Explore
Explore Validate
Validate Learn
Learn13-7300
antibody from Invitrogen Antibodies
Targeting: CAMK2A
CAMKA, CaMKIINalpha, CaMKIIα, KIAA0968
 Western blot
Western blotAntibody data
- Antibody Data
- Antigen structure
- References [21]
- Comments [0]
- Validations
- Western blot [4]
- Other assay [17]
Submit
Validation data
Reference
Comment
Report error
- Product number
- 13-7300 - Provider product page

- Provider
- Invitrogen Antibodies
- Product name
- CaMKII alpha Monoclonal Antibody (Cba-2)
- Antibody type
- Monoclonal
- Antigen
- Purifed from natural sources
- Description
- This antibody is specific for alpha subunit of CaM Kinase II. No significant reactivity was seen with the B, B1, yB, dA, dB, and dC isoforms. The epitope is located in the regulatory domain.
- Antibody clone number
- Cba-2
- Concentration
- 0.5 mg/mL
Submitted references CAMK2/CaMKII activates MLKL in short-term starvation to facilitate autophagic flux.
Dynamic bi-directional phosphorylation events associated with the reciprocal regulation of synapses during homeostatic up- and down-scaling.
The Developmental Shift of NMDA Receptor Composition Proceeds Independently of GluN2 Subunit-Specific GluN2 C-Terminal Sequences.
Calmodulin shuttling mediates cytonuclear signaling to trigger experience-dependent transcription and memory.
Reciprocal control of excitatory synapse numbers by Wnt and Wnt inhibitor PRR7 secreted on exosomes.
Synaptic GAP and GEF Complexes Cluster Proteins Essential for GTP Signaling.
Nogo-A regulates spatial learning as well as memory formation and modulates structural plasticity in the adult mouse hippocampus.
Dopamine neuronal loss contributes to memory and reward dysfunction in a model of Alzheimer's disease.
Chronic Hypobaric Hypoxia Induces Right Ventricular Hypertrophy and Apoptosis in Rats: Therapeutic Potential of Nanocurcumin in Improving Adaptation.
Melatonin stimulates dendrite formation and complexity in the hilar zone of the rat hippocampus: participation of the Ca++/Calmodulin complex.
Evolutionary and functional perspectives on signaling from neuronal surface to nucleus.
miR-26a and miR-384-5p are required for LTP maintenance and spine enlargement.
The schizophrenia susceptibility gene dysbindin regulates dendritic spine dynamics.
Calcium and calmodulin-dependent serine/threonine protein kinase type II (CaMKII)-mediated intramolecular opening of integrin cytoplasmic domain-associated protein-1 (ICAP-1α) negatively regulates β1 integrins.
Dual regulation of miRNA biogenesis generates target specificity in neurotrophin-induced protein synthesis.
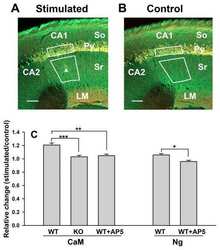
Stimulation-mediated translocation of calmodulin and neurogranin from soma to dendrites of mouse hippocampal CA1 pyramidal neurons.
Delayed treatment with a novel neurotrophic compound reduces behavioral deficits in rabbit ischemic stroke.
Netrin-1 induces axon branching in developing cortical neurons by frequency-dependent calcium signaling pathways.
Calcium-calmodulin-dependent protein kinase II contributes to spinal cord central sensitization.
PSD-95 assembles a ternary complex with the N-methyl-D-aspartic acid receptor and a bivalent neuronal NO synthase PDZ domain.
PSD-95 assembles a ternary complex with the N-methyl-D-aspartic acid receptor and a bivalent neuronal NO synthase PDZ domain.
Zhan Q, Jeon J, Li Y, Huang Y, Xiong J, Wang Q, Xu TL, Li Y, Ji FH, Du G, Zhu MX
Autophagy 2022 Apr;18(4):726-744
Autophagy 2022 Apr;18(4):726-744
Dynamic bi-directional phosphorylation events associated with the reciprocal regulation of synapses during homeostatic up- and down-scaling.
Desch K, Langer JD, Schuman EM
Cell reports 2021 Aug 24;36(8):109583
Cell reports 2021 Aug 24;36(8):109583
The Developmental Shift of NMDA Receptor Composition Proceeds Independently of GluN2 Subunit-Specific GluN2 C-Terminal Sequences.
McKay S, Ryan TJ, McQueen J, Indersmitten T, Marwick KFM, Hasel P, Kopanitsa MV, Baxter PS, Martel MA, Kind PC, Wyllie DJA, O'Dell TJ, Grant SGN, Hardingham GE, Komiyama NH
Cell reports 2018 Oct 23;25(4):841-851.e4
Cell reports 2018 Oct 23;25(4):841-851.e4
Calmodulin shuttling mediates cytonuclear signaling to trigger experience-dependent transcription and memory.
Cohen SM, Suutari B, He X, Wang Y, Sanchez S, Tirko NN, Mandelberg NJ, Mullins C, Zhou G, Wang S, Kats I, Salah A, Tsien RW, Ma H
Nature communications 2018 Jun 22;9(1):2451
Nature communications 2018 Jun 22;9(1):2451
Reciprocal control of excitatory synapse numbers by Wnt and Wnt inhibitor PRR7 secreted on exosomes.
Lee SH, Shin SM, Zhong P, Kim HT, Kim DI, Kim JM, Heo WD, Kim DW, Yeo CY, Kim CH, Liu QS
Nature communications 2018 Aug 24;9(1):3434
Nature communications 2018 Aug 24;9(1):3434
Synaptic GAP and GEF Complexes Cluster Proteins Essential for GTP Signaling.
Wilkinson B, Li J, Coba MP
Scientific reports 2017 Jul 13;7(1):5272
Scientific reports 2017 Jul 13;7(1):5272
Nogo-A regulates spatial learning as well as memory formation and modulates structural plasticity in the adult mouse hippocampus.
Zagrebelsky M, Lonnemann N, Fricke S, Kellner Y, Preuß E, Michaelsen-Preusse K, Korte M
Neurobiology of learning and memory 2017 Feb;138:154-163
Neurobiology of learning and memory 2017 Feb;138:154-163
Dopamine neuronal loss contributes to memory and reward dysfunction in a model of Alzheimer's disease.
Nobili A, Latagliata EC, Viscomi MT, Cavallucci V, Cutuli D, Giacovazzo G, Krashia P, Rizzo FR, Marino R, Federici M, De Bartolo P, Aversa D, Dell'Acqua MC, Cordella A, Sancandi M, Keller F, Petrosini L, Puglisi-Allegra S, Mercuri NB, Coccurello R, Berretta N, D'Amelio M
Nature communications 2017 Apr 3;8:14727
Nature communications 2017 Apr 3;8:14727
Chronic Hypobaric Hypoxia Induces Right Ventricular Hypertrophy and Apoptosis in Rats: Therapeutic Potential of Nanocurcumin in Improving Adaptation.
Nehra S, Bhardwaj V, Kar S, Saraswat D
High altitude medicine & biology 2016 Dec;17(4):342-352
High altitude medicine & biology 2016 Dec;17(4):342-352
Melatonin stimulates dendrite formation and complexity in the hilar zone of the rat hippocampus: participation of the Ca++/Calmodulin complex.
Domínguez-Alonso A, Valdés-Tovar M, Solís-Chagoyán H, Benítez-King G
International journal of molecular sciences 2015 Jan 16;16(1):1907-27
International journal of molecular sciences 2015 Jan 16;16(1):1907-27
Evolutionary and functional perspectives on signaling from neuronal surface to nucleus.
Cohen SM, Li B, Tsien RW, Ma H
Biochemical and biophysical research communications 2015 Apr 24;460(1):88-99
Biochemical and biophysical research communications 2015 Apr 24;460(1):88-99
miR-26a and miR-384-5p are required for LTP maintenance and spine enlargement.
Gu QH, Yu D, Hu Z, Liu X, Yang Y, Luo Y, Zhu J, Li Z
Nature communications 2015 Apr 10;6:6789
Nature communications 2015 Apr 10;6:6789
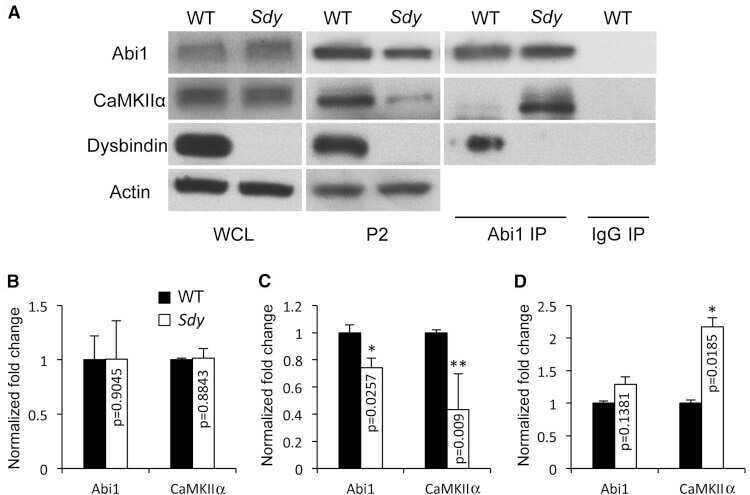
The schizophrenia susceptibility gene dysbindin regulates dendritic spine dynamics.
Jia JM, Hu Z, Nordman J, Li Z
The Journal of neuroscience : the official journal of the Society for Neuroscience 2014 Oct 8;34(41):13725-36
The Journal of neuroscience : the official journal of the Society for Neuroscience 2014 Oct 8;34(41):13725-36
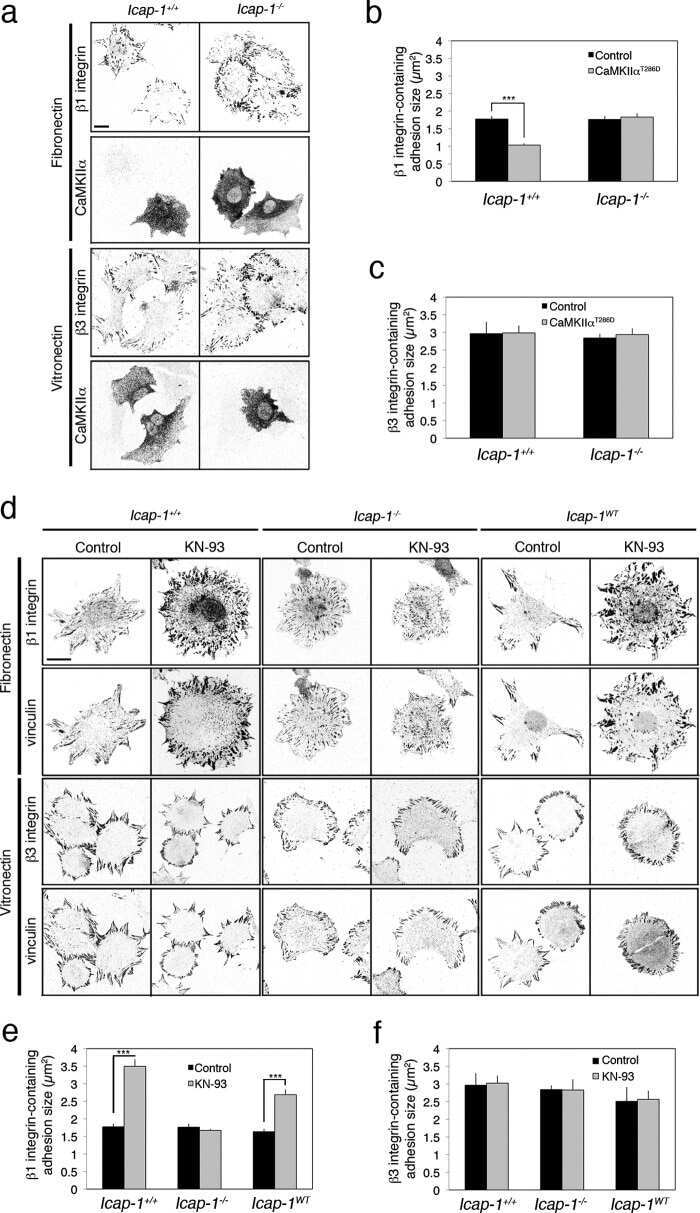
Calcium and calmodulin-dependent serine/threonine protein kinase type II (CaMKII)-mediated intramolecular opening of integrin cytoplasmic domain-associated protein-1 (ICAP-1α) negatively regulates β1 integrins.
Millon-Frémillon A, Brunner M, Abed N, Collomb E, Ribba AS, Block MR, Albigès-Rizo C, Bouvard D
The Journal of biological chemistry 2013 Jul 12;288(28):20248-60
The Journal of biological chemistry 2013 Jul 12;288(28):20248-60
Dual regulation of miRNA biogenesis generates target specificity in neurotrophin-induced protein synthesis.
Huang YW, Ruiz CR, Eyler EC, Lin K, Meffert MK
Cell 2012 Mar 2;148(5):933-46
Cell 2012 Mar 2;148(5):933-46
Stimulation-mediated translocation of calmodulin and neurogranin from soma to dendrites of mouse hippocampal CA1 pyramidal neurons.
Huang KP, Huang FL, Shetty PK
Neuroscience 2011 Mar 31;178:1-12
Neuroscience 2011 Mar 31;178:1-12
Delayed treatment with a novel neurotrophic compound reduces behavioral deficits in rabbit ischemic stroke.
Lapchak PA, Schubert DR, Maher PA
Journal of neurochemistry 2011 Jan;116(1):122-31
Journal of neurochemistry 2011 Jan;116(1):122-31
Netrin-1 induces axon branching in developing cortical neurons by frequency-dependent calcium signaling pathways.
Tang F, Kalil K
The Journal of neuroscience : the official journal of the Society for Neuroscience 2005 Jul 13;25(28):6702-15
The Journal of neuroscience : the official journal of the Society for Neuroscience 2005 Jul 13;25(28):6702-15
Calcium-calmodulin-dependent protein kinase II contributes to spinal cord central sensitization.
Fang L, Wu J, Lin Q, Willis WD
The Journal of neuroscience : the official journal of the Society for Neuroscience 2002 May 15;22(10):4196-204
The Journal of neuroscience : the official journal of the Society for Neuroscience 2002 May 15;22(10):4196-204
PSD-95 assembles a ternary complex with the N-methyl-D-aspartic acid receptor and a bivalent neuronal NO synthase PDZ domain.
Christopherson KS, Hillier BJ, Lim WA, Bredt DS
The Journal of biological chemistry 1999 Sep 24;274(39):27467-73
The Journal of biological chemistry 1999 Sep 24;274(39):27467-73
PSD-95 assembles a ternary complex with the N-methyl-D-aspartic acid receptor and a bivalent neuronal NO synthase PDZ domain.
Christopherson KS, Hillier BJ, Lim WA, Bredt DS
The Journal of biological chemistry 1999 Sep 24;274(39):27467-73
The Journal of biological chemistry 1999 Sep 24;274(39):27467-73
No comments: Submit comment
Supportive validation
- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
- Western blot analysis was performed on tissue extracts (30 µg lysate) of Rat Brain (Lane 1), and Mouse Brain (Lane 2). The blots were probed with Anti-CaM Kinase II alpha Mouse Monoclonal Antibody (Product # 13-7300, 0.5-1 µg/mL) and detected by chemiluminescence using Goat anti-Mouse IgG (H+L) Secondary Antibody, HRP conjµgate (Product # 62-6520, 1:4000 dilution). A ~ 54 kDa band corresponding to CaM Kinase II alpha was observed across tissues tested. Known quantity of protein samples were electrophoresed using Novex® NuPAGE® 12 % Bis-Tris gel (Product # NP0342BOX), XCell SureLock™ Electrophoresis System (Product # EI0002) and Novex® Sharp Pre-Stained Protein Standard (Product # LC5800). Resolved proteins were then transferred onto a nitrocellulose membrane with iBlot® 2 Dry Blotting System (Product # IB21001). The membrane was probed with the relevant primary and secondary Antibody following blocking with 5 % skimmed milk. Chemiluminescent detection was performed using Pierce™ ECL Western Blotting Substrate (Product # 32106).

- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
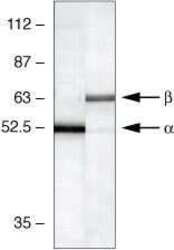
- Western blot using CaM Kinase II Alpha Monoclonal Antibody, Mouse: Western blot analysis of mouse brain lysates using CaM Kinase II Alpha Monoclonal Antibody, Mouse (lane 1, Product # 13-7300) and b-CAM Kinase II Monoclonal Antibody, Mouse (lane 2, Product # 13-9800).

- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
- Western blot analysis was performed on tissue extracts (30 µg lysate) of Rat Brain (Lane 1), and Mouse Brain (Lane 2). The blots were probed with Anti-CaM Kinase II alpha Mouse Monoclonal Antibody (Product # 13-7300, 0.5-1 µg/mL) and detected by chemiluminescence using Goat anti-Mouse IgG (H+L) Secondary Antibody, HRP conjµgate (Product # 62-6520, 1:4000 dilution). A ~ 54 kDa band corresponding to CaM Kinase II alpha was observed across tissues tested. Known quantity of protein samples were electrophoresed using Novex® NuPAGE® 12 % Bis-Tris gel (Product # NP0342BOX), XCell SureLock™ Electrophoresis System (Product # EI0002) and Novex® Sharp Pre-Stained Protein Standard (Product # LC5800). Resolved proteins were then transferred onto a nitrocellulose membrane with iBlot® 2 Dry Blotting System (Product # IB21001). The membrane was probed with the relevant primary and secondary Antibody following blocking with 5 % skimmed milk. Chemiluminescent detection was performed using Pierce™ ECL Western Blotting Substrate (Product # 32106).

- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
- Western blot was performed using Anti-CaMKII alpha Monoclonal Antibody (Cba-2) (Product # 13-7300) and a band at ~50 kDa corresponding to CaMKII alpha was observed in Mouse Brain in comparison to other tissues which are reported to be negative. Tissue extracts (30 µg lysate) of Mouse Brain (Lane 1), Mouse Skeletal Muscle (Lane 2) and Mouse Heart (Lane 3) were electrophoresed using Novex® NuPAGE® 4-12% % Bis-Tris gel (Product # NP0322BOX). Resolved proteins were then transferred onto a nitrocellulose membrane (Product # IB23001) by iBlot® 2 Dry Blotting System (Product # IB21001). The blot was probed with the primary antibody (1µg/ml) and detected by chemiluminescence with Goat anti-Mouse IgG (H+L) Superclonal™ Recombinant Secondary Antibody, HRP (Product # A28177, 1:4000 dilution) using the iBright FL 1000 (Product # A32752). Chemiluminescent detection was performed using Novex® ECL Chemiluminescent Substrate Reagent Kit (Product # WP20005)..

Supportive validation
- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
- NULL

- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL

- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
- NULL

- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL

- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
- NULL

- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
- NULL

- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL

- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
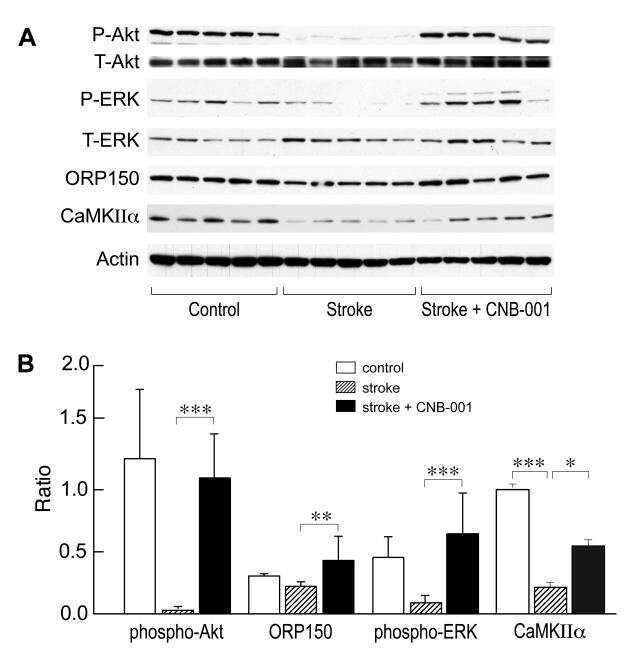
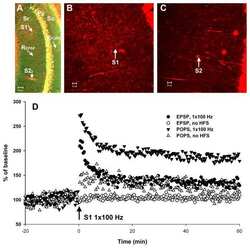
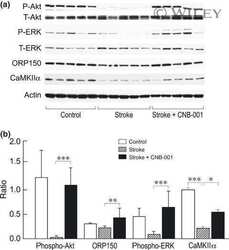
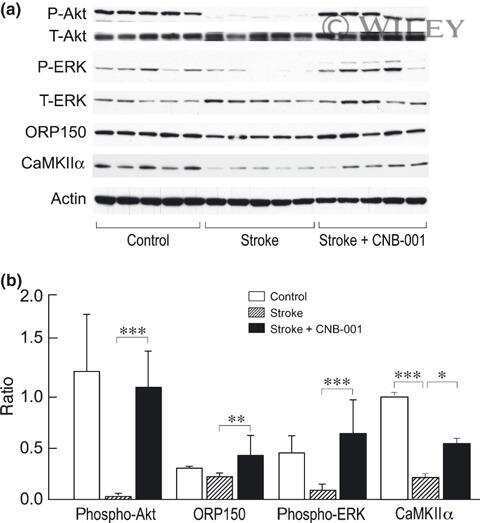
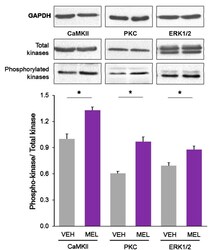
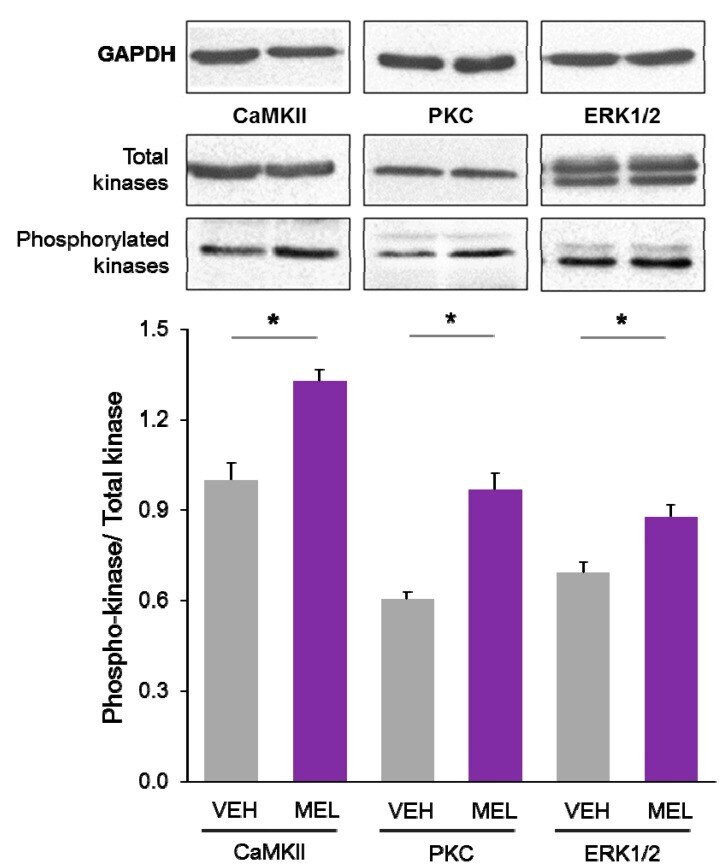
- Figure 4 Relative content of phosphorylated Ca 2+ /CaM Kinase II, Protein Kinase C and Extracellular Signal-Regulated Kinase 1/2 in hippocampal slices incubated with Melatonin. Hippocampal slices were incubated for 3 h with either the vehicle (VEH) or 100 nM melatonin (MEL). CaMKII and Thr286-phospho-CaMKII (P-CaMKII), PKC and phospho-PKC (P-PKC) as well as ERK1/2 and phospho-ERK1/2 (P-ERK1/2) were determined by Western blot. Upper panels show immunodetection of GAPDH as load control. Representative fluorograms of total and phosphorylated enzymes revealed by ECL are shown below. Histograms correspond to densitometric analysis of bands depicted immediately above. Results are expressed as the mean +- SEM of four densitometric scannings obtained from two independent experiments. Asterisks indicate significant differences ( p < 0.05).
- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
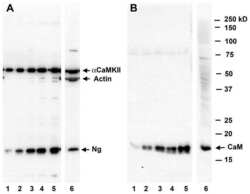
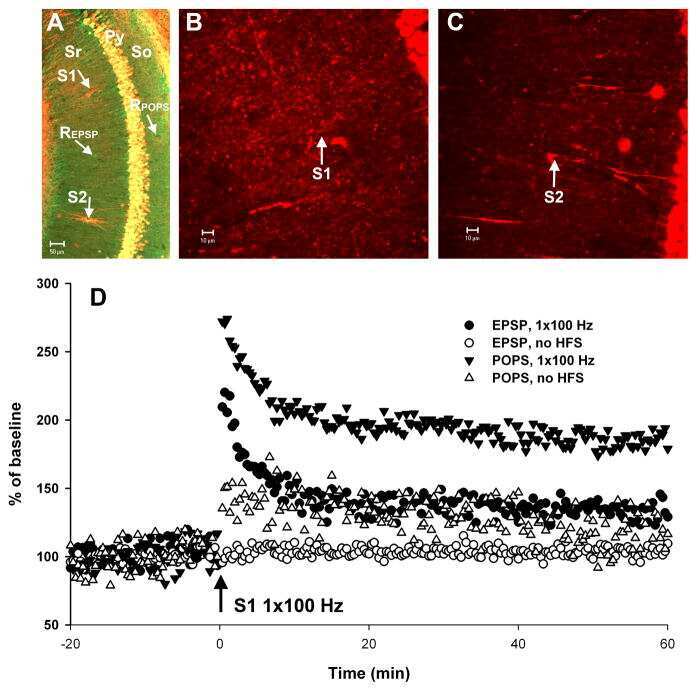
- Figure 5 Calmodulin content in hippocampal slices treated with Melatonin. Hippocampal slices were incubated during 6 h with either the vehicle (VEH) or melatonin (MEL). Calmodulin (CaM) in the homogenates ( A ); and in the soluble (SOL) and cytoskeletal fractions (CSK) separated by centrifugation ( B ); was determined by Western blot. Upper panels show Carbonic Anhydrase (CA) used as external load control and stained with Coomassie blue. Representative fluorograms of CaM are shown immediately below. First lane of both gel and fluorogram from panel A was loaded with pure CA (5 ug) and CaM (1 ug), respectively. CaM was recognized with a specific CaM antibody and ECL. Histograms correspond to densitometric analysis of the bands shown in the upper panels. Results are the mean +- SEM of four densitometric scannings obtained from two independent experiments. Asterisk indicates significant differences ( p < 0.05).

- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
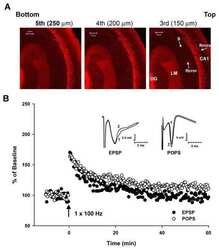
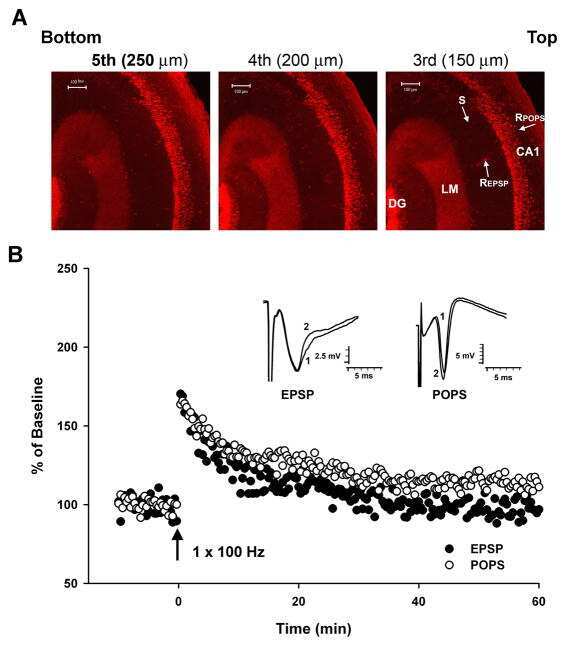
- Figure 4 Verification of phosphoregulated Camk2a, ERK1, and ERK2 by immunoblotting (A) Western blot (WB) images for pCamk2a (pThr 286 ) and total Camk2a. Analyses were performed using three independent biological replicates of Bic-treated, TTX-treated, and untreated (control) neurons for 5 min (top) or 24 h (bottom). (B) Bar graphs show quantification of pCamk2a detected by WB in comparison to the phosphopeptide's signals quantified via LC-MS/MS (MS) after 5 min (left) or 24 h (right) Bic or TTX stimulation. Protein signals of the blot were quantified based on the intensity ratio of the phosphorylated protein over total protein normalizing treated conditions to the untreated control. Log 2 -scaled MS intensity of the phosphorylated peptide is depicted as the fold change of treatment over control as well (see Method details ). Error bars represent the standard deviation between biological replicates (WB, n = 3; MS, n = 4). For statistical analysis of the WB data, a two-sided t test was performed: *** p > 0.001; ** p < 0.01; * p < 0.05;. p < 0.1; n.s., not significant. For MS-data, ** FDR < 0.01. (C) WB images for pERK1 (pThr 203 /pTyr 205 ) or pERK2 (pThr 183 /pTyr 185 ) and total ERK1 and ERK2. Analyses were performed using three independent biological replicates of Bic-treated, TTX-treated, and untreated (control) neurons for 5 min (left) or 24 h (right). (D) Bar graphs show quantification of pERK1 and pERK2 detected by WB in comparison to the phosphopeptides quantified via

- Submitted by
- Invitrogen Antibodies (provider)
- Main image
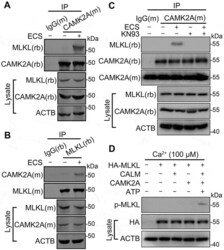
- Experimental details
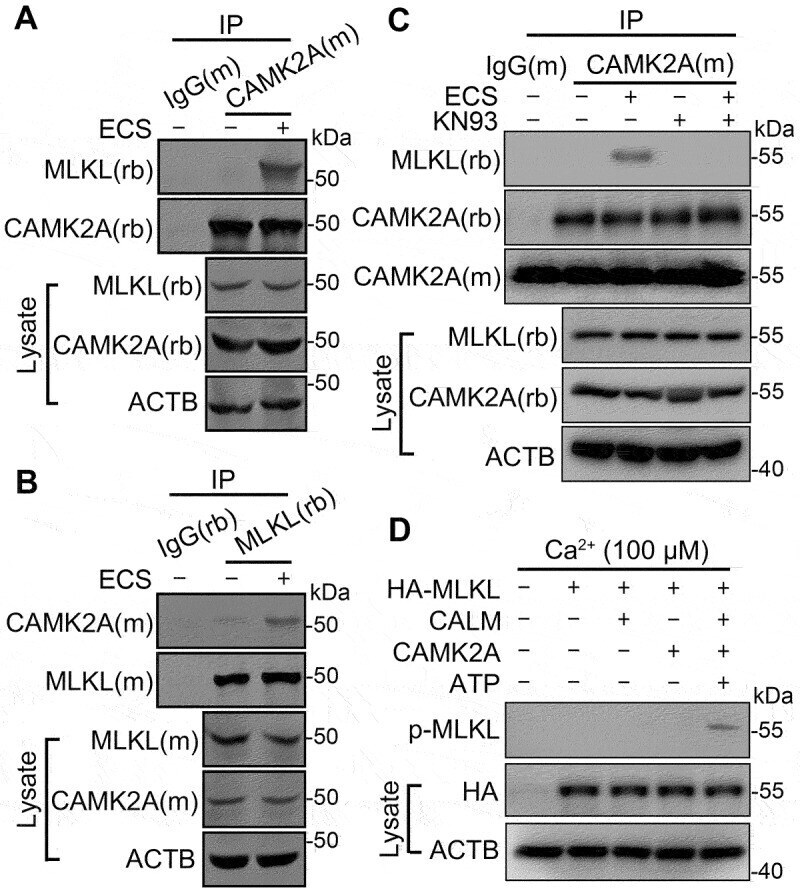
- Figure 3. Starvation induces CAMK2A-MLKL association in a protein complex. (A and B) Cell lysates from N2a cells untreated or treated with ECS for 1 h were subject to immunoprecipitation (IP) by IgG, or anti-CAMK2A (A) or anti-MLKL (B) antibody, followed by western blotting for MLKL and CAMK2A. (M) and (rb) denote the antibody host species as mouse and rabbit, respectively. The reciprocal co-immunoprecipitation showed increased association between CAMK2 and MLKL following starvation. (C) As in ( A ) but the cells were treated with KN93 (10 uM) as indicated. The blot by the mouse anti-CAMK2A antibody (CAMK2A[m]) shows the presence of antibody heavy chain in IP products from both IgG(m) and CAMK2A(m). However, the rabbit anti-CAMK2A antibody (CAMK2A[rb]) only revealed bands in this area for IP products from CAMK2A(m), but not that from IgG(m), indicating no cross reactivity of the secondary antibody. (D) In vitro kinase assay showing MLKL phosphorylation by recombinant CAMK2A. HA-MLKL purified by IP from transfected N2a cells was incubated with CALM, CAMK2A and ATP as indicated in a kinase buffer. The samples were subject to western blotting by the p-MLKL antibody. HA and ACTB were also measured from the cell lysates.