbumin Polyclonal Antibody (Product # PA1-933) shows staining in C6 Cells. Parvalbumin staining (green), F-Actin staining with Phalloidin (red) and nuclei with DAPI (blue) is shown. Cells were grown on chamber slides and fixed with formaldehyde prior to staining. Cells were probed without (control) or with or an antibody recognizing Parvalbumin (Product # PA1-933) at a dilution of 1:100 over night at 4°C, washed with PBS and incubated with a DyLight-488 conjugated secondary antibody (Product # 35552, Goat Anti-Rabbit). Images were taken at 60X magnification.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- Immunofluorescent analysis of MAP2 (green) and parvalbumin (red) on rat primary Hippocampal neurons (E18) (Product # A15587) cultured for 28 days in the B-27 Plus Neuronal Culture System (Product # A3653401). At day 28 the cells were fixed with 4% paraformaldehyde for 15 min, permeabilized with 0.1% triton x-100 for 30min, and blocked with 1% BSA for 30 min at room temperature. Cells were stained with anti-parvalbumin antibody (Product # PA1-933) at a dilution of 1:200, and anti-MAP2 (Product # 13-1500) at a dilution of1:4000, in 1% BSA staining buffer, overnight at 4C, and then incubated with Alexa Fluor 488 conjugated donkey anti-rabbit (Product # A-21206) and Alexa Fluor 594 donkey anti-mouse (Product # A-21203) antibodies at a dilution of 1:1000 for 30 min. at room temp. Wash 3 times with DPBS. Stain with DAPI for nucleus.
Supportive validation
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
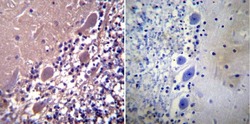
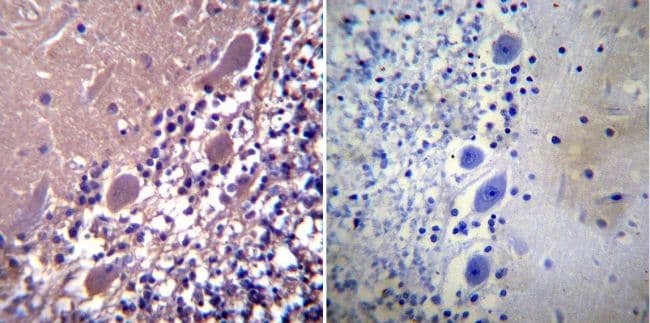
- Experimental details
- Immunohistochemistry was performed on normal biopsies of deparaffinized Human cerebellum tissue. To expose target proteins, heat induced antigen retrieval was performed using 10mM sodium citrate (pH6.0) buffer, microwaved for 8-15 minutes. Following antigen retrieval tissues were blocked in 3% BSA-PBS for 30 minutes at room temperature. Tissues were then probed at a dilution of 1:100 with a rabbit polyclonal antibody recognizing Parvalbumin (Product # PA1-933) or without primary antibody (negative control) overnight at 4°C in a humidified chamber. Tissues were washed extensively with PBST and endogenous peroxidase activity was quenched with a peroxidase suppressor. Detection was performed using a biotin-conjugated secondary antibody and SA-HRP, followed by colorimetric detection using DAB. Tissues were counterstained with hematoxylin and prepped for mounting.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
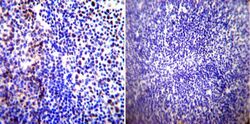
- Experimental details
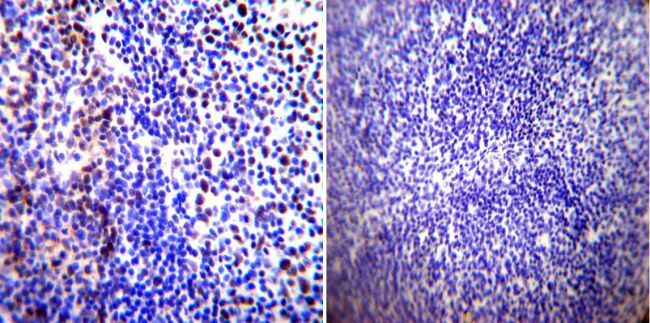
- Immunohistochemistry was performed on normal biopsies of deparaffinized Human tonsil tissue. To expose target proteins, heat induced antigen retrieval was performed using 10mM sodium citrate (pH6.0) buffer, microwaved for 8-15 minutes. Following antigen retrieval tissues were blocked in 3% BSA-PBS for 30 minutes at room temperature. Tissues were then probed at a dilution of 1:100 with a rabbit polyclonal antibody recognizing Parvalbumin (Product # PA1-933) or without primary antibody (negative control) overnight at 4°C in a humidified chamber. Tissues were washed extensively with PBST and endogenous peroxidase activity was quenched with a peroxidase suppressor. Detection was performed using a biotin-conjugated secondary antibody and SA-HRP, followed by colorimetric detection using DAB. Tissues were counterstained with hematoxylin and prepped for mounting.
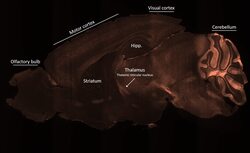
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
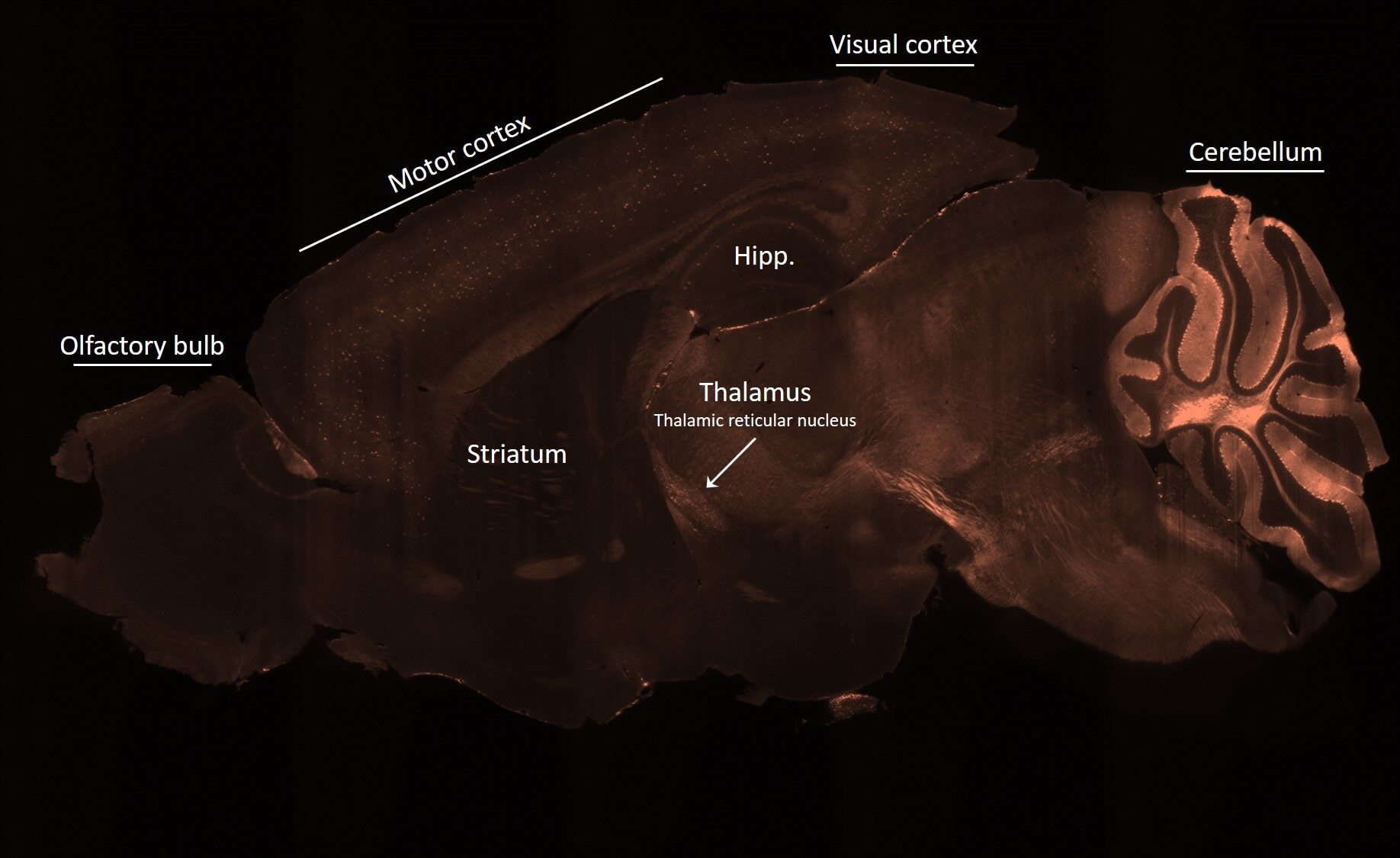
- Experimental details
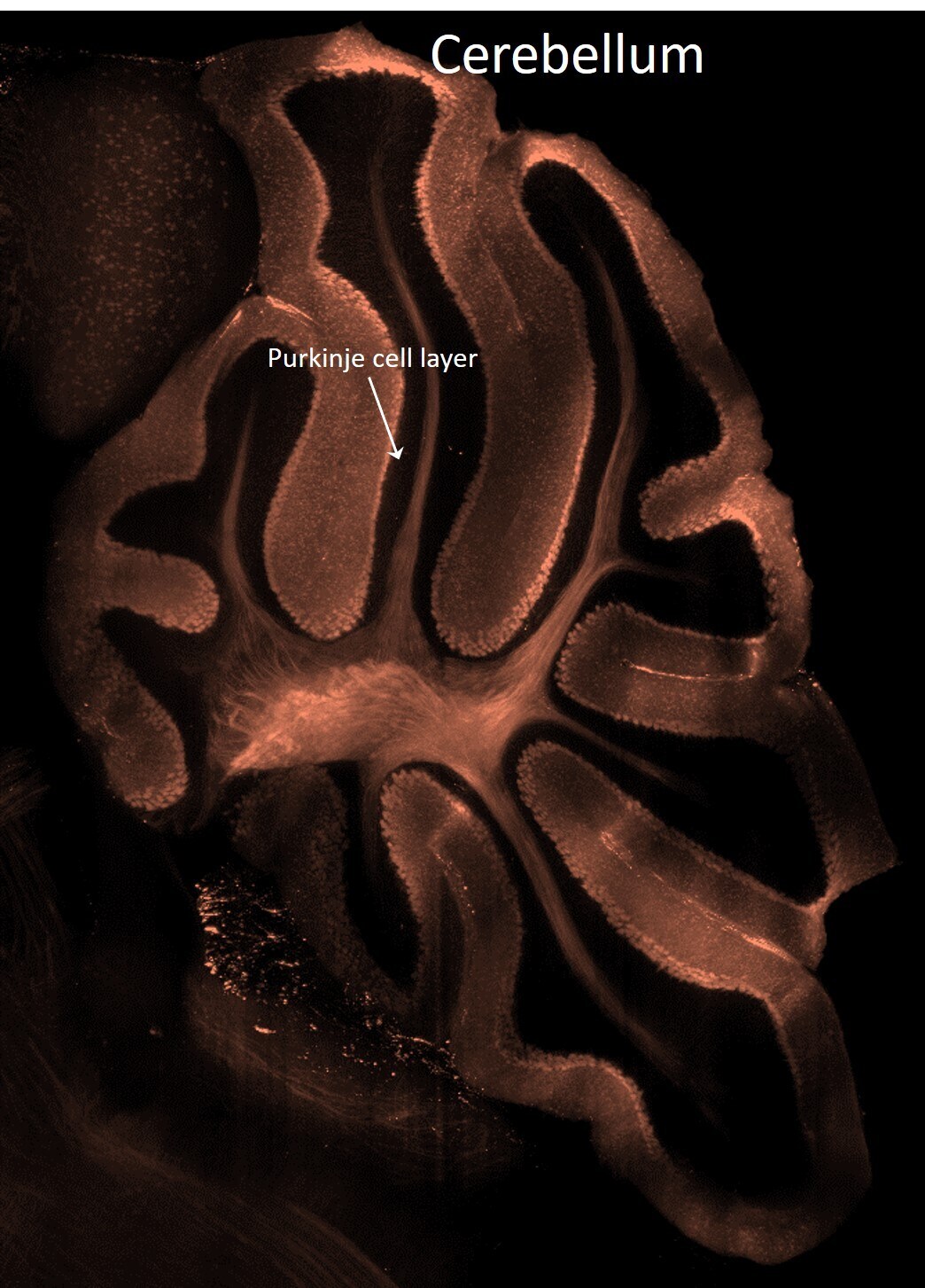
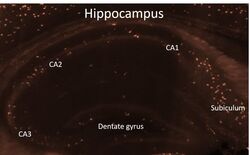
- Spatial expression of Parvalbumin in intact mouse brain hemispheres. Tissue was perfused with LifeCanvas’s SHIELD (Park et al., Nature Biotech, 2018) solution kit. Before antibody labeling, lipids were removed for best antibody diffusion. 20 µg anti-Parvalbumin (PV) polyclonal antibody (Product # PA1-933) was then used to actively label the intact tissue sample for < 2 days. A Rhodamine Red-X conjugated Fab fragment secondary antibody was also used in SmartLabel. After antibody labeling, tissue was then refractive index matched using EasyIndex and imaged at single-cell resolution (1.8 µm/pixel in XY with 4-µm Z-step; 561 nm laser line) on LifeCanvas’s SmartSPIM light-sheet microscope. Data courtesy of LifeCanvas Technologies.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
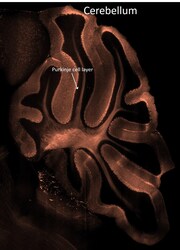
- Experimental details
- Spatial expression of Parvalbumin in intact mouse brain hemispheres. Tissue was perfused with LifeCanvas’s SHIELD (Park et al., Nature Biotech, 2018) solution kit. Before antibody labeling, lipids were removed for best antibody diffusion. 20 µg anti-Parvalbumin (PV) polyclonal antibody (Product # PA1-933) was then used to actively label the intact tissue sample for < 2 days. A Rhodamine Red-X conjugated Fab fragment secondary antibody was also used in SmartLabel. After antibody labeling, tissue was then refractive index matched using EasyIndex and imaged at single-cell resolution (1.8 µm/pixel in XY with 4-µm Z-step; 561 nm laser line) on LifeCanvas’s SmartSPIM light-sheet microscope. Data courtesy of LifeCanvas Technologies.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- Spatial expression of Parvalbumin in intact mouse brain hemispheres. Tissue was perfused with LifeCanvas’s SHIELD (Park et al., Nature Biotech, 2018) solution kit. Before antibody labeling, lipids were removed for best antibody diffusion. 20 µg anti-Parvalbumin (PV) polyclonal antibody (Product # PA1-933) was then used to actively label the intact tissue sample for < 2 days. A Rhodamine Red-X conjugated Fab fragment secondary antibody was also used in SmartLabel. After antibody labeling, tissue was then refractive index matched using EasyIndex and imaged at single-cell resolution (1.8 µm/pixel in XY with 4-µm Z-step; 561 nm laser line) on LifeCanvas’s SmartSPIM light-sheet microscope. Data courtesy of LifeCanvas Technologies.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- Spatial expression of Parvalbumin in intact mouse brain hemispheres. Tissue was perfused with LifeCanvas’s SHIELD (Park et al., Nature Biotech, 2018) solution kit. Before antibody labeling, lipids were removed for best antibody diffusion. 20 µg anti-Parvalbumin (PV) polyclonal antibody (Product # PA1-933) was then used to actively label the intact tissue sample for < 2 days. A Rhodamine Red-X conjugated Fab fragment secondary antibody was also used in SmartLabel. After antibody labeling, tissue was then refractive index matched using EasyIndex and imaged at single-cell resolution (1.8 µm/pixel in XY with 4-µm Z-step; 561 nm laser line) on LifeCanvas’s SmartSPIM light-sheet microscope. Data courtesy of LifeCanvas Technologies.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
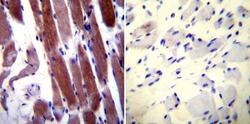
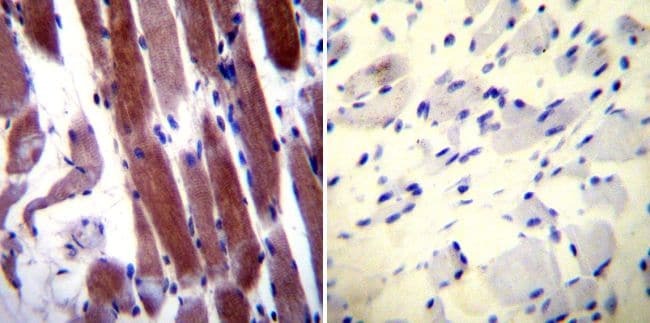
- Immunohistochemistry was performed on normal biopsies of deparaffinized Human skeletal muscle tissue. To expose target proteins, heat induced antigen retrieval was performed using 10mM sodium citrate (pH6.0) buffer, microwaved for 8-15 minutes. Following antigen retrieval tissues were blocked in 3% BSA-PBS for 30 minutes at room temperature. Tissues were then probed at a dilution of 1:20 with a rabbit polyclonal antibody recognizing Parvalbumin (Product # PA1-933) or without primary antibody (negative control) overnight at 4°C in a humidified chamber. Tissues were washed extensively with PBST and endogenous peroxidase activity was quenched with a peroxidase suppressor. Detection was performed using a biotin-conjugated secondary antibody and SA-HRP, followed by colorimetric detection using DAB. Tissues were counterstained with hematoxylin and prepped for mounting.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- Spatial expression of Parvalbumin in intact mouse brain hemispheres. Tissue was perfused with LifeCanvas’s SHIELD (Park et al., Nature Biotech, 2018) solution kit. Before antibody labeling, lipids were removed for best antibody diffusion. 20 µg anti-Parvalbumin (PV) polyclonal antibody (Product # PA1-933) was then used to actively label the intact tissue sample for < 2 days. A Rhodamine Red-X conjugated Fab fragment secondary antibody was also used in SmartLabel. After antibody labeling, tissue was then refractive index matched using EasyIndex and imaged at single-cell resolution (1.8 µm/pixel in XY with 4-µm Z-step; 561 nm laser line) on LifeCanvas’s SmartSPIM light-sheet microscope. Data courtesy of LifeCanvas Technologies.
Supportive validation
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
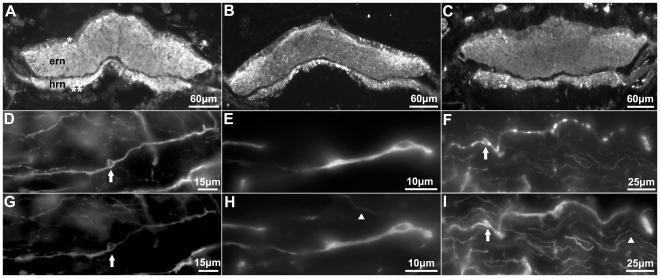
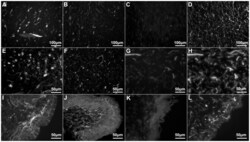
- Figure 1 Anti-calbindin 1, anti-parvalbumin, and anti-calbindin 2 immunoreactivity in the nervous system of Holothuria glaberrima (Holothuroidea). Transverse sections through the radial nerve cords showing immunoreactivity to anti-calbindin 1 (A), anti-parvalbumin (B), and anti-calbindin 2 (C) in the ectoneural and hyponeural components of the radial nerve. Double-labeling of anti-calbindin 1 (D) and RN1 (G), anti-parvalbumin (E) and RN1 (H), and anti-calbindin 2 (F) and RN1 (I) in cells (arrows) and fibers of the connective tissue plexus of the tube feet. Co-labeling is observed with the anti-CBPs markers and RN1 in the majority of the fibers, but not in all (arrowheads). ern, ectoneural component of the radial nerve cord; hrn, hyponeural component of the radial nerve cord; *, ectoneural component cell bodies; **, hyponeural component cell bodies.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
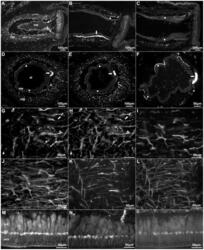
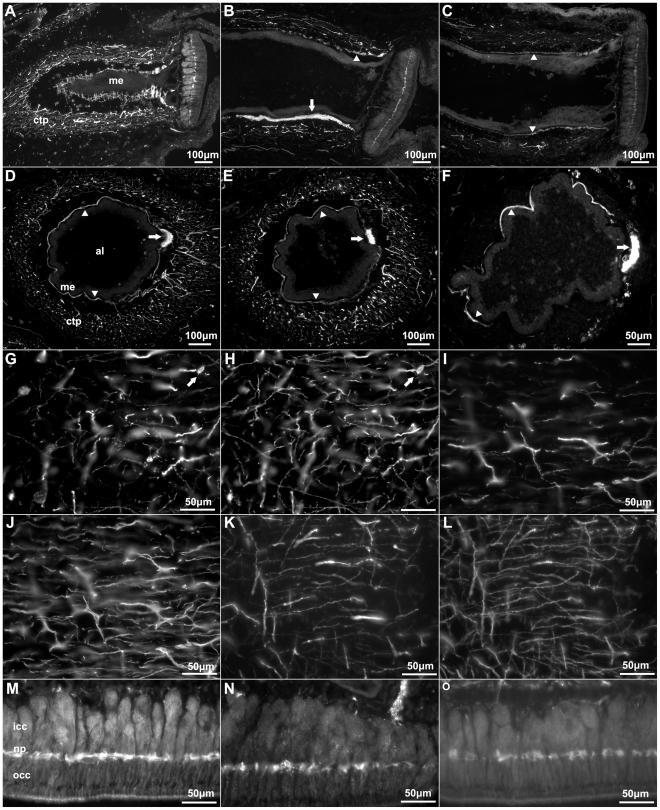
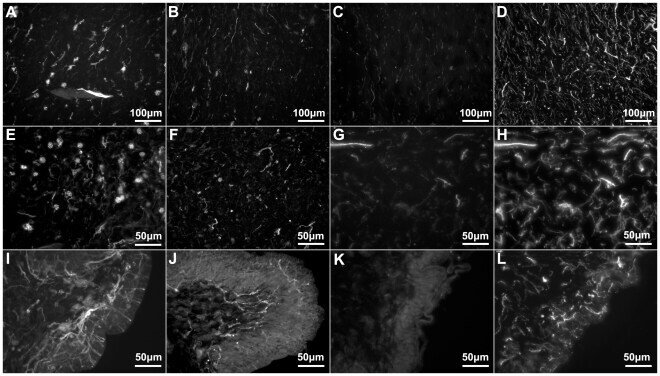
- Figure 4 Anti-calbindin 1, anti-parvalbumin, and anti-calbindin 2 immunoreactivity in the podial nervous system of Holothuria glaberrima (Holothuroidea). Longitudinal and transverse sections through the tube feet showing immunoreactivity to anti-calbindin 1 (A,D,G,M), anti-parvalbumin (B,E,I,N), and anti-calbindin 2 (C,F,K,O) in the different subdivisions of the podial nervous system. (A-F) Immunoreactivity was observed with all markers in the podial nerve (arrows) and in the podial cylindrical fenestrated sheath (arrowheads). (G-L) Co-labeling of anti-calbindin 1 (G) and RN1 (H), anti-parvalbumin (I) and RN1 (J), and anti-calbindin 2 (K) and RN1 (L) was observed in fibers and cells (arrows) of the connective tissue plexus. (M-O) Anti-calbindin 1 (M), anti-parvalbumin (N), and anti-calbindin 2 (O) immunoreactivity was also present in the nerve plate of the tube feet's disk. Al, ambulacral lumen; ctp, connective tissue plexus; icc, inner cluster of cells; me, mesothelium; np, nerve plate; occ, outer cluster of cells.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
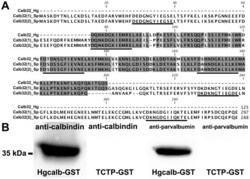
- Figure 7 Western blot of holothurian and echinoid radial nerve cords preparations using anti-calbindin 1 and anti-parvalbumin. (A) A 32 kDa immunoreactive band to anti-calbindin 1 was observed in H. glaberrima and L. variegatus radial nerve cords homogenates. A 28 kDa immunoreactive band corresponding to calbindin 1 was observed in the rat kidney positive control and no immunoreactive bands were observed in the synthetic peptide of an EF-hand domain-containing protein negative control. (B) A 32 kDa immunoreactive band to anti-calbindin 1 was observed in H. glaberrima and L. variegatus radial nerve cords homogenates. A 12 kDa major immunoreactive band corresponding to calbindin 1 was observed in the rat skeletal muscle positive control and no immunoreactive bands were observed in the synthetic peptide of an EF-hand domain-containing protein negative control.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
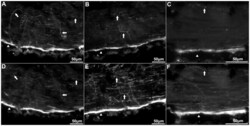
- Figure 2 Anti-calbindin 1, anti-parvalbumin, and anti-calbindin 2 immunoreactivity in the peripheral nerve and circular muscle of Holothuria glaberrima (Holothuroidea). Longitudinal sections through the body wall showing colabeling of anti-calbindin 1 (A) and RN1 (D), anti-parvalbumin (B) and RN1 (E), and anti-calbindin 2 (C) and RN1 (F). Most of the immunoreactivity was observed in the peripheral nerves (arrowheads), while only a minor supopulations of fibers were co-labeled by the anti-CBP and RN1 (arrows).
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
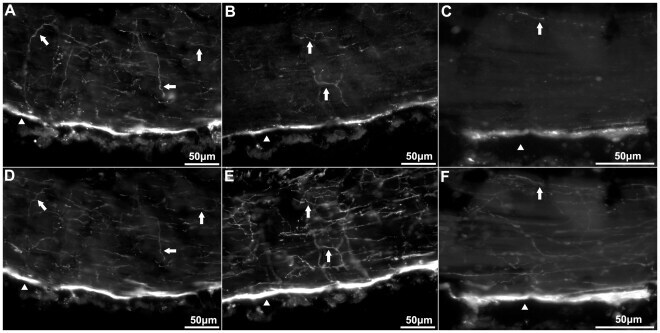
- Figure 3 Anti-calbindin 1, anti-parvalbumin, and anti-calbindin 2 immunoreactivity in the body wall nervous system of Holothuria glaberrima (Holothuroidea). Longitudinal sections through the body wall showing immunoreactivity to anti-calbindin 1 (A,E,I), anti-parvalbumin (B,F,J), and anti-calbindin 2 (C,G,K) as compared to RN1 (D,H,L) immunoreactivity in the different layers of the body wall. Immunoreactivity of the internal connective tissue plexus (A-D), external connective tissue (E-H) and epidermis (I-L) showed that the anti-CBP only labeled a minor subpopulation of fibers and cells within these plexi, as compared to RN1 which labeled a large subpopulations of cells and fibers of the body wall nervous system.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
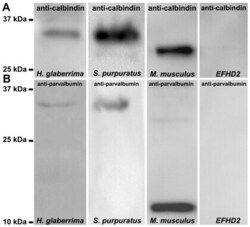
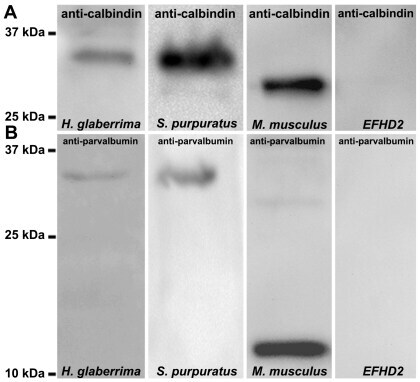
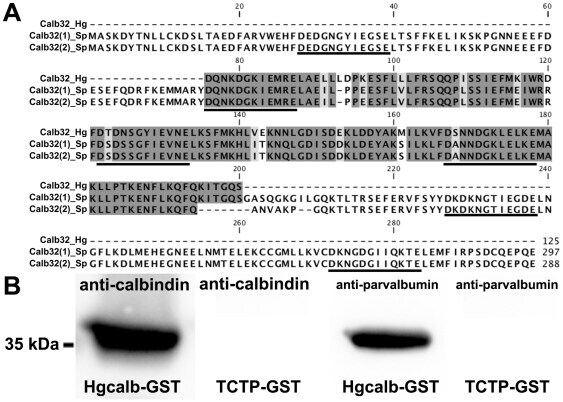
- Figure 8 Identification of calbindin-D32k as the anti-calbindin 1 and anti-parvalbumin epitope in echinoderms. (A) Sequence alignment of H. glaberrima calbindin-D32k fragment (Calb32_Hg) and S. purpuratus calbindin-D32k isoform 1 (Calb32(1)_Sp) and isoform 2 (Calb32(2)_Sp), showing a high degree of homology. Alignment of the proteins was made using the CLUSTALW multiple sequences alignment. Dark gray represent conserved residues, light gray represent conservation of strong groups, and black lines below the residues represents the presence of an EF hand domain. (B) A 36 kDa immunoreactive band to anti-calbindin 1 and anti-parvalbumin was observed in the H. glaberrima GST-tagged calbindin-D32k fragment peptide (Calb32_Hg), and no immunoreactive band to anti-calbindin 1 and anti-parvalbumin was observed in the negative control, GST-tagged translationally controlled tumor protein (TCTP).
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
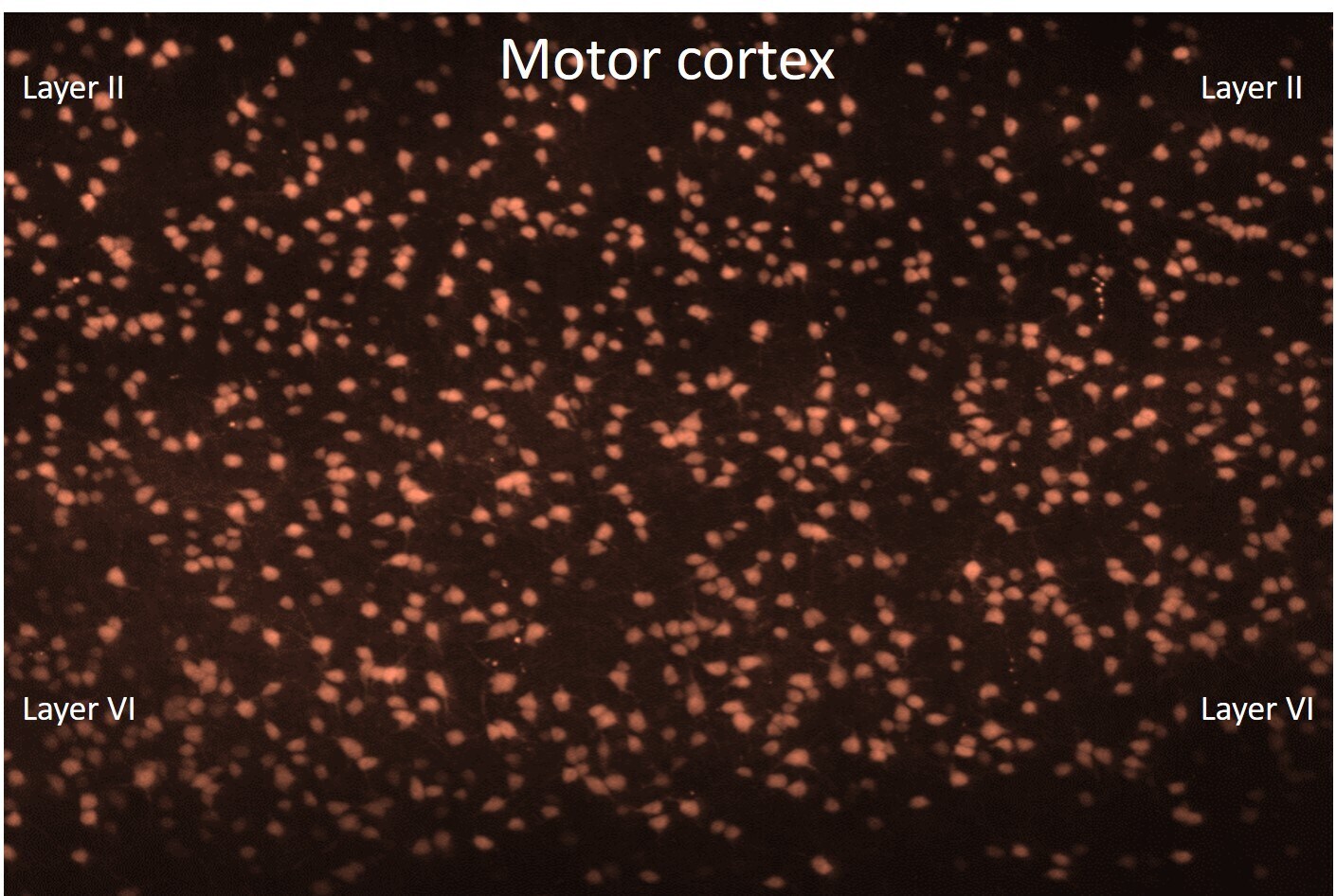
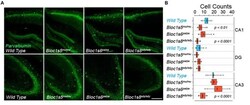
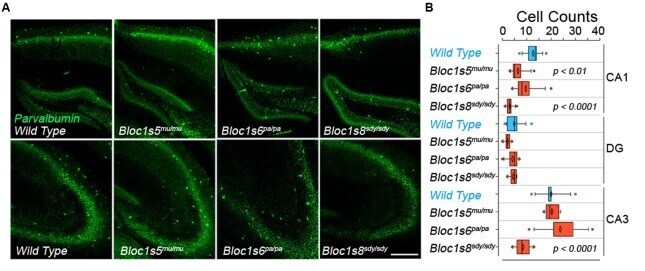
- FIGURE 1 Differential Effects of BLOC-1 Complex Mutations in the Number of Parvalbumin-Positive Neurons. Hippocampal sections from wild type, Bloc1s8 sdy/sdy , Bloc1s5 mu/mu , and Bloc1s6 pa/pa postnatal day 50 adults were stained with antibodies against parvalbumin and imaged by confocal microscopy. (A) Depicts sections of the dentate gyrus and CA1 (upper panels) and the CA3 area (bottom panels) for four genotypes. (B) Depicts quantitation of the number of parvalbumin positive cells per section in each one of the genotypes. Cell counts were performed blind to the genotype of the animal. Wild type ( n = 9), Bloc1s8 sdy/sdy ( n = 7), Bloc1s5 mu/mu ( n = 7), and Bloc1s6 pa/pa ( n = 6) hippocampi analyzed. One Way ANOVA followed by Dunnett's Multiple Comparison.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
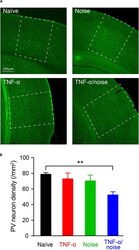
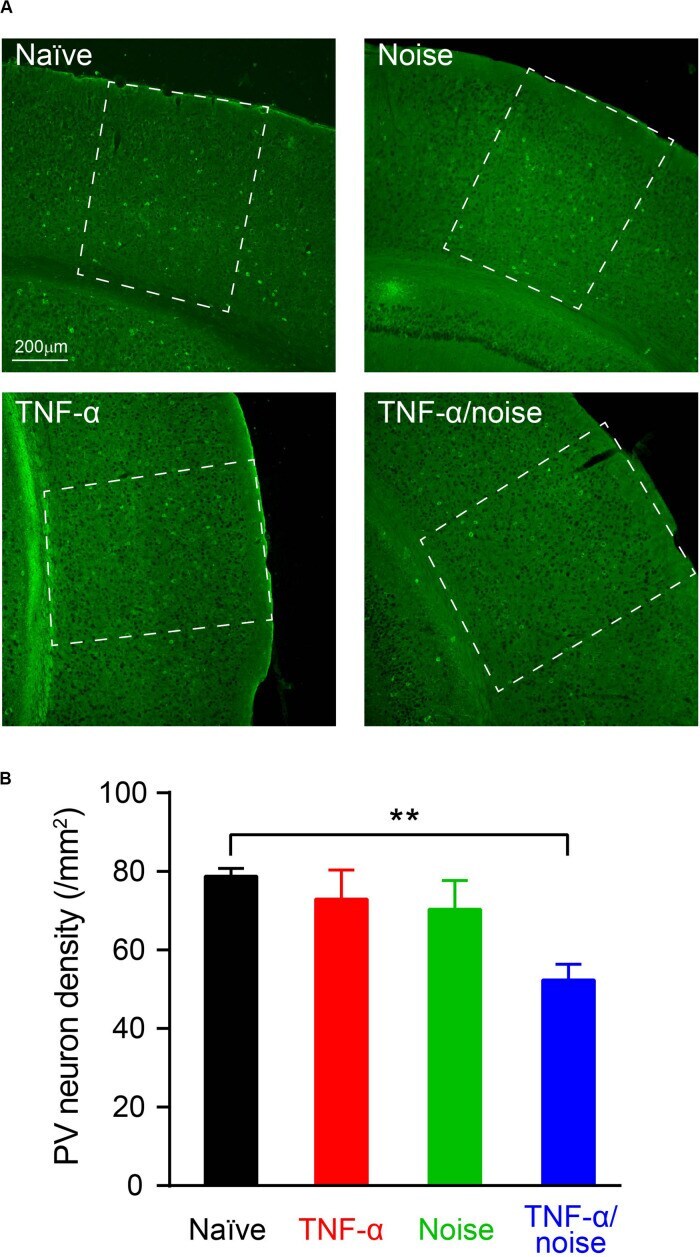
- Experimental details
- FIGURE 4 TNF-alpha infusion and noise exposure synergistically induced PV+ neuron loss. (A) Representative images of PV+ neurons stained with a parvalbumin antibody. (B) PV+ neuron density was reduced only in mice that had received both noise exposure and TNF-alpha infusion. All data are presented as mean +- SEM. ** indicates p < 0.01.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
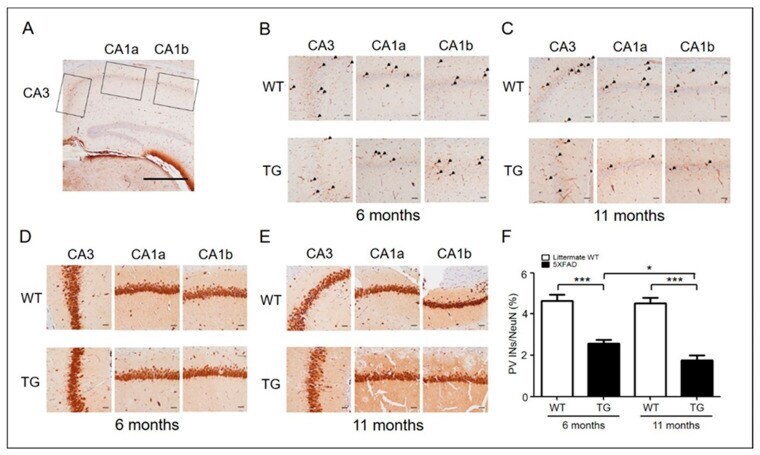
- Experimental details
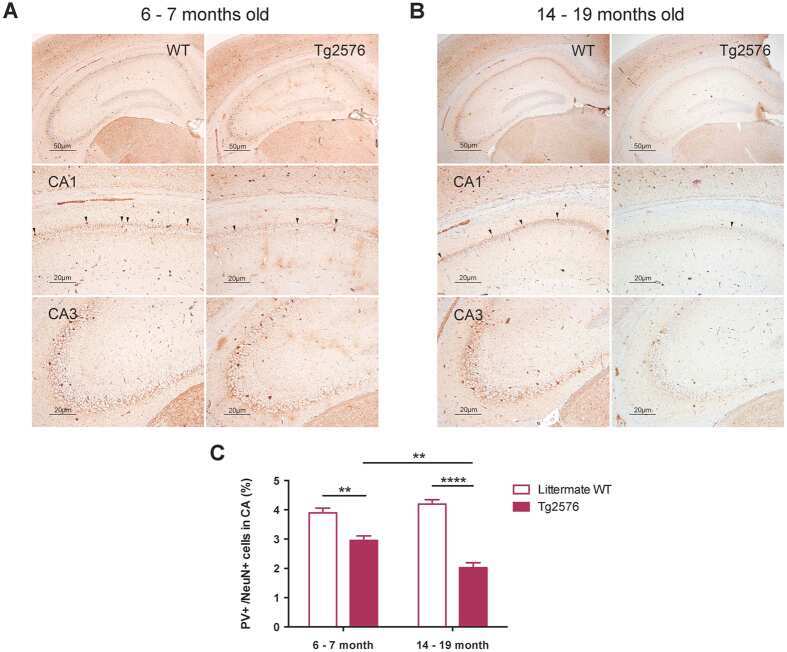
- Figure 1 PV interneurons in AD mice (6-month-old adult and 11-month-old aged) and wild type. ( A ) Representative PV and NeuN immunohistochemical stains of the hippocampus CA3-CA1 region (original magnification, x40) (scale bar = 500 mum). ( B ) Representative PV immunohistochemical stains of the hippocampus of 6-month-old (adult) WT ( n = 6) and 5xFAD ( n = 6) mice. PV-positive interneurons are shown in the hippocampus at a closer distance from CA1a-b (original magnification, x 200, pointed by arrowheads) and CA3 (original magnification, x200) (scale bar = 200 mum). ( C ) Representative PV immunohistochemical stains of the hippocampus of 11-month-old (aged) WT ( n = 6) and 5xFAD ( n = 6) mice. ( D ) Representative NeuN immunohistochemical stains of the hippocampi of 6-month-old (adult) WT ( n = 6) and 5xFAD ( n = 6) mice. NeuN-positive cells are shown in the hippocampus at a closer distance from CA1a-b (original magnification, x200, stained cells) and CA3 (original magnification, x200) (scale bar = 200 mum). ( E ) Representative NeuN immunohistochemical stains of the hippocampus of 11-month-old (aged) WT ( n = 6) and 5xFAD ( n = 6) mice. ( F ) The ratio of PV-positive cells to NeuN-positive cells in adult and aged 5xFAD mice (*, p < 0.05; ***, p < 0.0001 compared with the indicated group using unpaired t -test). AD, Alzheimer's disease; PV, parvalbumin; INs, interneurons; NeuN, neuronal nuclei; CA, cornu ammonis; TG, transgenic; WT, wild-type.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
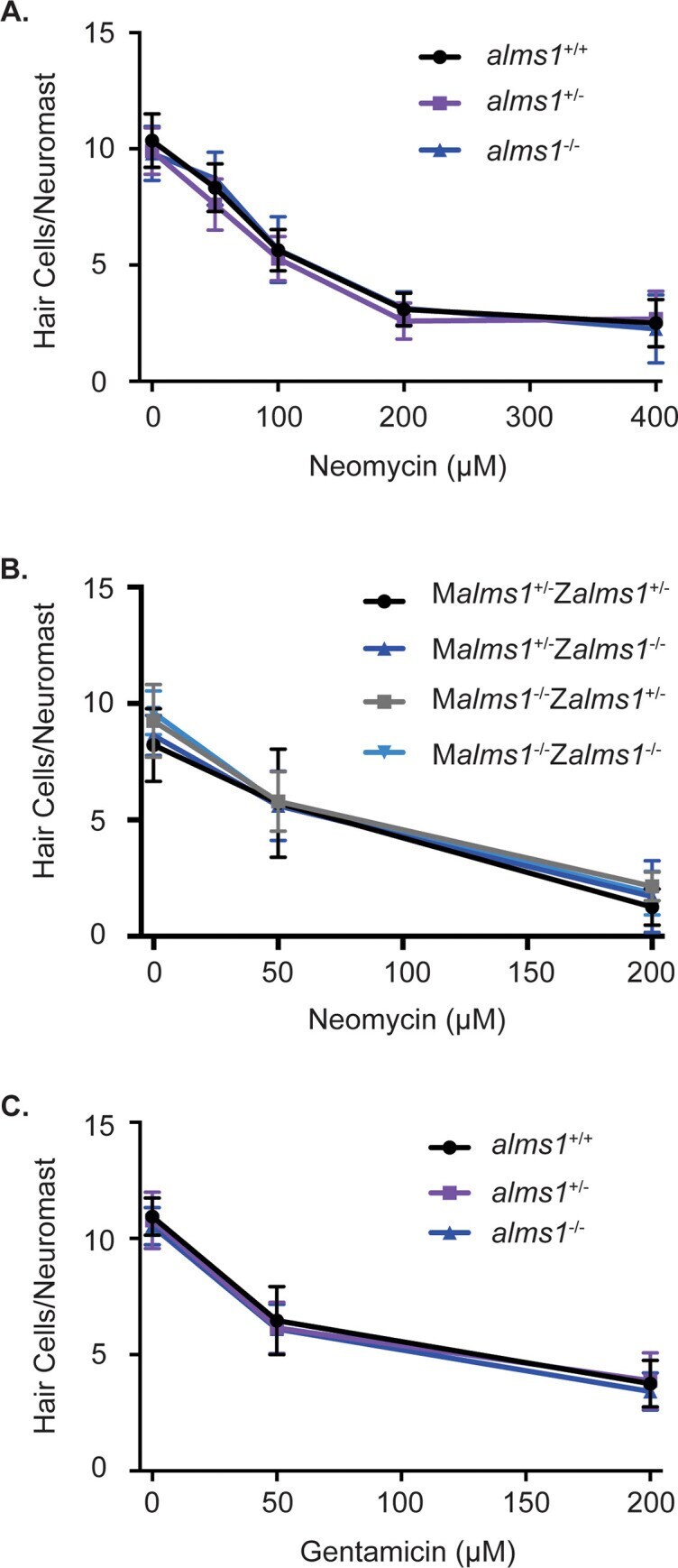
- 10.1371/journal.pone.0246844.g004 Fig 4 alms1 mutant larvae are not resistant to aminoglycoside-induced hair cell death. (A) Dose-response curve for wild-type siblings and alms1 mutants from an incross of heterozygous parents treated with 0-400 muM neomycin for 30 minutes with a one-hour recovery time. n = 7-19 fish per neomycin dose for homozygous wild-type and mutants and 20-25 for heterozygous fish. There were no significant differences due to genotype (p = 0.1273) or the interaction between genotype and neomycin (p = 0.3939) by two-way ANOVA. (B) Dose-response curve for fish that were either heterozygous or homozygous for the alms1 mutation from mothers also either heterozygous or homozygous for the alms1 mutation treated with 0-200 muM neomycin for 30 minutes with a one-hour recovery time. M = maternal genotype and Z = zygotic genotype. n = 7-16 fish per neomycin dose for each genotype. There were no significant differences due to genotype (p = 0.8060) or the interaction between genotype and neomycin (p = 0.2024) by two-way ANOVA. (C) Dose-response curve for wild-type siblings and alms1 mutants from parents heterozygous for the alms1 mutation treated with 0-200 muM gentamicin for 6 hours. n = 10-14 fish per gentamicin dose for homozygous wild-type and mutants and 21-25 for heterozygous fish. There were no significant differences due to genotype (p = 0.9401) or the interaction between genotype and neomycin (p = 0.2850) by two-way ANOVA. Data are shown as mean +/- standard
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
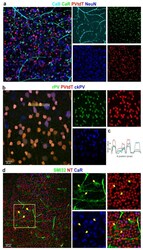
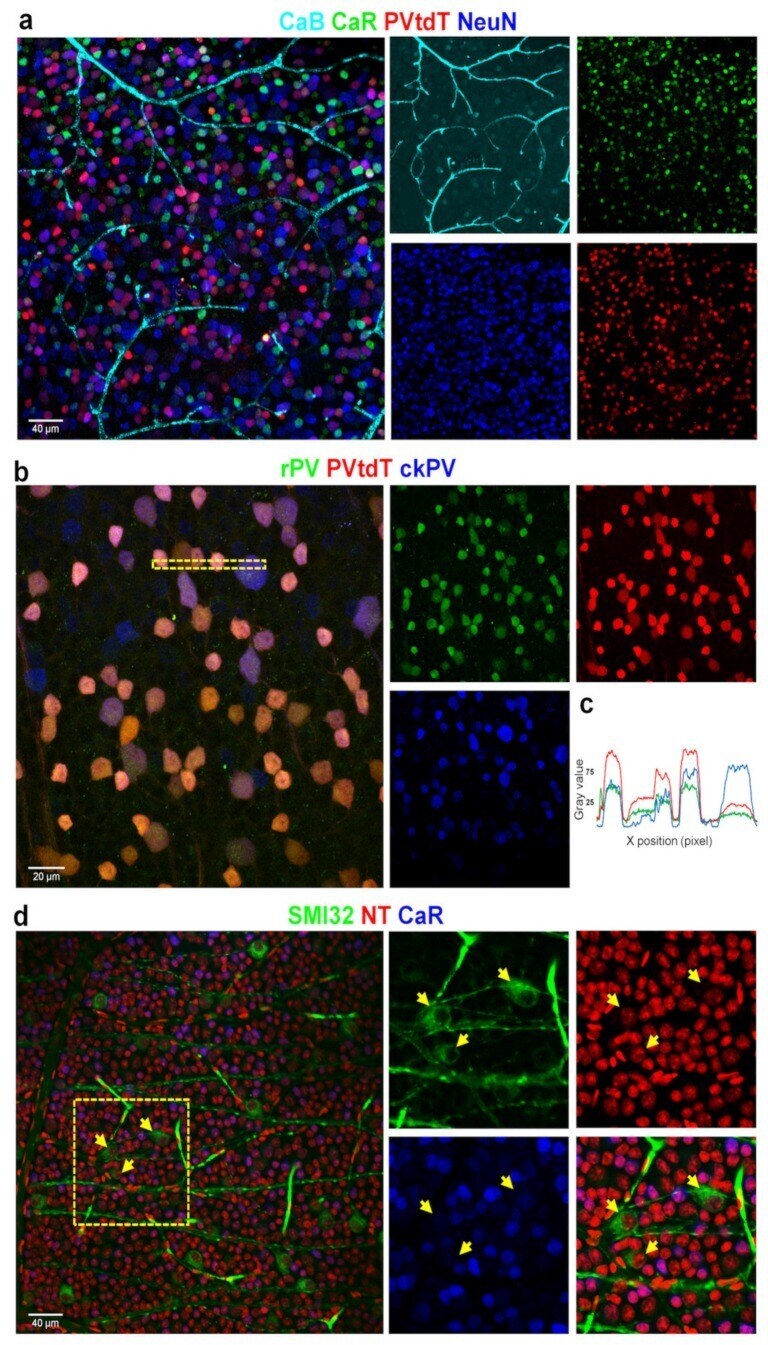
- Figure 8 Correlating multiple labels with CaBPs can help reveal individual differences between antibodies and GC identity. Panel ( a ) shows the parvalbumin-tdTomato (PV-tdT) GMO mouse retina with CaB, CaR, and NeuN labeling (composite image on the left and split into four channels on the right--all colors correspond to the label above). Panel ( b ) displays the comparison between PV-tdT, rabbit-PV (rPV), and chicken-PV (ckPV) antibody-labeling performed on the same retina. ( c ) A histogram profile, acquired from the dashed area in b shows a variance in normalized relative intensities of the PV-tdT (GMO, red) and r- (green), ckPV (blue) labels. ( d ) ON and OFF GCs (ganglion cells) are brightly labeled with SMI32 (arrows) but not by CaR. CaR, on the other hand, labels smaller GCs not labeled by SMI32. Neurons and vascular cells are all labeled with the fluoro-Nissl stain Neurotrace-640/660 (NT).
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
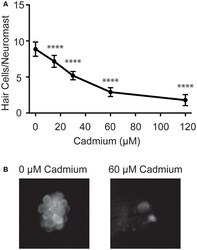
- Figure 1 Cadmium kills hair cells in zebrafish larvae in a dose-dependent manner. (A) Treating 5-days post-fertilization (dpf) zebrafish with cadmium chloride doses ranging from 0 to 120 muM for 3 h showed an increasing amount of hair cell death with increasing cadmium chloride dose. Data are displayed as mean +- standard deviation. **** p < 0.0001 as compared to 0 muM cadmium chloride control by ANOVA and Dunnett's post hoc test. Fish were fixed and stained with an anti-parvalbumin antibody and hair cells from six neuromasts from each fish were counted (OP1, M2, IO4, O2, MI2, and MI1) and averaged. n = 10 fish for the 0, 15, 60 and 120 muM cadmium chloride groups and n = 7 for the 30 muM cadmium chloride group. (B) Representative images of the MI1 neuromast following treatment with either 0 (left) or 60 (right) muM cadmium chloride.
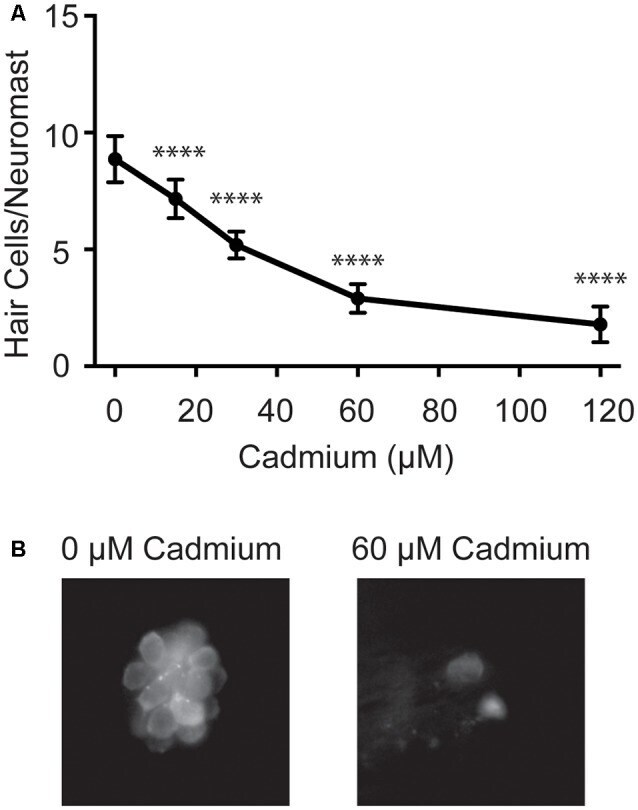
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
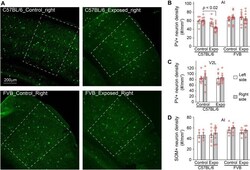
- Experimental details
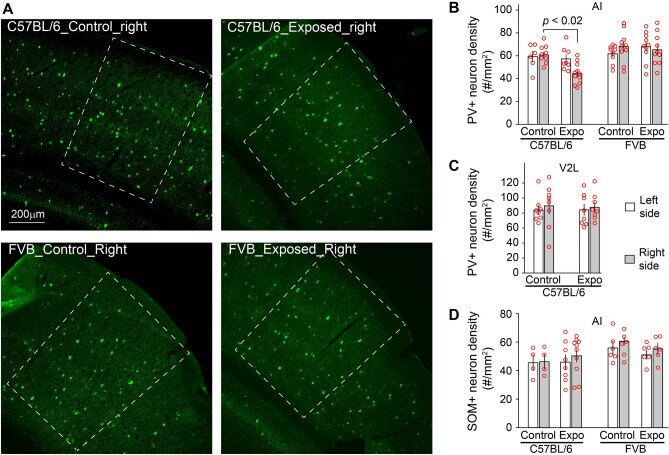
- Figure 4 Strain differences in noise-induced PV+ neuron loss. ( A ) Example images of PV+ neurons in AI of the right hemisphere, which is delimited with white dashed lines. ( B ) The density of PV+ neurons was significantly reduced in the right AI of C57BL/6 but not FVB mice 5 days after noise exposure. ( C ) The reduction of PV+ neuron density was not observed in lateral area of the secondary visual cortex (V2L). ( D ) The density of SOM+ neurons was not significantly altered in AI of C57BL/6 or FVB mice 5 days after noise exposure.