Explore
Explore Validate
Validate Learn
Learn Western blot
Western blot Immunocytochemistry
Immunocytochemistry Immunoprecipitation
ImmunoprecipitationAntibody data
- Antibody Data
- Antigen structure
- References [45]
- Comments [0]
- Validations
- Immunocytochemistry [8]
- Immunohistochemistry [2]
- Other assay [24]
Submit
Validation data
Reference
Comment
Report error
- Product number
- MA1-514 - Provider product page

- Provider
- Invitrogen Antibodies
- Product name
- AHR Monoclonal Antibody (RPT1)
- Antibody type
- Monoclonal
- Antigen
- Synthetic peptide
- Description
- MA1-514 detects the aryl hydrocarbon receptor (AhR) from human, mouse, and rat cells. MA1-514 has been successfully used in Western blotting, immunoprecipitation, immunofluorescence and immunohistochemistry procedures. By Western blot, this antibody detects an ~96 kDa protein representing AhR, with some minor non-specific bands observed in certain samples. MA1-514 immunogen is residues 12-31 from the human AhR protein with amino acids 18-21 being omitted.
- Reactivity
- Human, Mouse, Rat
- Host
- Mouse
- Isotype
- IgG
- Antibody clone number
- RPT1
- Vial size
- 100 μL
- Concentration
- 1 mg/mL
- Storage
- -20°C, Avoid Freeze/Thaw Cycles
Submitted references Intestinal Osteopontin Protects From Alcohol-induced Liver Injury by Preserving the Gut Microbiome and the Intestinal Barrier Function.
Anakinra restores cellular proteostasis by coupling mitochondrial redox balance to autophagy.
Impact of Epithelial-Mesenchymal Immunophenotype on Local Aggressiveness in Papillary Thyroid Carcinoma Invading the Airway.
Tomato and Olive Bioactive Compounds: A Natural Shield against the Cellular Effects Induced by β-Hexachlorocyclohexane-Activated Signaling Pathways.
β-Hexachlorocyclohexane: A Small Molecule with a Big Impact on Human Cellular Biochemistry.
Aryl Hydrocarbon Receptor in Post-Mortem Hippocampus and in Serum from Young, Elder, and Alzheimer's Patients.
Night Cream Containing Melatonin, Carnosine and Helichrysum italicum Extract Helps Reduce Skin Reactivity and Signs of Photodamage: Ex Vivo and Clinical Studies.
AHR is a Zika virus host factor and a candidate target for antiviral therapy.
Parkin is transcriptionally regulated by the aryl hydrocarbon receptor: Impact on α-synuclein protein levels.
Identification of the novel role of butyrate as AhR ligand in human intestinal epithelial cells.
Towards Resolving the Pro- and Anti-Tumor Effects of the Aryl Hydrocarbon Receptor.
Aryl Hydrocarbon Receptor Promotes Liver Polyploidization and Inhibits PI3K, ERK, and Wnt/β-Catenin Signaling.
Dioxins and related environmental contaminants increase TDP-43 levels.
Notch-regulated miR-223 targets the aryl hydrocarbon receptor pathway and increases cytokine production in macrophages from rheumatoid arthritis patients.
piRNA-associated proteins and retrotransposons are differentially expressed in murine testis and ovary of aryl hydrocarbon receptor deficient mice.
Detrusor induction of miR-132/212 following bladder outlet obstruction: association with MeCP2 repression and cell viability.
Induction of a chloracne phenotype in an epidermal equivalent model by 2,3,7,8-tetrachlorodibenzo-p-dioxin (TCDD) is dependent on aryl hydrocarbon receptor activation and is not reproduced by aryl hydrocarbon receptor knock down.
HER2 overexpression-mediated inflammatory signaling enhances mammosphere formation through up-regulation of aryl hydrocarbon receptor transcription.
Role of the Ah receptor in homeostatic control of fatty acid synthesis in the liver.
Aryl hydrocarbon receptor regulates the cholesterol biosynthetic pathway in a dioxin response element-independent manner.
Activation of the aryl hydrocarbon receptor represses mammosphere formation in MCF-7 cells.
Maternal smoking and fetal sex significantly affect metabolic enzyme expression in the human fetal liver.
Dioxin receptor and SLUG transcription factors regulate the insulator activity of B1 SINE retrotransposons via an RNA polymerase switch.
Suppression mechanisms of flavonoids on aryl hydrocarbon receptor-mediated signal transduction.
Antagonism of aryl hydrocarbon receptor signaling by 6,2',4'-trimethoxyflavone.
Evidence for ligand-mediated selective modulation of aryl hydrocarbon receptor activity.
Differential gene regulation by the human and mouse aryl hydrocarbon receptor.
Inhibition of P-glycoprotein enhances the suppressive effect of kaempferol on transformation of the aryl hydrocarbon receptor.
Cacao polyphenol extract suppresses transformation of an aryl hydrocarbon receptor in C57BL/6 mice.
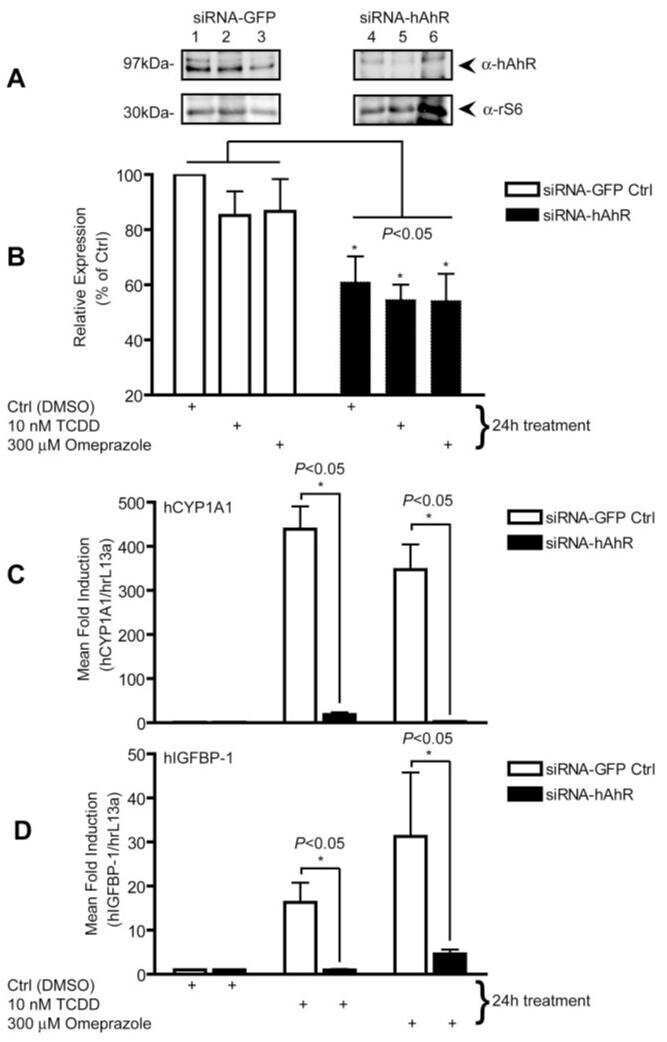
Omeprazole stimulates the induction of human insulin-like growth factor binding protein-1 through aryl hydrocarbon receptor activation.
The mouse and human Ah receptor differ in recognition of LXXLL motifs.
Characterization of the antiallergic drugs 3-[2-(2-phenylethyl) benzoimidazole-4-yl]-3-hydroxypropanoic acid and ethyl 3-hydroxy-3-[2-(2-phenylethyl)benzoimidazol-4-yl]propanoate as full aryl hydrocarbon receptor agonists.
Genome-wide B1 retrotransposon binds the transcription factors dioxin receptor and Slug and regulates gene expression in vivo.
AhR-agonist-induced transcriptional changes of genes involved in thyroid function in primary porcine thyrocytes.
Differentiation-specific factors modulate epidermal CYP1-4 gene expression in human skin in response to retinoic acid and classic aryl hydrocarbon receptor ligands.
Dioxin exerts anti-estrogenic actions in a novel dioxin-responsive telomerase-immortalized epithelial cell line of the porcine oviduct (TERT-OPEC).
Dioxin-induced immortalization of normal human keratinocytes and silencing of p53 and p16INK4a.
HES-1, a novel target gene for the aryl hydrocarbon receptor.
A new southwestern chemistry-based ELISA for detection of aryl hydrocarbon receptor transformation: application to the screening of its receptor agonists and antagonists.
Characterization of the phosphorylation status of the hepatitis B virus X-associated protein 2.
Characterization of the phosphorylation status of the hepatitis B virus X-associated protein 2.
Characterization of the AhR-hsp90-XAP2 core complex and the role of the immunophilin-related protein XAP2 in AhR stabilization.
Expression of the aryl hydrocarbon receptor is regulated by serum and mitogenic growth factors in murine 3T3 fibroblasts.
Expression of the aryl hydrocarbon receptor is regulated by serum and mitogenic growth factors in murine 3T3 fibroblasts.
Subunit composition of the heteromeric cytosolic aryl hydrocarbon receptor complex.
Das S, Song Z, Han H, Ge X, Desert R, Athavale D, Babu Komakula SS, Magdaleno F, Chen W, Lantvit D, Guzman G, Nieto N
Cellular and molecular gastroenterology and hepatology 2022;14(4):813-839
Cellular and molecular gastroenterology and hepatology 2022;14(4):813-839
Anakinra restores cellular proteostasis by coupling mitochondrial redox balance to autophagy.
van de Veerdonk FL, Renga G, Pariano M, Bellet MM, Servillo G, Fallarino F, De Luca A, Iannitti RG, Piobbico D, Gargaro M, Manni G, D'Onofrio F, Stincardini C, Sforna L, Borghi M, Castelli M, Pieroni S, Oikonomou V, Villella VR, Puccetti M, Giovagnoli S, Galarini R, Barola C, Maiuri L, Della Fazia MA, Cellini B, Talesa VN, Dinarello CA, Costantini C, Romani L
The Journal of clinical investigation 2022 Jan 18;132(2)
The Journal of clinical investigation 2022 Jan 18;132(2)
Impact of Epithelial-Mesenchymal Immunophenotype on Local Aggressiveness in Papillary Thyroid Carcinoma Invading the Airway.
Mandarano M, Andolfi M, Colella R, Monacelli M, Polistena A, Moretti S, Bellezza G, Puxeddu E, Sanguinetti A, Sidoni A, Avenia N, Puma F, Vannucci J
Journal of clinical medicine 2021 Sep 24;10(19)
Journal of clinical medicine 2021 Sep 24;10(19)
Tomato and Olive Bioactive Compounds: A Natural Shield against the Cellular Effects Induced by β-Hexachlorocyclohexane-Activated Signaling Pathways.
Rubini E, Minacori M, Paglia G, Macone A, Chichiarelli S, Altieri F, Eufemi M
Molecules (Basel, Switzerland) 2021 Nov 25;26(23)
Molecules (Basel, Switzerland) 2021 Nov 25;26(23)
β-Hexachlorocyclohexane: A Small Molecule with a Big Impact on Human Cellular Biochemistry.
Rubini E, Paglia G, Cannella D, Macone A, Di Sotto A, Gullì M, Altieri F, Eufemi M
Biomedicines 2020 Nov 16;8(11)
Biomedicines 2020 Nov 16;8(11)
Aryl Hydrocarbon Receptor in Post-Mortem Hippocampus and in Serum from Young, Elder, and Alzheimer's Patients.
Ramos-García NA, Orozco-Ibarra M, Estudillo E, Elizondo G, Gómez Apo E, Chávez Macías LG, Sosa-Ortiz AL, Torres-Ramos MA
International journal of molecular sciences 2020 Mar 14;21(6)
International journal of molecular sciences 2020 Mar 14;21(6)
Night Cream Containing Melatonin, Carnosine and Helichrysum italicum Extract Helps Reduce Skin Reactivity and Signs of Photodamage: Ex Vivo and Clinical Studies.
Granger C, Brown A, Aladren S, Narda M
Dermatology and therapy 2020 Dec;10(6):1315-1329
Dermatology and therapy 2020 Dec;10(6):1315-1329
AHR is a Zika virus host factor and a candidate target for antiviral therapy.
Giovannoni F, Bosch I, Polonio CM, Torti MF, Wheeler MA, Li Z, Romorini L, Rodriguez Varela MS, Rothhammer V, Barroso A, Tjon EC, Sanmarco LM, Takenaka MC, Modaresi SMS, Gutiérrez-Vázquez C, Zanluqui NG, Dos Santos NB, Munhoz CD, Wang Z, Damonte EB, Sherr D, Gehrke L, Peron JPS, Garcia CC, Quintana FJ
Nature neuroscience 2020 Aug;23(8):939-951
Nature neuroscience 2020 Aug;23(8):939-951
Parkin is transcriptionally regulated by the aryl hydrocarbon receptor: Impact on α-synuclein protein levels.
González-Barbosa E, García-Aguilar R, Vega L, Cabañas-Cortés MA, Gonzalez FJ, Segovia J, Morales-Lázaro SL, Cisneros B, Elizondo G
Biochemical pharmacology 2019 Oct;168:429-437
Biochemical pharmacology 2019 Oct;168:429-437
Identification of the novel role of butyrate as AhR ligand in human intestinal epithelial cells.
Marinelli L, Martin-Gallausiaux C, Bourhis JM, Béguet-Crespel F, Blottière HM, Lapaque N
Scientific reports 2019 Jan 24;9(1):643
Scientific reports 2019 Jan 24;9(1):643
Towards Resolving the Pro- and Anti-Tumor Effects of the Aryl Hydrocarbon Receptor.
Narasimhan S, Stanford Zulick E, Novikov O, Parks AJ, Schlezinger JJ, Wang Z, Laroche F, Feng H, Mulas F, Monti S, Sherr DH
International journal of molecular sciences 2018 May 7;19(5)
International journal of molecular sciences 2018 May 7;19(5)
Aryl Hydrocarbon Receptor Promotes Liver Polyploidization and Inhibits PI3K, ERK, and Wnt/β-Catenin Signaling.
Moreno-Marín N, Merino JM, Alvarez-Barrientos A, Patel DP, Takahashi S, González-Sancho JM, Gandolfo P, Rios RM, Muñoz A, Gonzalez FJ, Fernández-Salguero PM
iScience 2018 Jun 29;4:44-63
iScience 2018 Jun 29;4:44-63
Dioxins and related environmental contaminants increase TDP-43 levels.
Ash PEA, Stanford EA, Al Abdulatif A, Ramirez-Cardenas A, Ballance HI, Boudeau S, Jeh A, Murithi JM, Tripodis Y, Murphy GJ, Sherr DH, Wolozin B
Molecular neurodegeneration 2017 May 5;12(1):35
Molecular neurodegeneration 2017 May 5;12(1):35
Notch-regulated miR-223 targets the aryl hydrocarbon receptor pathway and increases cytokine production in macrophages from rheumatoid arthritis patients.
Ogando J, Tardáguila M, Díaz-Alderete A, Usategui A, Miranda-Ramos V, Martínez-Herrera DJ, de la Fuente L, García-León MJ, Moreno MC, Escudero S, Cañete JD, Toribio ML, Cases I, Pascual-Montano A, Pablos JL, Mañes S
Scientific reports 2016 Feb 3;6:20223
Scientific reports 2016 Feb 3;6:20223
piRNA-associated proteins and retrotransposons are differentially expressed in murine testis and ovary of aryl hydrocarbon receptor deficient mice.
Rico-Leo EM, Moreno-Marín N, González-Rico FJ, Barrasa E, Ortega-Ferrusola C, Martín-Muñoz P, Sánchez-Guardado LO, Llano E, Alvarez-Barrientos A, Infante-Campos A, Catalina-Fernández I, Hidalgo-Sánchez M, de Rooij DG, Pendás AM, Peña FJ, Merino JM, Fernández-Salguero PM
Open biology 2016 Dec;6(12)
Open biology 2016 Dec;6(12)
Detrusor induction of miR-132/212 following bladder outlet obstruction: association with MeCP2 repression and cell viability.
Sadegh MK, Ekman M, Krawczyk K, Svensson D, Göransson O, Dahan D, Nilsson BO, Albinsson S, Uvelius B, Swärd K
PloS one 2015;10(1):e0116784
PloS one 2015;10(1):e0116784
Induction of a chloracne phenotype in an epidermal equivalent model by 2,3,7,8-tetrachlorodibenzo-p-dioxin (TCDD) is dependent on aryl hydrocarbon receptor activation and is not reproduced by aryl hydrocarbon receptor knock down.
Forrester AR, Elias MS, Woodward EL, Graham M, Williams FM, Reynolds NJ
Journal of dermatological science 2014 Jan;73(1):10-22
Journal of dermatological science 2014 Jan;73(1):10-22
HER2 overexpression-mediated inflammatory signaling enhances mammosphere formation through up-regulation of aryl hydrocarbon receptor transcription.
Zhao S, Ohara S, Kanno Y, Midorikawa Y, Nakayama M, Makimura M, Park Y, Inouye Y
Cancer letters 2013 Mar 1;330(1):41-8
Cancer letters 2013 Mar 1;330(1):41-8
Role of the Ah receptor in homeostatic control of fatty acid synthesis in the liver.
Tanos R, Murray IA, Smith PB, Patterson A, Perdew GH
Toxicological sciences : an official journal of the Society of Toxicology 2012 Oct;129(2):372-9
Toxicological sciences : an official journal of the Society of Toxicology 2012 Oct;129(2):372-9
Aryl hydrocarbon receptor regulates the cholesterol biosynthetic pathway in a dioxin response element-independent manner.
Tanos R, Patel RD, Murray IA, Smith PB, Patterson AD, Perdew GH
Hepatology (Baltimore, Md.) 2012 Jun;55(6):1994-2004
Hepatology (Baltimore, Md.) 2012 Jun;55(6):1994-2004
Activation of the aryl hydrocarbon receptor represses mammosphere formation in MCF-7 cells.
Zhao S, Kanno Y, Nakayama M, Makimura M, Ohara S, Inouye Y
Cancer letters 2012 Apr 28;317(2):192-8
Cancer letters 2012 Apr 28;317(2):192-8
Maternal smoking and fetal sex significantly affect metabolic enzyme expression in the human fetal liver.
O'Shaughnessy PJ, Monteiro A, Bhattacharya S, Fowler PA
The Journal of clinical endocrinology and metabolism 2011 Sep;96(9):2851-60
The Journal of clinical endocrinology and metabolism 2011 Sep;96(9):2851-60
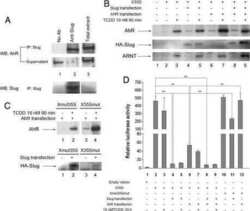
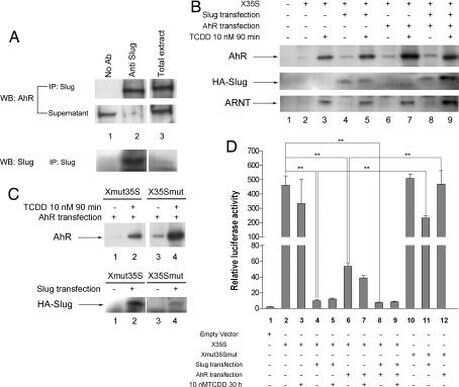
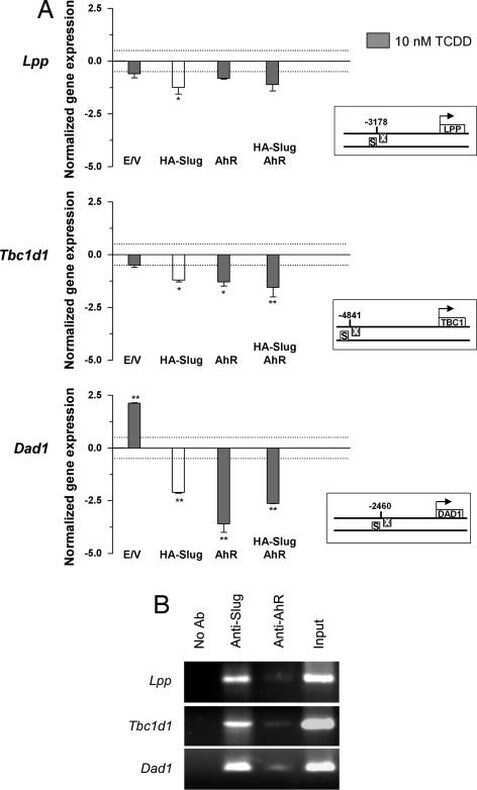
Dioxin receptor and SLUG transcription factors regulate the insulator activity of B1 SINE retrotransposons via an RNA polymerase switch.
Román AC, González-Rico FJ, Moltó E, Hernando H, Neto A, Vicente-Garcia C, Ballestar E, Gómez-Skarmeta JL, Vavrova-Anderson J, White RJ, Montoliu L, Fernández-Salguero PM
Genome research 2011 Mar;21(3):422-32
Genome research 2011 Mar;21(3):422-32
Suppression mechanisms of flavonoids on aryl hydrocarbon receptor-mediated signal transduction.
Mukai R, Shirai Y, Saito N, Fukuda I, Nishiumi S, Yoshida K, Ashida H
Archives of biochemistry and biophysics 2010 Sep 1;501(1):134-41
Archives of biochemistry and biophysics 2010 Sep 1;501(1):134-41
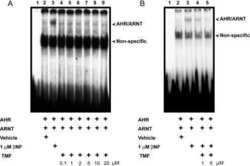
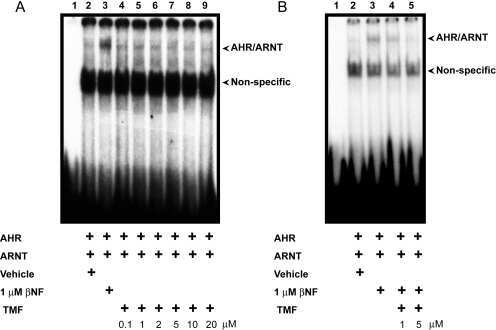
Antagonism of aryl hydrocarbon receptor signaling by 6,2',4'-trimethoxyflavone.
Murray IA, Flaveny CA, DiNatale BC, Chairo CR, Schroeder JC, Kusnadi A, Perdew GH
The Journal of pharmacology and experimental therapeutics 2010 Jan;332(1):135-44
The Journal of pharmacology and experimental therapeutics 2010 Jan;332(1):135-44
Evidence for ligand-mediated selective modulation of aryl hydrocarbon receptor activity.
Murray IA, Morales JL, Flaveny CA, Dinatale BC, Chiaro C, Gowdahalli K, Amin S, Perdew GH
Molecular pharmacology 2010 Feb;77(2):247-54
Molecular pharmacology 2010 Feb;77(2):247-54
Differential gene regulation by the human and mouse aryl hydrocarbon receptor.
Flaveny CA, Murray IA, Perdew GH
Toxicological sciences : an official journal of the Society of Toxicology 2010 Apr;114(2):217-25
Toxicological sciences : an official journal of the Society of Toxicology 2010 Apr;114(2):217-25
Inhibition of P-glycoprotein enhances the suppressive effect of kaempferol on transformation of the aryl hydrocarbon receptor.
Mukai R, Satsu H, Shimizu M, Ashida H
Bioscience, biotechnology, and biochemistry 2009 Jul;73(7):1635-9
Bioscience, biotechnology, and biochemistry 2009 Jul;73(7):1635-9
Cacao polyphenol extract suppresses transformation of an aryl hydrocarbon receptor in C57BL/6 mice.
Mukai R, Fukuda I, Nishiumi S, Natsume M, Osakabe N, Yoshida K, Ashida H
Journal of agricultural and food chemistry 2008 Nov 12;56(21):10399-405
Journal of agricultural and food chemistry 2008 Nov 12;56(21):10399-405
Omeprazole stimulates the induction of human insulin-like growth factor binding protein-1 through aryl hydrocarbon receptor activation.
Murray IA, Perdew GH
The Journal of pharmacology and experimental therapeutics 2008 Mar;324(3):1102-10
The Journal of pharmacology and experimental therapeutics 2008 Mar;324(3):1102-10
The mouse and human Ah receptor differ in recognition of LXXLL motifs.
Flaveny C, Reen RK, Kusnadi A, Perdew GH
Archives of biochemistry and biophysics 2008 Mar 15;471(2):215-23
Archives of biochemistry and biophysics 2008 Mar 15;471(2):215-23
Characterization of the antiallergic drugs 3-[2-(2-phenylethyl) benzoimidazole-4-yl]-3-hydroxypropanoic acid and ethyl 3-hydroxy-3-[2-(2-phenylethyl)benzoimidazol-4-yl]propanoate as full aryl hydrocarbon receptor agonists.
Morales JL, Krzeminski J, Amin S, Perdew GH
Chemical research in toxicology 2008 Feb;21(2):472-82
Chemical research in toxicology 2008 Feb;21(2):472-82
Genome-wide B1 retrotransposon binds the transcription factors dioxin receptor and Slug and regulates gene expression in vivo.
Roman AC, Benitez DA, Carvajal-Gonzalez JM, Fernandez-Salguero PM
Proceedings of the National Academy of Sciences of the United States of America 2008 Feb 5;105(5):1632-7
Proceedings of the National Academy of Sciences of the United States of America 2008 Feb 5;105(5):1632-7
AhR-agonist-induced transcriptional changes of genes involved in thyroid function in primary porcine thyrocytes.
Pocar P, Klonisch T, Brandsch C, Eder K, Fröhlich C, Hoang-Vu C, Hombach-Klonisch S
Toxicological sciences : an official journal of the Society of Toxicology 2006 Feb;89(2):408-14
Toxicological sciences : an official journal of the Society of Toxicology 2006 Feb;89(2):408-14
Differentiation-specific factors modulate epidermal CYP1-4 gene expression in human skin in response to retinoic acid and classic aryl hydrocarbon receptor ligands.
Du L, Neis MM, Ladd PA, Keeney DS
The Journal of pharmacology and experimental therapeutics 2006 Dec;319(3):1162-71
The Journal of pharmacology and experimental therapeutics 2006 Dec;319(3):1162-71
Dioxin exerts anti-estrogenic actions in a novel dioxin-responsive telomerase-immortalized epithelial cell line of the porcine oviduct (TERT-OPEC).
Hombach-Klonisch S, Pocar P, Kauffold J, Klonisch T
Toxicological sciences : an official journal of the Society of Toxicology 2006 Apr;90(2):519-28
Toxicological sciences : an official journal of the Society of Toxicology 2006 Apr;90(2):519-28
Dioxin-induced immortalization of normal human keratinocytes and silencing of p53 and p16INK4a.
Ray SS, Swanson HI
The Journal of biological chemistry 2004 Jun 25;279(26):27187-93
The Journal of biological chemistry 2004 Jun 25;279(26):27187-93
HES-1, a novel target gene for the aryl hydrocarbon receptor.
Thomsen JS, Kietz S, Ström A, Gustafsson JA
Molecular pharmacology 2004 Jan;65(1):165-71
Molecular pharmacology 2004 Jan;65(1):165-71
A new southwestern chemistry-based ELISA for detection of aryl hydrocarbon receptor transformation: application to the screening of its receptor agonists and antagonists.
Fukuda I, Nishiumi S, Yabushita Y, Mukai R, Kodoi R, Hashizume K, Mizuno M, Hatanaka Y, Ashida H
Journal of immunological methods 2004 Apr;287(1-2):187-201
Journal of immunological methods 2004 Apr;287(1-2):187-201
Characterization of the phosphorylation status of the hepatitis B virus X-associated protein 2.
Dull AB, Carlson DB, Petrulis JR, Perdew GH
Archives of biochemistry and biophysics 2002 Oct 15;406(2):209-21
Archives of biochemistry and biophysics 2002 Oct 15;406(2):209-21
Characterization of the phosphorylation status of the hepatitis B virus X-associated protein 2.
Dull AB, Carlson DB, Petrulis JR, Perdew GH
Archives of biochemistry and biophysics 2002 Oct 15;406(2):209-21
Archives of biochemistry and biophysics 2002 Oct 15;406(2):209-21
Characterization of the AhR-hsp90-XAP2 core complex and the role of the immunophilin-related protein XAP2 in AhR stabilization.
Meyer BK, Perdew GH
Biochemistry 1999 Jul 13;38(28):8907-17
Biochemistry 1999 Jul 13;38(28):8907-17
Expression of the aryl hydrocarbon receptor is regulated by serum and mitogenic growth factors in murine 3T3 fibroblasts.
Vaziri C, Schneider A, Sherr DH, Faller DV
The Journal of biological chemistry 1996 Oct 18;271(42):25921-7
The Journal of biological chemistry 1996 Oct 18;271(42):25921-7
Expression of the aryl hydrocarbon receptor is regulated by serum and mitogenic growth factors in murine 3T3 fibroblasts.
Vaziri C, Schneider A, Sherr DH, Faller DV
The Journal of biological chemistry 1996 Oct 18;271(42):25921-7
The Journal of biological chemistry 1996 Oct 18;271(42):25921-7
Subunit composition of the heteromeric cytosolic aryl hydrocarbon receptor complex.
Chen HS, Perdew GH
The Journal of biological chemistry 1994 Nov 4;269(44):27554-8
The Journal of biological chemistry 1994 Nov 4;269(44):27554-8
No comments: Submit comment
Supportive validation
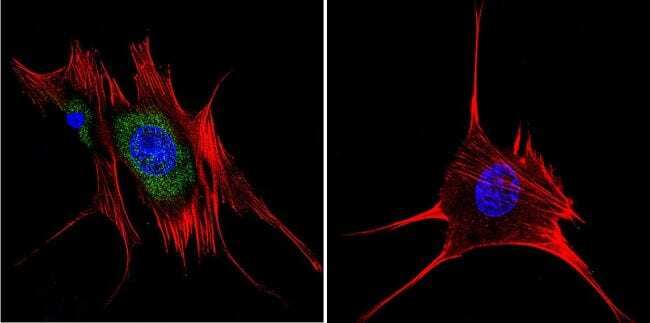
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
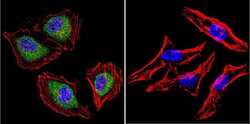
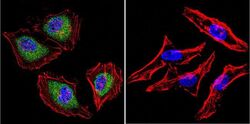
- Immunofluorescent analysis of Aryl Hydrocarbon Receptor in HeLa cells. Cells were grown on chamber slides and fixed with formaldehyde prior to staining. Cells were probed without (control) or with a Aryl Hydrocarbon Receptor monoclonal antibody (Product # MA1-514) at a dilution of 1:200 overnight at 4 C, washed with PBS and incubated with a DyLight-488 conjugated secondary antibody (Product # 35503). Aryl Hydrocarbon Receptor staining (green), F-Actin staining with Phalloidin (red) and nuclei with DAPI (blue) is shown. Images were taken at 60X magnification.
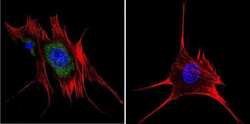
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
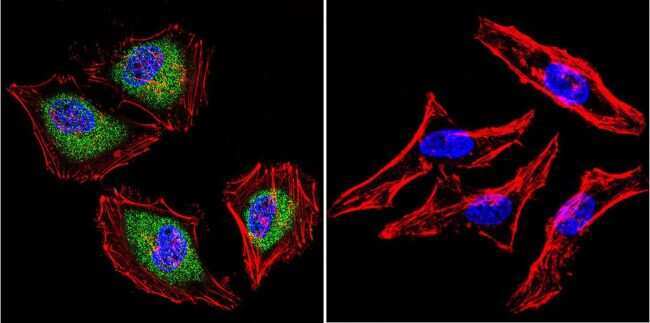
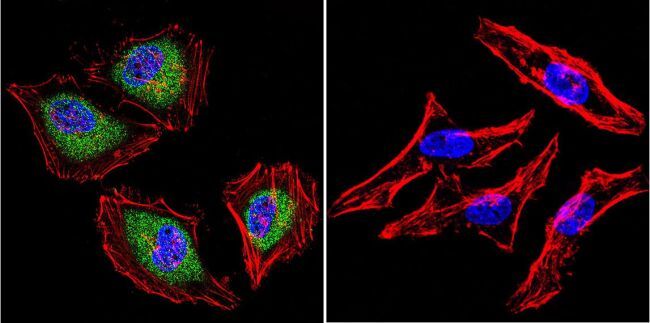
- Immunofluorescent analysis of Aryl Hydrocarbon Receptor in MCF-7 cells. Cells were grown on chamber slides and fixed with formaldehyde prior to staining. Cells were probed without (control) or with a Aryl Hydrocarbon Receptor monoclonal antibody (Product # MA1-514) at a dilution of 1:100 overnight at 4 C, washed with PBS and incubated with a DyLight-488 conjugated secondary antibody (Product # 35503). Aryl Hydrocarbon Receptor staining (green), F-Actin staining with Phalloidin (red) and nuclei with DAPI (blue) is shown. Images were taken at 60X magnification.
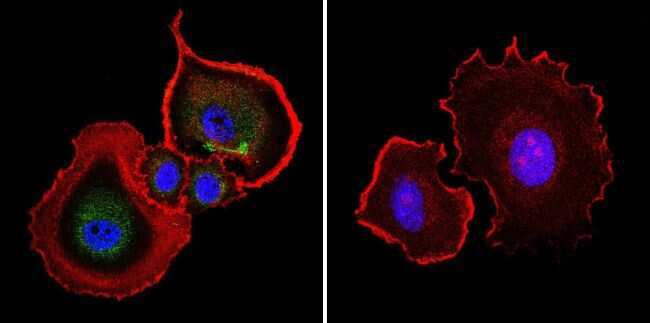
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
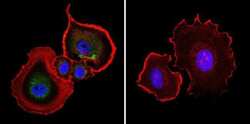
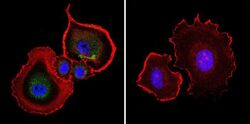
- Immunofluorescent analysis of Aryl Hydrocarbon Receptor in NIH-3T3 cells. Cells were grown on chamber slides and fixed with formaldehyde prior to staining. Cells were probed without (control) or with a Aryl Hydrocarbon Receptor monoclonal antibody (Product # MA1-514) at a dilution of 1:200 overnight at 4 C, washed with PBS and incubated with a DyLight-488 conjugated secondary antibody (Product # 35503). Aryl Hydrocarbon Receptor staining (green), F-Actin staining with Phalloidin (red) and nuclei with DAPI (blue) is shown. Images were taken at 60X magnification.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
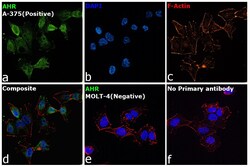
- Experimental details
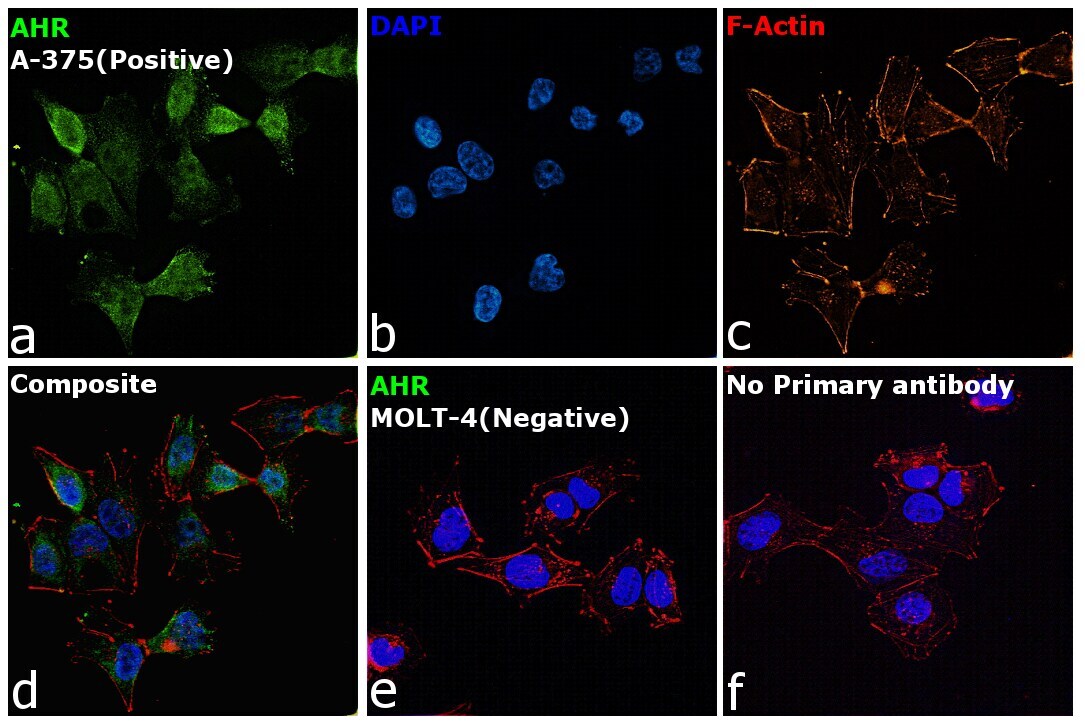
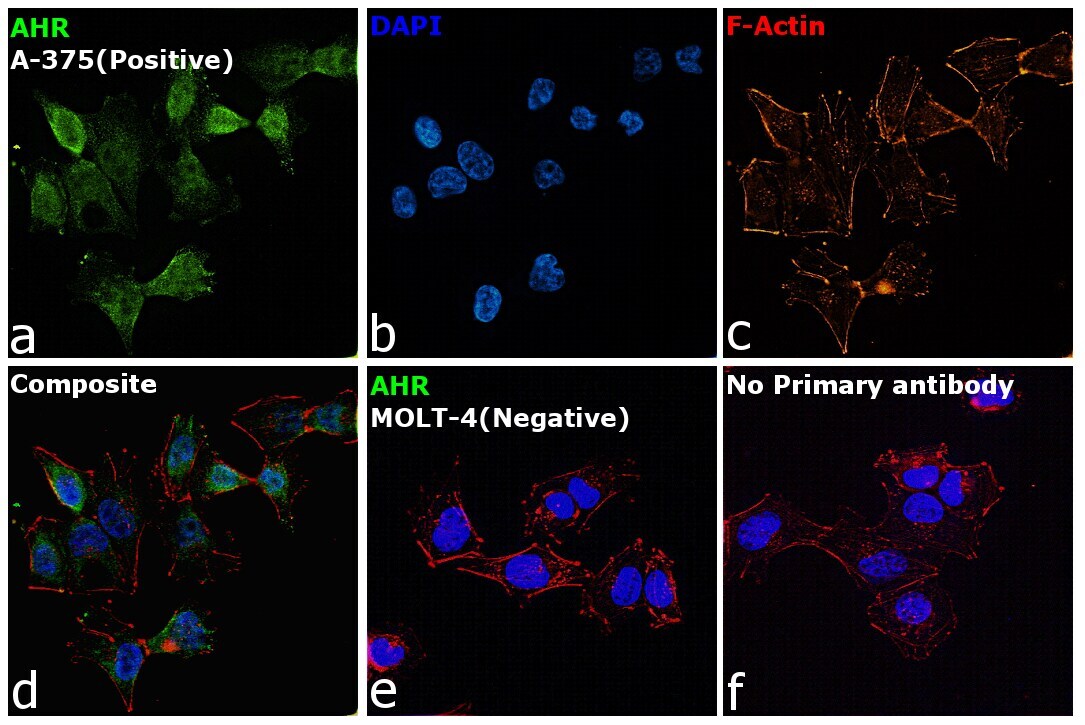
- Immunofluorescence analysis of AHR was performed using 70% confluent log phase A-375 cells. The cells were fixed with 4% Paraformaldehyde for 10 minutes, permeabilized with 0.1% Triton™ X-100 for 10 minutes, and blocked with 2% BSA for 10 minutes at room temperature. The cells were labeled with AHR Monoclonal Antibody (RPT1) (Product # MA1-514) at 1:200 dilution in 0.1% BSA, incubated at 4 degree Celsius overnight and then labeled with Goat anti-Mouse IgG (H+L), Superclonal™ Recombinant Secondary Antibody, Alexa Fluor 488 (Product # A28175), (1:2000 dilution) for 45 minutes at room temperature (Panel a: Green). Nuclei (Panel b: Blue) were stained with SlowFade® Gold Antifade Mountant with DAPI (Product # S36938). F-actin (Panel c: Red) was stained with Rhodamine Phalloidin (Product # R415, 1:300). Panel d represents the merged image showing Cytoplasmic localization. Panel e represents MOLT-4 cells having no expression of AHR. Panel f represents control cells with no primary antibody to assess background. The images were captured at 60X magnification.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
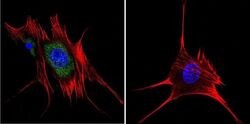
- Immunofluorescent analysis of Aryl Hydrocarbon Receptor in HeLa cells. Cells were grown on chamber slides and fixed with formaldehyde prior to staining. Cells were probed without (control) or with a Aryl Hydrocarbon Receptor monoclonal antibody (Product # MA1-514) at a dilution of 1:200 overnight at 4 C, washed with PBS and incubated with a DyLight-488 conjugated secondary antibody (Product # 35503). Aryl Hydrocarbon Receptor staining (green), F-Actin staining with Phalloidin (red) and nuclei with DAPI (blue) is shown. Images were taken at 60X magnification.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
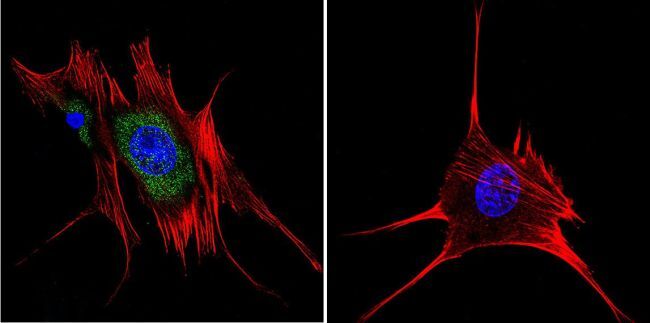
- Immunofluorescent analysis of Aryl Hydrocarbon Receptor in MCF-7 cells. Cells were grown on chamber slides and fixed with formaldehyde prior to staining. Cells were probed without (control) or with a Aryl Hydrocarbon Receptor monoclonal antibody (Product # MA1-514) at a dilution of 1:100 overnight at 4 C, washed with PBS and incubated with a DyLight-488 conjugated secondary antibody (Product # 35503). Aryl Hydrocarbon Receptor staining (green), F-Actin staining with Phalloidin (red) and nuclei with DAPI (blue) is shown. Images were taken at 60X magnification.
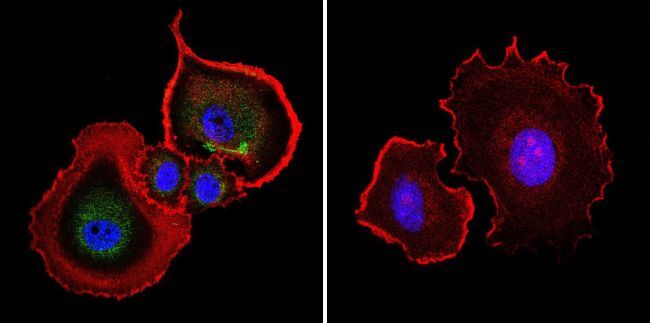
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- Immunofluorescent analysis of Aryl Hydrocarbon Receptor in NIH-3T3 cells. Cells were grown on chamber slides and fixed with formaldehyde prior to staining. Cells were probed without (control) or with a Aryl Hydrocarbon Receptor monoclonal antibody (Product # MA1-514) at a dilution of 1:200 overnight at 4 C, washed with PBS and incubated with a DyLight-488 conjugated secondary antibody (Product # 35503). Aryl Hydrocarbon Receptor staining (green), F-Actin staining with Phalloidin (red) and nuclei with DAPI (blue) is shown. Images were taken at 60X magnification.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
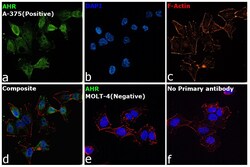
- Experimental details
- Immunofluorescence analysis of AHR was performed using 70% confluent log phase A-375 cells. The cells were fixed with 4% Paraformaldehyde for 10 minutes, permeabilized with 0.1% Triton™ X-100 for 10 minutes, and blocked with 2% BSA for 10 minutes at room temperature. The cells were labeled with AHR Monoclonal Antibody (RPT1) (Product # MA1-514) at 1:200 dilution in 0.1% BSA, incubated at 4 degree Celsius overnight and then labeled with Goat anti-Mouse IgG (H+L), Superclonal™ Recombinant Secondary Antibody, Alexa Fluor 488 (Product # A28175), (1:2000 dilution) for 45 minutes at room temperature (Panel a: Green). Nuclei (Panel b: Blue) were stained with SlowFade® Gold Antifade Mountant with DAPI (Product # S36938). F-actin (Panel c: Red) was stained with Rhodamine Phalloidin (Product # R415, 1:300). Panel d represents the merged image showing Cytoplasmic localization. Panel e represents MOLT-4 cells having no expression of AHR. Panel f represents control cells with no primary antibody to assess background. The images were captured at 60X magnification.
Supportive validation
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
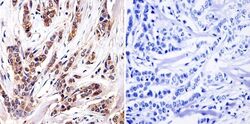
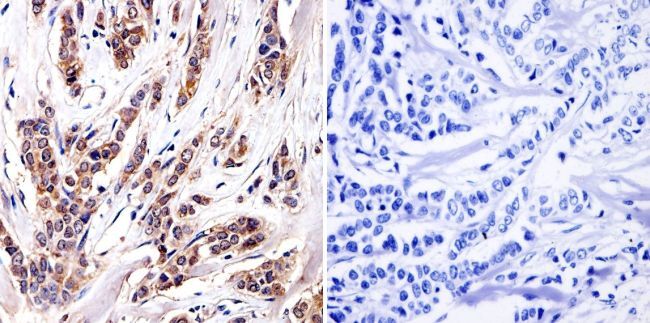
- Immunohistochemistry was performed on cancer biopsies of deparaffinized Human breast carcinoma tissue. To expose target proteins, heat induced antigen retrieval was performed using 10mM sodium citrate (pH6.0) buffer, microwaved for 8-15 minutes. Following antigen retrieval tissues were blocked in 3% BSA-PBS for 30 minutes at room temperature and probed with a Aryl Hydrocarbon Receptor monoclonal antibody (Product # MA1-514) at a dilution of 1:100 or without primary antibody (negative control) overnight at 4°C in a humidified chamber. Tissues were washed with PBST and endogenous peroxidase activity was quenched with a peroxidase suppressor. Detection was performed using a biotin-conjugated secondary antibody and SA-HRP, followed by colorimetric detection using DAB. Tissues were counterstained with hematoxylin and prepped for mounting.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
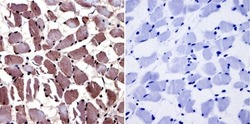
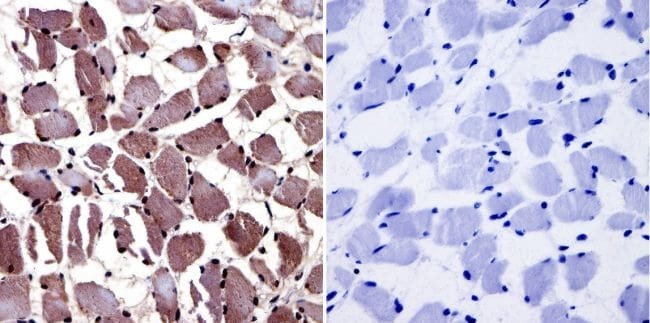
- Immunohistochemistry was performed on normal biopsies of deparaffinized Human skeletal muscle tissue. To expose target proteins, heat induced antigen retrieval was performed using 10mM sodium citrate (pH6.0) buffer, microwaved for 8-15 minutes. Following antigen retrieval tissues were blocked in 3% BSA-PBS for 30 minutes at room temperature and probed with a Aryl Hydrocarbon Receptor monoclonal antibody (Product # MA1-514) at a dilution of 1:20 or without primary antibody (negative control) overnight at 4°C in a humidified chamber. Tissues were washed with PBST and endogenous peroxidase activity was quenched with a peroxidase suppressor. Detection was performed using a biotin-conjugated secondary antibody and SA-HRP, followed by colorimetric detection using DAB. Tissues were counterstained with hematoxylin and prepped for mounting.
Supportive validation
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
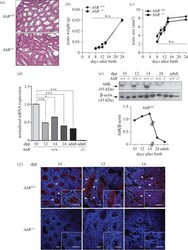
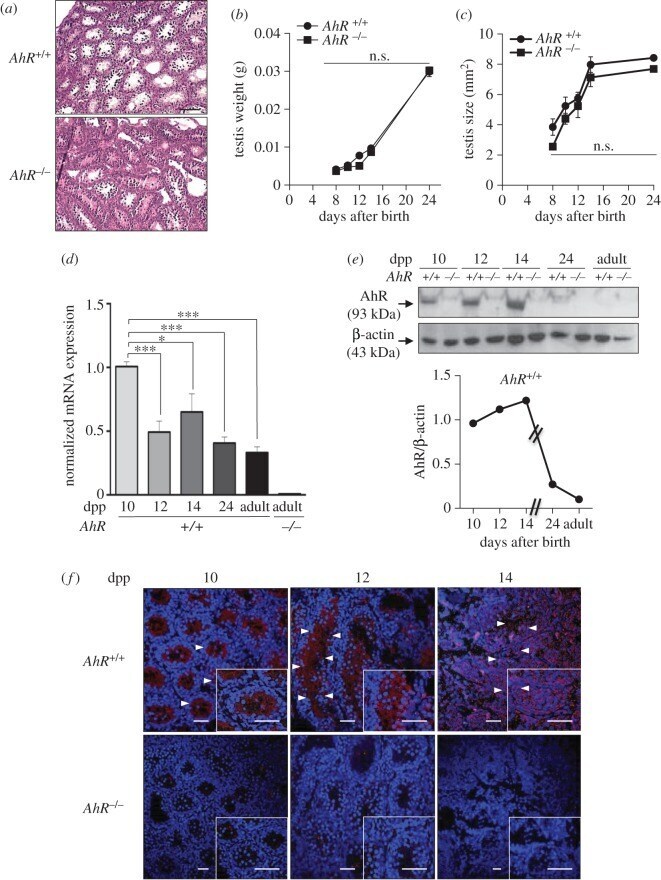
- Figure 1. AhR is expressed in mouse testis and it does not significantly affect testis weight or size. ( a ) Testes from 24 dpp AhR +/+ and AhR -/- mice were extracted, processed histologically and stained with haematoxylin-eosin. ( b , c ) Testes weight ( b ) and size ( c ) were determined in mice of each genotype at the indicated times after birth. Testis size was calculated using the formula [length x width 2 x 0.4]. ( d ) AhR mRNA expression was quantified by RT-qPCR in AhR +/+ testes at the indicated ages, using total RNA and the specific primers indicated in Material and methods. RNA isolated from adult AhR -/- testes was used as negative control. RT-qPCR data were normalized by the expression of Gapdh and represented as 2 -DeltaDeltaCt . ( e ) AhR protein expression was determined by immunoblotting in AhR +/+ testes at the indicated ages using total protein extracts. The expression of beta-actin was used to normalize protein loading. The graph represents the profile of AhR protein expression over time. ( f ) Immunofluorescence analysis of AhR expression in AhR +/+ and AhR -/- mouse testes at 10, 12 and 14 dpp. Arrowheads mark the presence of AhR (red fluorescence) in the seminiferous tubules. Testes extracted from AhR -/- mice were used as negative controls. DAPI was used to stain cell nuclei. Details of the micrographs are shown in the insets. Four biological replicates and two experimental duplicates were done for panels ( d,e ). At least four individual mice of each
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
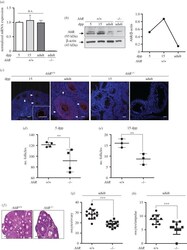
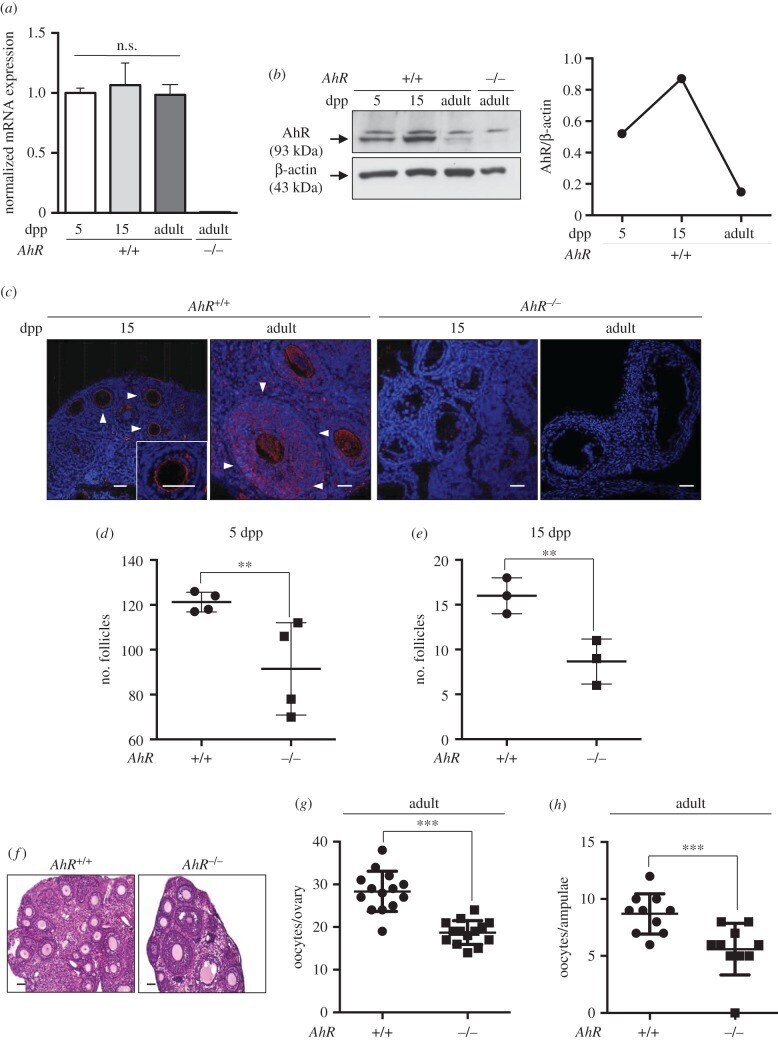
- Figure 7. AhR deficiency reduces the number of follicles in the ovary. ( a ) Ovaries were extracted from 5 and 15 dpp, and adult (six to eight weeks) AhR +/+ mice and used to obtain total RNA. AhR mRNA expression was quantified by RT-qPCR as indicated in Material and methods. Gene expression was normalized by Gapdh and represented as 2 -DeltaDeltaCt . Ovaries from adult AhR -/- mice were used as negative control. ( b ) Ovaries from AhR +/+ mice at the ages indicated above were used to prepare total protein extracts. AhR protein expression was determined by immunoblotting as indicated in Material and methods. Ovaries from adult AhR -/- mice were used as negative control. ( c ) AhR +/+ and AhR -/- ovaries were processed for immunofluorescence and stained with an AhR-specific antibody. Alexa-633-labelled secondary antibody was used to detect AhR expression. DAPI staining was added to label cell nuclei. Arrowheads mark AhR expression. ( d , e ) The number of follicles was quantified in 5 ( d ) and 15 dpp ( e ) AhR +/+ and AhR -/- ovaries. ( f ) Haematoxylin-eosin staining was performed to analyse the follicles present in adult AhR +/+ and AhR -/- ovaries. ( g,h ) Oocytes were extracted from the ovary ( g ) or the ampulae of the oviduct ( h ) of adult mice of both genotypes. At least five biological replicates were analysed for each genotype. Duplicate or triplicate experimental determinations were done. Data in panels ( a,d,e,g,h ) are shown as means +- s.d. ** p < 0.01, *** p
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
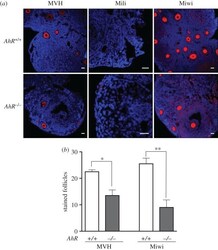
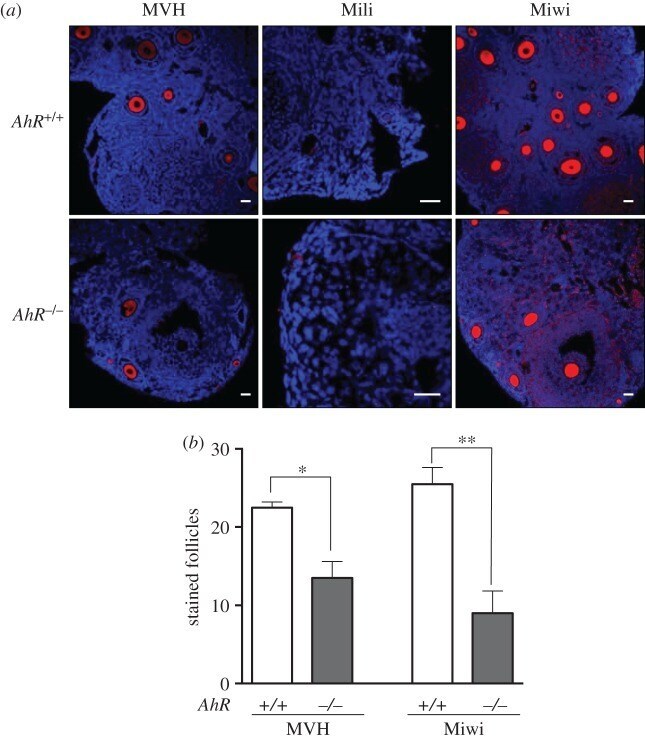
- Figure 9. Adult AhR- mice have reduced numbers of MVH- and Miwi-positive ovarian follicles. ( a ) Ovaries from adult (five to six weeks) AhR +/+ and AhR -/- mice were extracted and processed for the detection of MVH, Mili and Miwi by immunofluorescence. An Alexa-633-labelled secondary antibody has been used. ( b ) The number of positive follicles was quantified for each individual ovary of each mouse genotype. Ovaries from at least four AhR +/+ and AhR -/- mice were used, and immunofluorescences were done in triplicate. Data in panel ( b ) are shown as mean +- s.d. * p < 0.05, ** p < 0.01. Adult mice were 12-13 weeks of age. Scale bar, 50 um.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
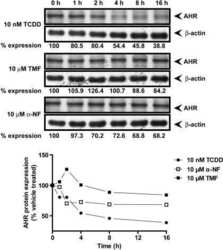
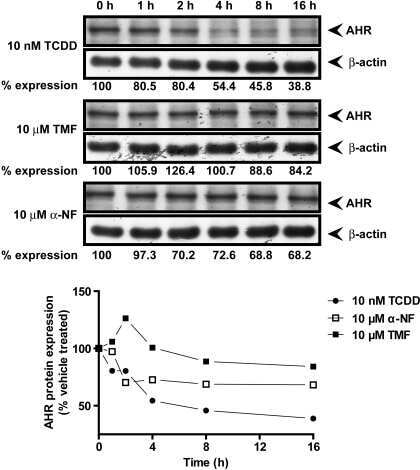
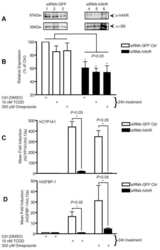
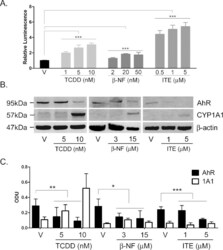
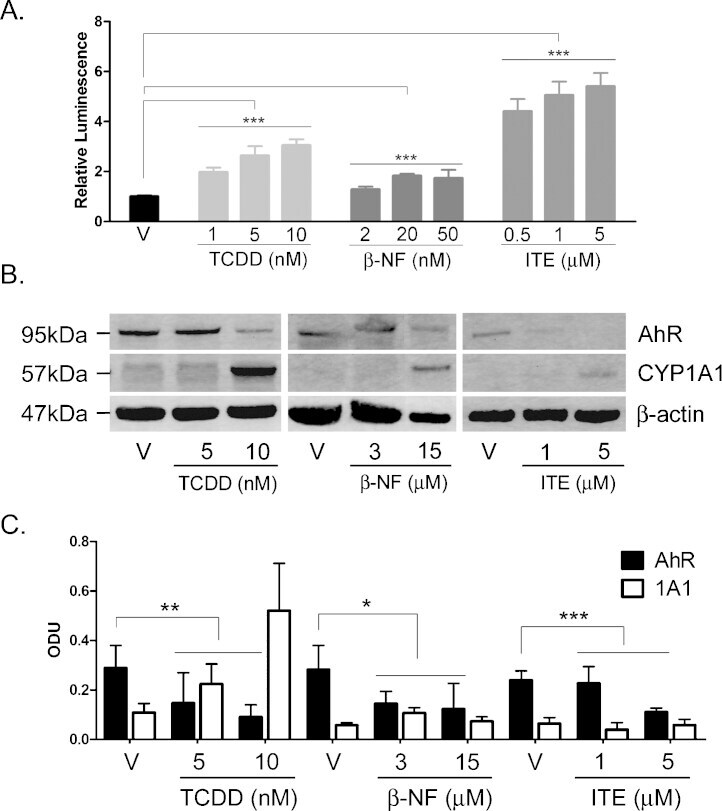
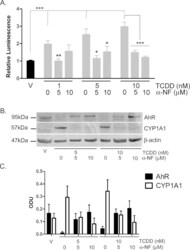
- Fig. 1 TCDD, beta-NF and ITE induce AhR activation in NHEK, (A) Primary normal human epidermal keratinocytes (NHEKs) were co-transfected with XRE4-SV40-luciferase and renilla-luciferase constructs and treated with vehicle, TCDD, beta-NF or ITE as indicated for 48 h. XRE-luciferase activity was measured and firefly:renilla luciferase ratio was normalised to vehicle. Data shown are mean +- sem, n = 9 (triplicate wells from 3 donors). Analysis of trend comparing vehicle to ligand, *** P < 0.0004. (B) Primary NHEKs were treated every 48 h for 8 days (on days 0, 2, 4 and 6) with vehicle or ligand as indicated. Samples were lysed and Western blotting performed. (C) Densitometry was carried out on blots probed with antibodies against AhR and CYP1A1 and normalised to beta-actin. Densitometry represents mean +- sem from 3 donors; graph shows results of one-way ANOVA. Effects of TCDD on AhR: one-way ANOVA, ** P = 0.007, analysis of trend P = 0.02. Effects of beta-NF on AhR: one-way ANOVA, * P = 0.02, analysis of trend, NS. Effects of ITE on AhR: one-way ANOVA, *** P = 0.0002, analysis of trend P < 0.02.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
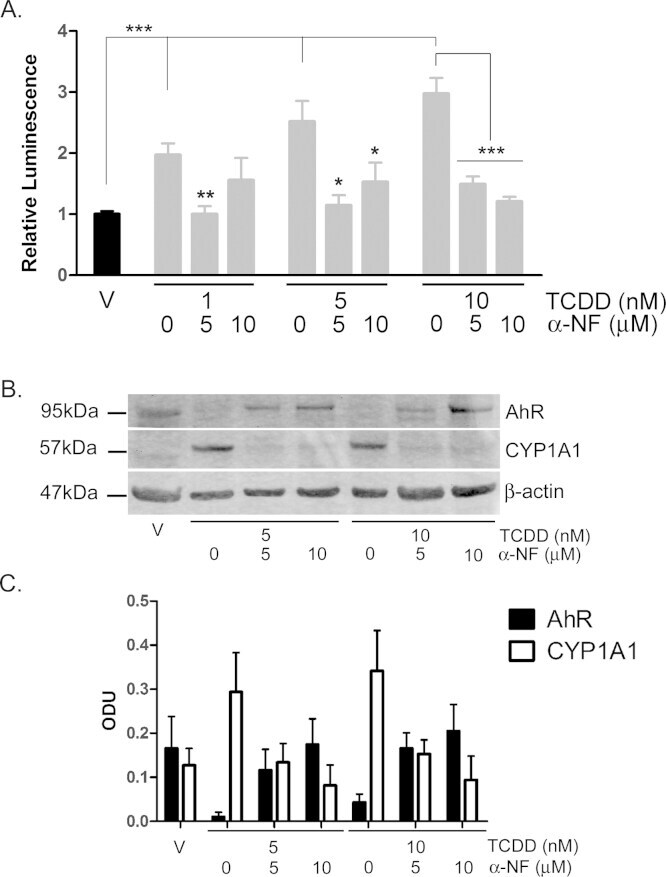
- Fig. 6 TCDD-induced AhR activation is inhibited by alpha-NF in primary NHEKs. (A) Primary NHEKs were co-transfected with XRE4-SV40-luciferase and renilla-luciferase control constructs and treated with vehicle or TCDD and/or alpha-NF as indicated for 48 h. Luciferase activity was measured and the firefly:renilla luciferase ratio was normalised to vehicle. Data shown are mean (+-sem), n = 9 (triplicate wells from 3 donors). Two-way ANOVA: post hoc tests comparing vehicle to TCDD +- alpha-NF, Dunnett's: **/*, analysis of trend: *** P < 0.0001. (B) Primary NHEKs were treated with vehicle, TCDD and/or alpha-NF every 48 h as indicated for 8 days. Cells were lysed and proteins separated by Western blotting. Blots were probed with AhR and CYP1A1 antibodies with beta-actin as loading control. Western blot is representative of duplicate blots from 3 donors. (C) Densitometry was performed on blots from 3 donors probed with antibodies against AhR (black bars) and CYP1A1 (white bars) and normalised to beta-actin. Two-way ANOVA and analysis of trend: effects of TCDD on AhR, * P = 0.02. Densitometry is represents mean (+-sem) from 3 donors.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
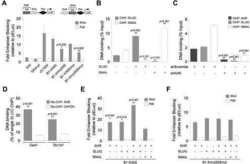
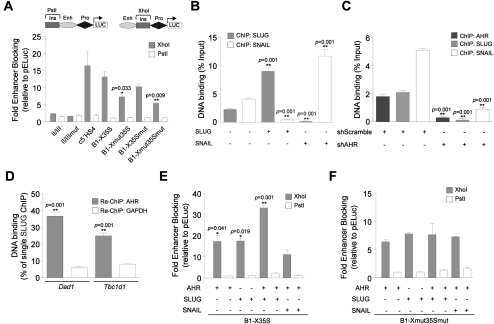
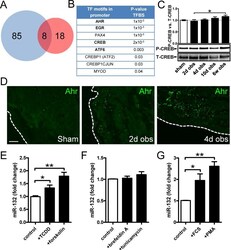
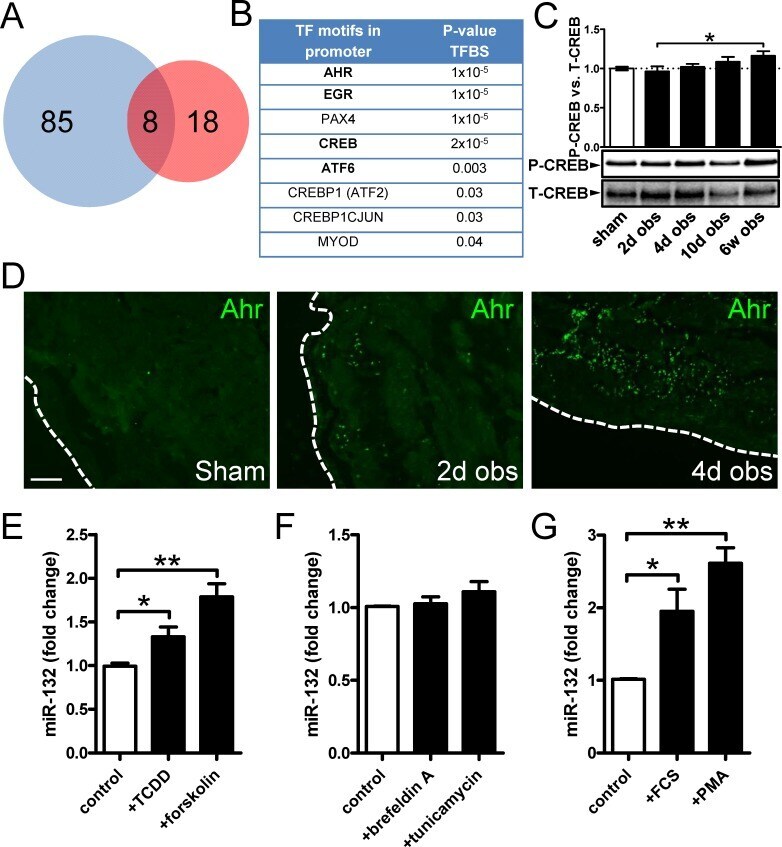
- Figure 3 Bioinformatics analysis points to the involvement of Ahr in miR-132/212 induction following outlet obstruction. Transcription factor binding site (TFBS) analysis identified significant enrichment of 93 transcription factor binding motifs in promoters of differentially expressed genes at 10 days of obstruction (blue circle in A ). These were cross-referenced against putative transcription factor binding sites in the miR-132/212 promoter (red circle in A ), yielding a list of putative mediators of miR-132/212 induction in outlet obstruction ( B ). Hits with low expression levels and those with raw p-values exceeding 0.01 are shown in thin lettering. Panel C shows phosphorylation of CREB (S133) in bladders from sham-operated rats and following various time of obstruction. Panel D shows immunofluorescence labeling (green) of the dioxin receptor Ahr in bladders from sham-operated and obstructed rats. Dotted lines indicate the outer surface of the urinary bladder. The scale bar to the left applies to all images and represents 100 mum. Panels E through G show miR-132 expression in human bladder smooth muscle cells stimulated with vehicle (control) and various pharmacological substances in vitro. TCDD: 2,3,7,8-tetrachlorodibenzo-p-dioxin; FCS: fetal calf serum; PMA: Phorbol 12-myristate 13-acetate. Further details are given in Materials and Methods .
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
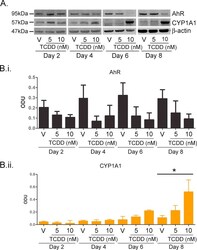
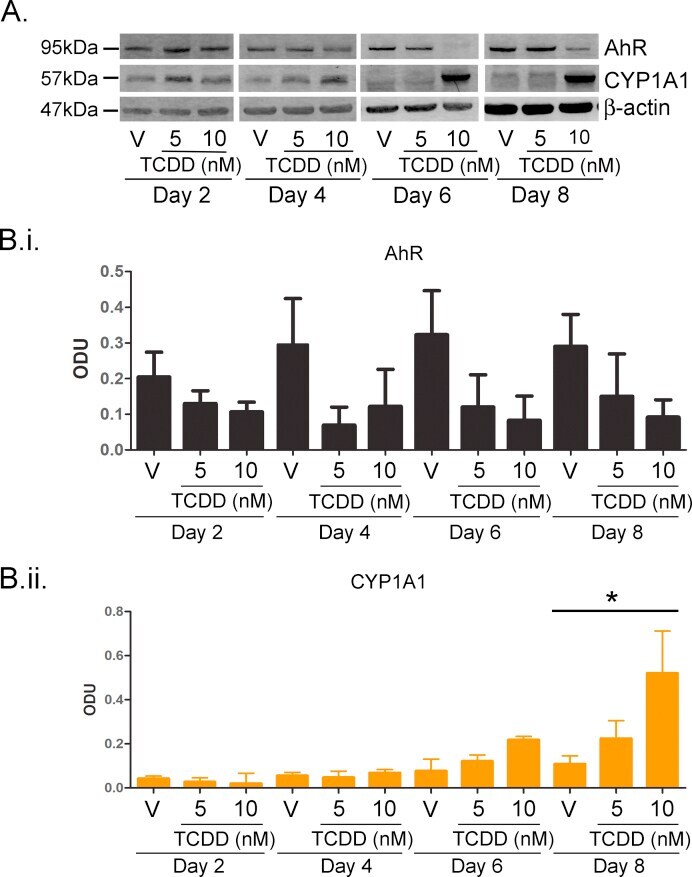
- Experimental details
- Supplementary Fig. I TCDD resulted in time and dose dependent AhR degradation and CYP1A1 induction between 2 and 8 days. (A) Primary NHEKs were treated for up to 8 days with vehicle, 5 or 10 nM TCDD as indicated. Samples were lysed and Western blotting performed. Densitometry was carried out on blots probed with antibodies against (B.i) AhR and (B.ii) CYP1A1 and normalised to beta-actin. Densitometry represents mean +- sem from 3 donors. Two-way ANOVA: analysis of trend on effects of TCDD on AhR: * P = 0.02. Effects of time and TCDD on CYP1A1 induction, analysis of trend: ** P < 0.002.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
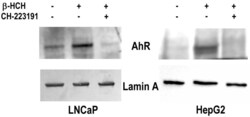
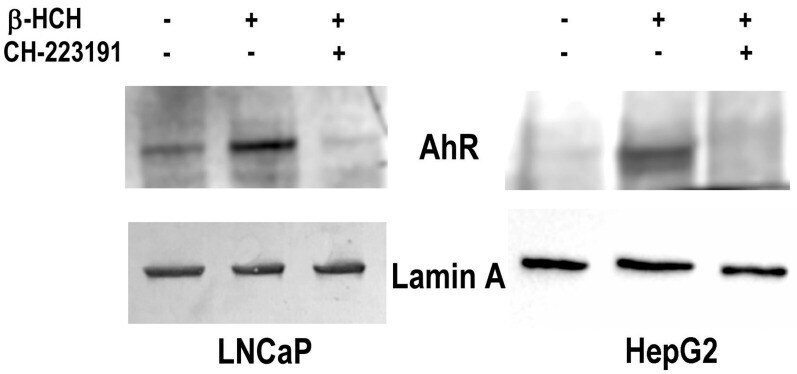
- Experimental details
- Figure 5 AhR nuclear localization in LNCaP and HepG2 cells. Western blot analysis was performed on nuclear protein extracts obtained from both prostate (LNCaP) and hepatic (HepG2) cancer cellular models treated with 10 uM beta-HCH following or not a 2 h pretreatment with 150 nM CH223191. The presence of AhR in the nuclear fraction is more noticeable in the samples corresponding to beta-HCH stimulated cells with respect to control untreated cells. The amount of nuclear AhR is similar to control if cells are treated with 10 uM beta-HCH in the presence of the AhR antagonist CH223191. Lamin A was used as a specific nuclear marker and normalization protein. The experiment was repeated three times and a representative blot is reported.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
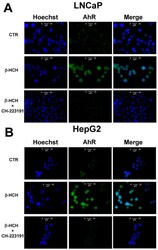
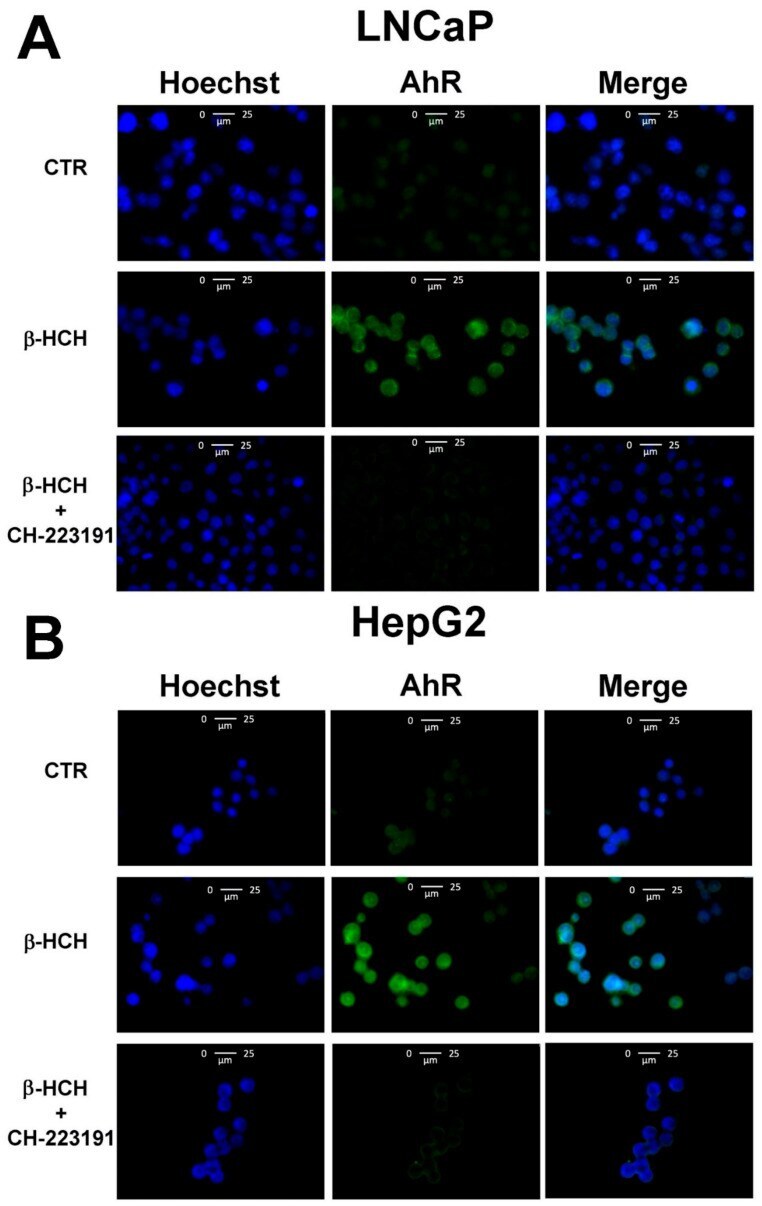
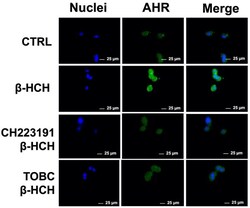
- Figure 6 Cellular distribution of AhR followed by immunofluorescence in LNCaP ( A ) and HepG2 ( B ) cells. CTR: Control untreated cells; beta-HCH: Cells after 4 h of 10 uM beta-HCH stimulation; beta-HCH + CH223191: Cells after 2 h pre-incubation with 150 nM CH223191 followed by 4 h of 10 uM beta-HCH stimulation.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
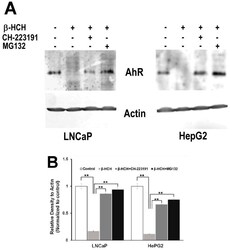
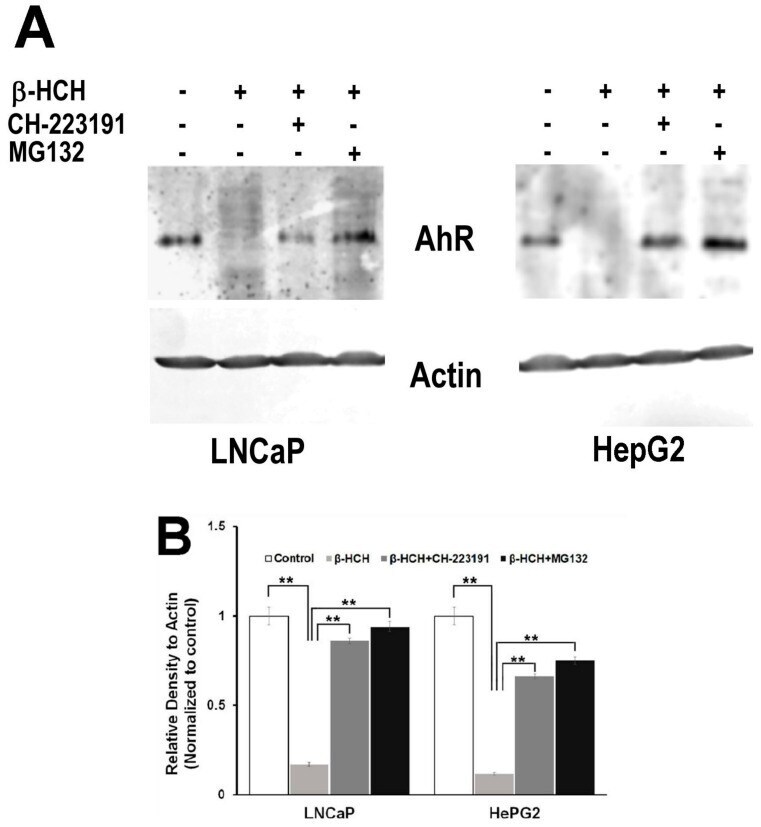
- Figure 7 AhR proteasomal degradation is induced by beta-HCH in both LNCaP and HepG2 cells. ( A ) Total protein extracts were subjected to Western blot analysis. Samples obtained from cells treated with 10 uM beta-HCH alone exhibit a weaker signal compared to the samples from cells treated with both the inhibitors. The 2 h pretreatment step with 2 uM MG132 or 150 nM CH22131 is able to maintain AhR at the same levels as the control. ( B ) Quantification of protein levels by densitometric analysis. Values for each sample were normalized against the beta-actin level present in the same sample and taken as a housekeeping control. The experiment was repeated three times and a representative blot is reported. Statistically significant differences (** p < 0.01) are marked with asterisks.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
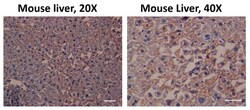
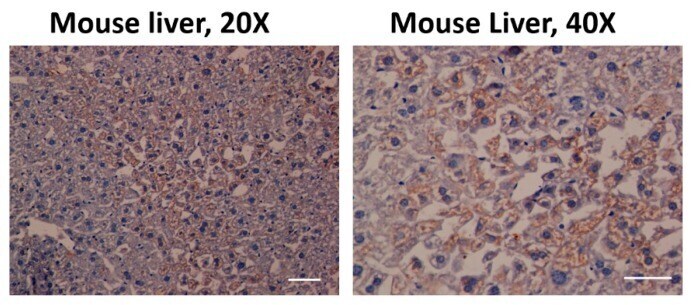
- Figure 2 Aryl hydrocarbon receptor (AHR) expression in C57BL/6 mouse liver. The antibody used against AHR was effective (coffee stain). Likewise, the samples were also stained with hematoxylin (blue) to contrast the cell nuclei Scale bar = 100 mum.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
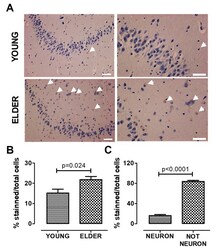
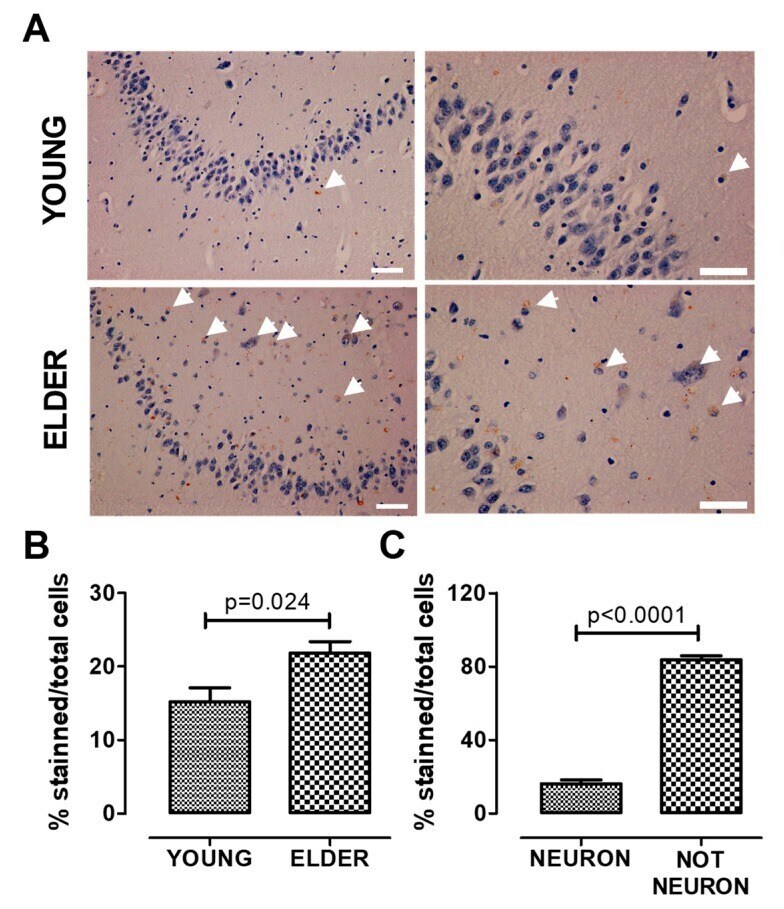
- Figure 3 AHR expression increases in the old hippocampus. We determined AHR expression by immunostaining and found that AHR expression increases in the elderly. The higher expression was found in non-neuronal cells. Representative images ( A ), total quantification ( B ), and AHR staining quantification in neurons and non-neuronal cells from the elderly ( C ). Arrows show the AHR staining. Values are mean +- SEM; Statistics were performed by unpaired t-test. Young (n = 7): 20-30 years old. Elder ( n = 6): >60 years old. Scale bar = 50 mum.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
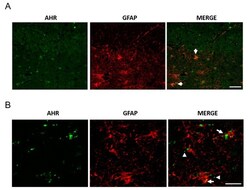
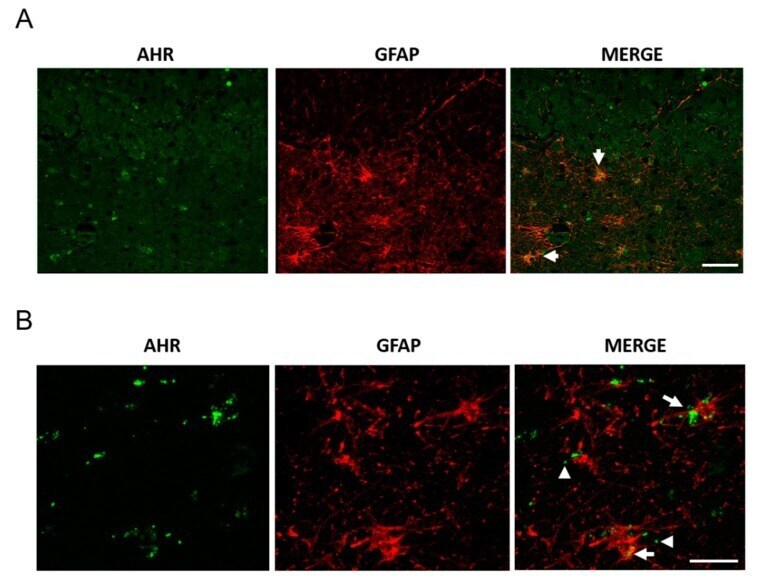
- Figure 4 AHR and GFAP colocalization in post-mortem tissue. We determined AHR and GFAP expression by immunostaining and found AHR in astrocytes, apparently as microvesicles. Representative images from a 94 year-old female donor ( A , scale bar = 50 mum) and a 100 year-old Alzheimer''s disease (AD) female donor ( B , scale bar = 20 mum). The arrows show the AHR staining in astrocytes. The arrowheads show AHR with the appearance of microvesicles. We analyzed samples from three donors.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
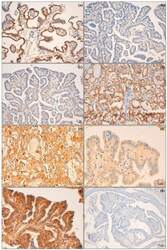
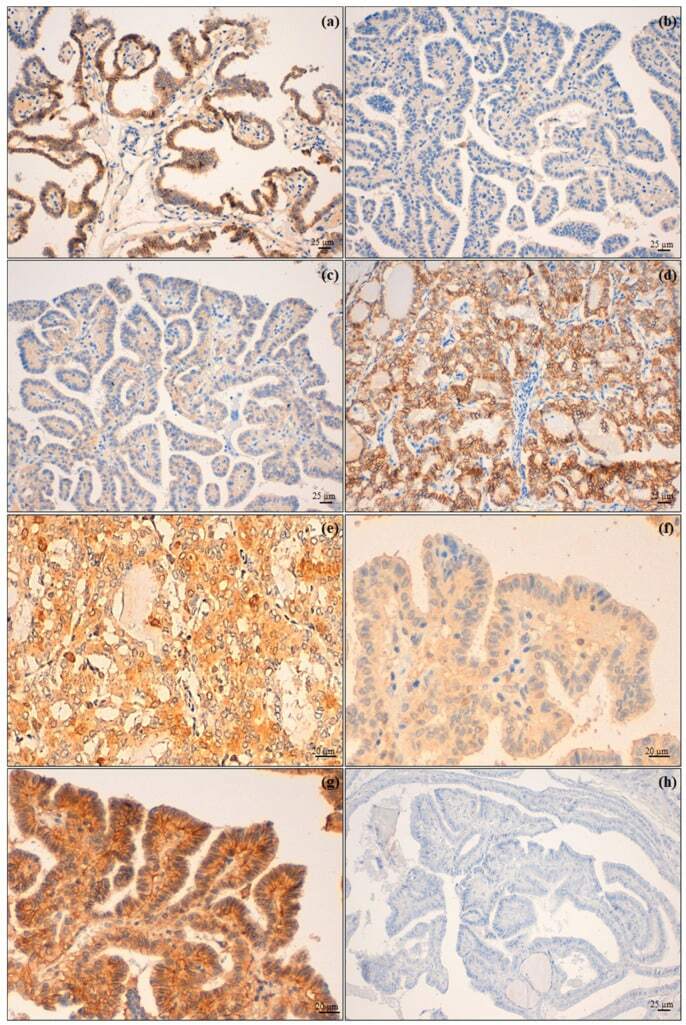
- Figure 2 Biomarker immunohistochemical expression. ( a ) Membranous high expression of N-cadherin on tumour cells of PTC infiltrating the airway (PTC-A) compared with ( b ) an intrathyroid PTC (PTC-B); ( c ) PTC-A presented a low expression of E-cadherin and ( d ) PTC-B (follicular variant) with retained expression of this molecule; ( e ) PTC-A showing a high expression of AhR in contrast with ( f ), which shows low expression of the same immunomarker in a PTC-B; ( g ) CD147 high expression of PTC-B and ( h ) absent expression in PTC-A. Original magnification: ( a - d , h ) 200x; ( e - g ) 400x.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
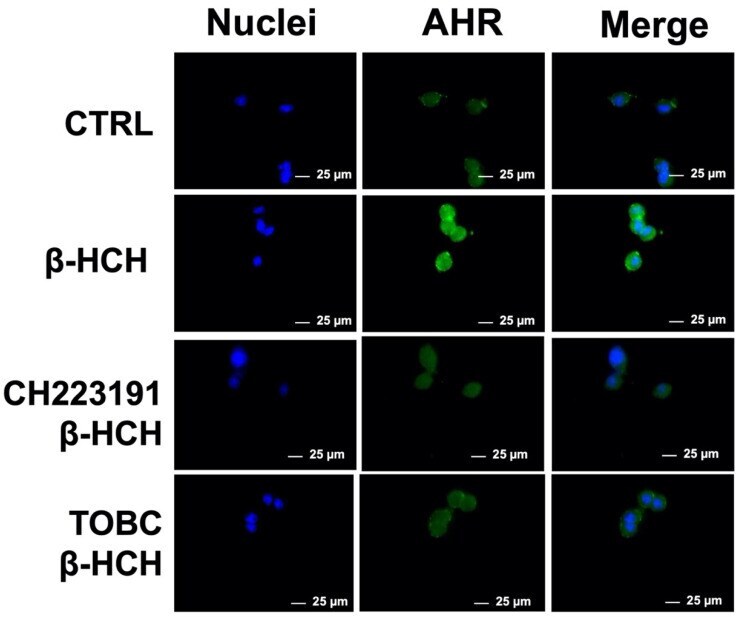
- Figure 4 Cellular distribution of AhR followed by immunofluorescence in LNCaP cells. As displayed in the figures, receptor is only present in nuclei of cells treated with beta-HCH alone, but not in presence of CH223191 nor TOBC. Selected images are representative of three independent experiments and were captured under same acquisition parameters CTR: control untreated cells; beta-HCH: cells subjected to a 4 h stimulation with 10 uM beta-HCH; beta-HCH + CH223191: cells after 2 h pre-incubation with 150 nM CH223191, followed by 4 h of 10 uM beta-HCH stimulation; TOBC+ beta-HCH: cells pretreated 3 h with 1 mg/mL TOBC, and then for 4 h with 10 uM beta-HCH. CH223191 was used as a positive control for AhR inhibition.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
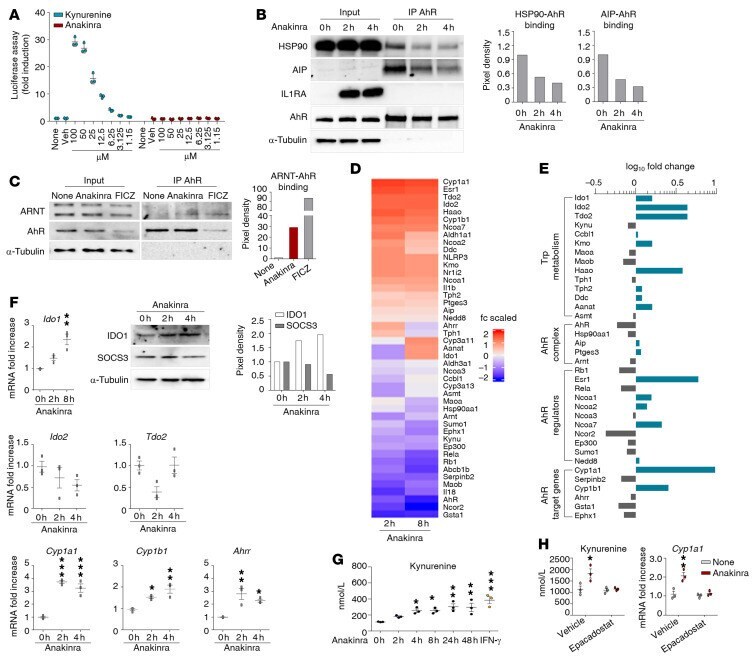
- Figure 5 Anakinra activates a xenobiotic sensing pathway via IDO1. ( A ) H1L1 cells were treated with different doses of kynurenine and anakinra for 6 hours and assessed for luciferase assay. ( B ) Representative immunoblots of IL1RA, HSP90, AIP, and AhR and ( C ) ARNT and AhR in cell lysates in which AhR was immunoprecipitated from Il1r1 -/- MEF cells treated with 10 mug/mL anakinra or 10 muM FICZ. In B and C , data are representative of 1 out of 2 independent experiments and the relative densitometric analysis are reported. ( D and E ) Il1r1 -/- MEF cells were treated with 10 mug/mL anakinra for 2 and 8 hours and analyzed for gene expression by a custom QuantiGene plex gene expression assay. Fold changes are reported as heatmap for 2 and 8 hours ( D ) and histograms for 8 hours ( E ) (data are representative of 1 out of 2 independent experiments). ( F ) Il1r1 -/- MEF cells were treated with 10 mug/mL anakinra and analyzed for mRNA expression of selected genes by RT-PCR ( n = 3 independent samples) and protein expression of IDO1 and SOCS3 by immunoblotting (representative experiment). The relative densitometric analysis is reported. ( G ) Il1r1 -/- MEF cells were treated with either 10 mug/mL anakinra for different times or 10 ng/mL IFN-gamma as positive control for 48 hours and assessed for kynurenine production by ELISA. ( H ) Il1r1 -/- MEF cells were treated with 10 mug/mL anakinra in the presence or absence of 10 muM epacadostat and assessed for kynurenine production by
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
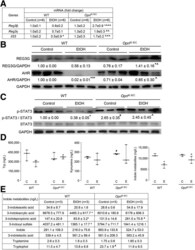
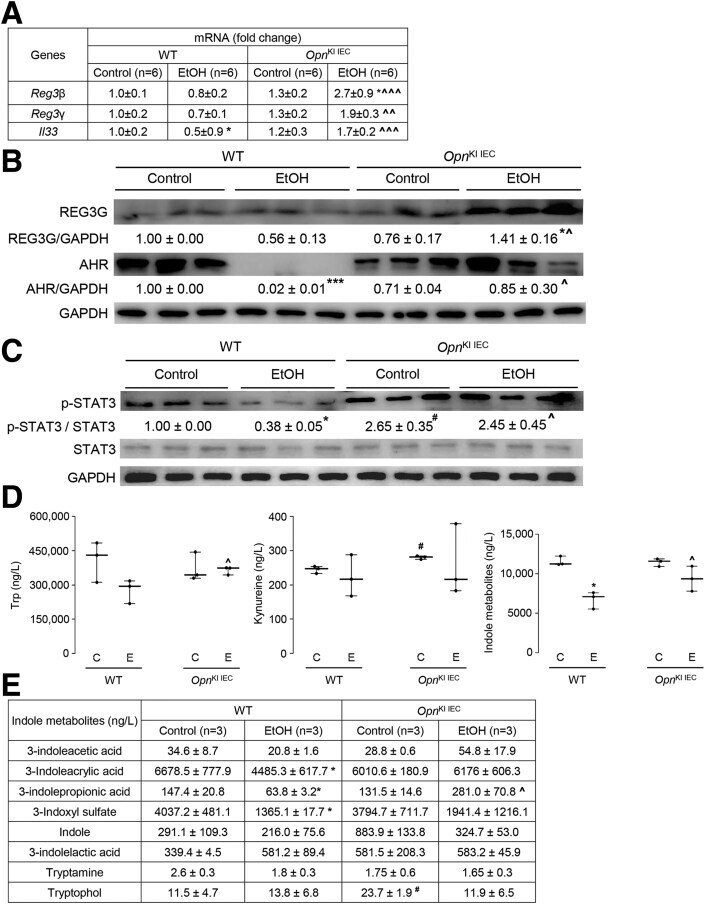
- Experimental details
- Figure 6 OPN preserves the gut microbiome by inducing AMPs expression in IECs, which maintains intestinal barrier function and protects from ALD. WT and Opn KI IEC mice were fed control or ethanol diet for 6 weeks to provoke ALD. mRNA expression of Reg3beta , Reg3gamma , and Il33 in IECs isolated from jejunum. Data were normalized with Gapdh as housekeeping gene and FC was calculated against WT control (n = 6 [3 males + 3 females]/group) ( A ). Western blot of REG3G and total AHR in IECs isolated from jejunum (n = 3 males/group) ( B ). Western blot of STAT3 and pSTAT3 in IECs isolated from jejunum (n = 3 males/group) ( C ). Levels of Trp, its metabolites kynureine and indole metabolites (3-indoleacetic acid + 3-indoleacrylic acid + 3-indolepropionic acid + 3-indoxyl sulfate + indole + indole-3-lactic acid + tryptamine + tryptophol) in portal serum (n = 3 males/group) ( D-E ). Levels of SCFAs (acetic acid + propionic acid + butyric acid) in portal serum (n = 3 males/group) ( F-G ). mRNA expression of Ahr , Occludin , Claudin3 , JamA , and Jam4 in IECs from jejunum. Data were normalized with Gapdh as housekeeping gene and FC was calculated against WT control (n = 6 [3 males +3 females]/group) ( H ). Data are expressed as mean +- standard error of the mean (SEM). * P < .05; ** P < .01; and *** P < .001 vs control; # P < .01 vs WT control; ^ P < .05; ^^ P < .01; and ^^^ P < .01 vs WT ethanol. WT and Opn DeltaIEC mice were fed control or ethanol diet for 6 weeks to provoke ALD. mR
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
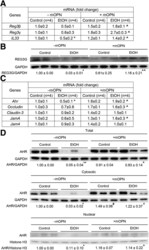
- Experimental details
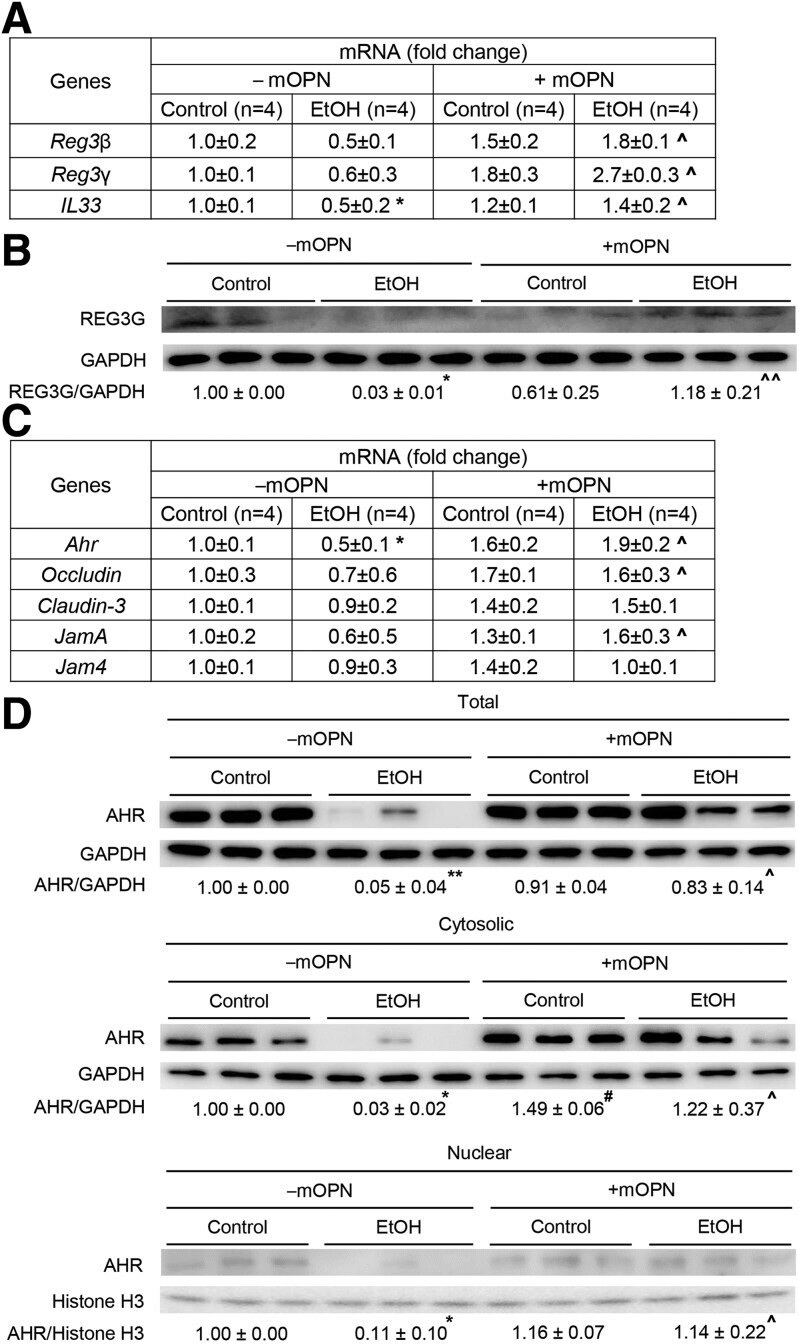
- Oral administration of mOPN induces AMPs in IECs, which preserve the gut microbiota and the intestinal barrier function. WT mice were fed control or ethanol diet for 6 weeks in the presence or absence of mOPN. mRNA expression of Reg3beta , Reg3gamma , and Il33 in IECs from jejunum. Data were normalized with Gapdh as housekeeping gene, and FC was calculated against without mOPN control (n = 4/group) ( A ). REG3G protein expression in IECs from jejunum (n = 3/group) ( B ). mRNA expression of Ahr , Occludin , Claudin3 , JamA , and Jam4 in IECs from jejunum. Data were normalized with Gapdh as housekeeping gene, and FC was calculated against without mOPN control (n = 4/group) ( C ). Western blot of total, cytosolic and nuclear AHR in IECs from jejunum (n = 3/group) ( D ). Levels of Trp, its metabolites kynureine and indole metabolites (3-indoleacrylic acid + 3-indoleacrylic acid + 3-indolepropionic acid + 3-indoxyl sulfate + indole + indole-3-lactic acid + tryptamine + tryptophol) in portal serum (n = 3/group) ( E-F ). Level of SCFAs (acetic acid + propionic acid + butyric acid) in portal serum (n = 4/group) ( G-H ). Data are expressed as mean +- standard error of the mean. * P