Explore
Explore Validate
Validate Learn
Learn Western blot
Western blotAntibody data
- Antibody Data
- Antigen structure
- References [27]
- Comments [0]
- Validations
- Western blot [1]
- Immunohistochemistry [1]
- Other assay [12]
Submit
Validation data
Reference
Comment
Report error
- Product number
- MA1-2014 - Provider product page

- Provider
- Invitrogen Antibodies
- Product name
- NMDAR2B Monoclonal Antibody (NR2B)
- Antibody type
- Monoclonal
- Antigen
- Recombinant full-length protein
- Description
- MA1-2014 detects the N-methyl-D-aspartate (NMDA) receptor type 2B in human and mouse samples. MA1-2014 has been used successfully in Western blot, IHC (P) and immunocytochemistry procedures. In Western blot analysis of mouse brain tissue this antibody detects a ~166 kDa protein representing NMDA receptor type 2B. The MA1-2014 immunogen is a recombinant protein composed of amino acid residues 934-1457 of the rat NMDA receptor type 2B.
- Reactivity
- Human, Mouse, Rat
- Host
- Mouse
- Isotype
- IgG
- Antibody clone number
- NR2B
- Vial size
- 100 µg
- Concentration
- 1 mg/mL
- Storage
- -20° C, Avoid Freeze/Thaw Cycles
Submitted references Transient neuroinflammation following surgery contributes to long-lasting cognitive decline in elderly rats via dysfunction of synaptic NMDA receptor.
A novel bio-inspired strategy to prevent amyloidogenesis and synaptic damage in Alzheimer's disease.
The σ1 Receptor and the HINT1 Protein Control α2δ1 Binding to Glutamate NMDA Receptors: Implications in Neuropathic Pain.
TRPV4 Regulates Soman-Induced Status Epilepticus and Secondary Brain Injury via NMDA Receptor and NLRP3 Inflammasome.
DAPK1 Promotes Extrasynaptic GluN2B Phosphorylation and Striatal Spine Instability in the YAC128 Mouse Model of Huntington Disease.
Tau knockout exacerbates degeneration of parvalbumin-positive neurons in substantia nigra pars reticulata in Parkinson's disease-related α-synuclein A53T mice.
Altered Regulation of Striatal Neuronal N-Methyl-D-Aspartate Receptor Trafficking by Palmitoylation in Huntington Disease Mouse Model.
AMPA Receptor Dysregulation and Therapeutic Interventions in a Mouse Model of CDKL5 Deficiency Disorder.
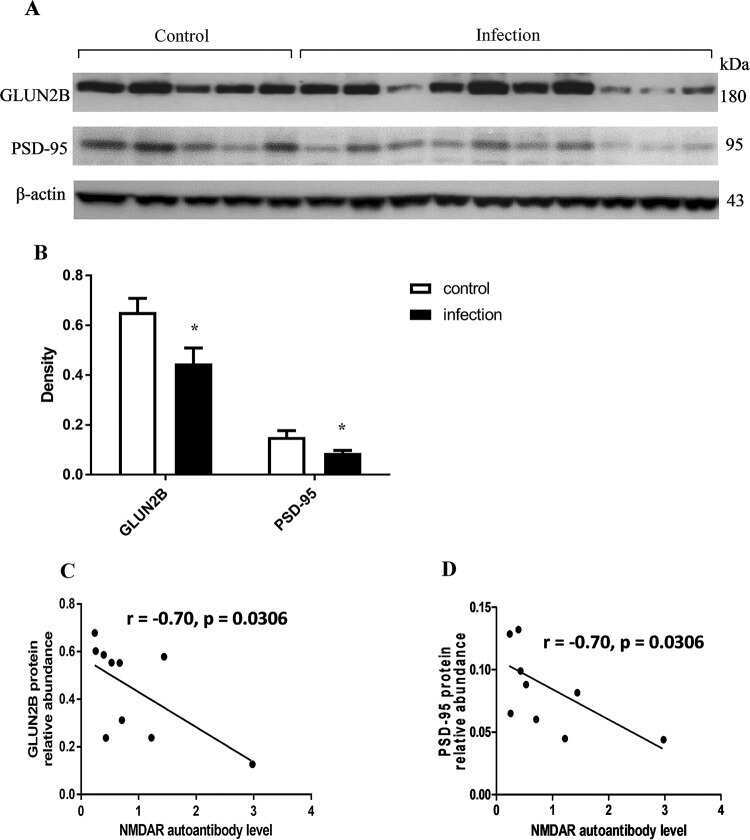
Chronic Toxoplasma gondii Infection Induces Anti-N-Methyl-d-Aspartate Receptor Autoantibodies and Associated Behavioral Changes and Neuropathology.
Disruption of Ninjurin1 Leads to Repetitive and Anxiety-Like Behaviors in Mice.
Overexpression of human NR2B receptor subunit in LMAN causes stuttering and song sequence changes in adult zebra finches.
Neurodegeneration in Amygdala Precedes Hippocampus in the APPswe/ PS1dE9 Mouse Model of Alzheimer's Disease.
Disulfide-containing high mobility group box-1 promotes N-methyl-D-aspartate receptor function and excitotoxicity by activating Toll-like receptor 4-dependent signaling in hippocampal neurons.
Environmental enrichment mitigates the sex-specific effects of gestational inflammation on social engagement and the hypothalamic pituitary adrenal axis-feedback system.
The influence of neuropathology on brain inflammation in human and experimental temporal lobe epilepsy.
Mitigation of augmented extrasynaptic NMDAR signaling and apoptosis in cortico-striatal co-cultures from Huntington's disease mice.
P38 MAPK is involved in enhanced NMDA receptor-dependent excitotoxicity in YAC transgenic mouse model of Huntington disease.
N-methyl-D-aspartate receptor subunit- and neuronal-type dependence of excitotoxic signaling through post-synaptic density 95.
TRPV1 acts as a synaptic protein and regulates vesicle recycling.
Age and meloxicam attenuate the ischemia/reperfusion-induced down-regulation in the NMDA receptor genes.
Dopamine D(1) receptor-mediated enhancement of NMDA receptor trafficking requires rapid PKC-dependent synaptic insertion in the prefrontal neurons.
Interaction of postsynaptic density protein-95 with NMDA receptors influences excitotoxicity in the yeast artificial chromosome mouse model of Huntington's disease.
Neuronal survival and resistance to HIV-1 Tat toxicity in the primary culture of rat fetal neurons.
Nonspecific interaction of prefibrillar amyloid aggregates with glutamatergic receptors results in Ca2+ increase in primary neuronal cells.
Cell-specific expression of N-methyl-D-aspartate receptor subunits in Müller glia and neurons from the chick retina.
Characterization of NMDA receptor subunit-specific antibodies: distribution of NR2A and NR2B receptor subunits in rat brain and ontogenic profile in the cerebellum.
Characterization of NMDA receptor subunit-specific antibodies: distribution of NR2A and NR2B receptor subunits in rat brain and ontogenic profile in the cerebellum.
Chen B, Qin G, Xiao J, Deng X, Lin A, Liu H
Journal of neuroinflammation 2022 Jul 13;19(1):181
Journal of neuroinflammation 2022 Jul 13;19(1):181
A novel bio-inspired strategy to prevent amyloidogenesis and synaptic damage in Alzheimer's disease.
Catania M, Colombo L, Sorrentino S, Cagnotto A, Lucchetti J, Barbagallo MC, Vannetiello I, Vecchi ER, Favagrossa M, Costanza M, Giaccone G, Salmona M, Tagliavini F, Di Fede G
Molecular psychiatry 2022 Dec;27(12):5227-5234
Molecular psychiatry 2022 Dec;27(12):5227-5234
The σ1 Receptor and the HINT1 Protein Control α2δ1 Binding to Glutamate NMDA Receptors: Implications in Neuropathic Pain.
Rodríguez-Muñoz M, Cortés-Montero E, Onetti Y, Sánchez-Blázquez P, Garzón-Niño J
Biomolecules 2021 Nov 12;11(11)
Biomolecules 2021 Nov 12;11(11)
TRPV4 Regulates Soman-Induced Status Epilepticus and Secondary Brain Injury via NMDA Receptor and NLRP3 Inflammasome.
Wang S, He H, Long J, Sui X, Yang J, Lin G, Wang Q, Wang Y, Luo Y
Neuroscience bulletin 2021 Jul;37(7):905-920
Neuroscience bulletin 2021 Jul;37(7):905-920
DAPK1 Promotes Extrasynaptic GluN2B Phosphorylation and Striatal Spine Instability in the YAC128 Mouse Model of Huntington Disease.
Schmidt ME, Caron NS, Aly AE, Lemarié FL, Dal Cengio L, Ko Y, Lazic N, Anderson L, Nguyen B, Raymond LA, Hayden MR
Frontiers in cellular neuroscience 2020;14:590569
Frontiers in cellular neuroscience 2020;14:590569
Tau knockout exacerbates degeneration of parvalbumin-positive neurons in substantia nigra pars reticulata in Parkinson's disease-related α-synuclein A53T mice.
Jiao L, Zheng M, Duan J, Wu T, Li Z, Liu L, Xiang X, Tang X, He J, Li X, Zhang G, Ding J, Cai H, Lin X
FASEB journal : official publication of the Federation of American Societies for Experimental Biology 2020 Sep;34(9):12239-12254
FASEB journal : official publication of the Federation of American Societies for Experimental Biology 2020 Sep;34(9):12239-12254
Altered Regulation of Striatal Neuronal N-Methyl-D-Aspartate Receptor Trafficking by Palmitoylation in Huntington Disease Mouse Model.
Kang R, Wang L, Sanders SS, Zuo K, Hayden MR, Raymond LA
Frontiers in synaptic neuroscience 2019;11:3
Frontiers in synaptic neuroscience 2019;11:3
AMPA Receptor Dysregulation and Therapeutic Interventions in a Mouse Model of CDKL5 Deficiency Disorder.
Yennawar M, White RS, Jensen FE
The Journal of neuroscience : the official journal of the Society for Neuroscience 2019 Jun 12;39(24):4814-4828
The Journal of neuroscience : the official journal of the Society for Neuroscience 2019 Jun 12;39(24):4814-4828
Chronic Toxoplasma gondii Infection Induces Anti-N-Methyl-d-Aspartate Receptor Autoantibodies and Associated Behavioral Changes and Neuropathology.
Li Y, Viscidi RP, Kannan G, McFarland R, Pletnikov MV, Severance EG, Yolken RH, Xiao J
Infection and immunity 2018 Oct;86(10)
Infection and immunity 2018 Oct;86(10)
Disruption of Ninjurin1 Leads to Repetitive and Anxiety-Like Behaviors in Mice.
Le H, Ahn BJ, Lee HS, Shin A, Chae S, Lee SY, Shin MW, Lee EJ, Cha JH, Son T, Seo JH, Wee HJ, Lee HJ, Jang Y, Lo EH, Jeon S, Oh GT, Kim D, Kim KW
Molecular neurobiology 2017 Nov;54(9):7353-7368
Molecular neurobiology 2017 Nov;54(9):7353-7368
Overexpression of human NR2B receptor subunit in LMAN causes stuttering and song sequence changes in adult zebra finches.
Chakraborty M, Chen LF, Fridel EE, Klein ME, Senft RA, Sarkar A, Jarvis ED
Scientific reports 2017 Apr 21;7(1):942
Scientific reports 2017 Apr 21;7(1):942
Neurodegeneration in Amygdala Precedes Hippocampus in the APPswe/ PS1dE9 Mouse Model of Alzheimer's Disease.
Lin TW, Liu YF, Shih YH, Chen SJ, Huang TY, Chang CY, Lien CH, Yu L, Chen SH, Kuo YM
Current Alzheimer research 2015;12(10):951-63
Current Alzheimer research 2015;12(10):951-63
Disulfide-containing high mobility group box-1 promotes N-methyl-D-aspartate receptor function and excitotoxicity by activating Toll-like receptor 4-dependent signaling in hippocampal neurons.
Balosso S, Liu J, Bianchi ME, Vezzani A
Antioxidants & redox signaling 2014 Oct 20;21(12):1726-40
Antioxidants & redox signaling 2014 Oct 20;21(12):1726-40
Environmental enrichment mitigates the sex-specific effects of gestational inflammation on social engagement and the hypothalamic pituitary adrenal axis-feedback system.
Connors EJ, Shaik AN, Migliore MM, Kentner AC
Brain, behavior, and immunity 2014 Nov;42:178-90
Brain, behavior, and immunity 2014 Nov;42:178-90
The influence of neuropathology on brain inflammation in human and experimental temporal lobe epilepsy.
Aalbers MW, Rijkers K, Majoie HJ, Dings JT, Schijns OE, Schipper S, De Baets MH, Kessels A, Vles JS, Hoogland G
Journal of neuroimmunology 2014 Jun 15;271(1-2):36-42
Journal of neuroimmunology 2014 Jun 15;271(1-2):36-42
Mitigation of augmented extrasynaptic NMDAR signaling and apoptosis in cortico-striatal co-cultures from Huntington's disease mice.
Milnerwood AJ, Kaufman AM, Sepers MD, Gladding CM, Zhang L, Wang L, Fan J, Coquinco A, Qiao JY, Lee H, Wang YT, Cynader M, Raymond LA
Neurobiology of disease 2012 Oct;48(1):40-51
Neurobiology of disease 2012 Oct;48(1):40-51
P38 MAPK is involved in enhanced NMDA receptor-dependent excitotoxicity in YAC transgenic mouse model of Huntington disease.
Fan J, Gladding CM, Wang L, Zhang LY, Kaufman AM, Milnerwood AJ, Raymond LA
Neurobiology of disease 2012 Mar;45(3):999-1009
Neurobiology of disease 2012 Mar;45(3):999-1009
N-methyl-D-aspartate receptor subunit- and neuronal-type dependence of excitotoxic signaling through post-synaptic density 95.
Fan J, Vasuta OC, Zhang LY, Wang L, George A, Raymond LA
Journal of neurochemistry 2010 Nov;115(4):1045-56
Journal of neurochemistry 2010 Nov;115(4):1045-56
TRPV1 acts as a synaptic protein and regulates vesicle recycling.
Goswami C, Rademacher N, Smalla KH, Kalscheuer V, Ropers HH, Gundelfinger ED, Hucho T
Journal of cell science 2010 Jun 15;123(Pt 12):2045-57
Journal of cell science 2010 Jun 15;123(Pt 12):2045-57
Age and meloxicam attenuate the ischemia/reperfusion-induced down-regulation in the NMDA receptor genes.
Montori S, Dos-Anjos S, Martínez-Villayandre B, Regueiro-Purriños MM, Gonzalo-Orden JM, Ruano D, Fernández-López A
Neurochemistry international 2010 Jul;56(8):878-85
Neurochemistry international 2010 Jul;56(8):878-85
Dopamine D(1) receptor-mediated enhancement of NMDA receptor trafficking requires rapid PKC-dependent synaptic insertion in the prefrontal neurons.
Li YC, Liu G, Hu JL, Gao WJ, Huang YQ
Journal of neurochemistry 2010 Jul;114(1):62-73
Journal of neurochemistry 2010 Jul;114(1):62-73
Interaction of postsynaptic density protein-95 with NMDA receptors influences excitotoxicity in the yeast artificial chromosome mouse model of Huntington's disease.
Fan J, Cowan CM, Zhang LY, Hayden MR, Raymond LA
The Journal of neuroscience : the official journal of the Society for Neuroscience 2009 Sep 2;29(35):10928-38
The Journal of neuroscience : the official journal of the Society for Neuroscience 2009 Sep 2;29(35):10928-38
Neuronal survival and resistance to HIV-1 Tat toxicity in the primary culture of rat fetal neurons.
Aksenova MV, Aksenov MY, Adams SM, Mactutus CF, Booze RM
Experimental neurology 2009 Feb;215(2):253-63
Experimental neurology 2009 Feb;215(2):253-63
Nonspecific interaction of prefibrillar amyloid aggregates with glutamatergic receptors results in Ca2+ increase in primary neuronal cells.
Pellistri F, Bucciantini M, Relini A, Nosi D, Gliozzi A, Robello M, Stefani M
The Journal of biological chemistry 2008 Oct 31;283(44):29950-60
The Journal of biological chemistry 2008 Oct 31;283(44):29950-60
Cell-specific expression of N-methyl-D-aspartate receptor subunits in Müller glia and neurons from the chick retina.
Lamas M, Lee-Rivera I, López-Colomé AM
Investigative ophthalmology & visual science 2005 Oct;46(10):3570-7
Investigative ophthalmology & visual science 2005 Oct;46(10):3570-7
Characterization of NMDA receptor subunit-specific antibodies: distribution of NR2A and NR2B receptor subunits in rat brain and ontogenic profile in the cerebellum.
Wang YH, Bosy TZ, Yasuda RP, Grayson DR, Vicini S, Pizzorusso T, Wolfe BB
Journal of neurochemistry 1995 Jul;65(1):176-83
Journal of neurochemistry 1995 Jul;65(1):176-83
Characterization of NMDA receptor subunit-specific antibodies: distribution of NR2A and NR2B receptor subunits in rat brain and ontogenic profile in the cerebellum.
Wang YH, Bosy TZ, Yasuda RP, Grayson DR, Vicini S, Pizzorusso T, Wolfe BB
Journal of neurochemistry 1995 Jul;65(1):176-83
Journal of neurochemistry 1995 Jul;65(1):176-83
No comments: Submit comment
Supportive validation
- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
- Western blot detection of NMDA receptor type 2B in mouse brain tissue using Product # MA1-2014.

Supportive validation
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
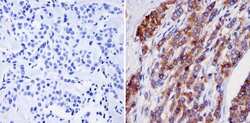
- Experimental details
- Immunohistochemistry analysis of NMDA Receptor 2B showing staining in the cytoplasm and membrane of paraffin-embedded human breast tissue (right) compared with a negative control without primary antibody (left). To expose target proteins, antigen retrieval was performed using 10mM sodium citrate (pH 6.0), microwaved for 8-15 min. Following antigen retrieval, tissues were blocked in 3% H2O2-methanol for 15 min at room temperature, washed with ddH2O and PBS, and then probed with a NMDA Receptor 2B monoclonal antibody (Product # MA1-2014) diluted in 3% BSA-PBS at a dilution of 1:20 overnight at 4°C in a humidified chamber. Tissues were washed extensively in PBST and detection was performed using an HRP-conjugated secondary antibody followed by colorimetric detection using a DAB kit. Tissues were counterstained with hematoxylin and dehydrated with ethanol and xylene to prep for mounting.
Supportive validation
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
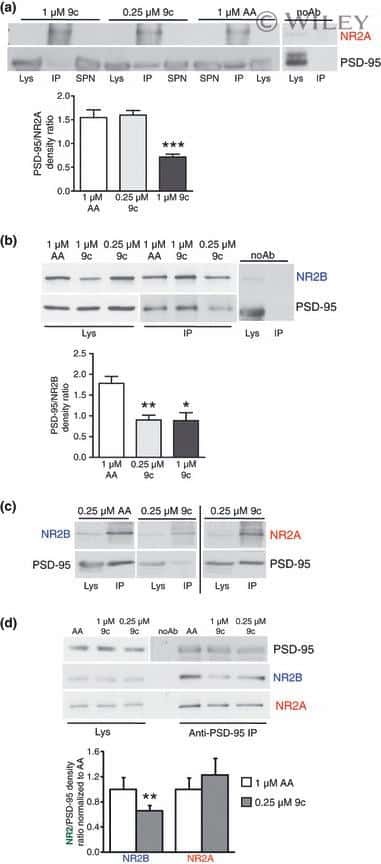
- Figure 2 Design of lentiviral vectors for NR2B manipulation in LMAN. ( a ) Diagram of the eGFPhNR2B vector to overexpress human NR2B . ( b ) Diagram of the eGFP vector for control injections. ( c ) Western blots showing no human NR2B protein in HEK293FT cells without transfection (Lane 1), and greater NR2B protein (* ~180 kD band) in cells transfected with the eGFPhNR2B plasmid (possible degraded or unfinished NR2B protein product is detected as an additional lower band) (Lane 2). ( d ) HEK293FT cells transfected with the eGFP plasmid show GFP-expressing cells labeled using a GFP antibody (green). ( e ) Cells transfected with the eGFP plasmid stained with an anti-NR2B antibody show no labeled cells. ( f ) Merged image of ( d ) and ( e ). ( g ) Cells transfected with the eGFPhNR2B plasmid show GFP-expressing cells (localized in cell membranes) labeled using a GFP antibody (green). ( h ) Cells transfected with the eGFPhNR2B plasmid show NR2B-expressing cells (localized in cell membranes) stained with anti-NR2B antibody (red). ( i ) Merged image of ( g ) and ( h ) showing complete overlap of the human NR2B tagged with eGFP (yellow). ( j ) Darkfield view of a brain section showing the injection site in LMAN. ( k ) Cells expressing GFP-tagged (green) human NR2B ~3 weeks after injection of the eGFPhNR2B lentivirus in LMAN. ( l ) Darkfield photograph of coronal sections showing drawings of LMAN and part of nidopallium that was microdissected for protein analysis. ( m ) Western blots
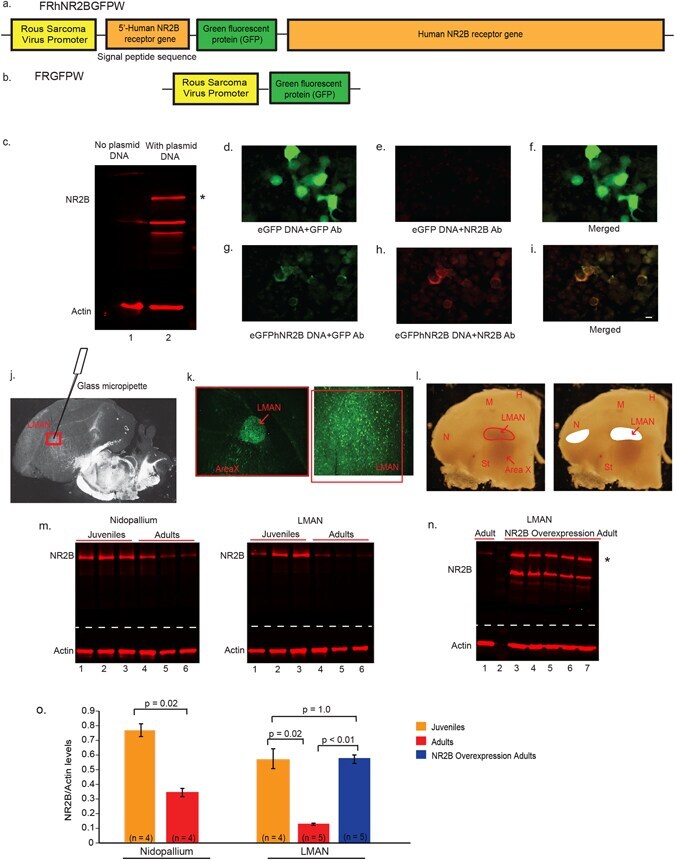
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
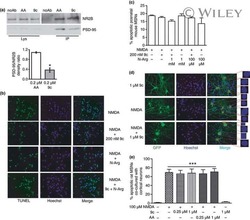
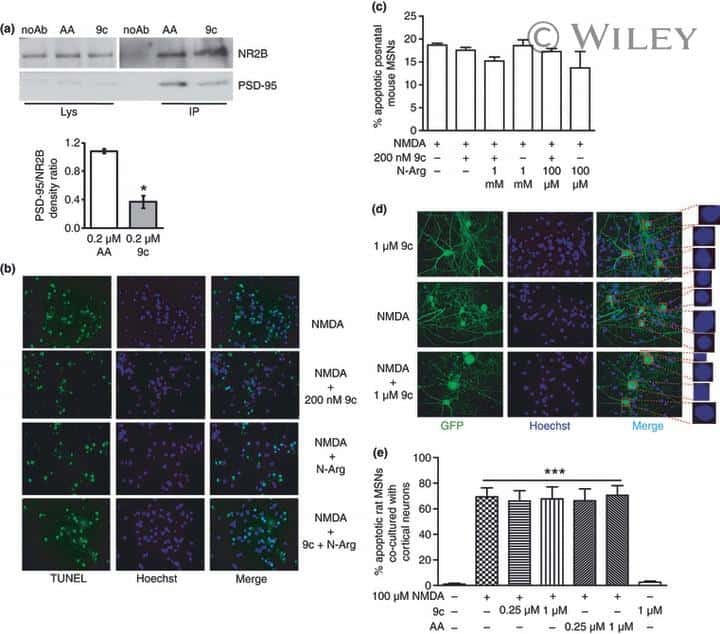
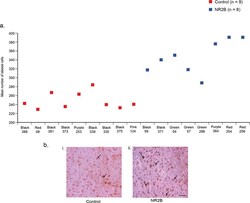
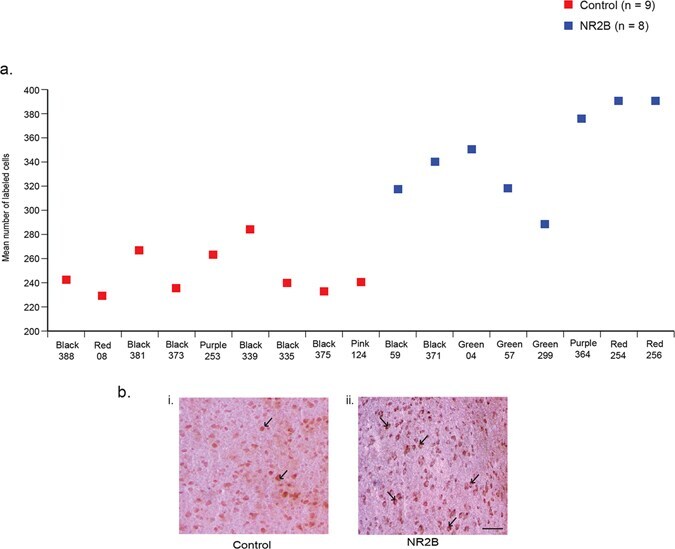
- Figure 3 Immunostaining of NR2B protein in LMAN cells. ( a ) Quantification showing higher levels of labeled cells in birds injected with the eGFPhNR2B lentivirus in LMAN (blue) compared to control birds with injections of the eGFPhNR2B lentivirus in nidopallium or the eGFP lentivirus in LMAN (red). Control-injected animals did not differ in number of labeled cells (p = 0.73, two sample Wilcoxon test). ( b ) Examples of labeled LMAN cells in eGFP-injected animals (i), and in eGFPhNR2B-injected animals (ii). Scale bar, 200 mum.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
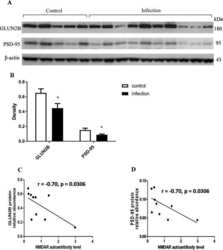
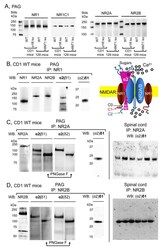
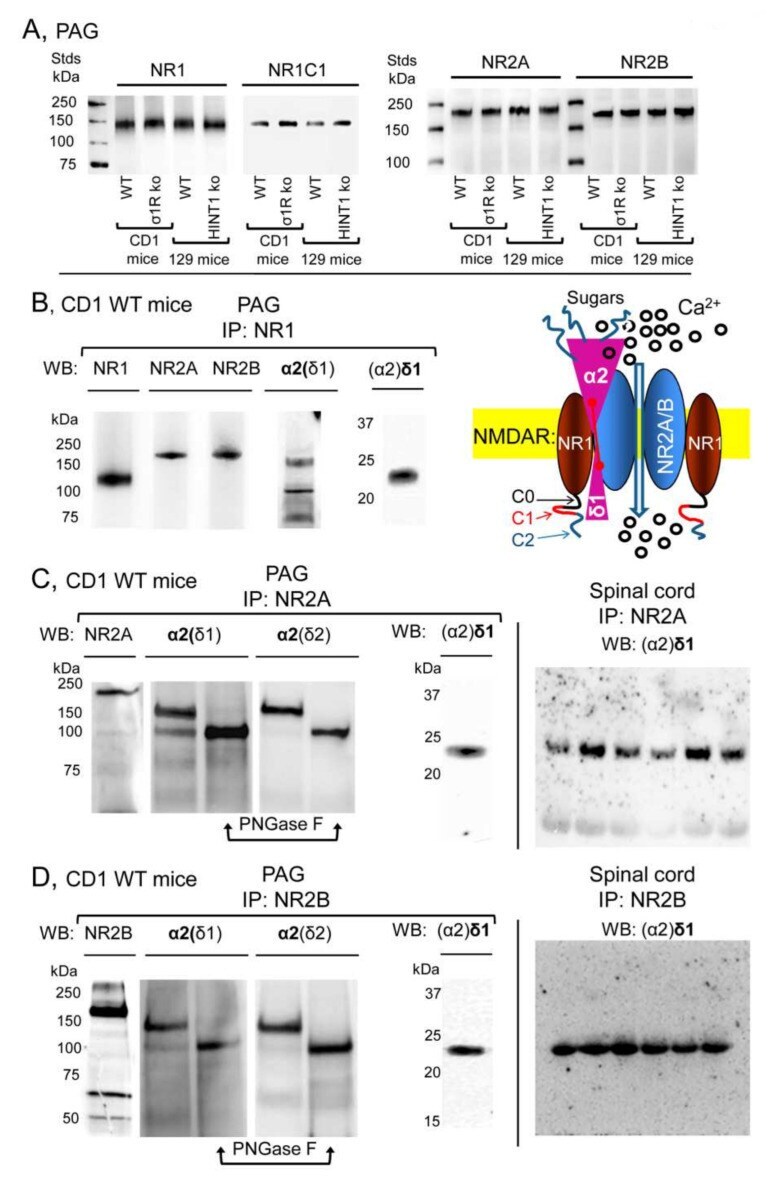
- Figure 2 Expression of NR1, NR2A, and NR2B subunits of glutamate NMDARs in mouse PAG: Co-precipitation with alpha2(delta1), alpha2(delta2), and (alpha2)delta1 proteins. ( A ) PAG P2 fractions enriched in synaptosomes from the CD1 WT, CD1 sigma1R -/- , 129 WT, and 129 HINT1 -/- mice of Figure 1 B,C, were used. About 60 mug protein/lane was resolved by SDS-PAGE and examined in Western blots that were probed with antibodies against NR1, NR1 C1, NR2A, and NR2B NMDAR subunits, as described in Methods. The assay was repeated twice with comparable outcomes. Immunoprecipitation assays on CD1 WT mice ( n = 12): PAG and SC membranes from CD1 WT mice were solubilized with 1% NP-40 and incubated overnight at 4 degC with affinity-purified biotinylated IgGs raised against the ( B ) NR1, ( C ) NR2A, or ( D ) NR2B subunits. Protein complexes were immunoprecipitated (IP) with streptavidin agarose, resolved by SDS-PAGE and visualized in Western blots. The expected size of the alpha2(delta1) and alpha2(delta2) is approximately 100 kDa; however, the bands detected usually appeared as a doublet of 100-140 kDa. ( C , D ), The material associated with the NR2A or NR2B subunits was subjected to deglycosylation with PNGase F, which depleted the alpha2(delta) 140 kDa band in favor of a 100 kDa band. In SC synaptosomes, (alpha2)delta1-2 immunosignals were enriched at NR2A/B subunits (each lane was loaded with solubilized spinal cord tissue from a single CD1 WT mouse, for more details see Section 2 ). I
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
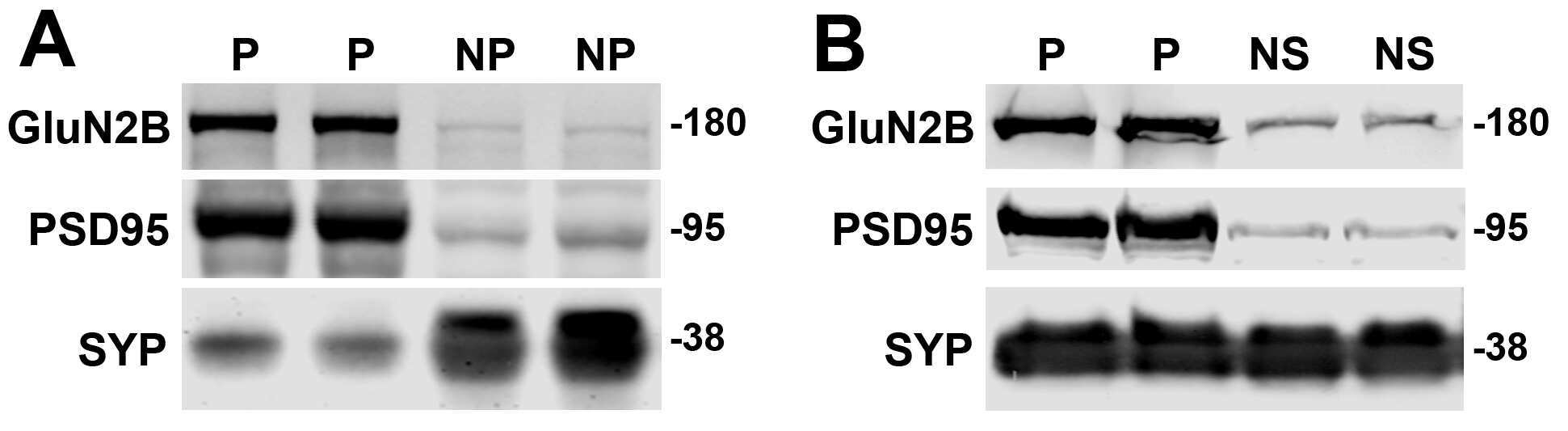
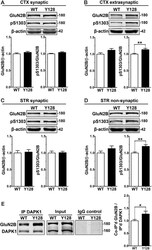
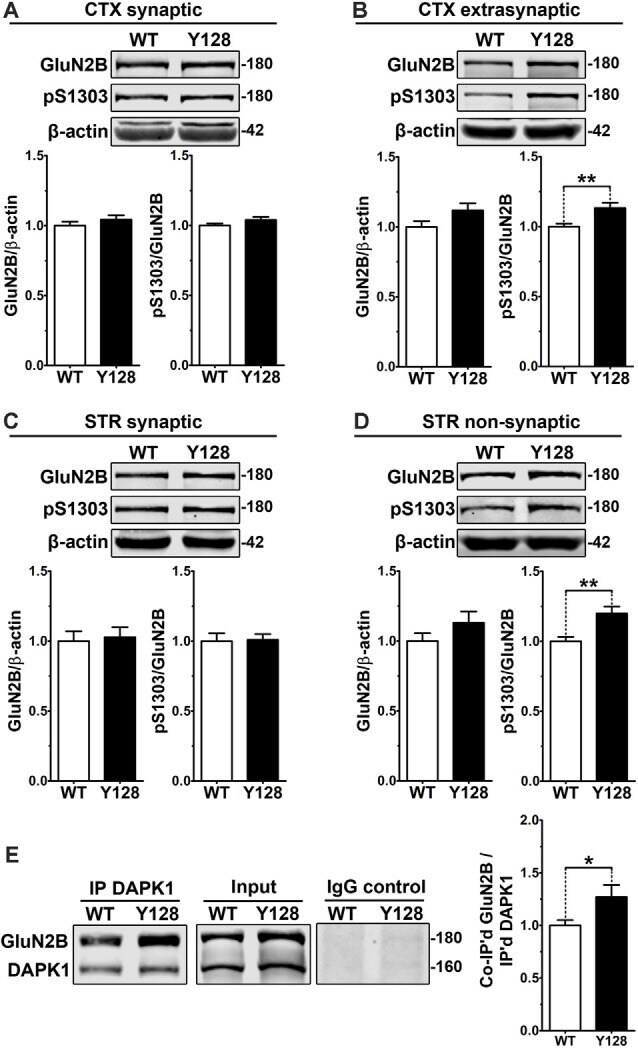
- Figure 2 Extrasynaptic GluN2B S1303 phosphorylation and interaction with DAPK1 are elevated in the YAC128 brain. Cortical or striatal tissues were subjected to subcellular fractionation to yield (A,C) synaptic (PSD), (B) cortical extrasynaptic (non-PSD), or (D) striatal ""non-synaptic"" membrane fractions, which were run by SDS-PAGE and Western blotting for GluN2B expression and S1303 phosphorylation ( n = 6-14 biological replicates, two technical replicates each; Student's t -test, * p < 0.05, ** p < 0.01). (E) DAPK1 was immunoprecipitated from WT and YAC128 cortical lysates, followed by the detection of co-immunoprecipitated GluN2B by Western blot. All sample blot images in panel (E) are cropped from the same Western blot membrane which contained multiple biological replicates run side-by-side. Data are normalized to WT values ( n = 14 biological replicates, Student's t -test, * p < 0.05, ** p < 0.01).
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
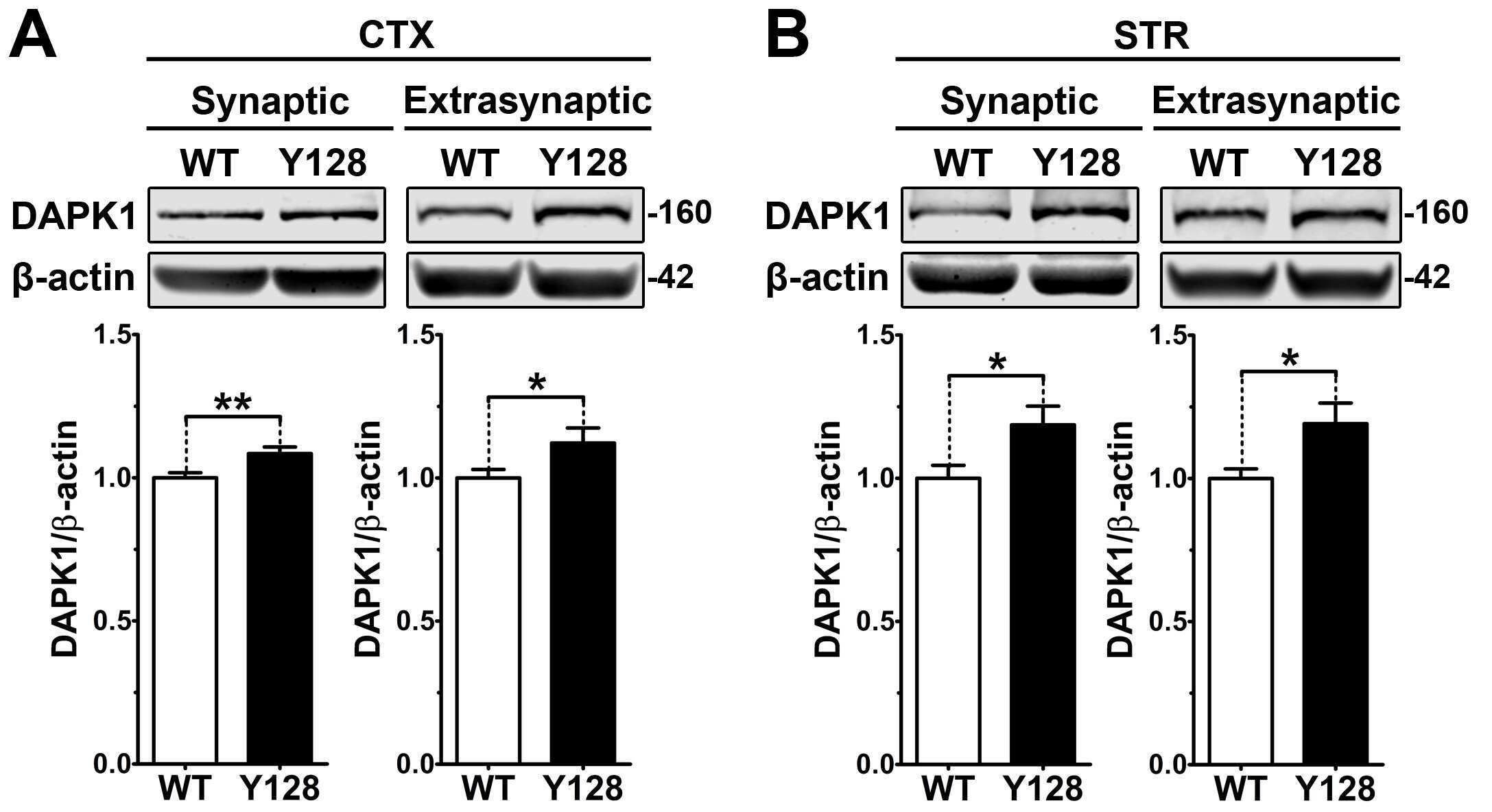
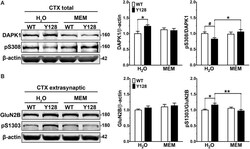
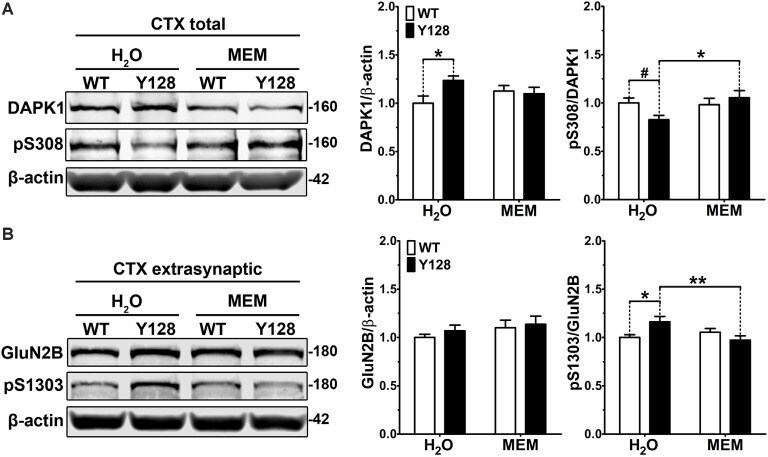
- Figure 3 Low-dose memantine normalizes cortical DAPK1 activation and extrasynaptic pS1303 levels in YAC128 mice. Cortical tissues from 1-month-old WT and YAC128 mice treated with low-dose memantine from conception were processed to obtain (A) total or (B) extrasynaptic (non-PSD) membrane protein fractions. Samples were processed for DAPK1 or GluN2B protein expression and phosphorylation levels by Western blot. Data are normalized to WT H 2 O values ( n = 8 biological replicates, two technical replicates each; two-way ANOVA with Bonferroni post hoc analysis, * p < 0.05, ** p < 0.01; Student's t -test, # p < 0.05; two-way ANOVA interaction * p < 0.05 for DAPK1 protein level and pS308, two-way ANOVA interaction ** p < 0.01 for pS1303).
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
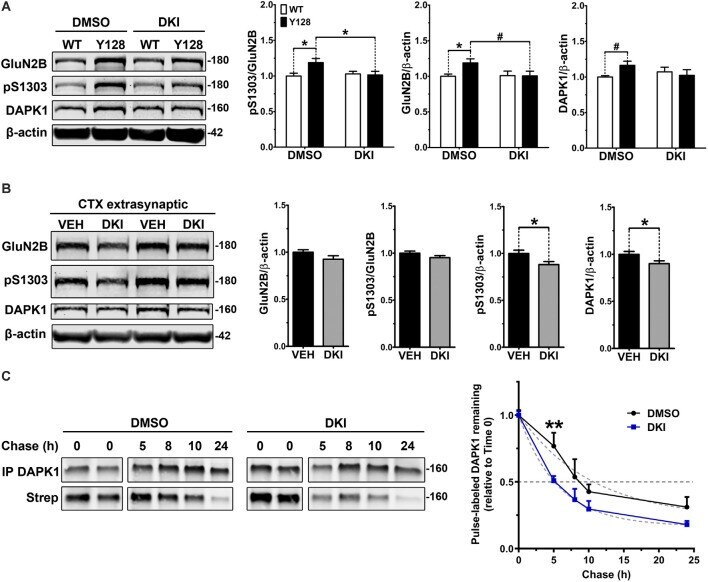
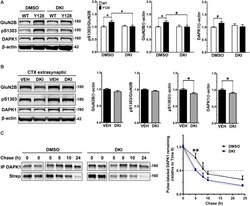
- Figure 4 DAPK1 inhibition normalizes extrasynaptic GluN2B phosphorylation and DAPK1 protein levels in the YAC128 model. (A) Corticostriatal primary cultures from WT and YAC128 embryos were treated at DIV21 with DMSO (0.1%) or a DAPK1 inhibitor (DKI, 10 muM) for 1 h followed by lysis in a gentle buffer to solubilize non-synaptic protein. Lysates were processed by Western blot to detect GluN2B, pS1303, and DAPK1 levels. Data are normalized to WT DMSO values ( n = 8 biological replicates; two-way ANOVA with Bonferroni post hoc analysis, * p < 0.05; Student's t -test, # p < 0.05). (B) Four-week-old YAC128 mice were treated intranasally with 50 nmol of DAPK1 inhibitor (DKI; TC-DAPK6) or vehicle control (VEH). After 6 h, cortical extrasynaptic fractions were isolated to measure GluN2B, pS1303, and DAPK1 levels ( n = 9 biological replicates, two technical replicates each; Student's t -test, * p < 0.05). (C) Quantitative measurement of DAPK1-FLAG protein turnover using bio-orthogonal labeling, FLAG immunoprecipitation, CLICK chemistry, and detection of AHA/Streptavidin (Strep)-labeled DAPK1 in transiently transfected COS-7 cells treated with DMSO or 10 muM DKI (TC-DAPK6) for 24 h ( n = 3 for 8 h and 10 h chase time points, n = 7 for all other chase time points; two-way ANOVA ** p < 0.01 for DKI treatment effect; Sidak's multiple comparisons test, ** p < 0.01).