Explore
Explore Validate
Validate Learn
Learn Western blot
Western blot ELISA
ELISA Other assay
Other assayAntibody data
- Antibody Data
- Antigen structure
- References [74]
- Comments [0]
- Validations
- Other assay [38]
Submit
Validation data
Reference
Comment
Report error
- Product number
- 13-8100 - Provider product page

- Provider
- Invitrogen Antibodies
- Product name
- Connexin 26 Monoclonal Antibody (CX-12H10)
- Antibody type
- Monoclonal
- Antigen
- Synthetic peptide
- Description
- 13-8100 reacts strongly with Connexin-26, which has a predicted molecular weight of 26.5 kDa. Reactivity of this antibody on western blots has been confirmed by using extracts from mouse liver, mouse brain, and rat brain and it appears to exhibit minimal cross-reaction with Cx30 by western blotting. The degree of cross-reaction with Cx30 by IHC is uncertain, but may be influenced by fixation conditions.
- Reactivity
- Human, Mouse, Rat
- Host
- Mouse
- Isotype
- IgG
- Antibody clone number
- CX-12H10
- Vial size
- 100 μg
- Concentration
- 0.5 mg/mL
- Storage
- -20°C
Submitted references Purification of Fibroblasts From the Spiral Ganglion.
Dysregulation of Connexin Expression Plays a Pivotal Role in Psoriasis.
Dynamic Spatiotemporal Expression Changes in Connexins of the Developing Primate's Cochlea.
GJB2 Mutations Linked to Hearing Loss Exhibit Differential Trafficking and Functional Defects as Revealed in Cochlear-Relevant Cells.
A heterozygous mutation in GJB2 (Cx26F142L) associated with deafness and recurrent skin rashes results in connexin assembly deficiencies.
Allele-Specific Small Interfering RNA Corrects Aberrant Cellular Phenotype in Keratitis-Ichthyosis-Deafness Syndrome Keratinocytes.
Moderate Changes in CO(2) Modulate the Firing of Neurons in the VTA and Substantia Nigra.
Detecting CO(2)-Sensitive Hemichannels in Neurons in Acute Brain Slices.
Characterisation of medullary astrocytic populations in respiratory nuclei and alterations in sudden unexpected death in epilepsy.
Connexin 43 maintains tissue polarity and regulates mitotic spindle orientation in the breast epithelium.
Early Hearing Loss upon Disruption of Slc4a10 in C57BL/6 Mice.
Connexin 43 plays an important role in the transformation of cholangiocytes with Clonochis sinensis excretory-secretory protein and N-nitrosodimethylamine.
Intrinsic Oncogenic Function of Intracellular Connexin26 Protein in Head and Neck Squamous Cell Carcinoma Cells.
Immunofluorescence reveals unusual patterns of labelling for connexin43 localized to calbindin-D28K-positive interstitial cells in the pineal gland.
Disease-linked connexin26 S17F promotes volar skin abnormalities and mild wound healing defects in mice.
Essential Roles of L-Type Amino Acid Transporter 1 in Syncytiotrophoblast Development by Presenting Fusogenic 4F2hc.
Overlapping expression of anion exchangers in the cochlea of a non-human primate suggests functional compensation.
Altered CO2 sensitivity of connexin26 mutant hemichannels in vitro.
Expression of connexin 26 and connexin 43 is reduced in Hirschsprung's disease.
CO2-evoked release of PGE2 modulates sighs and inspiration as demonstrated in brainstem organotypic culture.
Distinct Expression Patterns Of Causative Genes Responsible For Hereditary Progressive Hearing Loss In Non-Human Primate Cochlea.
Tanshinone IIA enhances bystander cell killing of cancer cells expressing Drosophila melanogaster deoxyribonucleoside kinase in nuclei and mitochondria.
OTX2 regulates the expression of TAp63 leading to macular and cochlear neuroepithelium development.
Cx26 knockout predisposes the mammary gland to primary mammary tumors in a DMBA-induced mouse model of breast cancer.
Reduced expression of Connexin26 and its DNA promoter hypermethylation in the inner ear of mimetic aging rats induced by d-galactose.
The Clouston syndrome mutation connexin30 A88V leads to hyperproliferation of sebaceous glands and hearing impairments in mice.
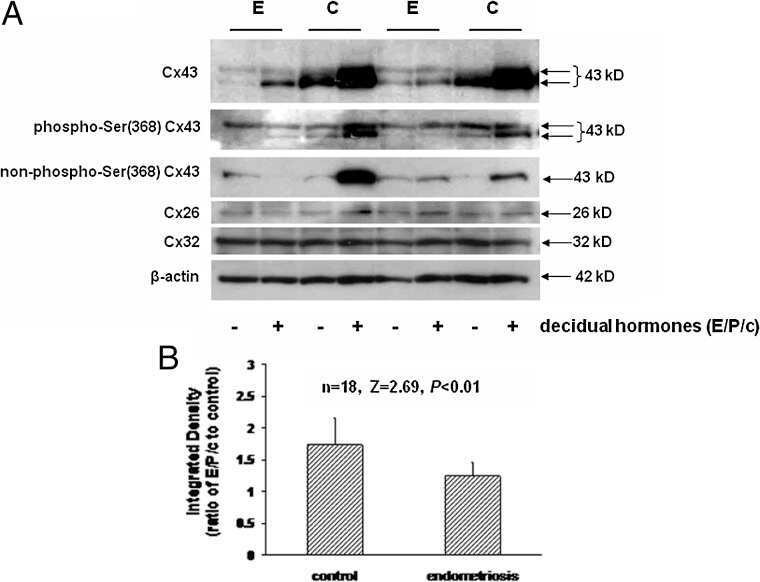
Reduced connexin 43 in eutopic endometrium and cultured endometrial stromal cells from subjects with endometriosis.
Expression of gap junction proteins connexins 26, 30, and 43 in Dupuytren's disease.
Tanshinone IIA increases the bystander effect of herpes simplex virus thymidine kinase/ganciclovir gene therapy via enhanced gap junctional intercellular communication.
Phosphorylation on Ser-279 and Ser-282 of connexin43 regulates endocytosis and gap junction assembly in pancreatic cancer cells.
Genetic ablation of the fatty acid-binding protein FABP5 suppresses HER2-induced mammary tumorigenesis.
Role of p63 and the Notch pathway in cochlea development and sensorineural deafness.
Connexin32 can restore hearing in connexin26 deficient mice.
Different domains are critical for oligomerization compatibility of different connexins.
The connexin26 S17F mouse mutant represents a model for the human hereditary keratitis-ichthyosis-deafness syndrome.
Connexin 26 expression correlates with less aggressive phenotype of intestinal type-gastric carcinomas.
Clinical significance of the expression of connexin26 in colorectal cancer.
Consortin, a trans-Golgi network cargo receptor for the plasma membrane targeting and recycling of connexins.
The human deafness-associated connexin 30 T5M mutation causes mild hearing loss and reduces biochemical coupling among cochlear non-sensory cells in knock-in mice.
Pannexin1 and Pannexin2 channels show quaternary similarities to connexons and different oligomerization numbers from each other.
Connexin30 null and conditional connexin26 null mice display distinct pattern and time course of cellular degeneration in the cochlea.
Cx36 makes channels coupling human pancreatic beta-cells, and correlates with insulin expression.
Neuronal connexin expression in the cochlear nucleus of big brown bats.
Aberrant expression of connexin 26 is associated with lung metastasis of colorectal cancer.
Connexin hemichannel composition determines the FGF-1-induced membrane permeability and free [Ca2+]i responses.
A murine living skin equivalent amenable to live-cell imaging: analysis of the roles of connexins in the epidermis.
Role of gap junctions in spontaneous activity of the rat bladder.
Impaired gap junction formation and intercellular calcium signaling in urinary bladder cancer cells can be improved by Gö6976.
In differentiating prefusion myoblasts connexin43 gap junction coupling is upregulated before myoblast alignment then reduced in post-mitotic cells.
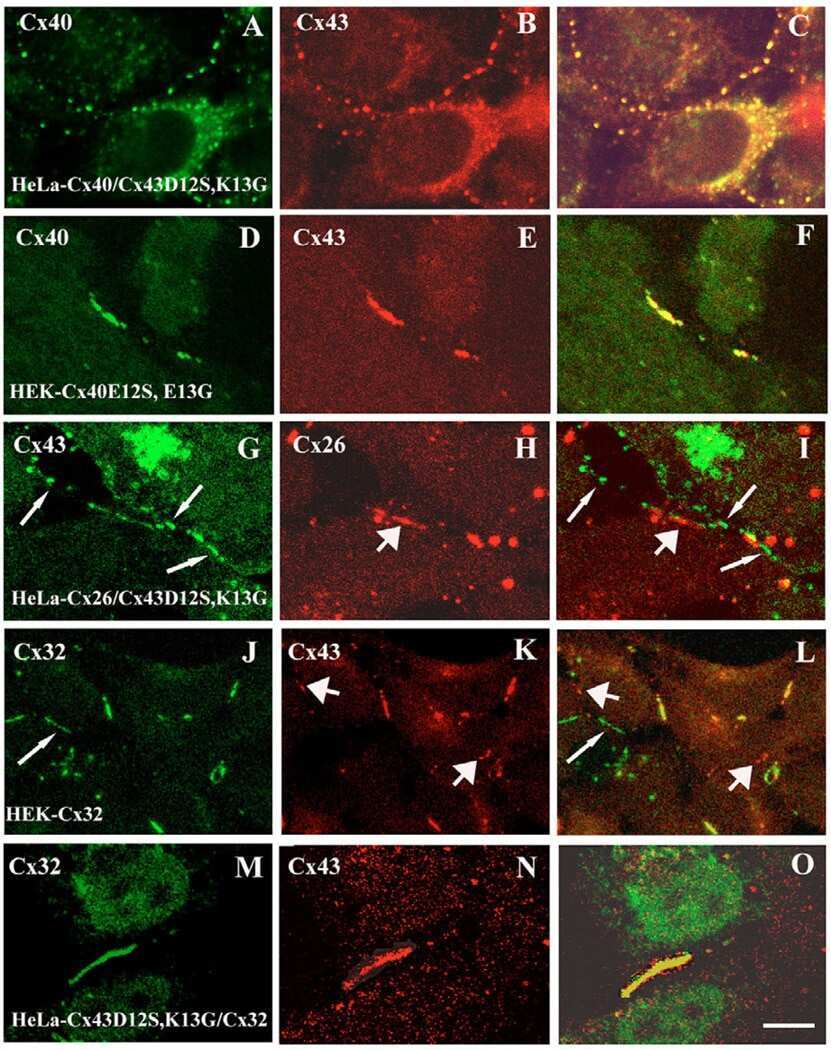
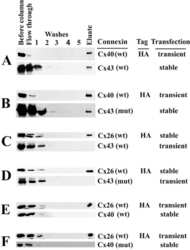
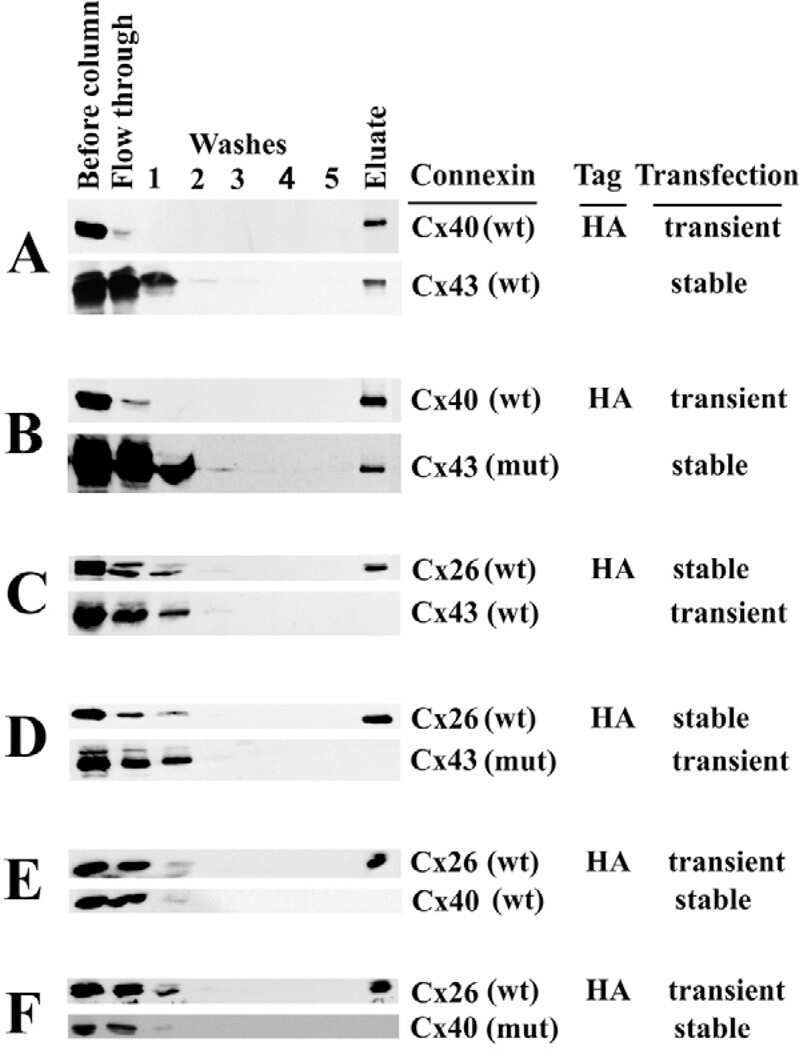
N-terminal residues in Cx43 and Cx40 determine physiological properties of gap junction channels, but do not influence heteromeric assembly with each other or with Cx26.
Contacts and cooperation between cells depend on the hormone ouabain.
Differential induction of connexins 26 and 30 in skin tumors and their adjacent epidermis.
Restoration of functional gap junctions through internal ribosome entry site-dependent synthesis of endogenous connexins in density-inhibited cancer cells.
Transient upregulation of connexin43 gap junctions and synchronized cell cycle control precede myoblast fusion in regenerating skeletal muscle in vivo.
Connexin43 with a cytoplasmic loop deletion inhibits the function of several connexins.
Connexin43 and connexin26 form gap junctions, but not heteromeric channels in co-expressing cells.
Overexpression of estrogen receptor-alpha gene suppresses gap junctional intercellular communication in endometrial carcinoma cells.
Functional domain mapping and selective trans-dominant effects exhibited by Cx26 disease-causing mutations.
Sequential changes of connexin32 and connexin26 in ischemia-reperfusion of the liver in rats.
The relationship between connexins, gap junctions, tissue architecture and tumour invasion, as studied in a novel in vitro model of HPV-16-associated cervical cancer progression.
Suppression of gap junctional intercellular communication via 5' CpG island methylation in promoter region of E-cadherin gene in endometrial cancer cells.
Connexin immunoreactivity in glial cells of the rat retina.
Role of gap junctions in fluid secretion of lacrimal glands.
Cell-specific expression of connexins and evidence of restricted gap junctional coupling between glial cells and between neurons.
Cell-specific expression of connexins and evidence of restricted gap junctional coupling between glial cells and between neurons.
trans-dominant inhibition of connexin-43 by mutant connexin-26: implications for dominant connexin disorders affecting epidermal differentiation.
Connexin 43 hemi channels mediate Ca2+-regulated transmembrane NAD+ fluxes in intact cells.
Connexin 26 and basic fibroblast growth factor are expressed primarily in the subpial and subependymal layers in adult brain parenchyma: roles in stem cell proliferation and morphological plasticity?
Localization of connexin26 and connexin32 in putative CO(2)-chemosensitive brainstem regions in rat.
Localization of connexin26 and connexin32 in putative CO(2)-chemosensitive brainstem regions in rat.
A role for heterologous gap junctions between melanoma and endothelial cells in metastasis.
Transport function and hepatocellular localization of mrp6 in rat liver.
Biochemical and functional characterization of intercellular adhesion and gap junctions in fibroblasts.
Biochemical and functional characterization of intercellular adhesion and gap junctions in fibroblasts.
Anacker A, Esser KH, Lenarz T, Paasche G
Frontiers in neurology 2022;13:877342
Frontiers in neurology 2022;13:877342
Dysregulation of Connexin Expression Plays a Pivotal Role in Psoriasis.
O'Shaughnessy EM, Duffy W, Garcia-Vega L, Hussey K, Burden AD, Zamiri M, Martin PE
International journal of molecular sciences 2021 Jun 4;22(11)
International journal of molecular sciences 2021 Jun 4;22(11)
Dynamic Spatiotemporal Expression Changes in Connexins of the Developing Primate's Cochlea.
Hosoya M, Fujioka M, Murayama AY, Ogawa K, Okano H, Ozawa H
Genes 2021 Jul 16;12(7)
Genes 2021 Jul 16;12(7)
GJB2 Mutations Linked to Hearing Loss Exhibit Differential Trafficking and Functional Defects as Revealed in Cochlear-Relevant Cells.
Beach R, Abitbol JM, Allman BL, Esseltine JL, Shao Q, Laird DW
Frontiers in cell and developmental biology 2020;8:215
Frontiers in cell and developmental biology 2020;8:215
A heterozygous mutation in GJB2 (Cx26F142L) associated with deafness and recurrent skin rashes results in connexin assembly deficiencies.
Albuloushi A, Lovgren ML, Steel A, Yeoh Y, Waters A, Zamiri M, Martin PE
Experimental dermatology 2020 Oct;29(10):970-979
Experimental dermatology 2020 Oct;29(10):970-979
Allele-Specific Small Interfering RNA Corrects Aberrant Cellular Phenotype in Keratitis-Ichthyosis-Deafness Syndrome Keratinocytes.
Lee MY, Wang HZ, White TW, Brooks T, Pittman A, Halai H, Petrova A, Xu D, Hart SL, Kinsler VA, Di WL
The Journal of investigative dermatology 2020 May;140(5):1035-1044.e7
The Journal of investigative dermatology 2020 May;140(5):1035-1044.e7
Moderate Changes in CO(2) Modulate the Firing of Neurons in the VTA and Substantia Nigra.
Hill E, Dale N, Wall MJ
iScience 2020 Jul 24;23(7):101343
iScience 2020 Jul 24;23(7):101343
Detecting CO(2)-Sensitive Hemichannels in Neurons in Acute Brain Slices.
Hill E, Dale N, Wall MJ
STAR protocols 2020 Dec 18;1(3):100139
STAR protocols 2020 Dec 18;1(3):100139
Characterisation of medullary astrocytic populations in respiratory nuclei and alterations in sudden unexpected death in epilepsy.
Patodia S, Paradiso B, Ellis M, Somani A, Sisodiya SM, Devinsky O, Thom M
Epilepsy research 2019 Nov;157:106213
Epilepsy research 2019 Nov;157:106213
Connexin 43 maintains tissue polarity and regulates mitotic spindle orientation in the breast epithelium.
Bazzoun D, Adissu HA, Wang L, Urazaev A, Tenvooren I, Fostok SF, Chittiboyina S, Sturgis J, Hodges K, Chandramouly G, Vidi PA, Talhouk RS, Lelièvre SA
Journal of cell science 2019 May 16;132(10)
Journal of cell science 2019 May 16;132(10)
Early Hearing Loss upon Disruption of Slc4a10 in C57BL/6 Mice.
Huebner AK, Maier H, Maul A, Nietzsche S, Herrmann T, Praetorius J, Hübner CA
Journal of the Association for Research in Otolaryngology : JARO 2019 Jun;20(3):233-245
Journal of the Association for Research in Otolaryngology : JARO 2019 Jun;20(3):233-245
Connexin 43 plays an important role in the transformation of cholangiocytes with Clonochis sinensis excretory-secretory protein and N-nitrosodimethylamine.
Kim EM, Bae YM, Choi MH, Hong ST
PLoS neglected tropical diseases 2019 Apr;13(4):e0006843
PLoS neglected tropical diseases 2019 Apr;13(4):e0006843
Intrinsic Oncogenic Function of Intracellular Connexin26 Protein in Head and Neck Squamous Cell Carcinoma Cells.
Iikawa N, Yamamoto Y, Kawasaki Y, Nishijima-Matsunobu A, Suzuki M, Yamada T, Omori Y
International journal of molecular sciences 2018 Jul 23;19(7)
International journal of molecular sciences 2018 Jul 23;19(7)
Immunofluorescence reveals unusual patterns of labelling for connexin43 localized to calbindin-D28K-positive interstitial cells in the pineal gland.
Tsao DD, Wang SG, Lynn BD, Nagy JI
The European journal of neuroscience 2017 Jun;45(12):1553-1569
The European journal of neuroscience 2017 Jun;45(12):1553-1569
Disease-linked connexin26 S17F promotes volar skin abnormalities and mild wound healing defects in mice.
Press E, Alaga KC, Barr K, Shao Q, Bosen F, Willecke K, Laird DW
Cell death & disease 2017 Jun 1;8(6):e2845
Cell death & disease 2017 Jun 1;8(6):e2845
Essential Roles of L-Type Amino Acid Transporter 1 in Syncytiotrophoblast Development by Presenting Fusogenic 4F2hc.
Ohgaki R, Ohmori T, Hara S, Nakagomi S, Kanai-Azuma M, Kaneda-Nakashima K, Okuda S, Nagamori S, Kanai Y
Molecular and cellular biology 2017 Jun 1;37(11)
Molecular and cellular biology 2017 Jun 1;37(11)
Overlapping expression of anion exchangers in the cochlea of a non-human primate suggests functional compensation.
Hosoya M, Fujioka M, Kobayashi R, Okano H, Ogawa K
Neuroscience research 2016 Sep;110:1-10
Neuroscience research 2016 Sep;110:1-10
Altered CO2 sensitivity of connexin26 mutant hemichannels in vitro.
de Wolf E, van de Wiel J, Cook J, Dale N
Physiological reports 2016 Nov;4(22)
Physiological reports 2016 Nov;4(22)
Expression of connexin 26 and connexin 43 is reduced in Hirschsprung's disease.
Coyle D, Doyle B, Murphy JM, O'Donnell AM, Gillick J, Puri P
The Journal of surgical research 2016 Nov;206(1):242-251
The Journal of surgical research 2016 Nov;206(1):242-251
CO2-evoked release of PGE2 modulates sighs and inspiration as demonstrated in brainstem organotypic culture.
Forsberg D, Horn Z, Tserga E, Smedler E, Silberberg G, Shvarev Y, Kaila K, Uhlén P, Herlenius E
eLife 2016 Jul 5;5
eLife 2016 Jul 5;5
Distinct Expression Patterns Of Causative Genes Responsible For Hereditary Progressive Hearing Loss In Non-Human Primate Cochlea.
Hosoya M, Fujioka M, Ogawa K, Okano H
Scientific reports 2016 Feb 26;6:22250
Scientific reports 2016 Feb 26;6:22250
Tanshinone IIA enhances bystander cell killing of cancer cells expressing Drosophila melanogaster deoxyribonucleoside kinase in nuclei and mitochondria.
Jiang H, Zhao L, Dong X, He A, Zheng C, Johansson M, Karlsson A, Zheng X
Oncology reports 2015 Sep;34(3):1487-93
Oncology reports 2015 Sep;34(3):1487-93
OTX2 regulates the expression of TAp63 leading to macular and cochlear neuroepithelium development.
Palombo R, Porta G, Bruno E, Provero P, Serra V, Neduri K, Viziano A, Alessandrini M, Micarelli A, Ottaviani F, Melino G, Terrinoni A
Aging 2015 Nov;7(11):928-36
Aging 2015 Nov;7(11):928-36
Cx26 knockout predisposes the mammary gland to primary mammary tumors in a DMBA-induced mouse model of breast cancer.
Stewart MK, Bechberger JF, Welch I, Naus CC, Laird DW
Oncotarget 2015 Nov 10;6(35):37185-99
Oncotarget 2015 Nov 10;6(35):37185-99
Reduced expression of Connexin26 and its DNA promoter hypermethylation in the inner ear of mimetic aging rats induced by d-galactose.
Wu X, Wang Y, Sun Y, Chen S, Zhang S, Shen L, Huang X, Lin X, Kong W
Biochemical and biophysical research communications 2014 Sep 26;452(3):340-6
Biochemical and biophysical research communications 2014 Sep 26;452(3):340-6
The Clouston syndrome mutation connexin30 A88V leads to hyperproliferation of sebaceous glands and hearing impairments in mice.
Bosen F, Schütz M, Beinhauer A, Strenzke N, Franz T, Willecke K
FEBS letters 2014 May 2;588(9):1795-801
FEBS letters 2014 May 2;588(9):1795-801
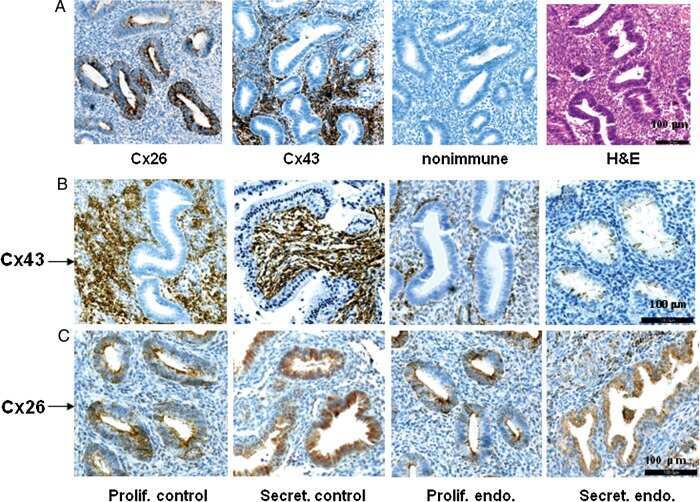
Reduced connexin 43 in eutopic endometrium and cultured endometrial stromal cells from subjects with endometriosis.
Yu J, Boicea A, Barrett KL, James CO, Bagchi IC, Bagchi MK, Nezhat C, Sidell N, Taylor RN
Molecular human reproduction 2014 Mar;20(3):260-70
Molecular human reproduction 2014 Mar;20(3):260-70
Expression of gap junction proteins connexins 26, 30, and 43 in Dupuytren's disease.
Holzer LA, Cör A, Holzer G
Acta orthopaedica 2014 Feb;85(1):97-101
Acta orthopaedica 2014 Feb;85(1):97-101
Tanshinone IIA increases the bystander effect of herpes simplex virus thymidine kinase/ganciclovir gene therapy via enhanced gap junctional intercellular communication.
Xiao J, Zhang G, Qiu P, Liu X, Wu Y, Du B, Li J, Zhou J, Li J, Tan Y
PloS one 2013;8(7):e67662
PloS one 2013;8(7):e67662
Phosphorylation on Ser-279 and Ser-282 of connexin43 regulates endocytosis and gap junction assembly in pancreatic cancer cells.
Johnson KE, Mitra S, Katoch P, Kelsey LS, Johnson KR, Mehta PP
Molecular biology of the cell 2013 Mar;24(6):715-33
Molecular biology of the cell 2013 Mar;24(6):715-33
Genetic ablation of the fatty acid-binding protein FABP5 suppresses HER2-induced mammary tumorigenesis.
Levi L, Lobo G, Doud MK, von Lintig J, Seachrist D, Tochtrop GP, Noy N
Cancer research 2013 Aug 1;73(15):4770-80
Cancer research 2013 Aug 1;73(15):4770-80
Role of p63 and the Notch pathway in cochlea development and sensorineural deafness.
Terrinoni A, Serra V, Bruno E, Strasser A, Valente E, Flores ER, van Bokhoven H, Lu X, Knight RA, Melino G
Proceedings of the National Academy of Sciences of the United States of America 2013 Apr 30;110(18):7300-5
Proceedings of the National Academy of Sciences of the United States of America 2013 Apr 30;110(18):7300-5
Connexin32 can restore hearing in connexin26 deficient mice.
Degen J, Schütz M, Dicke N, Strenzke N, Jokwitz M, Moser T, Willecke K
European journal of cell biology 2011 Oct;90(10):817-24
European journal of cell biology 2011 Oct;90(10):817-24
Different domains are critical for oligomerization compatibility of different connexins.
Martínez AD, Maripillán J, Acuña R, Minogue PJ, Berthoud VM, Beyer EC
The Biochemical journal 2011 May 15;436(1):35-43
The Biochemical journal 2011 May 15;436(1):35-43
The connexin26 S17F mouse mutant represents a model for the human hereditary keratitis-ichthyosis-deafness syndrome.
Schütz M, Auth T, Gehrt A, Bosen F, Körber I, Strenzke N, Moser T, Willecke K
Human molecular genetics 2011 Jan 1;20(1):28-39
Human molecular genetics 2011 Jan 1;20(1):28-39
Connexin 26 expression correlates with less aggressive phenotype of intestinal type-gastric carcinomas.
Liu X, Furuya T, Li D, Xu J, Cao X, Li Q, Xu J, Xu Z, Sasaki K, Liu X
International journal of molecular medicine 2010 May;25(5):709-16
International journal of molecular medicine 2010 May;25(5):709-16
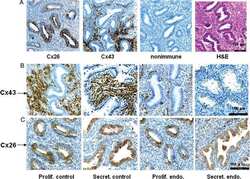
Clinical significance of the expression of connexin26 in colorectal cancer.
Nomura S, Maeda K, Noda E, Inoue T, Fukunaga S, Nagahara H, Hirakawa K
Journal of experimental & clinical cancer research : CR 2010 Jun 21;29(1):79
Journal of experimental & clinical cancer research : CR 2010 Jun 21;29(1):79
Consortin, a trans-Golgi network cargo receptor for the plasma membrane targeting and recycling of connexins.
del Castillo FJ, Cohen-Salmon M, Charollais A, Caille D, Lampe PD, Chavrier P, Meda P, Petit C
Human molecular genetics 2010 Jan 15;19(2):262-75
Human molecular genetics 2010 Jan 15;19(2):262-75
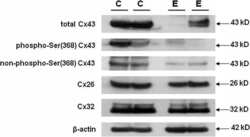
The human deafness-associated connexin 30 T5M mutation causes mild hearing loss and reduces biochemical coupling among cochlear non-sensory cells in knock-in mice.
Schütz M, Scimemi P, Majumder P, De Siati RD, Crispino G, Rodriguez L, Bortolozzi M, Santarelli R, Seydel A, Sonntag S, Ingham N, Steel KP, Willecke K, Mammano F
Human molecular genetics 2010 Dec 15;19(24):4759-73
Human molecular genetics 2010 Dec 15;19(24):4759-73
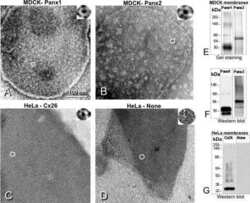
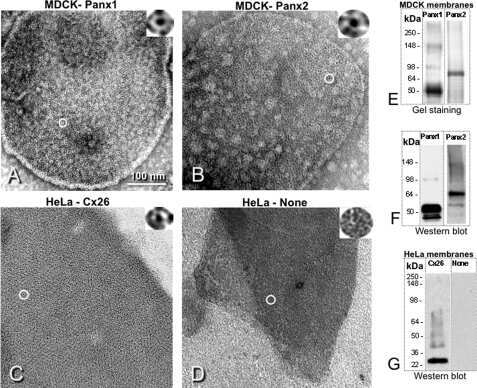
Pannexin1 and Pannexin2 channels show quaternary similarities to connexons and different oligomerization numbers from each other.
Ambrosi C, Gassmann O, Pranskevich JN, Boassa D, Smock A, Wang J, Dahl G, Steinem C, Sosinsky GE
The Journal of biological chemistry 2010 Aug 6;285(32):24420-31
The Journal of biological chemistry 2010 Aug 6;285(32):24420-31
Connexin30 null and conditional connexin26 null mice display distinct pattern and time course of cellular degeneration in the cochlea.
Sun Y, Tang W, Chang Q, Wang Y, Kong W, Lin X
The Journal of comparative neurology 2009 Oct 20;516(6):569-79
The Journal of comparative neurology 2009 Oct 20;516(6):569-79
Cx36 makes channels coupling human pancreatic beta-cells, and correlates with insulin expression.
Serre-Beinier V, Bosco D, Zulianello L, Charollais A, Caille D, Charpantier E, Gauthier BR, Diaferia GR, Giepmans BN, Lupi R, Marchetti P, Deng S, Buhler L, Berney T, Cirulli V, Meda P
Human molecular genetics 2009 Feb 1;18(3):428-39
Human molecular genetics 2009 Feb 1;18(3):428-39
Neuronal connexin expression in the cochlear nucleus of big brown bats.
Horowitz SS, Stamper SA, Simmons JA
Brain research 2008 Mar 4;1197:76-84
Brain research 2008 Mar 4;1197:76-84
Aberrant expression of connexin 26 is associated with lung metastasis of colorectal cancer.
Ezumi K, Yamamoto H, Murata K, Higashiyama M, Damdinsuren B, Nakamura Y, Kyo N, Okami J, Ngan CY, Takemasa I, Ikeda M, Sekimoto M, Matsuura N, Nojima H, Monden M
Clinical cancer research : an official journal of the American Association for Cancer Research 2008 Feb 1;14(3):677-84
Clinical cancer research : an official journal of the American Association for Cancer Research 2008 Feb 1;14(3):677-84
Connexin hemichannel composition determines the FGF-1-induced membrane permeability and free [Ca2+]i responses.
Schalper KA, Palacios-Prado N, Retamal MA, Shoji KF, Martínez AD, Sáez JC
Molecular biology of the cell 2008 Aug;19(8):3501-13
Molecular biology of the cell 2008 Aug;19(8):3501-13
A murine living skin equivalent amenable to live-cell imaging: analysis of the roles of connexins in the epidermis.
Kandyba EE, Hodgins MB, Martin PE
The Journal of investigative dermatology 2008 Apr;128(4):1039-49
The Journal of investigative dermatology 2008 Apr;128(4):1039-49
Role of gap junctions in spontaneous activity of the rat bladder.
Ikeda Y, Fry C, Hayashi F, Stolz D, Griffiths D, Kanai A
American journal of physiology. Renal physiology 2007 Oct;293(4):F1018-25
American journal of physiology. Renal physiology 2007 Oct;293(4):F1018-25
Impaired gap junction formation and intercellular calcium signaling in urinary bladder cancer cells can be improved by Gö6976.
Leinonen P, Aaltonen V, Koskela S, Lehenkari P, Korkiamäki T, Peltonen J
Cell communication & adhesion 2007 Oct;14(4):125-36
Cell communication & adhesion 2007 Oct;14(4):125-36
In differentiating prefusion myoblasts connexin43 gap junction coupling is upregulated before myoblast alignment then reduced in post-mitotic cells.
Gorbe A, Becker DL, Dux L, Krenacs L, Krenacs T
Histochemistry and cell biology 2006 Jun;125(6):705-16
Histochemistry and cell biology 2006 Jun;125(6):705-16
N-terminal residues in Cx43 and Cx40 determine physiological properties of gap junction channels, but do not influence heteromeric assembly with each other or with Cx26.
Gemel J, Lin X, Veenstra RD, Beyer EC
Journal of cell science 2006 Jun 1;119(Pt 11):2258-68
Journal of cell science 2006 Jun 1;119(Pt 11):2258-68
Contacts and cooperation between cells depend on the hormone ouabain.
Larre I, Ponce A, Fiorentino R, Shoshani L, Contreras RG, Cereijido M
Proceedings of the National Academy of Sciences of the United States of America 2006 Jul 18;103(29):10911-6
Proceedings of the National Academy of Sciences of the United States of America 2006 Jul 18;103(29):10911-6
Differential induction of connexins 26 and 30 in skin tumors and their adjacent epidermis.
Haass NK, Wladykowski E, Kief S, Moll I, Brandner JM
The journal of histochemistry and cytochemistry : official journal of the Histochemistry Society 2006 Feb;54(2):171-82
The journal of histochemistry and cytochemistry : official journal of the Histochemistry Society 2006 Feb;54(2):171-82
Restoration of functional gap junctions through internal ribosome entry site-dependent synthesis of endogenous connexins in density-inhibited cancer cells.
Lahlou H, Fanjul M, Pradayrol L, Susini C, Pyronnet S
Molecular and cellular biology 2005 May;25(10):4034-45
Molecular and cellular biology 2005 May;25(10):4034-45
Transient upregulation of connexin43 gap junctions and synchronized cell cycle control precede myoblast fusion in regenerating skeletal muscle in vivo.
Gorbe A, Becker DL, Dux L, Stelkovics E, Krenacs L, Bagdi E, Krenacs T
Histochemistry and cell biology 2005 Jun;123(6):573-83
Histochemistry and cell biology 2005 Jun;123(6):573-83
Connexin43 with a cytoplasmic loop deletion inhibits the function of several connexins.
Wang M, Martínez AD, Berthoud VM, Seul KH, Gemel J, Valiunas V, Kumari S, Brink PR, Beyer EC
Biochemical and biophysical research communications 2005 Aug 12;333(4):1185-93
Biochemical and biophysical research communications 2005 Aug 12;333(4):1185-93
Connexin43 and connexin26 form gap junctions, but not heteromeric channels in co-expressing cells.
Gemel J, Valiunas V, Brink PR, Beyer EC
Journal of cell science 2004 May 15;117(Pt 12):2469-80
Journal of cell science 2004 May 15;117(Pt 12):2469-80
Overexpression of estrogen receptor-alpha gene suppresses gap junctional intercellular communication in endometrial carcinoma cells.
Saito T, Tanaka R, Wataba K, Kudo R, Yamasaki H
Oncogene 2004 Feb 5;23(5):1109-16
Oncogene 2004 Feb 5;23(5):1109-16
Functional domain mapping and selective trans-dominant effects exhibited by Cx26 disease-causing mutations.
Thomas T, Telford D, Laird DW
The Journal of biological chemistry 2004 Apr 30;279(18):19157-68
The Journal of biological chemistry 2004 Apr 30;279(18):19157-68
Sequential changes of connexin32 and connexin26 in ischemia-reperfusion of the liver in rats.
Nakashima Y, Kohno H, EL-Assal ON, Dhar DK, Ono T, Yamanoi A, Nagasue N
Hepatology research : the official journal of the Japan Society of Hepatology 2003 Sep;27(1):67-75
Hepatology research : the official journal of the Japan Society of Hepatology 2003 Sep;27(1):67-75
The relationship between connexins, gap junctions, tissue architecture and tumour invasion, as studied in a novel in vitro model of HPV-16-associated cervical cancer progression.
Aasen T, Hodgins MB, Edward M, Graham SV
Oncogene 2003 Sep 11;22(39):7969-80
Oncogene 2003 Sep 11;22(39):7969-80
Suppression of gap junctional intercellular communication via 5' CpG island methylation in promoter region of E-cadherin gene in endometrial cancer cells.
Nishimura M, Saito T, Yamasaki H, Kudo R
Carcinogenesis 2003 Oct;24(10):1615-23
Carcinogenesis 2003 Oct;24(10):1615-23
Connexin immunoreactivity in glial cells of the rat retina.
Zahs KR, Kofuji P, Meier C, Dermietzel R
The Journal of comparative neurology 2003 Jan 20;455(4):531-46
The Journal of comparative neurology 2003 Jan 20;455(4):531-46
Role of gap junctions in fluid secretion of lacrimal glands.
Walcott B, Moore LC, Birzgalis A, Claros N, Valiunas V, Ott T, Willecke K, Brink PR
American journal of physiology. Cell physiology 2002 Mar;282(3):C501-7
American journal of physiology. Cell physiology 2002 Mar;282(3):C501-7
Cell-specific expression of connexins and evidence of restricted gap junctional coupling between glial cells and between neurons.
Rash JE, Yasumura T, Dudek FE, Nagy JI
The Journal of neuroscience : the official journal of the Society for Neuroscience 2001 Mar 15;21(6):1983-2000
The Journal of neuroscience : the official journal of the Society for Neuroscience 2001 Mar 15;21(6):1983-2000
Cell-specific expression of connexins and evidence of restricted gap junctional coupling between glial cells and between neurons.
Rash JE, Yasumura T, Dudek FE, Nagy JI
The Journal of neuroscience : the official journal of the Society for Neuroscience 2001 Mar 15;21(6):1983-2000
The Journal of neuroscience : the official journal of the Society for Neuroscience 2001 Mar 15;21(6):1983-2000
trans-dominant inhibition of connexin-43 by mutant connexin-26: implications for dominant connexin disorders affecting epidermal differentiation.
Rouan F, White TW, Brown N, Taylor AM, Lucke TW, Paul DL, Munro CS, Uitto J, Hodgins MB, Richard G
Journal of cell science 2001 Jun;114(Pt 11):2105-13
Journal of cell science 2001 Jun;114(Pt 11):2105-13
Connexin 43 hemi channels mediate Ca2+-regulated transmembrane NAD+ fluxes in intact cells.
Bruzzone S, Guida L, Zocchi E, Franco L, De Flora A
FASEB journal : official publication of the Federation of American Societies for Experimental Biology 2001 Jan;15(1):10-12
FASEB journal : official publication of the Federation of American Societies for Experimental Biology 2001 Jan;15(1):10-12
Connexin 26 and basic fibroblast growth factor are expressed primarily in the subpial and subependymal layers in adult brain parenchyma: roles in stem cell proliferation and morphological plasticity?
Mercier F, Hatton GI
The Journal of comparative neurology 2001 Feb 26;431(1):88-104
The Journal of comparative neurology 2001 Feb 26;431(1):88-104
Localization of connexin26 and connexin32 in putative CO(2)-chemosensitive brainstem regions in rat.
Solomon IC, Halat TJ, El-Maghrabi MR, O'Neal MH 3rd
Respiration physiology 2001 Dec;129(1-2):101-21
Respiration physiology 2001 Dec;129(1-2):101-21
Localization of connexin26 and connexin32 in putative CO(2)-chemosensitive brainstem regions in rat.
Solomon IC, Halat TJ, El-Maghrabi MR, O'Neal MH 3rd
Respiration physiology 2001 Dec;129(1-2):101-21
Respiration physiology 2001 Dec;129(1-2):101-21
A role for heterologous gap junctions between melanoma and endothelial cells in metastasis.
Ito A, Katoh F, Kataoka TR, Okada M, Tsubota N, Asada H, Yoshikawa K, Maeda S, Kitamura Y, Yamasaki H, Nojima H
The Journal of clinical investigation 2000 May;105(9):1189-97
The Journal of clinical investigation 2000 May;105(9):1189-97
Transport function and hepatocellular localization of mrp6 in rat liver.
Madon J, Hagenbuch B, Landmann L, Meier PJ, Stieger B
Molecular pharmacology 2000 Mar;57(3):634-41
Molecular pharmacology 2000 Mar;57(3):634-41
Biochemical and functional characterization of intercellular adhesion and gap junctions in fibroblasts.
Ko K, Arora P, Lee W, McCulloch C
American journal of physiology. Cell physiology 2000 Jul;279(1):C147-57
American journal of physiology. Cell physiology 2000 Jul;279(1):C147-57
Biochemical and functional characterization of intercellular adhesion and gap junctions in fibroblasts.
Ko K, Arora P, Lee W, McCulloch C
American journal of physiology. Cell physiology 2000 Jul;279(1):C147-57
American journal of physiology. Cell physiology 2000 Jul;279(1):C147-57
No comments: Submit comment
Supportive validation
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
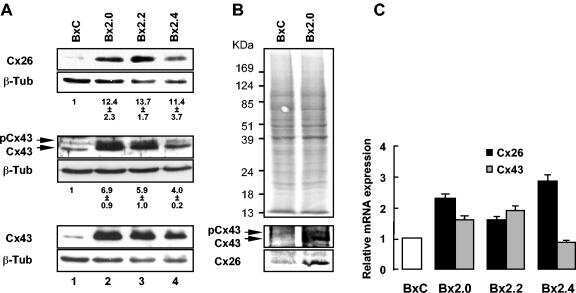
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
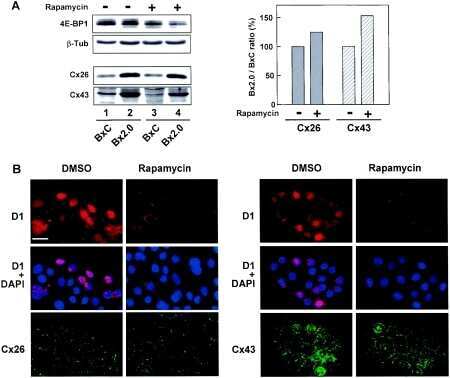
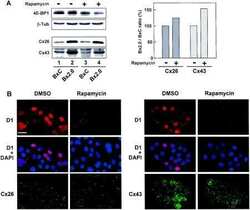
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL

- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
- NULL

- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
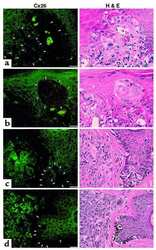
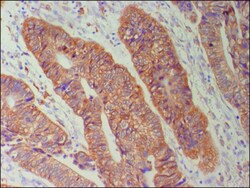
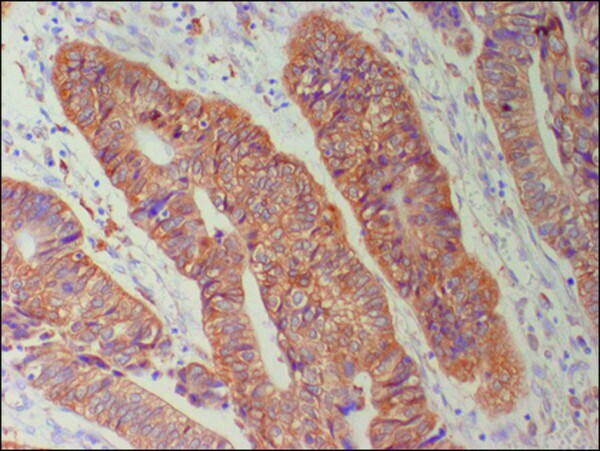
- Figure 1 Immunohistochemical staing for Cx26 in colorectal cancer. Cytoplasmic Cx26 expression was found (x200) .
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
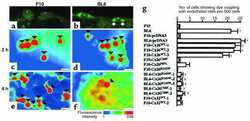
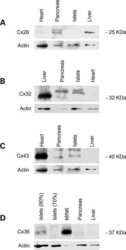
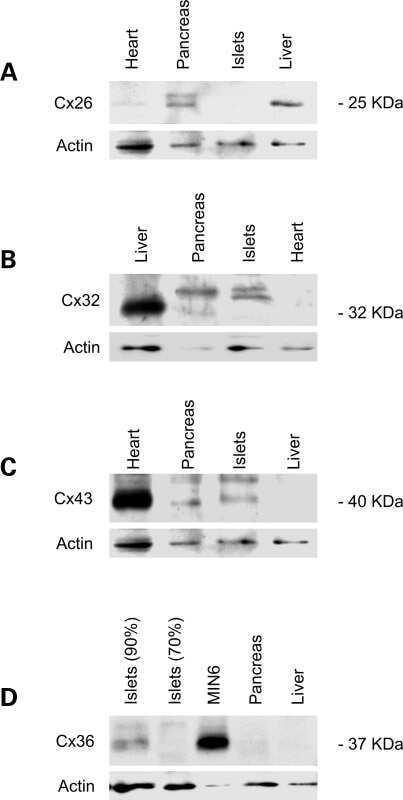
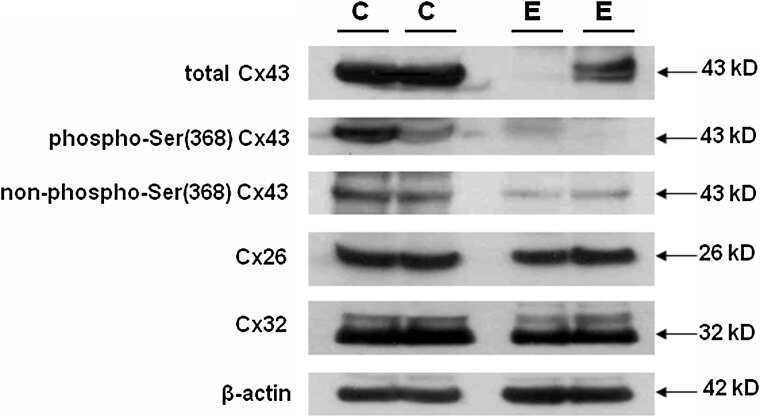
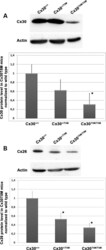
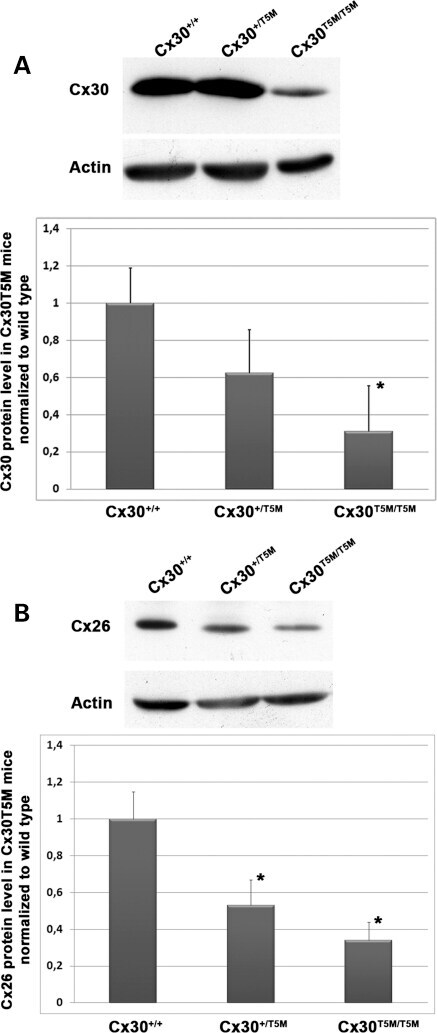
- Figure 4. Quantitative immunoblot analyses of connexin expression in the adult cochlea. Protein levels were normalized to their corresponding actin bands for quantification. Asterisk in ( A ) indicates a significant reduction in Cx30 in Cx30 T5M/T5M mice compared with Cx30 protein level in wild-type mice. Asterisks in ( B ) indicate a significant reduction in Cx26 in Cx30 T5M/T5M mice compared with Cx26 protein level in wild-type mice.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
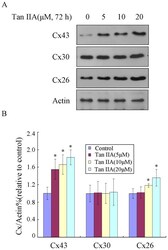
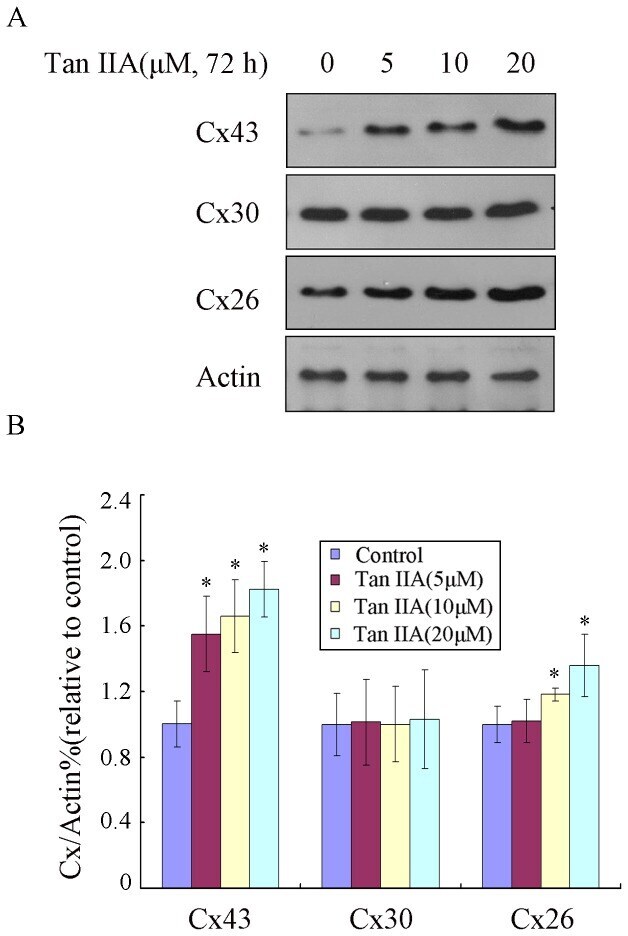
- Figure 1 Tan IIA treatment of B16 cells results in the upregulation of Cx26 and Cx43 proteins. B16 cells were treated with Tan IIA (0, 5, 10 or 20 uM) for 72 h. (A) Immunoblotting was performed using antibody against Cx26, Cx30 and Cx43. Actin was also tested as a loading control. (B) Relative quantification of the immunoblotting results as calculated by gray scanning (*p
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
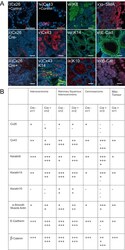
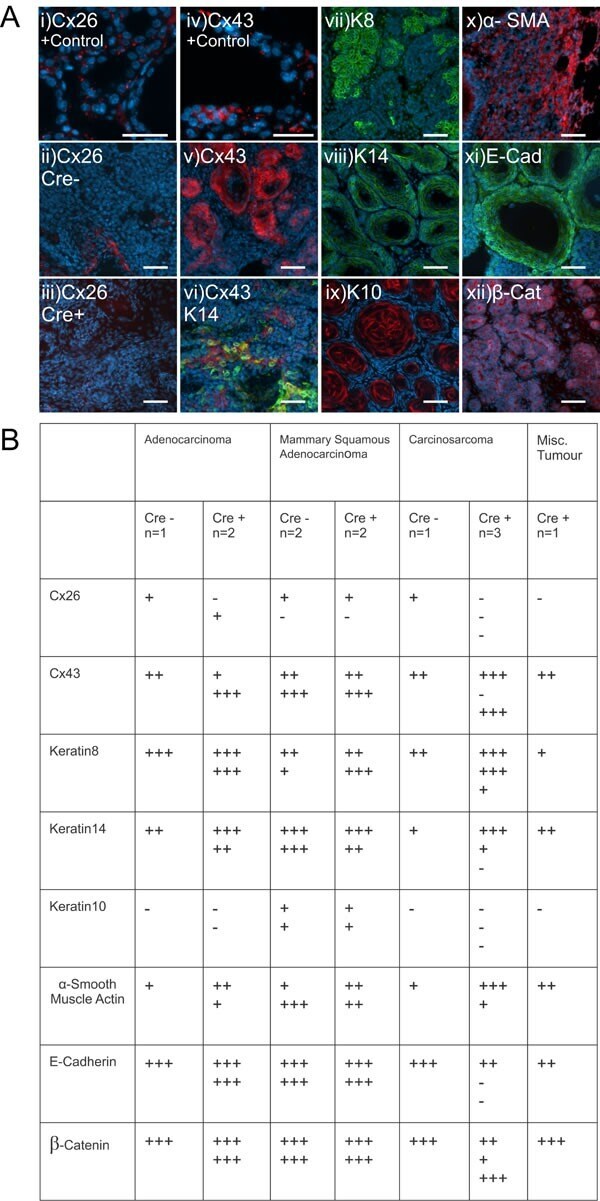
- Figure 5 Mammary tumors from Cx26 knockout mice express similar epithelial protein markers as control mice A. Representative images of paraffin-embedded primary mammary tumor sections immunolabelled with luminal and myoepithelial markers that included; Cx26 (i, ii, iii, red), Cx43 (iv, v, vi, red), keratin 14 (vi, viii green), keratin 8 (vii, green), keratin 10 (ix, red), alpha-smooth muscle actin (x, red), E-cadherin (xi, green) and beta-catenin (xii, red). Hoechst denotes nuclei. Scale bars=50 mum. B. Table indicates relative number of cells that are positive for the luminal and myoepithelial markers based on immunofluorescent labelling. +++50-100%, ++11-49%, +1-10%, -0%.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
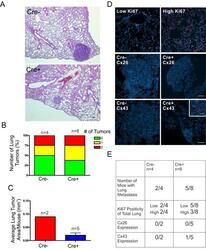
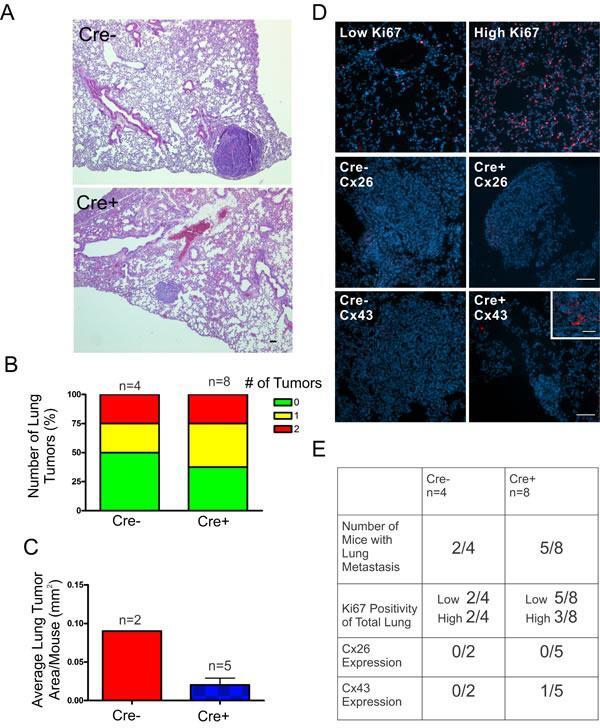
- Figure 6 DMBA-treated Cx26 knockout mice exhibit similar incidence of metastases to the lungs A. , B. Hematoxylin and eosin stained lung sections were evaluated for evidence of lung tumors revealing a similar proportion of mice that developed lung tumors in Cre+ and Cre- mice. C. Evaluation of average lung tumor area per mouse between Cre+ and Cre- mice revealed the likelihood of Cre- mice having larger lung tumor areas compared to Cre+ mice. D. Representative images of paraffin-embedded lung sections immunolabelled for Ki67 (Red), Cx26 (Red) and Cx43 (Red) revealed a similar percentage of lung tissue expressing high levels of Ki67 positivity between Cre- and Cre+ mice and tumors mostly negative, but not always (insert), for Cx26 and Cx43 expression. Hoechst denotes Nuclei. Scale bars = 50 mum. E. Quantification of Ki67, Cx26 and Cx43 immunofluorescent analysis.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
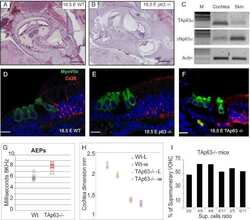
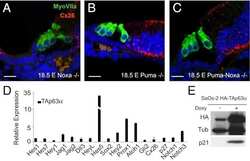
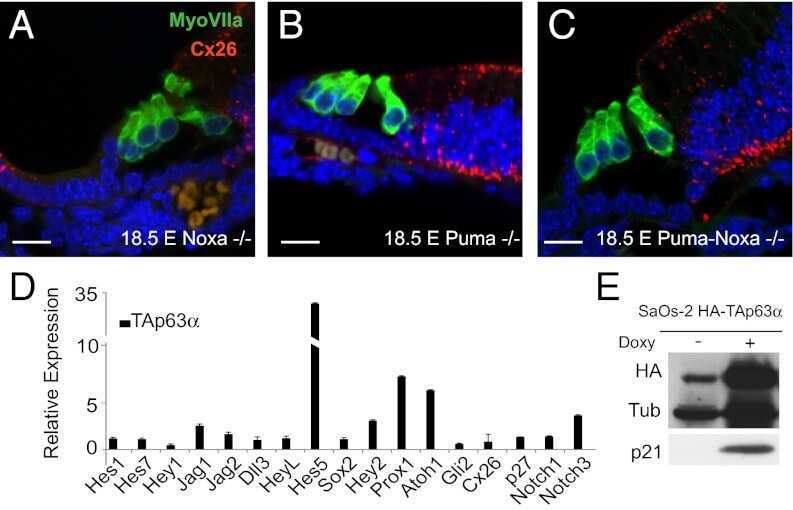
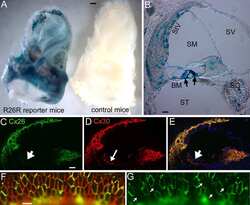
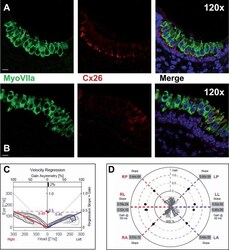
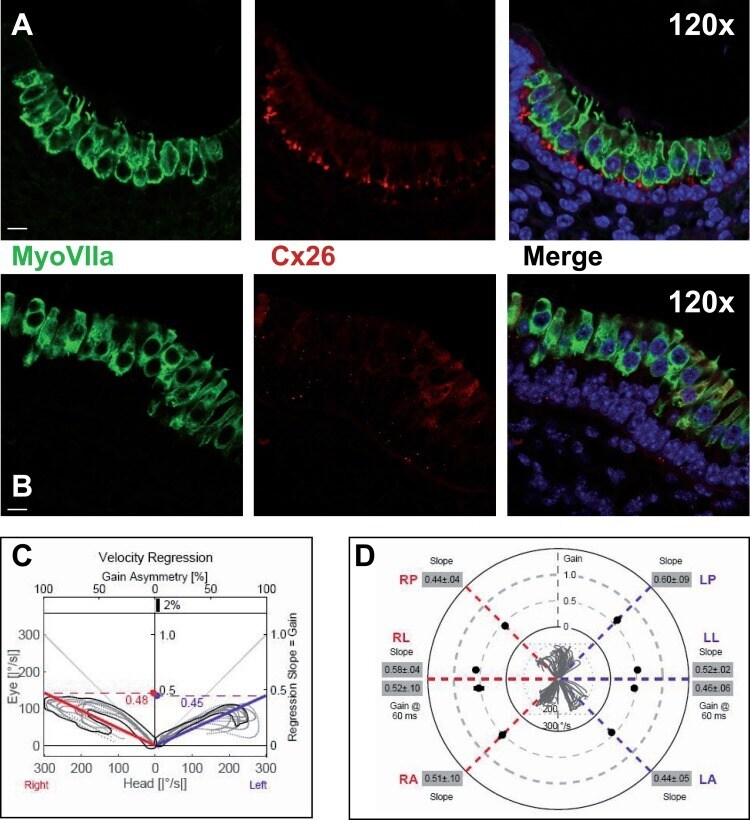
- Figure 3 Macular structure in p63 defective mice and vestibular function in EEC patient ( A ) Representative picture of a macula of the vestibule from mice normal controls stained with MyoVIIa and Connexin 26 antibody. Dapi has been used for nuclei staining. Bars =20 mum. ( B ) Macular neuroepithelium from p63-/- mice. ( C ) Results of vHIT testing of the lateral semicircular canal (LSC) for the patient described in the article; the regression analysis of the vestibulo-ocular reflex (VOR) gain (eye angular velocity/head angular velocity) shows values of 0.48 and 0.45 for the right and left LSC, respectively. This shows VOR hyporeflectivity (average gain between 0.8 and 1.1). ( D ) The vestibular hyporeflectivity for both sides was confirmed by studying VOR gain for all the semicircular canals, with instant gain at 60 milliseconds for the lateral semicircular canals: gain values were all under average, without any significant asymmetry involving one side with respect to the other. Canals positioned on the same plane are connected by lines; RA= right anterior, RP= right posterior, RL=right lateral; LA= left anterior, LP= left posterior, LL=left lateral.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
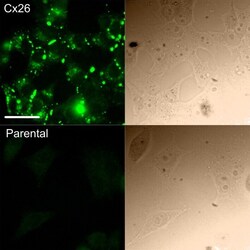
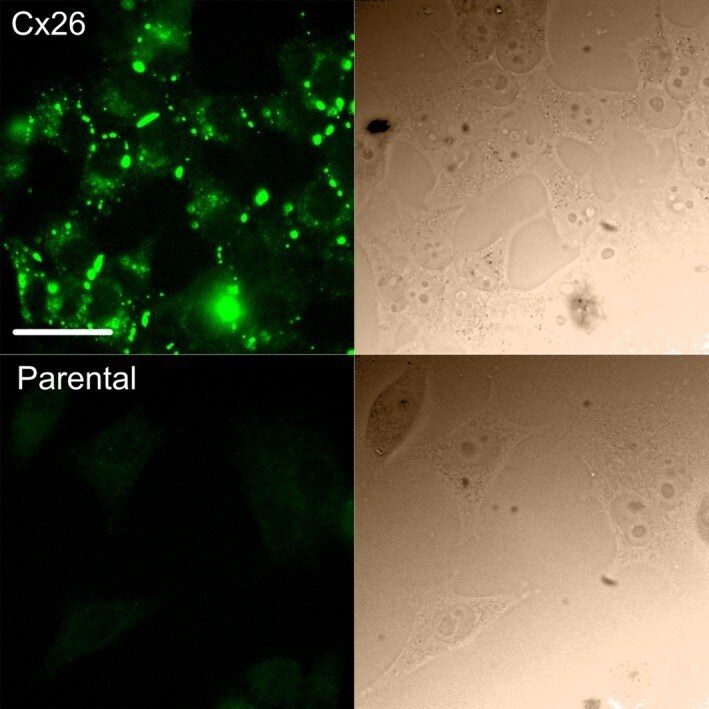
- Figure 1 Expression of Cx26 in HeLa cells. Top row, immunofluorescent staining for Cx26 in HeLa cells stably expressing Cx26. This demonstrates abundant expression of fluorescent puncta representing Cx26 hemichannels and gap junctions. Bottom row, parental HeLa cells do not exhibit any immunofluorescent puncta. In both rows, the corresponding bright-field image is shown on the right. Scale bar 40 mu m.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
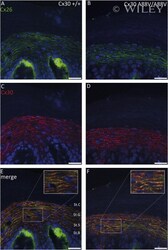
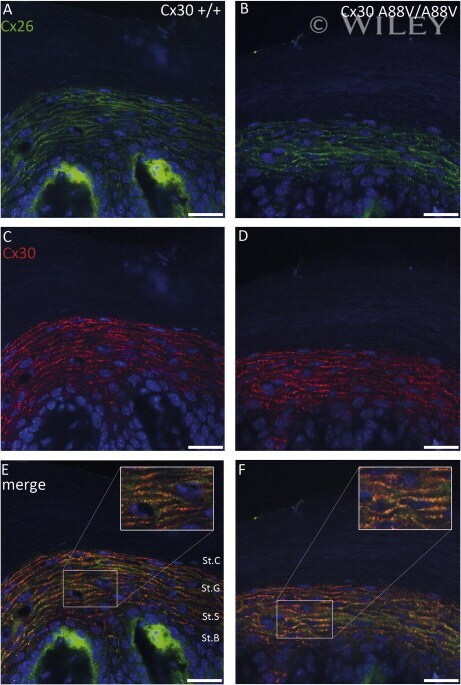
- The Cx30A88V protein forms gap junction plaques in the plasma membrane of the epidermis. Immunostainings of Cx26 in green (A and B) and Cx30 in red (C and D) in sole skin sections of wildtype and homozygous Cx30A88V mice show that both connexins are expressed in the stratum spinosum and stratum granulosum of the thick epidermis. Both wildtype connexins co-localize (yellow staining in the merged images (E and F)). The mutated Cx30A88V also co-localizes with Cx26 in the plasma membrane (detail view in (F)). Nuclei are in blue. Epidermal layers: St.B: stratum basale, St.S: stratum spinosum, StG: stratum granulosum, St.C: stratum corneum. Scale bars: 20 mum.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
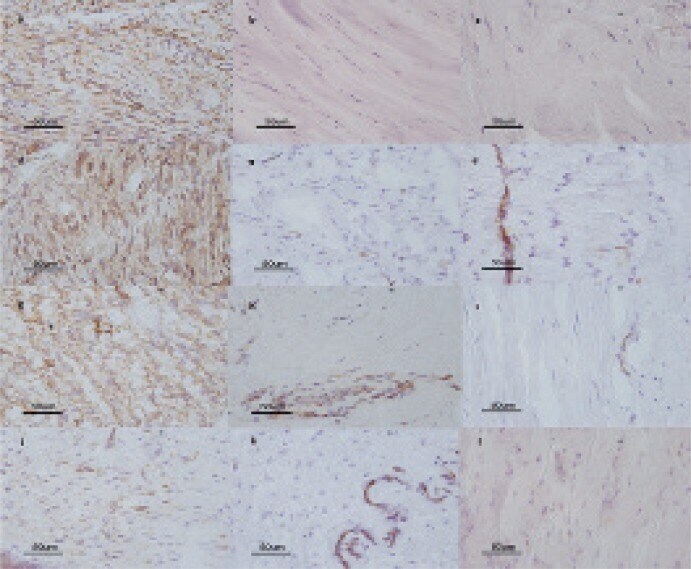
- Immunohistochemical expression of alpha-SMA (a, b, c); Cx26 (d, e, f); Cx43 (g, h, i), and Cx30 (j, k, l) in tissue sections from the involutional phase of DD (a, d, g, j); in tissue sections from the residual phase of DD (b, e, h, k), and in tissue sections from controls (patients with carpal tunnel syndrome) (c, f, i, l) (magnification 400x).
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
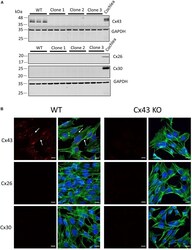
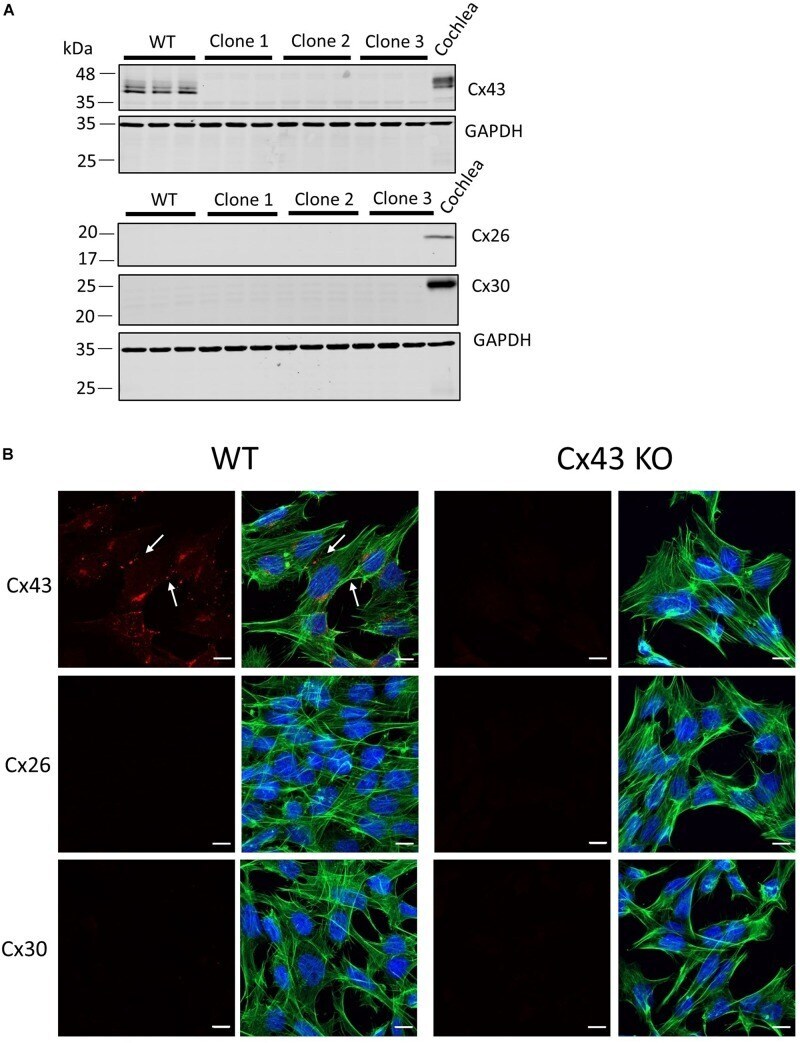
- FIGURE 1 Characterization of connexin expression and Cx43 ablation in HEI-OC1 cells. (A) Western blots for Cx43, Cx26, and Cx30 protein in wild type (WT) and three independent clones of CRISPR-Cas9 Cx43-knockout (Cx43-KO) HEI-OC1 cells where adult mouse cochlear lysate was used as a positive control. Molecular weight standards are denoted in kDa. (B) Immunolabeling revealed Cx43 gap junctions only in WT cells denoted by white arrows. Note Cx26 and Cx30 were not detected in WT or Cx43 KO cells (A,B) . Red = Cx43, Cx26, and Cx30, green = phalloidin staining of actin filaments, blue = Hoechst stained nuclei. Bars = 10 mum.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
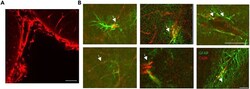
- Figure 5 Representative Examples of Co-localization Imaging for Connexin 26 with a Cell Group of Interest (A) A positive control for successful connexin 26 staining (leptomeninges, red). (B) Co-localization staining of connexin 26 (red; 594) with glial cells (GFAP; green; 488). Images were acquired on a Leica 880 confocal and processed using Zen Black software following the above immunohistochemistry protocol detailed above. Scale bars, 50 mum. Figure reproduced in part using content previously published in ; iScience on a CC-BY 4.0 license.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
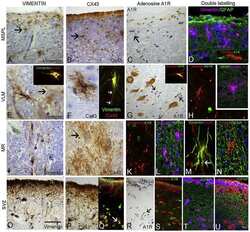
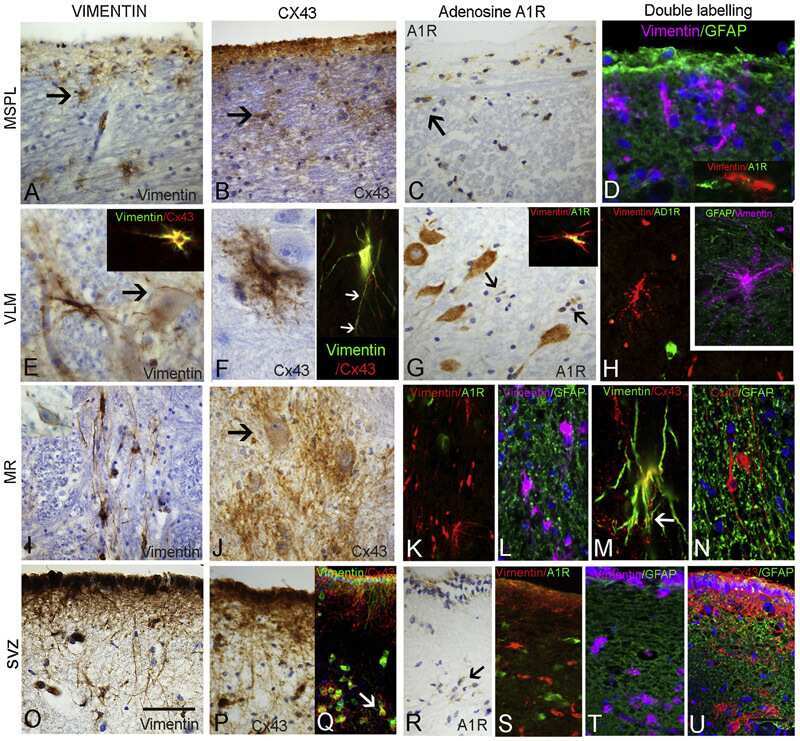
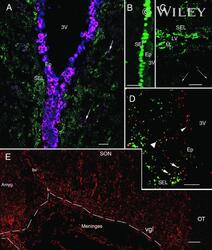
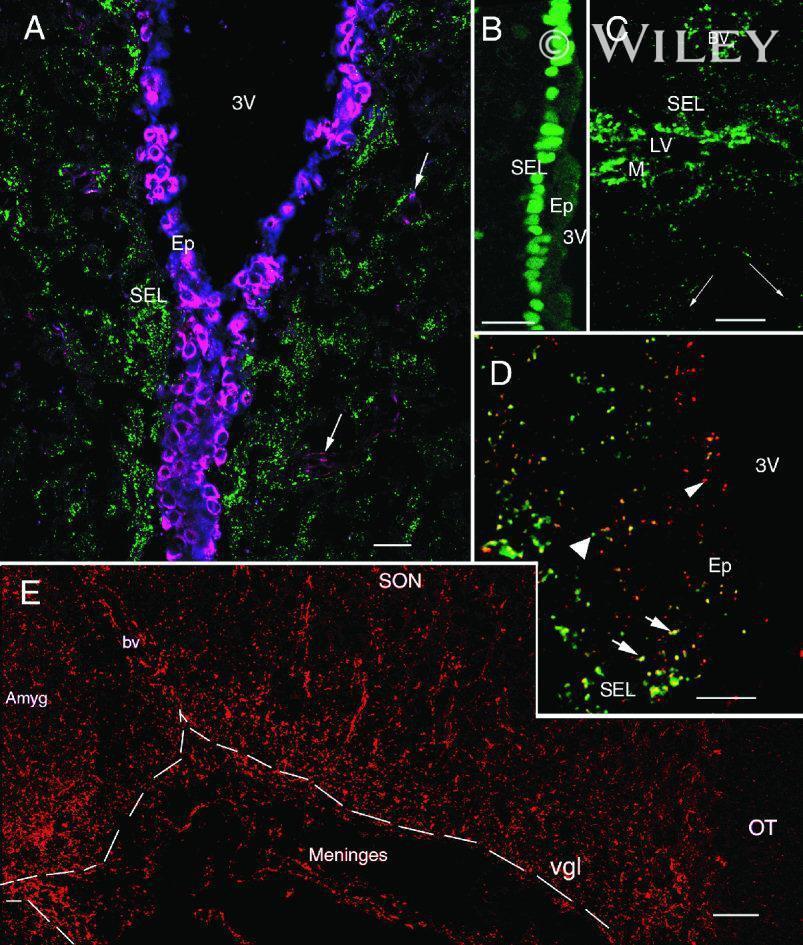
- 4 Distribution of Cx26 in subependymal and subpial zones. A: Intense Cx26-ir punctation (antibody 71-0500; green) in the subependymal layer (SEL) of the third ventricle (3V). Note the decreasing gradient of intensity from the ependymal borderline. Ependymocytes (Ep) and blood vessels (arrows) appear pink due to immunoreactivity for both S100beta (red) and vimentin (blue). B: Intense immunoreactivity for basic fibroblast growth factor (bFGF) in the nuclei of astrocytes in the SEL of 3V. C: The presence of numerous Cx26-ir puncta (antibody 51-2800) in meningeal projections (M) of the lateral ventricle (LV). Puncta were small in the SEL but large on the periphery of blood vessels (BV). Note the lines of small, Cx26-ir puncta along the arrows. D: Immunoreactivity for Cx26 (antibody 51-2800; green) and Cx43 (antibody 13-8300; red) was detected either as double-labeled puncta (arrows) or as single puncta (arrowheads) in the SEL of 3V. The ependyma was primarily Cx43 immunoreactive. E: Distribution of Cx26 immunoreactivity (antibody 13-8100) in the supraoptic nucleus (SON) and underlying meninges. Note the similar concentration of Cx26-ir puncta in the meninges and ventral glial limitans (vgl). White dashed lines indicate the brain surface. Cx26 immunoreactivity was intense in proximity to BV. OT, optic tract; Amyg, amygdala. Scale bars = 20 mum in A; 25 mum in B-D; 50 mum in E.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
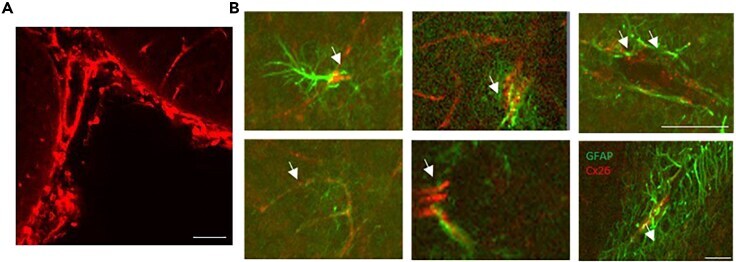
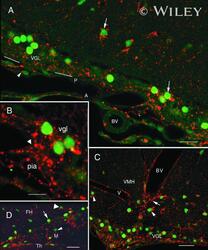
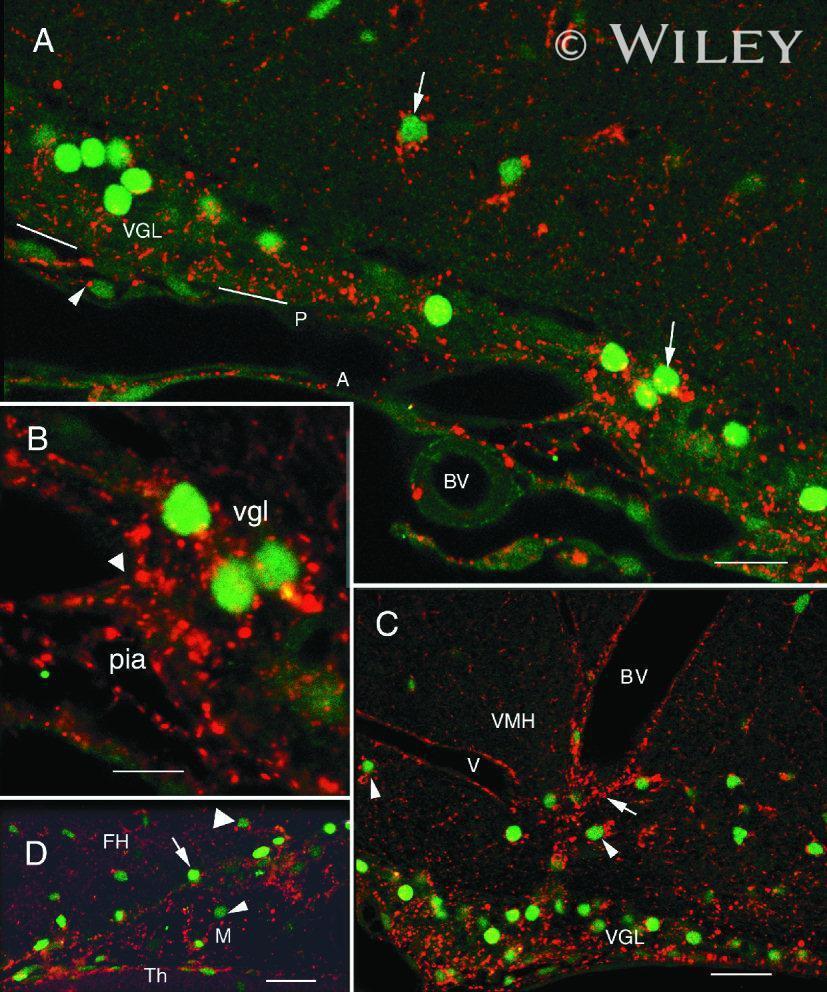
- 6 Association of Cx26 and bFGF in meningeal, subpial, and perivascular layers. All images show Cx26 immunoreactivity (antibody 13-8100; red) and bFGF immunoreactivity (green). A: Section of the periamygdaloid cortex showing an association of Cx26 immunoreactivity and bFGF immunoreactivity in the VGL (arrows), pia (P), and arachnoid (A). White lines indicate the brain surface. Note a Cx26-ir punctum in proximity to the nucleus of a pial fibroblast (arrowhead). BV, blood vessel. B: Magnification of the previous field. Note the continuum of Cx26-ir puncta between the pia and the vgl. An arrowhead indicates the brain surface. C: High concentrations of immunoreactivity to bFGF and Cx26 in the vgl of the ventromedial hypothalamus (VMH) and in the periphery of BV. Note that Cx26-ir puncta were associated (arrowheads) or were not associated (arrow) with nuclear bFGF immunoreactivity. D: Cx26-ir puncta and bFGF immunoreactivity in a meningeal projection (M) between the thalamus (Th) and the fimbria hippocampus (FH). Note the intense nuclear bFGF immunoreactivity in astrocytes that face the meningeal projection (arrow). bFGF immunoreactivity was less intense in astrocytes that were distant from the border zone (large arrowhead) and in fibroblasts (arrowhead). Scale bars = 20 mum in A; 10 mum in B; 25 mum in C,D.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
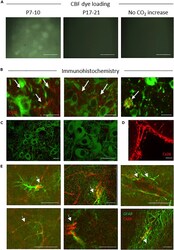
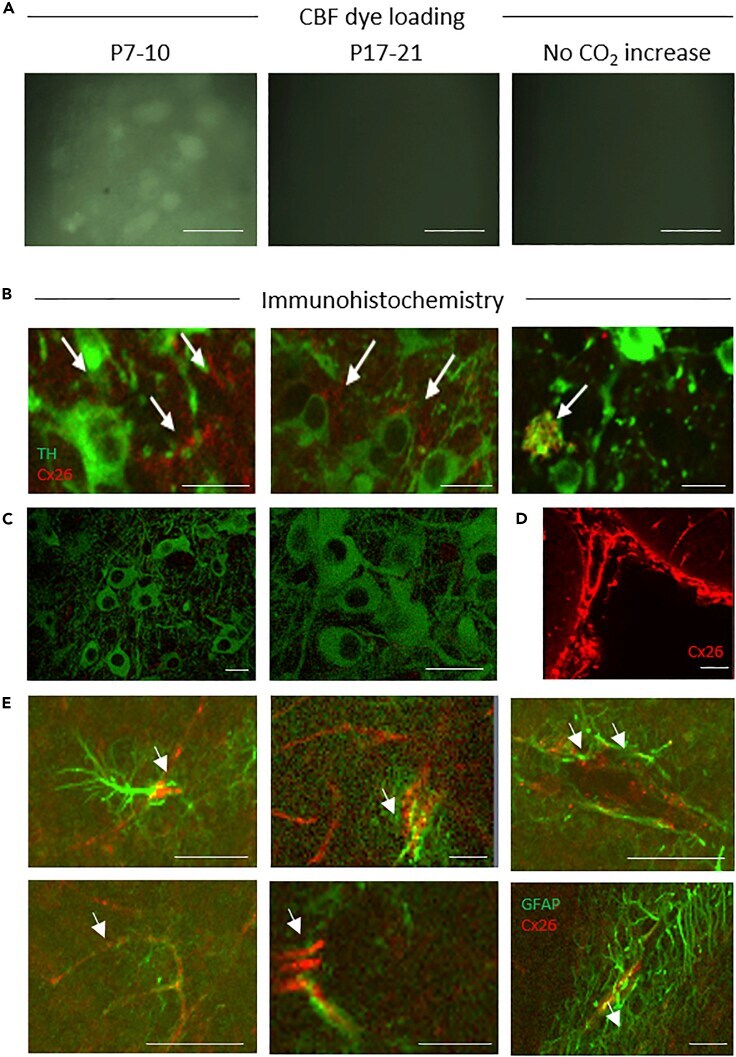
- Figure 3 Dye Loading and Cx26 Expression in Substantia Nigra (A) Carboxyfluorescein (CBF) dye loading following hypercapnia in P7-10 slices (cell bodies are clearly labeled). No dye loading occurred if CO 2 was not changed or in P17-21 slices; scale bar, 50 muM. (B) Immunofluorescent staining of P7-10 SN for Cx26 (red, arrows) in TH + neurons (green). (C) No co-localization of Cx26 (red) in TH + neurons (green) in the SN at P17-21; scale bars, 30 muM. Staining was deemed successful due to the positive leptomeninges staining from corresponding sections of the same brain. (D) Scale bar, 50 muM. (E) Cx26 (red, arrows) co-localized with GFAP (green, glial cell marker) in P17-21 SN; scale bar, 50 muM.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
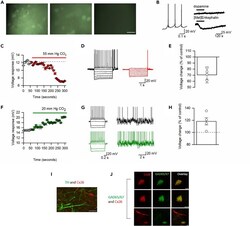
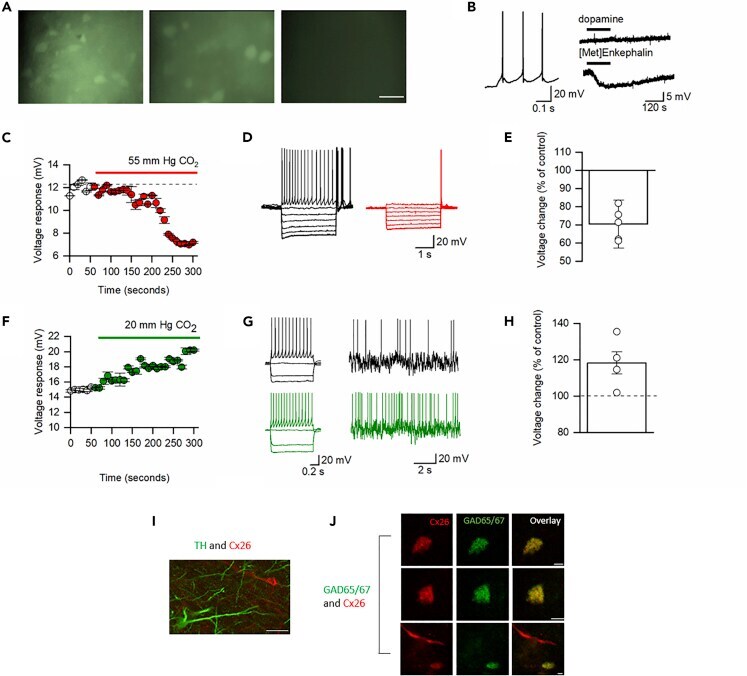
- Figure 4 GABAergic Neurons in the VTA Are Sensitive to CO 2 (A) CBF dye loading of VTA neurons in response to hypercapnia occurs at both P7-10 and P17-21 but does not occur without the increase in CO 2 (hypercapnia), scale bar, 50 muM. (B) Characteristics of CO 2 -sensitive VTA neurons: firing pattern, hyperpolarization to the opioid receptor agonist [Met5]Enkephalin (10 muM) but not to dopamine (30 muM). (C) Time course of changes in voltage response (CO 2 increased from 35 to 55 mm Hg, each point is a mean of six current steps, error bars are SEM). (D) Voltage responses to step currents at indicated time points in (C). (E) Quantification of changes in voltage response to increased CO 2 . (F) Time course of changes in voltage response (CO 2 decreased from 35 to 20 mm Hg, each point is a mean of six current steps, error bars are SEM). (G) Voltage responses to step and fluctuating current inputs (as in ; see Methods ) at indicated time points in (F) demonstrating increased input resistance and firing rate. (H) Quantification of changes in voltage response to decreased CO 2. (I and J) Representative single optical planes immunohistochemistry images. (I) Immunofluorescent staining of P17-21 VTA for Cx26 (red), which is not expressed by TH + neurons (green, no co-localization); scale bar, 50 muM. (J) Co-localization of Cx26 (red) with the soma of three individual GAD + neurons (green) in the VTA (scale bar, 20 muM).
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
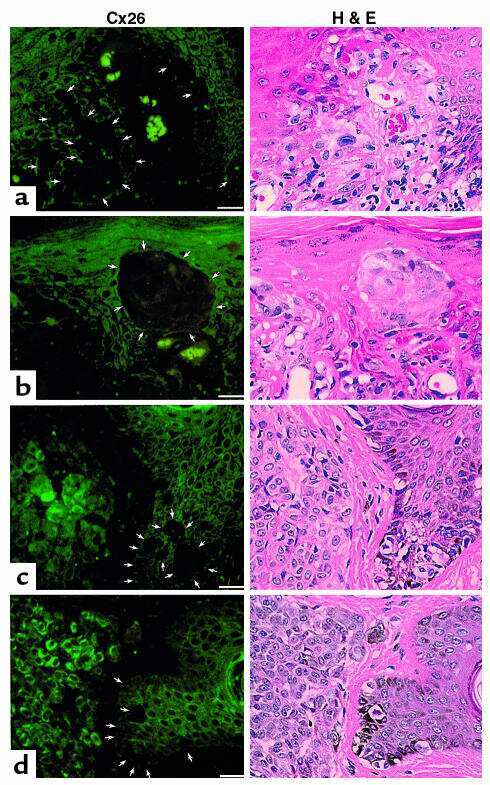
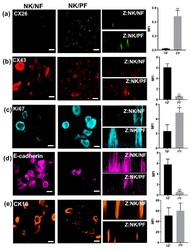
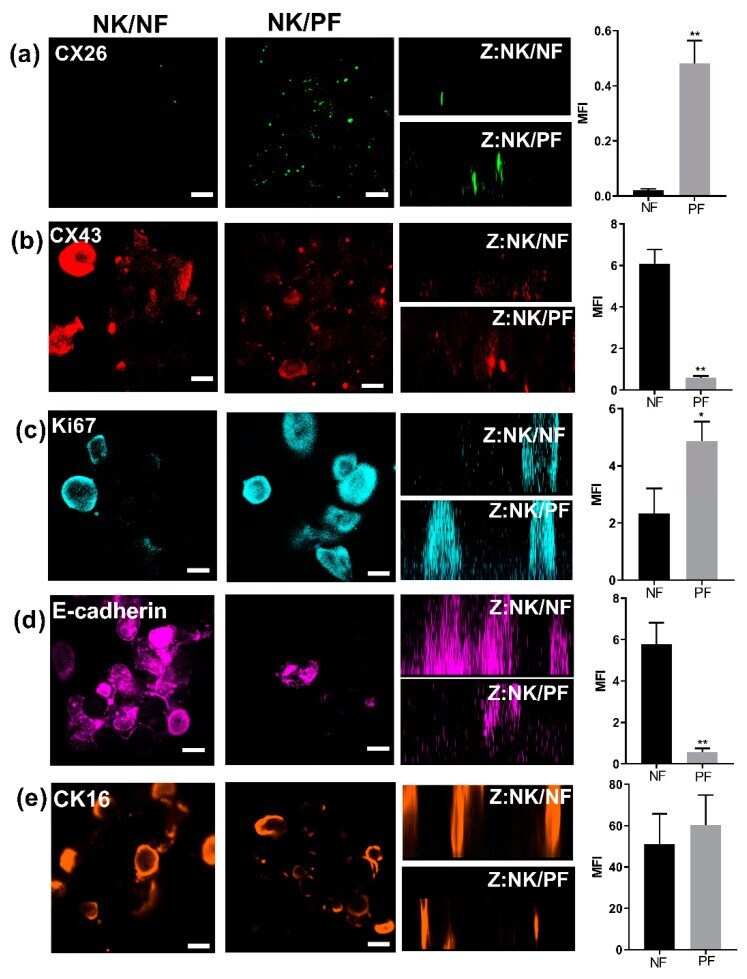
- Figure 5 3D Organotypic epidermal model. Keratinocytes grown in transwells on either a normal (column NK/NF) or on a psoriatic (column NK/PF) fibroblast feeder layer were fixed and stained after 12 days at the ALI with relevant antibodies followed by 3D z-stack reconstruction (Z: NK/NF). IHC analysis determined expression of ( a ) CX26, ( b ) CX43, ( c ) Ki67, ( d ) E-cadherin and ( e ) CK16 protein expression. ( a - e ) bar = 100 mum. The mean fluorescent intensity of multiple regions from different cultures was extracted (right hand panel). Statistical analysis was performed by Student t -test N = 3. * p < 0.05; ** p < 0.001.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
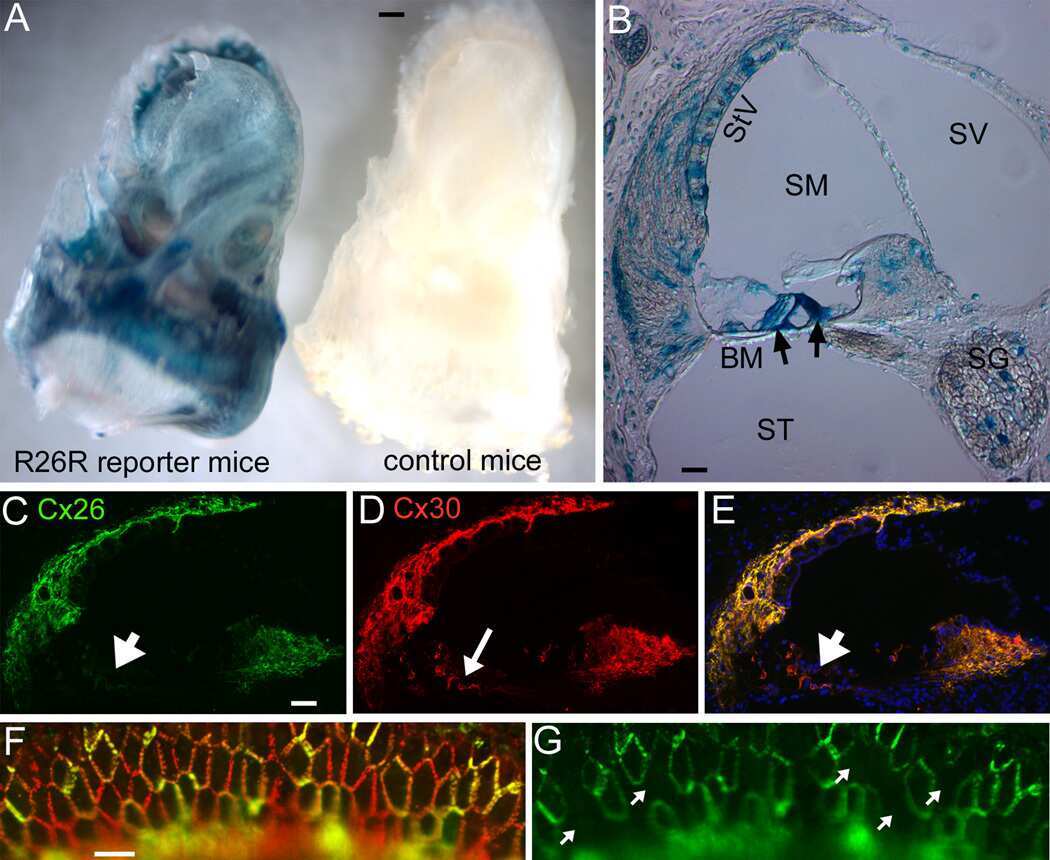
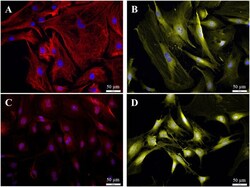
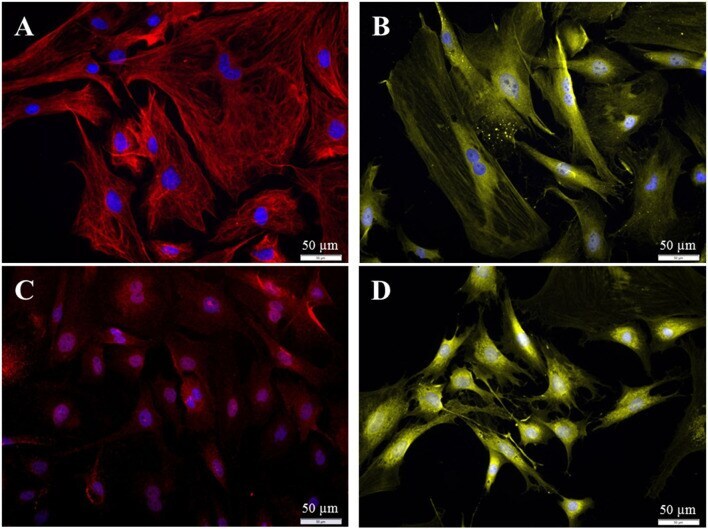
- Immuncytochemical characterization of the purified fibroblasts (P3 after 72 h cultivation). (A) Anti-Vimentin, clone V9 (Cy5, red, 3 s). (B) Anti-S100 (Cy3, yellow, 400 ms). (C) Anti-Cx26 (Cy5, red, 5 s). (D) Anti-carbonic-anhydrase II (Cy3, yellow, 100 ms). All preparations were additionally stained for DAPI (blue, 1 s).
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
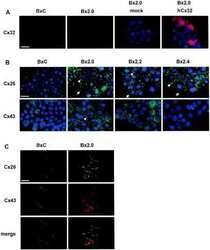
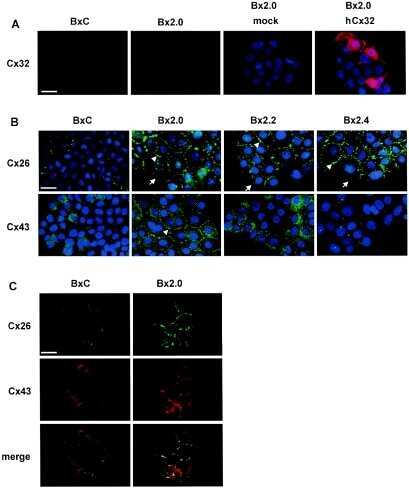
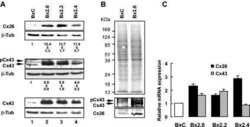
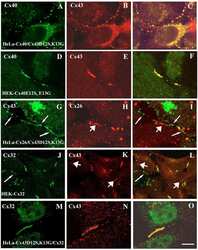
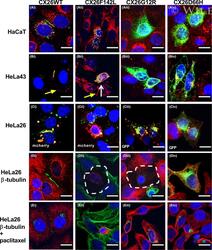
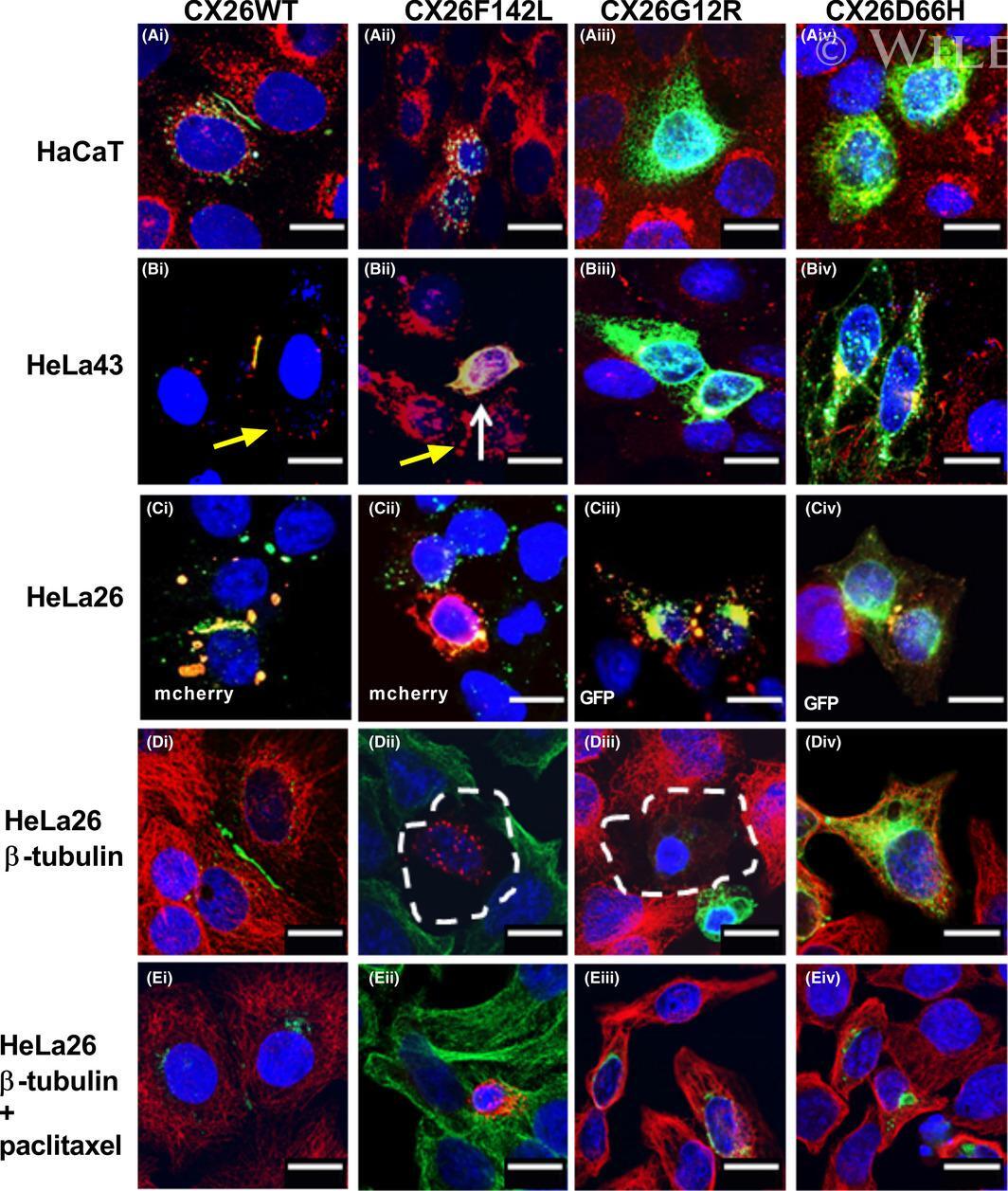
- 2 FIGURE Localization of CX26 mutations and the impact on CX43 and the microtubule network. HaCaT cells (A) and HeLa43 (B) were transfected with CX26WT-GFP (i), CX26F142L-GFP (ii), CX26G12R-GFP (iii) or CX26D66H-GFP (iv) (Cx -green) and co-stained for Cx43 (red). HeLa26 cells (C) were transfected with CX26WT-mCherry (red) (i), CX26F142L-mCherry (red) (ii), CX26G12R-GFP (green) (iii) or CX26D66H-GFP (green) (iv) and co-stained CX26 (stained green in Ci, Cii and red in Ciii and Civ). Areas of co-localization stained yellow. White arrow indicates trapped F142L, and yellow arrow indicates Cx43 Gap Junction staining. HeLa26 cells (D) were transfected with CX26WT-GFP (green) (i), CX26F142L-mCherry (red) (ii), CX26G12R-GFP (green) (iii) or CX26D66H-GFP (green) (iv) and were co-stained for -tubulin (stained red Di, Diii, Div and green in Dii). HeLa26 cells (E) were transfected with CX26WT-GFP(i), CX26F142L-mCherry (ii), CX26G12R-GFP (iii) or CX26D66H-GFP (iv) and treated with paclitaxel throughout the transfection period followed by co-staining with beta-tubulin (stained red Ei, Eiii, Eiv and green in Eii). Nuclei were stained with DAPI (blue). Scale bar = 10 um
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
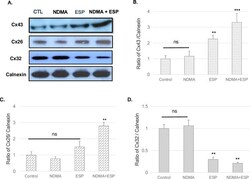
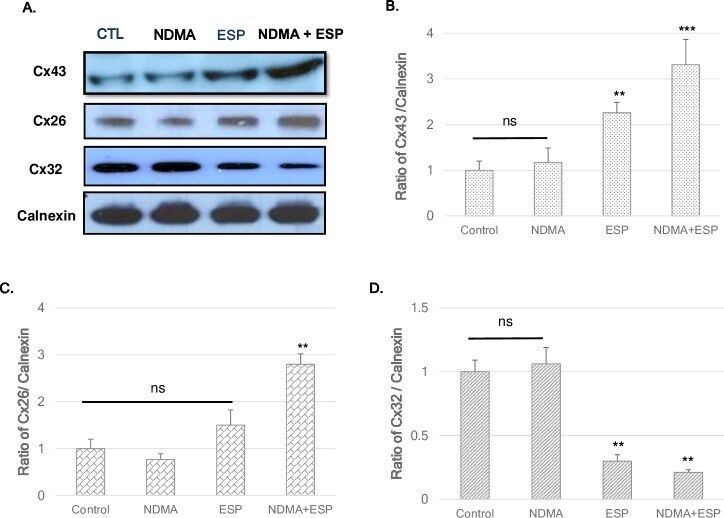
- Fig 3 Expression of the gap-junction proteins connexin 26, connexin 32, and connexin 43 in H69 cells after treatment with NDMA and/or ESP, as determined by western blotting. H69 cells were incubated with either PBS (vehicle) or NDMA and/or ESP for 72 h, and the cells were collected for protein extraction. A. The blots of each groups were run under same experimental conditions and the images were cropped from different parts of the same gels. B-D. Quantification of relative Cx43 Cx26, and Cx32 expression in each group. The each proteins level are indicated as normalization of the ratio of Cx43/Calnexin, Cx26/Calnexin, and Cx32/Calnexin, and Cox-2/Calnexin. DATA represent the mean +- SE of five independent experiments. * P < 0.05, ** P < 0.01 and *** P < 0.001 versus control.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
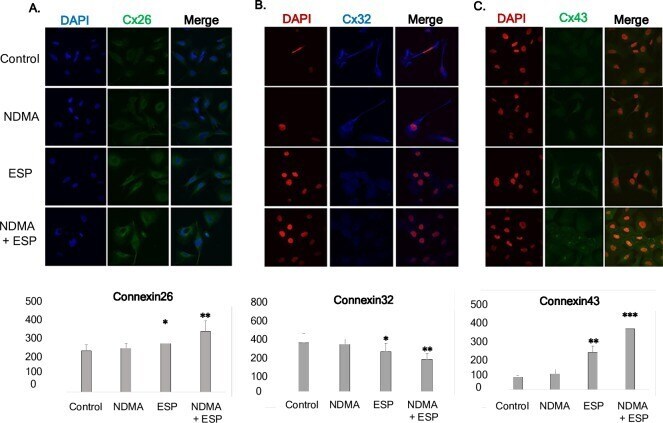
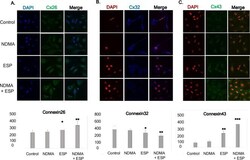
- Fig 4 Concentration of intracellular connexin 26, connexin 32, and connexin 43 in human cholangiocytes (H69 cells). After treatment with NDMA and ESP for 72 h, the concentration of intracellular connexin 26, 32, and 43 in H69 cells was measured by laser scanning microscopy (x4,000). A-C: Immunofluorescence confocal microscopy indicated that the intracellular concentrations of Cx26 (green)/DAPI (blue), Cx32 (blue)/ DAPI (red), and Cx43 (green)/ DAPI (red) increased in cells treated with NDMA + ESP. Scale bar = 25 mum. Data represent the mean +- SE of five independent experiments. * P < 0.05, ** P < 0.01 and *** P < 0.001 versus control.