Explore
Explore Validate
Validate Learn
Learn Immunocytochemistry
ImmunocytochemistryAntibody data
- Antibody Data
- Antigen structure
- References [14]
- Comments [0]
- Validations
- Immunocytochemistry [2]
- Immunohistochemistry [1]
- Flow cytometry [5]
- Other assay [5]
Submit
Validation data
Reference
Comment
Report error
- Product number
- MA5-14112 - Provider product page

- Provider
- Invitrogen Antibodies
- Product name
- CD36 Monoclonal Antibody (185-1G2)
- Antibody type
- Monoclonal
- Antigen
- Other
- Description
- MA5-14112 targets CD36 in FACS, immunofluorescence, and immunohistochemistry (frozen) applications and shows reactivity with human and mouse samples. The MA5-14112 immunogen is stimulated human leukocytes.
- Reactivity
- Human, Mouse
- Host
- Mouse
- Isotype
- IgG
- Antibody clone number
- 185-1G2
- Vial size
- 500 μL
- Concentration
- 0.2 mg/mL
- Storage
- 4°C
Submitted references Advanced glycation end-products reduce lipopolysaccharide uptake by macrophages.
Empagliflozin Ameliorates Free Fatty Acid Induced-Lipotoxicity in Renal Proximal Tubular Cells via the PPARγ/CD36 Pathway in Obese Mice.
Expanded IL-22(+) Group 3 Innate Lymphoid Cells and Role of Oxidized LDL-C in the Pathogenesis of Axial Spondyloarthritis with Dyslipidaemia.
SR-B1 drives endothelial cell LDL transcytosis via DOCK4 to promote atherosclerosis.
Effects of scavenger receptors-1 class A stimulation on macrophage morphology and highly modified advanced glycation end product-protein phagocytosis.
EphrinA1-Fc attenuates myocardial ischemia/reperfusion injury in mice.
Route of antigen delivery impacts the immunostimulatory activity of dendritic cell-based vaccines for hepatocellular carcinoma.
Histopathologies, immunolocalization, and a glycan binding screen provide insights into Plasmodium falciparum interactions with the human placenta.
Implication of scavenger receptors in the interactions between diesel exhaust particles and immature or mature dendritic cells.
The cysteine-rich interdomain region from the highly variable plasmodium falciparum erythrocyte membrane protein-1 exhibits a conserved structure.
CD47 is necessary for inhibition of nitric oxide-stimulated vascular cell responses by thrombospondin-1.
Differential expression of a gene signature for scavenger/lectin receptors by endothelial cells and macrophages in human lymph node sinuses, the primary sites of regional metastasis.
Inhibition of endothelial cell migration by thrombospondin-1 type-1 repeats is mediated by beta1 integrins.
Phosphatidylserine regulates the maturation of human dendritic cells.
Kitaura A, Nishinaka T, Hamasaki S, Hatipoglu OF, Wake H, Nishibori M, Mori S, Nakao S, Takahashi H
PloS one 2021;16(1):e0245957
PloS one 2021;16(1):e0245957
Empagliflozin Ameliorates Free Fatty Acid Induced-Lipotoxicity in Renal Proximal Tubular Cells via the PPARγ/CD36 Pathway in Obese Mice.
Huang CC, Chou CA, Chen WY, Yang JL, Lee WC, Chen JB, Lee CT, Li LC
International journal of molecular sciences 2021 Nov 17;22(22)
International journal of molecular sciences 2021 Nov 17;22(22)
Expanded IL-22(+) Group 3 Innate Lymphoid Cells and Role of Oxidized LDL-C in the Pathogenesis of Axial Spondyloarthritis with Dyslipidaemia.
Min HK, Moon J, Lee SY, Lee AR, Lee CR, Lee J, Kwok SK, Cho ML, Park SH
Immune network 2021 Dec;21(6):e43
Immune network 2021 Dec;21(6):e43
SR-B1 drives endothelial cell LDL transcytosis via DOCK4 to promote atherosclerosis.
Huang L, Chambliss KL, Gao X, Yuhanna IS, Behling-Kelly E, Bergaya S, Ahmed M, Michaely P, Luby-Phelps K, Darehshouri A, Xu L, Fisher EA, Ge WP, Mineo C, Shaul PW
Nature 2019 May;569(7757):565-569
Nature 2019 May;569(7757):565-569
Effects of scavenger receptors-1 class A stimulation on macrophage morphology and highly modified advanced glycation end product-protein phagocytosis.
Hamasaki S, Kobori T, Yamazaki Y, Kitaura A, Niwa A, Nishinaka T, Nishibori M, Mori S, Nakao S, Takahashi H
Scientific reports 2018 Apr 12;8(1):5901
Scientific reports 2018 Apr 12;8(1):5901
EphrinA1-Fc attenuates myocardial ischemia/reperfusion injury in mice.
DuSablon A, Parks J, Whitehurst K, Estes H, Chase R, Vlahos E, Sharma U, Wert D, Virag J
PloS one 2017;12(12):e0189307
PloS one 2017;12(12):e0189307
Route of antigen delivery impacts the immunostimulatory activity of dendritic cell-based vaccines for hepatocellular carcinoma.
Pardee AD, Yano H, Weinstein AM, Ponce AA, Ethridge AD, Normolle DP, Vujanovic L, Mizejewski GJ, Watkins SC, Butterfield LH
Journal for immunotherapy of cancer 2015;3:32
Journal for immunotherapy of cancer 2015;3:32
Histopathologies, immunolocalization, and a glycan binding screen provide insights into Plasmodium falciparum interactions with the human placenta.
Hromatka BS, Ngeleza S, Adibi JJ, Niles RK, Tshefu AK, Fisher SJ
Biology of reproduction 2013 Jun;88(6):154
Biology of reproduction 2013 Jun;88(6):154
Implication of scavenger receptors in the interactions between diesel exhaust particles and immature or mature dendritic cells.
Taront S, Dieudonné A, Blanchard S, Jeannin P, Lassalle P, Delneste Y, Gosset P
Particle and fibre toxicology 2009 Mar 13;6:9
Particle and fibre toxicology 2009 Mar 13;6:9
The cysteine-rich interdomain region from the highly variable plasmodium falciparum erythrocyte membrane protein-1 exhibits a conserved structure.
Klein MM, Gittis AG, Su HP, Makobongo MO, Moore JM, Singh S, Miller LH, Garboczi DN
PLoS pathogens 2008 Sep 5;4(9):e1000147
PLoS pathogens 2008 Sep 5;4(9):e1000147
CD47 is necessary for inhibition of nitric oxide-stimulated vascular cell responses by thrombospondin-1.
Isenberg JS, Ridnour LA, Dimitry J, Frazier WA, Wink DA, Roberts DD
The Journal of biological chemistry 2006 Sep 8;281(36):26069-80
The Journal of biological chemistry 2006 Sep 8;281(36):26069-80
Differential expression of a gene signature for scavenger/lectin receptors by endothelial cells and macrophages in human lymph node sinuses, the primary sites of regional metastasis.
Martens JH, Kzhyshkowska J, Falkowski-Hansen M, Schledzewski K, Gratchev A, Mansmann U, Schmuttermaier C, Dippel E, Koenen W, Riedel F, Sankala M, Tryggvason K, Kobzik L, Moldenhauer G, Arnold B, Goerdt S
The Journal of pathology 2006 Mar;208(4):574-89
The Journal of pathology 2006 Mar;208(4):574-89
Inhibition of endothelial cell migration by thrombospondin-1 type-1 repeats is mediated by beta1 integrins.
Short SM, Derrien A, Narsimhan RP, Lawler J, Ingber DE, Zetter BR
The Journal of cell biology 2005 Feb 14;168(4):643-53
The Journal of cell biology 2005 Feb 14;168(4):643-53
Phosphatidylserine regulates the maturation of human dendritic cells.
Chen X, Doffek K, Sugg SL, Shilyansky J
Journal of immunology (Baltimore, Md. : 1950) 2004 Sep 1;173(5):2985-94
Journal of immunology (Baltimore, Md. : 1950) 2004 Sep 1;173(5):2985-94
No comments: Submit comment
Supportive validation
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
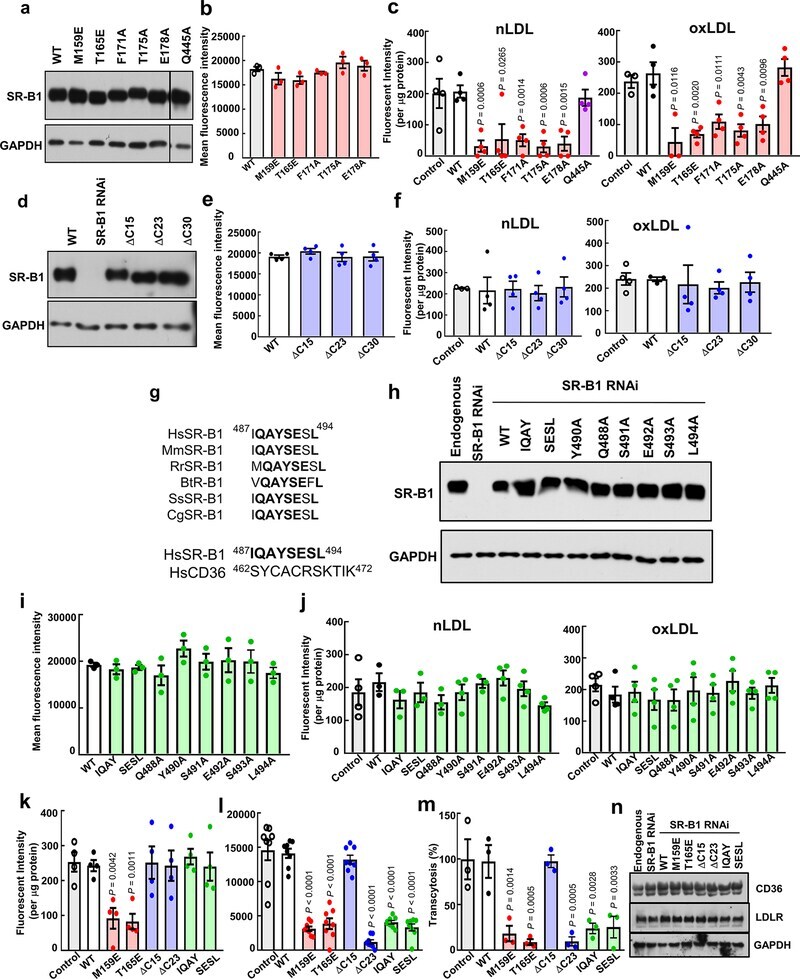
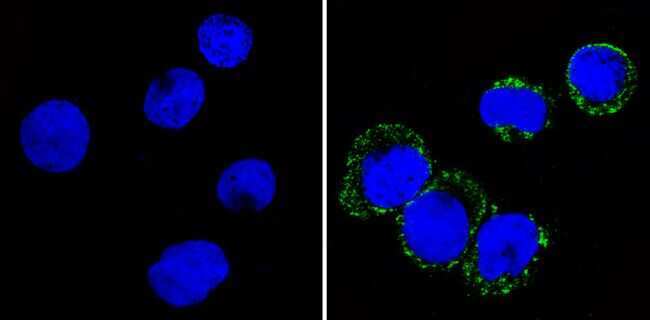
- Experimental details
- Immunofluorescent analysis of CD36 (green) showing staining in the membrane and cytoplasm of THP-1 cells (right) compared to a negative control without primary antibody (left). Formalin-fixed cells were permeabilized with 0.1% Triton X-100 in TBS for 5-10 minutes and blocked with 3% BSA-PBS for 30 minutes at room temperature. Cells were probed with a CD36 monoclonal antibody (Product # MA5-14112) in 3% BSA-PBS at a dilution of 1:20 and incubated overnight at 4ºC in a humidified chamber. Cells were washed with PBST and incubated with a DyLight-conjugated secondary antibody in PBS at room temperature in the dark. F-actin (red) was stained with a fluorescent red phalloidin and nuclei (blue) were stained with Hoechst or DAPI. Images were taken at a magnification of 60x.
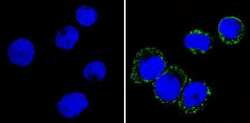
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
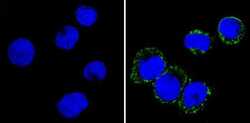
- Experimental details
- Immunofluorescent analysis of CD36 (green) showing staining in the membrane and cytoplasm of THP-1 cells (right) compared to a negative control without primary antibody (left). Formalin-fixed cells were permeabilized with 0.1% Triton X-100 in TBS for 5-10 minutes and blocked with 3% BSA-PBS for 30 minutes at room temperature. Cells were probed with a CD36 monoclonal antibody (Product # MA5-14112) in 3% BSA-PBS at a dilution of 1:20 and incubated overnight at 4ºC in a humidified chamber. Cells were washed with PBST and incubated with a DyLight-conjugated secondary antibody in PBS at room temperature in the dark. F-actin (red) was stained with a fluorescent red phalloidin and nuclei (blue) were stained with Hoechst or DAPI. Images were taken at a magnification of 60x.
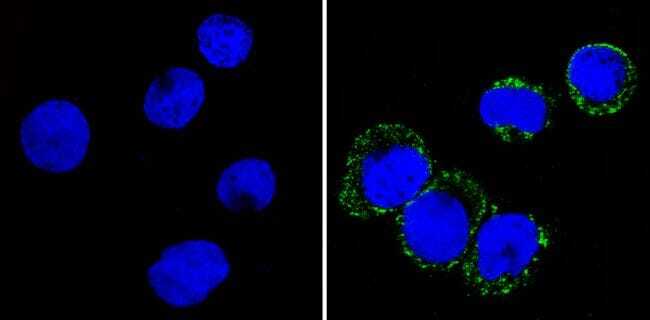
Supportive validation
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
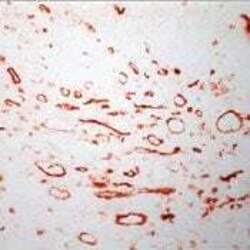
- Experimental details
- Frozen human tonsil section stained with CD36 antibody using UltraVision LP and AEC chromogen. Note staining of monocytes, marcrophages and endothelial cells.
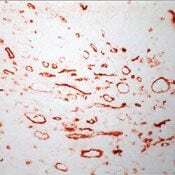
Supportive validation
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
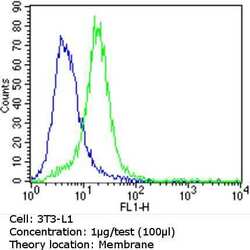
- Experimental details
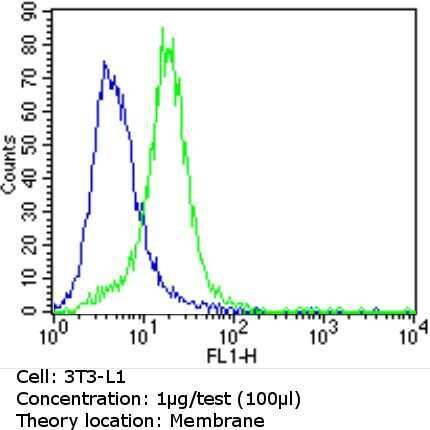
- Flow cytometry analysis of CD36 in 3T3-L1 cells (green) compared to an isotype control (blue). Cells were harvested, adjusted to a concentration of 1-5x10^6 cells/mL, fixed with 2% paraformaldehyde and washed with PBS. Cells were blocked with a 2% solution of BSA-PBS for 30 min at room temperature and incubated with a CD36 monoclonal antibody (Product # MA5-14112) at a dilution of 1 µg/test for 60 min at room temperature. Cells were then incubated for 40 min at room temperature in the dark using a Dylight 488-conjugated goat anti-mouse IgG (H+L) secondary antibody and re-suspended in PBS for FACS analysis.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
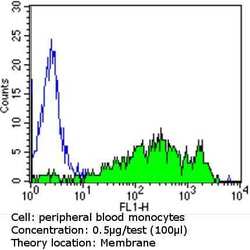
- Experimental details
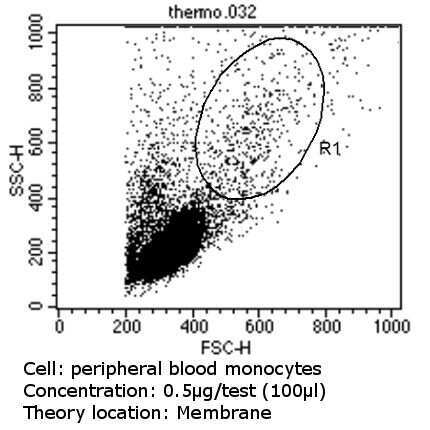
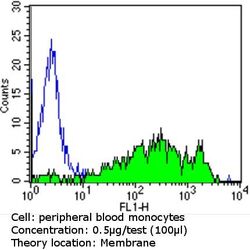
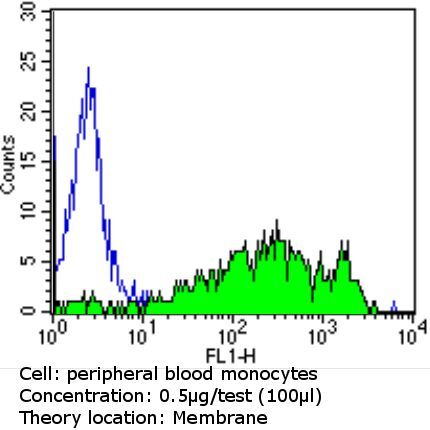
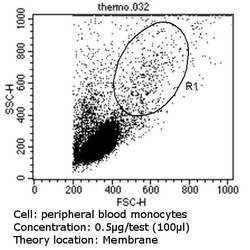
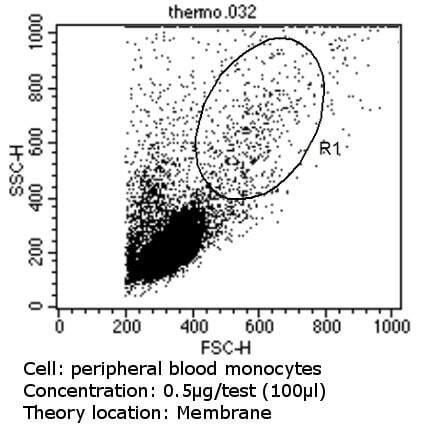
- Flow cytometry analysis of CD36 in PBMC cells (green) compared to an isotype control (blue). Human blood was collected, combined with a hydrophilic polysaccharide, centrifuged, transferred to a conical tube and washed with PBS. 50 µL of cell solution was added to each tube at a dilution of 2x10^7 cells/mL, followed by the addition of 50 µL of isotype control and primary antibody (Product # MA5-14112) at a dilution of 0.5 µg/test. Cells were incubated for 30 min at 4ºC and washed with a cell buffer, followed by incubation with a DyLight 488-conjugated goat anti-mouse IgG (H+L) secondary for 30 min at 4ºC in the dark. FACS analysis was performed using 400 µL of cell buffer.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
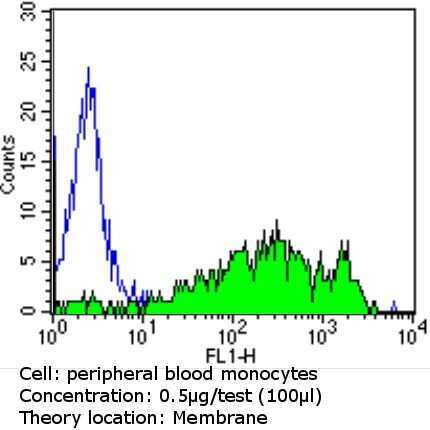
- Experimental details
- Flow cytometry analysis of CD36 in PBMC cells (green) compared to an isotype control (blue). Human blood was collected, combined with a hydrophilic polysaccharide, centrifuged, transferred to a conical tube and washed with PBS. 50 µL of cell solution was added to each tube at a dilution of 2x10^7 cells/mL, followed by the addition of 50 µL of isotype control and primary antibody (Product # MA5-14112) at a dilution of 0.5 µg/test. Cells were incubated for 30 min at 4ºC and washed with a cell buffer, followed by incubation with a DyLight 488-conjugated goat anti-mouse IgG (H+L) secondary for 30 min at 4ºC in the dark. FACS analysis was performed using 400 µL of cell buffer.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- Flow cytometry analysis of CD36 in PBMC cells (green) compared to an isotype control (blue). Human blood was collected, combined with a hydrophilic polysaccharide, centrifuged, transferred to a conical tube and washed with PBS. 50 µL of cell solution was added to each tube at a dilution of 2x10^7 cells/mL, followed by the addition of 50 µL of isotype control and primary antibody (Product # MA5-14112) at a dilution of 0.5 µg/test. Cells were incubated for 30 min at 4ºC and washed with a cell buffer, followed by incubation with a DyLight 488-conjugated goat anti-mouse IgG (H+L) secondary for 30 min at 4ºC in the dark. FACS analysis was performed using 400 µL of cell buffer.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- Flow cytometry analysis of CD36 in PBMC cells (green) compared to an isotype control (blue). Human blood was collected, combined with a hydrophilic polysaccharide, centrifuged, transferred to a conical tube and washed with PBS. 50 µL of cell solution was added to each tube at a dilution of 2x10^7 cells/mL, followed by the addition of 50 µL of isotype control and primary antibody (Product # MA5-14112) at a dilution of 0.5 µg/test. Cells were incubated for 30 min at 4ºC and washed with a cell buffer, followed by incubation with a DyLight 488-conjugated goat anti-mouse IgG (H+L) secondary for 30 min at 4ºC in the dark. FACS analysis was performed using 400 µL of cell buffer.
Supportive validation
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
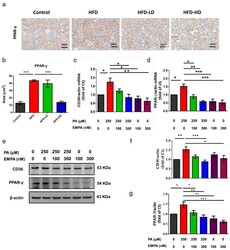
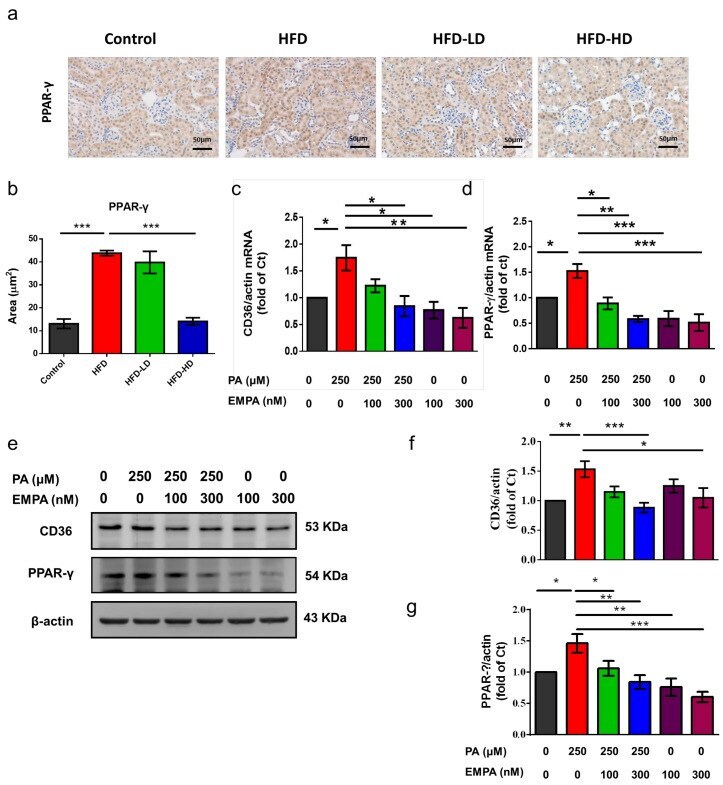
- Figure 6 Empagliflozin treatment downregulates PPAR-gamma expression in the kidney of HFD-fed mice and PA-treated HK-2 cells. ( a , b ) In the mouse kidney, high-dose empagliflozin attenuates PPAR-gamma expression, as shown by immunohistochemical staining. ( c ) HK-2 cells were cultured in medium with 1% bovine serum albumin, PA (250 muM), and low-dose (100 nM) or high-dose (300 nM) empagliflozin. High-dose empagliflozin significantly reduces the expression levels of CD36 mRNA ( c ) and protein ( e , f ). Empagliflozin also reduces PPAR-gamma mRNA ( d ) and protein expression ( e , g ). The histograms represent mean +- standard error (SE) from 6 experiments. Statistical analysis was performed with one-way ANOVA. * p < 0.05, ** p < 0.01, *** p < 0.001. EMPA, empagliflozin; PA, palmitic acid; PPAR, peroxisome proliferator-activated receptor.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
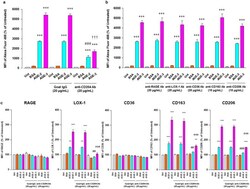
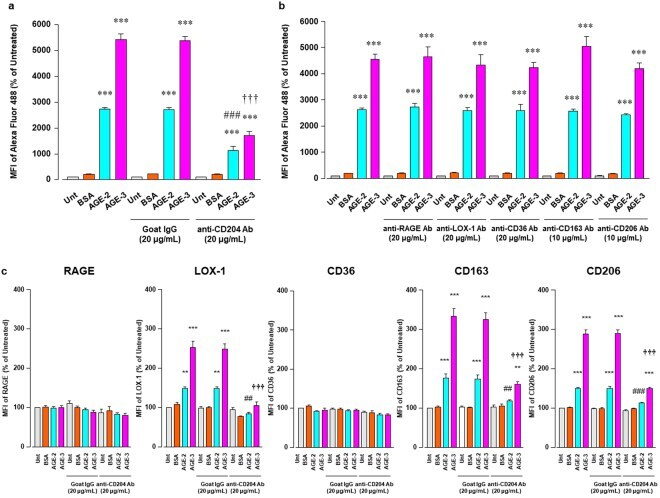
- Figure 5 Inhibitory effect of Abs against scavenger receptors for AGE uptake RAW264.7 cells seeded at 2 x 10 5 per well were pre-incubated with or without neutralizing Abs against scavenger receptors at 20 or 10 mug/ml for 1 h followed by culturing with BSA, AGE-2, or AGE-3 at 200 mug/ml for 1 h. ( a ) Neutralizing effect of anti-CD204 Ab on the uptake of fluorescent AGE-2, AGE-3, and BSA was determined by flow cytometry (n = 3). Goat IgG was used as an isotype-matched control of anti-CD204 Ab. ( b ) Neutralizing effect of each Ab against RAGE, LOX-1, CD36, CD163, or CD206 on the uptake of fluorescent BSA, AGE-2, or AGE-3 was determined by flow cytometry (n = 3-10). ( c ) Effect of anti-CD204 Ab on the expression of RAGE, LOX-1, CD36, CD163, and CD206 was determined by flow cytometry (n = 4-5). Each column represents the MFIs of the respective surface membrane proteins relative to the medium only group (Unt, Untreated) after 1 h incubation, which was arbitrarily defined as 100%. Data are presented as the means +- SEM and were analysed using one-way ANOVA followed by Tukey's test. *** p < 0.001, ** p < 0.01 compared with the value for Unt. ### p < 0.001, ## p < 0.01 compared with the value for AGE-2 alone. +++ p < 0.001 compared with the value for AGE-3 alone.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
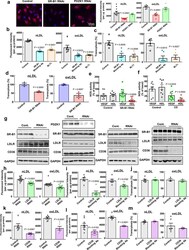
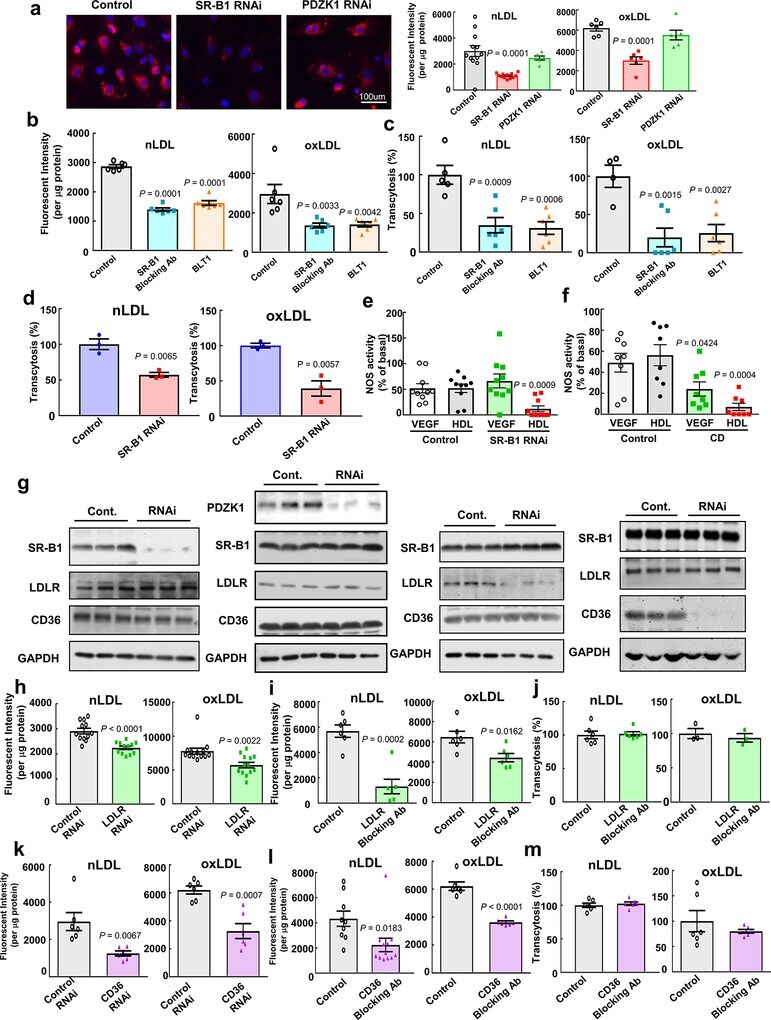
- Extended Data Figure 8. SR-B1 governs endothelial cell LDL transcytosis independent of effects on caveolae function. a , Dil-nLDL and diI-oxLDL uptake in endothelial cells after RNAi knockdown of SR-B1 or PDZK1. In left panel, diI-oxLDL visualization is shown in red, and DAPI-stained nuclei are in blue. N=6/group. b,c , Dil-nLDL and dil-oxLDL uptake ( b ) and transcytosis ( c ) in cells treated with control IgG or SR-B1 blocking antibody or BLT1. N=6/group. d , Dil-nLDL and diI-oxLDL transcytosis in endothelial cells after RNAi knockdown of SR-B1. N=3/group. e,f , NOS activation by VEGF (100 ng/mL) or HDL (20 µg/mL) with or without RNAi knockdown of SR-B1 ( e , n=10/group) or caveolae disruption by methyl-beta-cyclodextrin treatment ( f , 10mM for 60 min, n=8/group). g , Abundance of target protein following RNAi knockdown of SR-B1, PDZK1, LDLR or CD36 in HAEC. Findings for 3 samples per condition are shown. In all studies SR-B1, LDLR and CD36 expression were evaluated. h,i , DiI-nLDL and diI-oxLDL uptake in cells depleted of LDLR by RNAi ( h , n=12 for nLDL and 13 for oxLDL) or treated with control versus LDLR blocking antibody ( i , n=6/group). j , DiI-nLDL and diI-oxLDL transcytosis in cells treated with control versus LDLR blocking antibody. N=6 for nLDL and 3 for oxLDL. k,l , DiI-nLDL and diI-oxLDL uptake in cells depleted of CD36 by RNAi ( k , n=6/group) or treated with control versus CD36 blocking antibody ( l , n=9 and 12 for nLDL and 6 for oxLDL). m , DiI-nLDL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
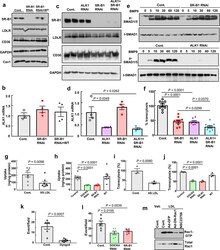
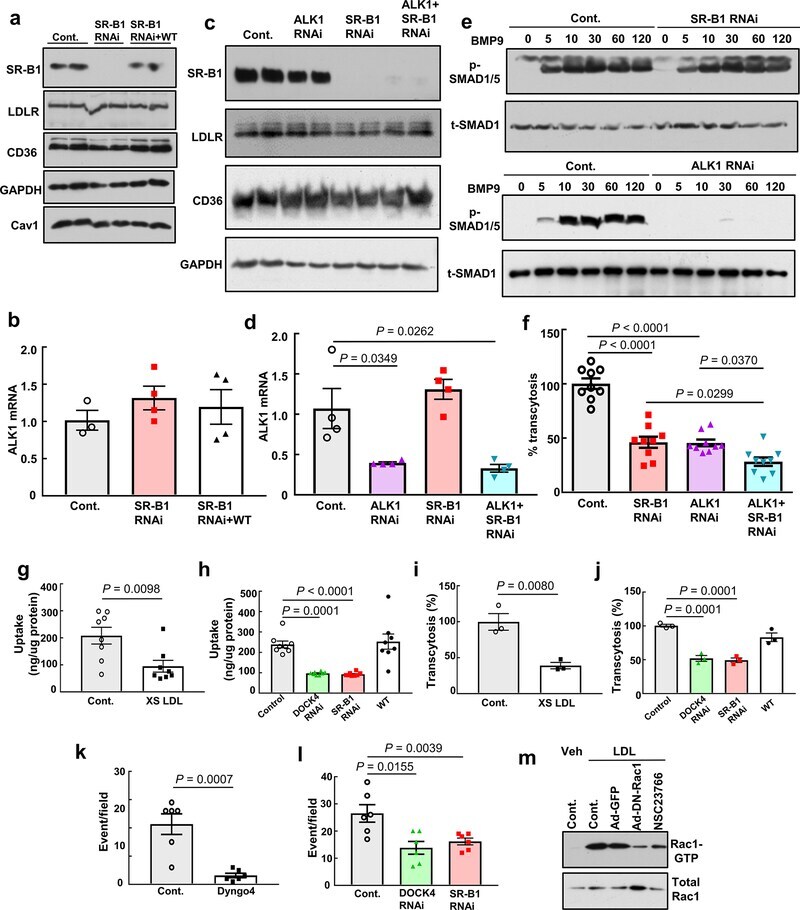
- Extended Data Figure 9. Roles of SR-B1, ALK1 and DOCK4 in endothelial cell LDL transcytosis. a , Abundance of SR-B1, LDLR, and CD36 protein following RNAi knockdown of SR-B1, or following reconstitution of wild-type SR-B1 expression in cells depleted of endogenous receptor. Caveolin-1 (Cav1) expression was also evaluated. Findings for 2 samples per condition are shown. b , ALK1 transcript levels in cells manipulated as in a. N=4/group. c , Abundance of SR-B1, LDLR, and CD36 protein following RNAi knockdown of ALK1, SR-B1, or ALK1 and SR-B1. Findings for 2 samples per condition are shown. d , ALK1 transcript levels in cells manipulated as in c. N=4/group. e , SMAD1/5 phosphorylation in response to BMP9 (10 ng/mL, for 0-120 min) following RNAi knockdown of SR-B1 or ALK1. Abundance of SMAD1/5 Ser463/465 phosphorylation and total SMAD1 were evaluated by immunoblotting. f , DiI-nLDL transcytosis following RNAi knockdown of ALK1, SR-B1, or ALK1 and SR-B1. N=9/group. g,h , nLDL uptake was evaluated using 125 I-nLDL in the absence or presence of 50-fold excess unlabeled nLDL ( g ), and h , following RNAi knockdown of SR-B1 or DOCK4, or reconstitution of wild-type SR-B1 expression in cells depleted of endogenous receptor. N=8/group. i,j , nLDL transcytosis was evaluated using 125 I-nLDL in the absence versus presence of 50-fold excess unlabeled nLDL ( i ), and j , following manipulation of SR-B1 or DOCK4 expression as in h. N=3/group. k,l , nLDL transcytosis was evaluated using total
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- Figure 10. Lentiviral reconstitution of wild-type and mutant SR-B1 expression in human endothelial cells. a-c , Studies of reconstituted wild-type SR-B1 (WT), extracellular point mutant SR-B1, or SR-B1-Q445A. The extracellular point mutants were: SR-B1-M159E, T165E, F171A, T175A, and E178A. Whole cell lysate abundance ( a ), cell surface abundance ( b ; except for Q445A, which was previously evaluated), and nLDL and oxLDL binding ( c ) were evaluated. d-f , Studies of reconstituted WT or C-terminal cytoplasmic tail deletion mutant SR-B1. The mutants were: SR-B1-DeltaC15 (Delta495-509), DeltaC23 (Delta487-509) and DeltaC30 (Delta480-509). Whole cell lysate abundance ( d ), cell surface abundance ( e ), and nLDL and oxLDL binding ( f ) were evaluated. g , Upper Panel: Sequence alignment of amino acids in the C-terminal cytoplasmic tail of SR-B1 homologs (residues 487-494) from human ( Homo sapien , Hs, Q8WTVO, Swiss-Prot), mouse ( Mus musculus , Mm, Q61009, Swiss- Prot), rat ( Rattus norvegius , Rr, P97943, Swiss-Prot), bovine ( Bos Taurus , Bt, O18824, Swiss-Prot), pig ( Sus scrofa , Ss, Q8SQC1, Swiss-Prot), and Chinese hamster ( Cricetulus griseus , Cg, Q60417, Swiss-Prot). Fully conserved residues are shown in bold. Lower Panel: Comparison of human SR-B1 residues 487-494 and entire human CD36 C-terminal cytoplasmic tail. Residues of SR-B1 not shared with CD36 are shown in bold. h-j , Studies of reconstituted WT or C-terminal cytoplasmic tail substitution mutant SR-B1. The mu