 Explore
Explore Validate
Validate Learn
Learn Western blot
Western blot Other assay
Other assayAntibody data
- Antibody Data
- Antigen structure
- References [41]
- Comments [0]
- Validations
- Other assay [17]
Submit
Validation data
Reference
Comment
Report error
- Product number
- 13-8200 - Provider product page

- Provider
- Invitrogen Antibodies
- Product name
- Connexin 32 Monoclonal Antibody (CX-2C2)
- Antibody type
- Monoclonal
- Antigen
- Synthetic peptide
- Reactivity
- Human, Mouse, Rat
- Host
- Mouse
- Isotype
- IgG
- Antibody clone number
- CX-2C2
- Vial size
- 100 µg
- Concentration
- 0.5 mg/mL
- Storage
- -20°C
Submitted references Assembly and Function of a Bioengineered Human Liver for Transplantation Generated Solely from Induced Pluripotent Stem Cells.
Haploinsufficient TNAP Mice Display Decreased Extracellular ATP Levels and Expression of Pannexin-1 Channels.
On the adhesion-cohesion balance and oxygen consumption characteristics of liver organoids.
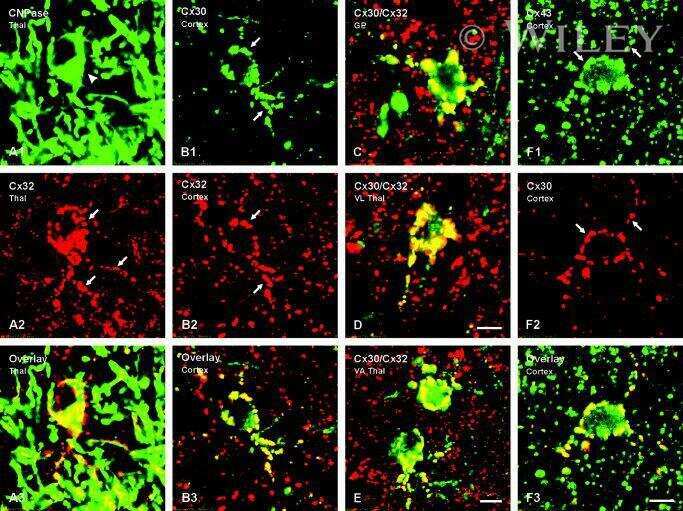
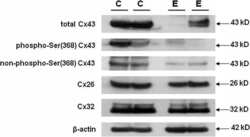
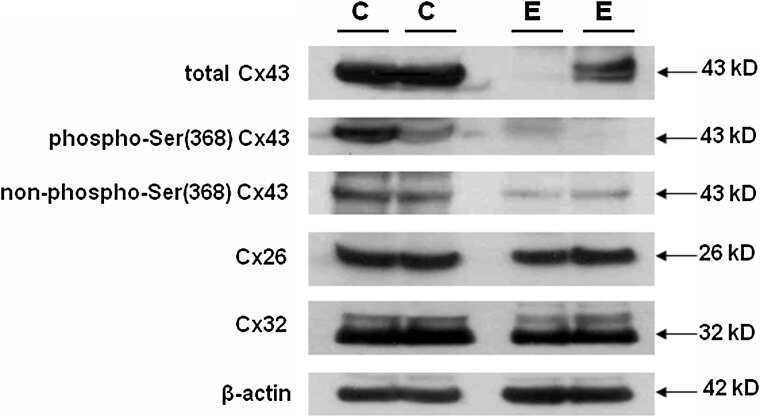
Differential expression of astrocytic connexins in a mouse model of prenatal alcohol exposure.
Cadherin expression, vectorial active transport, and metallothionein isoform 3 mediated EMT/MET responses in cultured primary and immortalized human proximal tubule cells.
Primary tumor- and metastasis-derived colon cancer cells differently modulate connexin expression and function in human capillary endothelial cells.
Maternal treatment with glucocorticoids modulates gap junction protein expression in the ovine fetal brain.
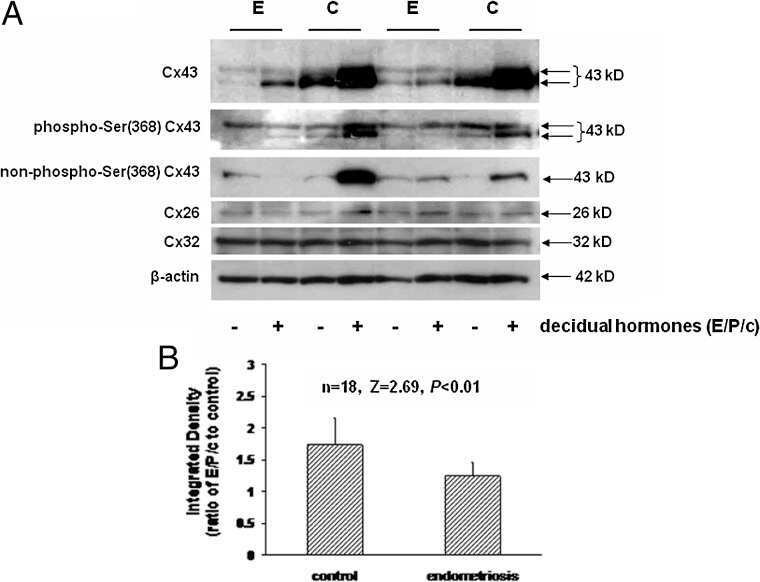
Reduced connexin 43 in eutopic endometrium and cultured endometrial stromal cells from subjects with endometriosis.
Nf1 loss and Ras hyperactivation in oligodendrocytes induce NOS-driven defects in myelin and vasculature.
The liver connexin32 interactome is a novel plasma membrane-mitochondrial signaling nexus.
Semi-automatic counting of connexin 32s immunolocalized in cultured fetal rat hepatocytes using image processing.
Connexin32 can restore hearing in connexin26 deficient mice.
Functional heterotypic interactions between astrocyte and oligodendrocyte connexins.
Cytoplasmic accumulation of connexin32 expands cancer stem cell population in human HuH7 hepatoma cells by enhancing its self-renewal.
Connexin32 is expressed in vascular endothelial cells and participates in gap-junction intercellular communication.
Tissue-specific cross-reactivity of connexin32 antibodies: problems and solutions unique to the central nervous system.
Neuronal connexin expression in the cochlear nucleus of big brown bats.
Impaired gap junction formation and intercellular calcium signaling in urinary bladder cancer cells can be improved by Gö6976.
Gene expression profiling of extracellular matrix as an effector of human hepatocyte phenotype in primary cell culture.
Gap junctional intercellular communication capacity by gap-FRAP technique: a comparative study.
Cytoplasmic accumulation of connexin32 protein enhances motility and metastatic ability of human hepatoma cells in vitro and in vivo.
Gap junction protein connexin 43 serves as a negative marker for a stem cell-containing population of human limbal epithelial cells.
Contacts and cooperation between cells depend on the hormone ouabain.
Astroblastoma: immunohistochemical and ultrastructural study of distinctive epithelial and probable tanycytic differentiation.
Astroblastoma: immunohistochemical and ultrastructural study of distinctive epithelial and probable tanycytic differentiation.
Restoration of functional gap junctions through internal ribosome entry site-dependent synthesis of endogenous connexins in density-inhibited cancer cells.
Transient upregulation of connexin43 gap junctions and synchronized cell cycle control precede myoblast fusion in regenerating skeletal muscle in vivo.
Overexpression of estrogen receptor-alpha gene suppresses gap junctional intercellular communication in endometrial carcinoma cells.
Suppression of gap junctional intercellular communication via 5' CpG island methylation in promoter region of E-cadherin gene in endometrial cancer cells.
Connexin immunoreactivity in glial cells of the rat retina.
Coupling of astrocyte connexins Cx26, Cx30, Cx43 to oligodendrocyte Cx29, Cx32, Cx47: Implications from normal and connexin32 knockout mice.
Expression of connexin 43 and connexin 32 gap-junction proteins in epilepsy-associated brain tumors and in the perilesional epileptic cortex.
Expression of connexin 43 and connexin 32 gap-junction proteins in epilepsy-associated brain tumors and in the perilesional epileptic cortex.
Cell-specific expression of connexins and evidence of restricted gap junctional coupling between glial cells and between neurons.
Cell-specific expression of connexins and evidence of restricted gap junctional coupling between glial cells and between neurons.
Localization of connexin26 and connexin32 in putative CO(2)-chemosensitive brainstem regions in rat.
Localization of connexin26 and connexin32 in putative CO(2)-chemosensitive brainstem regions in rat.
Immunogold evidence that neuronal gap junctions in adult rat brain and spinal cord contain connexin-36 but not connexin-32 or connexin-43.
Immunogold evidence that neuronal gap junctions in adult rat brain and spinal cord contain connexin-36 but not connexin-32 or connexin-43.
Biochemical and functional characterization of intercellular adhesion and gap junctions in fibroblasts.
Biochemical and functional characterization of intercellular adhesion and gap junctions in fibroblasts.
Takeishi K, Collin de l'Hortet A, Wang Y, Handa K, Guzman-Lepe J, Matsubara K, Morita K, Jang S, Haep N, Florentino RM, Yuan F, Fukumitsu K, Tobita K, Sun W, Franks J, Delgado ER, Shapiro EM, Fraunhoffer NA, Duncan AW, Yagi H, Mashimo T, Fox IJ, Soto-Gutierrez A
Cell reports 2020 Jun 2;31(9):107711
Cell reports 2020 Jun 2;31(9):107711
Haploinsufficient TNAP Mice Display Decreased Extracellular ATP Levels and Expression of Pannexin-1 Channels.
Sebastián-Serrano Á, de Diego-García L, Henshall DC, Engel T, Díaz-Hernández M
Frontiers in pharmacology 2018;9:170
Frontiers in pharmacology 2018;9:170
On the adhesion-cohesion balance and oxygen consumption characteristics of liver organoids.
Mattei G, Magliaro C, Giusti S, Ramachandran SD, Heinz S, Braspenning J, Ahluwalia A
PloS one 2017;12(3):e0173206
PloS one 2017;12(3):e0173206
Differential expression of astrocytic connexins in a mouse model of prenatal alcohol exposure.
Ramani M, Mylvaganam S, Krawczyk M, Wang L, Zoidl C, Brien J, Reynolds JN, Kapur B, Poulter MO, Zoidl G, Carlen PL
Neurobiology of disease 2016 Jul;91:83-93
Neurobiology of disease 2016 Jul;91:83-93
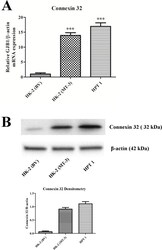
Cadherin expression, vectorial active transport, and metallothionein isoform 3 mediated EMT/MET responses in cultured primary and immortalized human proximal tubule cells.
Slusser A, Bathula CS, Sens DA, Somji S, Sens MA, Zhou XD, Garrett SH
PloS one 2015;10(3):e0120132
PloS one 2015;10(3):e0120132
Primary tumor- and metastasis-derived colon cancer cells differently modulate connexin expression and function in human capillary endothelial cells.
Thuringer D, Berthenet K, Cronier L, Solary E, Garrido C
Oncotarget 2015 Oct 6;6(30):28800-15
Oncotarget 2015 Oct 6;6(30):28800-15
Maternal treatment with glucocorticoids modulates gap junction protein expression in the ovine fetal brain.
Sadowska GB, Stonestreet BS
Neuroscience 2014 Sep 5;275:248-58
Neuroscience 2014 Sep 5;275:248-58
Reduced connexin 43 in eutopic endometrium and cultured endometrial stromal cells from subjects with endometriosis.
Yu J, Boicea A, Barrett KL, James CO, Bagchi IC, Bagchi MK, Nezhat C, Sidell N, Taylor RN
Molecular human reproduction 2014 Mar;20(3):260-70
Molecular human reproduction 2014 Mar;20(3):260-70
Nf1 loss and Ras hyperactivation in oligodendrocytes induce NOS-driven defects in myelin and vasculature.
Mayes DA, Rizvi TA, Titus-Mitchell H, Oberst R, Ciraolo GM, Vorhees CV, Robinson AP, Miller SD, Cancelas JA, Stemmer-Rachamimov AO, Ratner N
Cell reports 2013 Sep 26;4(6):1197-212
Cell reports 2013 Sep 26;4(6):1197-212
The liver connexin32 interactome is a novel plasma membrane-mitochondrial signaling nexus.
Fowler SL, Akins M, Zhou H, Figeys D, Bennett SA
Journal of proteome research 2013 Jun 7;12(6):2597-610
Journal of proteome research 2013 Jun 7;12(6):2597-610
Semi-automatic counting of connexin 32s immunolocalized in cultured fetal rat hepatocytes using image processing.
Takeuchi A, Fukazawa S, Chida K, Taguchi M, Shirataka M, Ikeda N
Acta histochemica 2012 Jul;114(4):318-26
Acta histochemica 2012 Jul;114(4):318-26
Connexin32 can restore hearing in connexin26 deficient mice.
Degen J, Schütz M, Dicke N, Strenzke N, Jokwitz M, Moser T, Willecke K
European journal of cell biology 2011 Oct;90(10):817-24
European journal of cell biology 2011 Oct;90(10):817-24
Functional heterotypic interactions between astrocyte and oligodendrocyte connexins.
Magnotti LM, Goodenough DA, Paul DL
Glia 2011 Jan;59(1):26-34
Glia 2011 Jan;59(1):26-34
Cytoplasmic accumulation of connexin32 expands cancer stem cell population in human HuH7 hepatoma cells by enhancing its self-renewal.
Kawasaki Y, Omori Y, Li Q, Nishikawa Y, Yoshioka T, Yoshida M, Ishikawa K, Enomoto K
International journal of cancer 2011 Jan 1;128(1):51-62
International journal of cancer 2011 Jan 1;128(1):51-62
Connexin32 is expressed in vascular endothelial cells and participates in gap-junction intercellular communication.
Okamoto T, Akiyama M, Takeda M, Gabazza EC, Hayashi T, Suzuki K
Biochemical and biophysical research communications 2009 May 1;382(2):264-8
Biochemical and biophysical research communications 2009 May 1;382(2):264-8
Tissue-specific cross-reactivity of connexin32 antibodies: problems and solutions unique to the central nervous system.
Fowler SL, McLean AC, Bennett SA
Cell communication & adhesion 2009 Dec;16(5-6):117-30
Cell communication & adhesion 2009 Dec;16(5-6):117-30
Neuronal connexin expression in the cochlear nucleus of big brown bats.
Horowitz SS, Stamper SA, Simmons JA
Brain research 2008 Mar 4;1197:76-84
Brain research 2008 Mar 4;1197:76-84
Impaired gap junction formation and intercellular calcium signaling in urinary bladder cancer cells can be improved by Gö6976.
Leinonen P, Aaltonen V, Koskela S, Lehenkari P, Korkiamäki T, Peltonen J
Cell communication & adhesion 2007 Oct;14(4):125-36
Cell communication & adhesion 2007 Oct;14(4):125-36
Gene expression profiling of extracellular matrix as an effector of human hepatocyte phenotype in primary cell culture.
Page JL, Johnson MC, Olsavsky KM, Strom SC, Zarbl H, Omiecinski CJ
Toxicological sciences : an official journal of the Society of Toxicology 2007 Jun;97(2):384-97
Toxicological sciences : an official journal of the Society of Toxicology 2007 Jun;97(2):384-97
Gap junctional intercellular communication capacity by gap-FRAP technique: a comparative study.
Abbaci M, Barberi-Heyob M, Stines JR, Blondel W, Dumas D, Guillemin F, Didelon J
Biotechnology journal 2007 Jan;2(1):50-61
Biotechnology journal 2007 Jan;2(1):50-61
Cytoplasmic accumulation of connexin32 protein enhances motility and metastatic ability of human hepatoma cells in vitro and in vivo.
Li Q, Omori Y, Nishikawa Y, Yoshioka T, Yamamoto Y, Enomoto K
International journal of cancer 2007 Aug 1;121(3):536-46
International journal of cancer 2007 Aug 1;121(3):536-46
Gap junction protein connexin 43 serves as a negative marker for a stem cell-containing population of human limbal epithelial cells.
Chen Z, Evans WH, Pflugfelder SC, Li DQ
Stem cells (Dayton, Ohio) 2006 May;24(5):1265-73
Stem cells (Dayton, Ohio) 2006 May;24(5):1265-73
Contacts and cooperation between cells depend on the hormone ouabain.
Larre I, Ponce A, Fiorentino R, Shoshani L, Contreras RG, Cereijido M
Proceedings of the National Academy of Sciences of the United States of America 2006 Jul 18;103(29):10911-6
Proceedings of the National Academy of Sciences of the United States of America 2006 Jul 18;103(29):10911-6
Astroblastoma: immunohistochemical and ultrastructural study of distinctive epithelial and probable tanycytic differentiation.
Kubota T, Sato K, Arishima H, Takeuchi H, Kitai R, Nakagawa T
Neuropathology : official journal of the Japanese Society of Neuropathology 2006 Feb;26(1):72-81
Neuropathology : official journal of the Japanese Society of Neuropathology 2006 Feb;26(1):72-81
Astroblastoma: immunohistochemical and ultrastructural study of distinctive epithelial and probable tanycytic differentiation.
Kubota T, Sato K, Arishima H, Takeuchi H, Kitai R, Nakagawa T
Neuropathology : official journal of the Japanese Society of Neuropathology 2006 Feb;26(1):72-81
Neuropathology : official journal of the Japanese Society of Neuropathology 2006 Feb;26(1):72-81
Restoration of functional gap junctions through internal ribosome entry site-dependent synthesis of endogenous connexins in density-inhibited cancer cells.
Lahlou H, Fanjul M, Pradayrol L, Susini C, Pyronnet S
Molecular and cellular biology 2005 May;25(10):4034-45
Molecular and cellular biology 2005 May;25(10):4034-45
Transient upregulation of connexin43 gap junctions and synchronized cell cycle control precede myoblast fusion in regenerating skeletal muscle in vivo.
Gorbe A, Becker DL, Dux L, Stelkovics E, Krenacs L, Bagdi E, Krenacs T
Histochemistry and cell biology 2005 Jun;123(6):573-83
Histochemistry and cell biology 2005 Jun;123(6):573-83
Overexpression of estrogen receptor-alpha gene suppresses gap junctional intercellular communication in endometrial carcinoma cells.
Saito T, Tanaka R, Wataba K, Kudo R, Yamasaki H
Oncogene 2004 Feb 5;23(5):1109-16
Oncogene 2004 Feb 5;23(5):1109-16
Suppression of gap junctional intercellular communication via 5' CpG island methylation in promoter region of E-cadherin gene in endometrial cancer cells.
Nishimura M, Saito T, Yamasaki H, Kudo R
Carcinogenesis 2003 Oct;24(10):1615-23
Carcinogenesis 2003 Oct;24(10):1615-23
Connexin immunoreactivity in glial cells of the rat retina.
Zahs KR, Kofuji P, Meier C, Dermietzel R
The Journal of comparative neurology 2003 Jan 20;455(4):531-46
The Journal of comparative neurology 2003 Jan 20;455(4):531-46
Coupling of astrocyte connexins Cx26, Cx30, Cx43 to oligodendrocyte Cx29, Cx32, Cx47: Implications from normal and connexin32 knockout mice.
Nagy JI, Ionescu AV, Lynn BD, Rash JE
Glia 2003 Dec;44(3):205-18
Glia 2003 Dec;44(3):205-18
Expression of connexin 43 and connexin 32 gap-junction proteins in epilepsy-associated brain tumors and in the perilesional epileptic cortex.
Aronica E, Gorter JA, Jansen GH, Leenstra S, Yankaya B, Troost D
Acta neuropathologica 2001 May;101(5):449-59
Acta neuropathologica 2001 May;101(5):449-59
Expression of connexin 43 and connexin 32 gap-junction proteins in epilepsy-associated brain tumors and in the perilesional epileptic cortex.
Aronica E, Gorter JA, Jansen GH, Leenstra S, Yankaya B, Troost D
Acta neuropathologica 2001 May;101(5):449-59
Acta neuropathologica 2001 May;101(5):449-59
Cell-specific expression of connexins and evidence of restricted gap junctional coupling between glial cells and between neurons.
Rash JE, Yasumura T, Dudek FE, Nagy JI
The Journal of neuroscience : the official journal of the Society for Neuroscience 2001 Mar 15;21(6):1983-2000
The Journal of neuroscience : the official journal of the Society for Neuroscience 2001 Mar 15;21(6):1983-2000
Cell-specific expression of connexins and evidence of restricted gap junctional coupling between glial cells and between neurons.
Rash JE, Yasumura T, Dudek FE, Nagy JI
The Journal of neuroscience : the official journal of the Society for Neuroscience 2001 Mar 15;21(6):1983-2000
The Journal of neuroscience : the official journal of the Society for Neuroscience 2001 Mar 15;21(6):1983-2000
Localization of connexin26 and connexin32 in putative CO(2)-chemosensitive brainstem regions in rat.
Solomon IC, Halat TJ, El-Maghrabi MR, O'Neal MH 3rd
Respiration physiology 2001 Dec;129(1-2):101-21
Respiration physiology 2001 Dec;129(1-2):101-21
Localization of connexin26 and connexin32 in putative CO(2)-chemosensitive brainstem regions in rat.
Solomon IC, Halat TJ, El-Maghrabi MR, O'Neal MH 3rd
Respiration physiology 2001 Dec;129(1-2):101-21
Respiration physiology 2001 Dec;129(1-2):101-21
Immunogold evidence that neuronal gap junctions in adult rat brain and spinal cord contain connexin-36 but not connexin-32 or connexin-43.
Rash JE, Staines WA, Yasumura T, Patel D, Furman CS, Stelmack GL, Nagy JI
Proceedings of the National Academy of Sciences of the United States of America 2000 Jun 20;97(13):7573-8
Proceedings of the National Academy of Sciences of the United States of America 2000 Jun 20;97(13):7573-8
Immunogold evidence that neuronal gap junctions in adult rat brain and spinal cord contain connexin-36 but not connexin-32 or connexin-43.
Rash JE, Staines WA, Yasumura T, Patel D, Furman CS, Stelmack GL, Nagy JI
Proceedings of the National Academy of Sciences of the United States of America 2000 Jun 20;97(13):7573-8
Proceedings of the National Academy of Sciences of the United States of America 2000 Jun 20;97(13):7573-8
Biochemical and functional characterization of intercellular adhesion and gap junctions in fibroblasts.
Ko K, Arora P, Lee W, McCulloch C
American journal of physiology. Cell physiology 2000 Jul;279(1):C147-57
American journal of physiology. Cell physiology 2000 Jul;279(1):C147-57
Biochemical and functional characterization of intercellular adhesion and gap junctions in fibroblasts.
Ko K, Arora P, Lee W, McCulloch C
American journal of physiology. Cell physiology 2000 Jul;279(1):C147-57
American journal of physiology. Cell physiology 2000 Jul;279(1):C147-57
No comments: Submit comment
Supportive validation
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
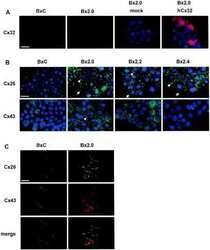
- Experimental details
- NULL
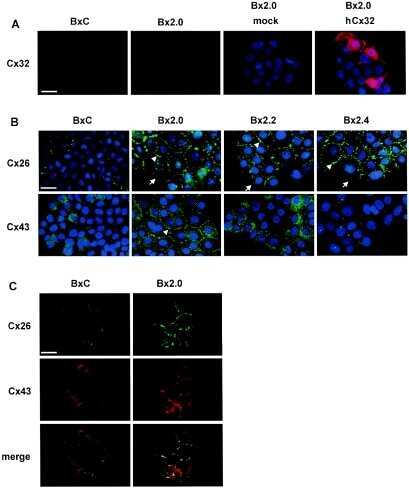
- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
- NULL

- Submitted by
- Invitrogen Antibodies (provider)
- Main image
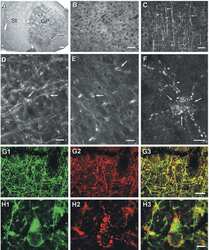
- Experimental details
- NULL

- Submitted by
- Invitrogen Antibodies (provider)
- Main image
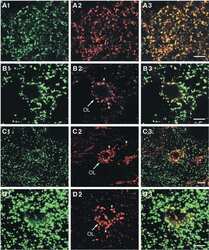
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL

- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
- NULL

- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
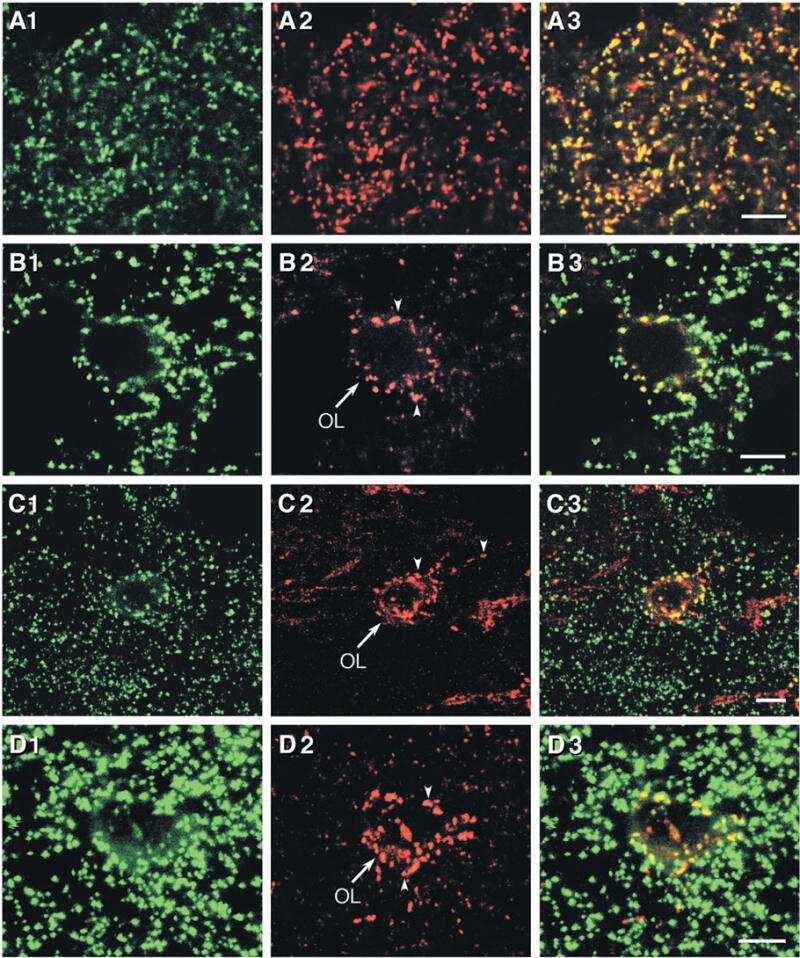
- Figure 1 Nf1 Loss or HRas Activation in Oligodendrocytes Causes Optic-Nerve Enlargement, Myelin Decompaction, Loss of Claudin-11, and Altered Cx32 Localization (A) Gross micrographs of optic nerves at the level of the chiasm in 12-month-old animals. Ruler shows 1 mm markings. (B) Toluidine-blue-stained semithin optic-nerve cross-sections, 1 mm rostral to the chiasm. (C) Optic-nerve g-ratio scatterplots (~5,000 axons/graph; measurements 1 mm rostral to the chiasm in three to five electron micrographs at 10,000x). Red box: 99% of WT g-ratios superimposed upon graphs of other genotypes. ANOVA, Tukey post hoc, p = 2.2 x 10 -16 . (D) Diagram of myelin compaction. Brown circle: axon cross-section; gray: major dense lines (MDLs); orange: Cx32 GJs between compact myelin layers; blue lines: claudin-11+ TJs in compact myelin. (E) Electron micrographs (scale bar, 10 nm) of WT, PLPCre;Nf1 fl / fl , and CNP-HRas optic-nerve myelin. White arrows: intraperiod lines (ILs); black arrows: MDLs. (F-I) Western blots of total optic-nerve lysate or whole-brain cytosolic, endosomal, or myelin fractions showing myelin proteins (F), claudin-11 (G), or Cx32 cellular localization (H and I). n = 3-5 animals/genotype/experiment. See also Figures S1, S2, S3, and S4 .

- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
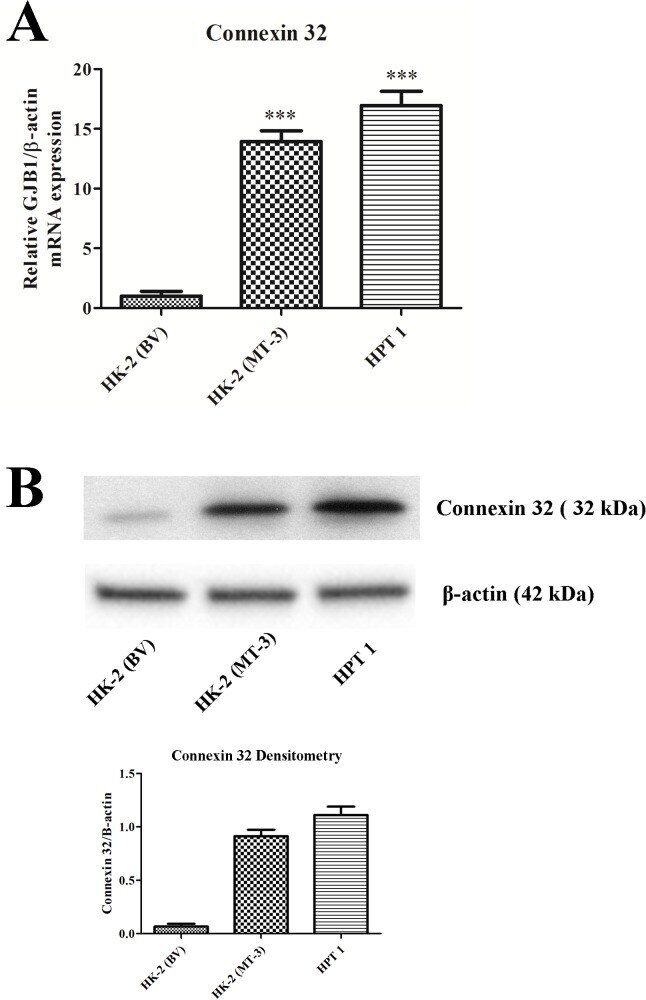
- Fig 8 Connexin 32 expression in HK-2, HPT, and HK-2 cells expressing MT3. Messenger RNA of connexin 32 was assessed with real-time PCR and expressed as a fold increase of the HK-2 cells stably transfected with the blank vector. The change in connexin 32 expression was normalized to the change in beta-actin expression. Western analysis of connexin 32 is shown below the graph.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
- Fig 11 Effect of altered domains of metallothionein-3 on the expression of connexin 32 in HK-2 cells. A) Messenger RNA of connexin 32 assessed with real-time PCR and expressed as fold change in expression versus HK-2 cells transfected with the blank vector. The change in connexin 32 expression was normalized to the change in beta-actin expression. Significant differences from HK-2 (BV) are designated as *** p < 0.0001. B) Western analysis of connexin 32 was conducted in identical cultures as in (A). beta-actin was used as a loading control and for densitometric normalization. Densitometry is shown beside the corresponding blot.

- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
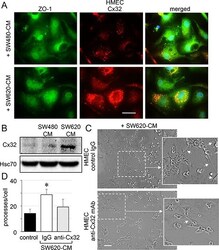
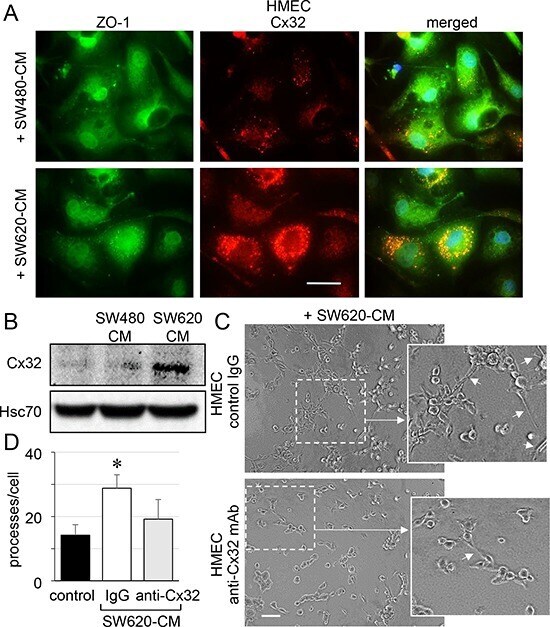
- Figure 4 SW620 cell-secreted factors overexpress the endothelial Cx32 favoring tubulogenesis A. Endothelial cell localization of Cx32 in CRC cell-conditioned media. HMEC were stimulated with SW480-CM or SW620-CM for 6 h and double-stained for ZO-1 and Cx32. Representative micrographs showing the strong labelling of Cx32 induced by SW620-CM and the combined image of co-localization with ZO-1 (yellow); DAPI staining of nuclei ( n = 3, bar 20 mum). B. SW620-CM increase the Cx32 expression in HMEC. A higher Cx32 protein level was detected in response to SW620-CM compared with SW480-CM by immune-blot analysis (no cell expression in unstimulated HMEC). Representative of 5 experiments (Hsc70 as loading control; 150 mug/lane). C-D. In vitro tubulogenesis assay of HMEC pretreated or not (control IgG) with inhibitory monoclonal antibody against Cx32 (anti-Cx32 mAb). HMEC were plated on Matrigel-coated 24-well plates, incubated with SW620-CM for 6 h, and photographed. C. Representative photos of tube formation in HMEC intracellularly delivered with 0.2 mug anti-Cx32 mAb or control IgG (Bar 80 mum). The dotted areas are enlarged in the inserts on the right. Arrows indicated branch points. D. Number of branch points per field of view was quantified (at least 80 single cells were scored; mean +- SD, n = 4; * P < 0.01 vs control).
- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
- Figure 2 Identification of novel Cx32 interactors by IP-HTMS ofCx32-enrichedliver membrane fractions and validation by co-IP of tissue lysates.(A) Silver stain of proteins found in sucrose gradient fractions 4-6(Figure 1 B) following IP with monoclonal anti-Cx32(Table 1 ). Negative controls included IP reactionsfrom KO fractions 4-6 (Supplemental Figure 1, Supporting Information ) and a no lysate (NL) mock IP reaction.Black boxes indicate gel regions excised for tryptic digest. Mobilityof precipitating IgG heavy and light chains are indicated by arrowheads.(A') Western blot of Cx32-enriched fractions 4-6 (F4-6,5 mug) and 1 muL (10%) of the corresponding IP productsfrom these same fractions analyzed in (A). Analysis confirmed presenceof bait (Cx32) in WT samples and lack of cross-reactivity in KO samples.Cx32-M indicates use of monoclonal anti-Cx32; Cx32-P indicates useof polyclonal anti-Cx32 antibody (Table 1 ).(B) IP-HTMS identified one known (Cx26) and 17 novel Cx32 interactingpartners in replicate screens of 1 mg and 5 mg input protein. Interactionnetwork generated by Osprey version 1.2.0 of proteins present in WTbut not KO IP-HTMS. Proteins identified in both the 1 mg and 5 mgscreens are represented with larger icons and thicker interactionlines than proteins identified only in the 5 mg screen. (C) Reciprocalco-IPs of liver lysates for Cx32 and Cx26. (D) Reciprocal co-IPs forCx32 and SFXN-1. Specificity was also verified in independent IPsusing an isotype IgG control

- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
- Figure 2. Immunostainings and fluorescence-activated cell sorting by GAP11. (A): Immunofluorescent staining with Cx32 and GAP11 antibodies (green) on frozen sections of human cornea and limbus with mouse liver as Cx32 positive control and PI counterstaining (red). (B): Flow cytometry and fluorescence-activated cell sorting of primary cultured human limbal epithelial cells by GAP11 (panels 2 and 3), with second Ab only as negative control (panel 1). A, negative cells; B, positive cells; C, selected negative cells (Cx43 dim ) with FITC intensity less than 0.2; D, selected positive cells (Cx43 bright ) with FITC intensity higher than 100. (C): Real-time PCR for Cx43 expression by ALL and selected Cx43 bright and Cx43 dim populations. Abbreviations: Ab, antibody; ALL, unfractioned; Cx, connexin; FITC, fluorescein isothiocyanate; PCR, polymerase chain reaction.
