Explore
Explore Validate
Validate Learn
Learn Immunohistochemistry
ImmunohistochemistryAntibody data
- Antibody Data
- Antigen structure
- References [34]
- Comments [0]
- Validations
- Immunohistochemistry [1]
- Other assay [44]
Submit
Validation data
Reference
Comment
Report error
- Product number
- 37-4600 - Provider product page

- Provider
- Invitrogen Antibodies
- Product name
- Connexin 36 Monoclonal Antibody (1E5H5)
- Antibody type
- Monoclonal
- Antigen
- Synthetic peptide
- Reactivity
- Human, Mouse
- Host
- Mouse
- Isotype
- IgG
- Antibody clone number
- 1E5H5
- Vial size
- 100 μg
- Concentration
- 0.5 mg/mL
- Storage
- -20°C
Submitted references PTEN Expression Regulates Gap Junction Connectivity in the Retina.
Synaptic protein interaction networks encode experience by assuming stimulus-specific and brain-region-specific states.
Amacrine Cells Forming Gap Junctions With Intrinsically Photosensitive Retinal Ganglion Cells: ipRGC Types, Neuromodulator Contents, and Connexin Isoform.
Phosphorylation of Connexin36 near the C-terminus switches binding affinities for PDZ-domain and 14-3-3 proteins in vitro.
Effects of Cone Connexin-36 Disruption on Light Adaptation and Circadian Regulation of the Photopic ERG.
Gap Junctions in A8 Amacrine Cells Are Made of Connexin36 but Are Differently Regulated Than Gap Junctions in AII Amacrine Cells.
Extrasynaptic NMDA Receptors on Rod Pathway Amacrine Cells: Molecular Composition, Activation, and Signaling.
Downregulation of Neuronal and Dendritic Connexin36-Made Electrical Synapses Without Glutamatergic Axon Terminals in Spinal Anterior Horn Cells From the Early Stage of Amyotrophic Lateral Sclerosis.
Differential Distribution of Retinal Ca(2+)/Calmodulin-Dependent Kinase II (CaMKII) Isoforms Indicates CaMKII-β and -δ as Specific Elements of Electrical Synapses Made of Connexin36 (Cx36).
Connexin30.2: In Vitro Interaction with Connexin36 in HeLa Cells and Expression in AII Amacrine Cells and Intrinsically Photosensitive Ganglion Cells in the Mouse Retina.
Expression of connexin 26 and connexin 43 is reduced in Hirschsprung's disease.
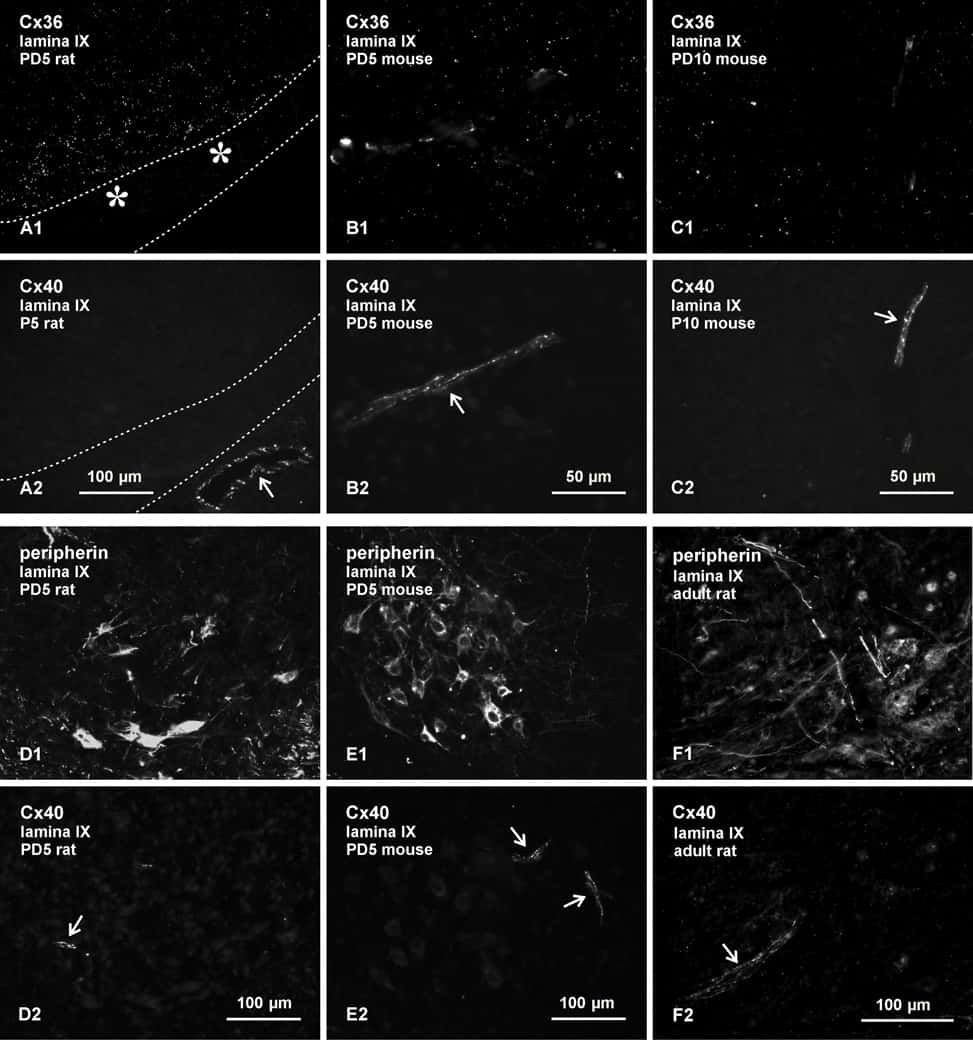
Expression and Localization of Connexins in the Outer Retina of the Mouse.
Abundance of gap junctions at glutamatergic mixed synapses in adult Mosquitofish spinal cord neurons.
Maternal treatment with glucocorticoids modulates gap junction protein expression in the ovine fetal brain.
Re-evaluation of connexins associated with motoneurons in rodent spinal cord, sexually dimorphic motor nuclei and trigeminal motor nucleus.
Molecular and functional asymmetry at a vertebrate electrical synapse.
Diurnal and circadian regulation of connexin 36 transcript and protein in the mammalian retina.
The rod pathway of the microbat retina has bistratified rod bipolar cells and tristratified AII amacrine cells.
Connexins and steroidogenesis in mouse Leydig cells.
Mixed Electrical-Chemical Synapses in Adult Rat Hippocampus are Primarily Glutamatergic and Coupled by Connexin-36.
Connexin and AMPA receptor expression changes over time in the rat olfactory bulb.
Subcellular distribution of connexin45 in OFF bipolar cells of the mouse retina.
Modafinil normalized hyperreflexia after spinal transection in adult rats.
Unique expression of connexins in the human cochlea.
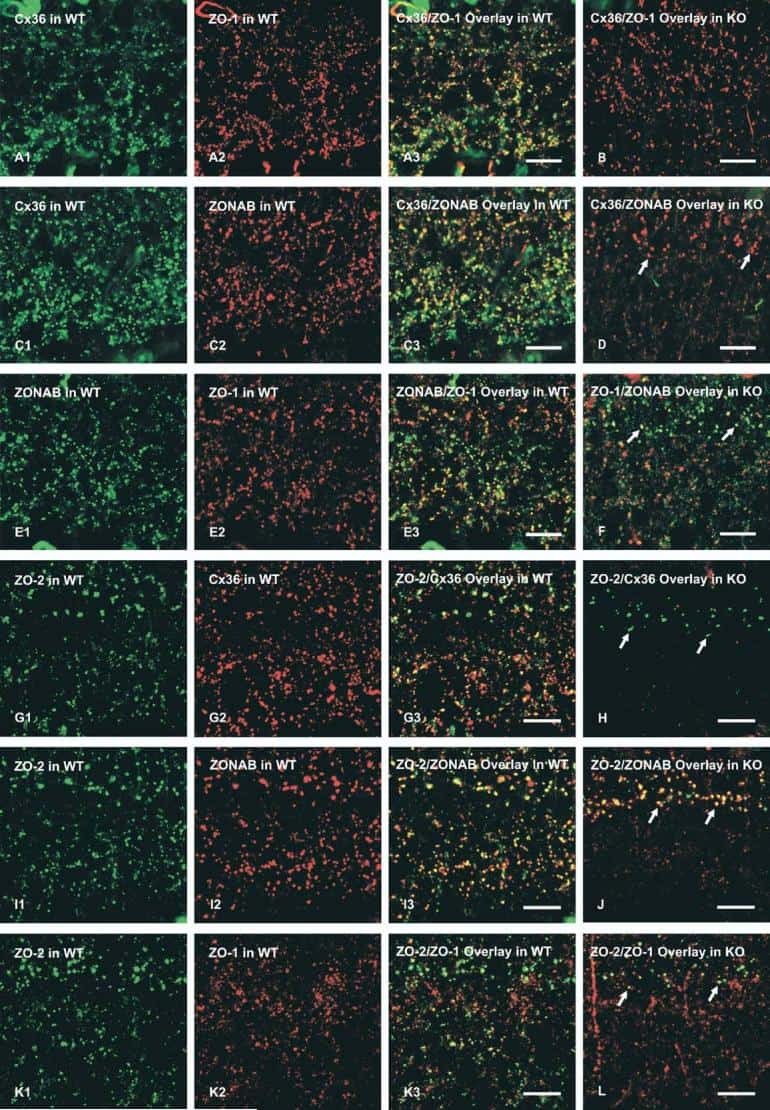
Connexin45-containing neuronal gap junctions in rodent retina also contain connexin36 in both apposing hemiplaques, forming bihomotypic gap junctions, with scaffolding contributed by zonula occludens-1.
The effects of passive exercise therapy initiated prior to or after the development of hyperreflexia following spinal transection.
Neuronal connexin expression in the cochlear nucleus of big brown bats.
The onset of hyperreflexia in the rat following complete spinal cord transection.
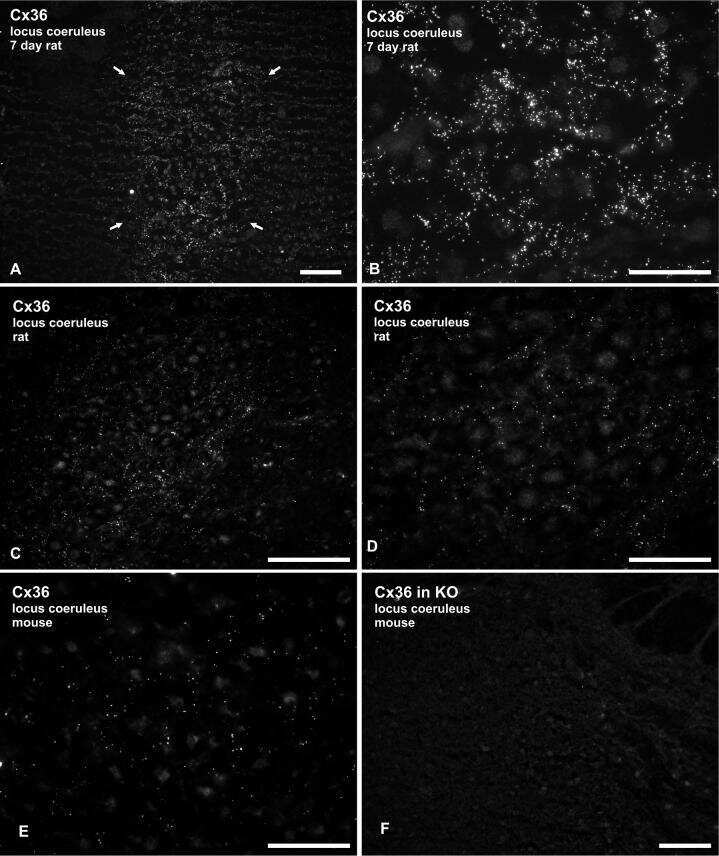
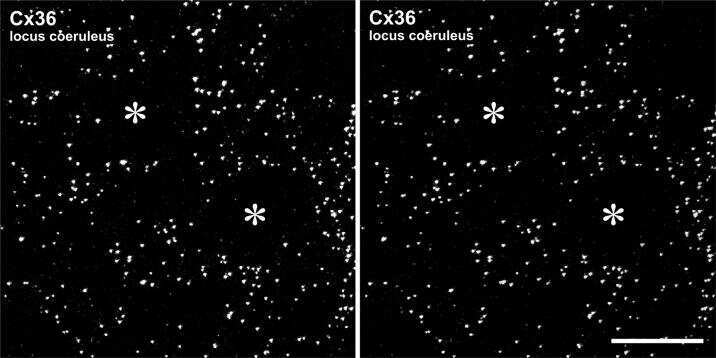
Identification of connexin36 in gap junctions between neurons in rodent locus coeruleus.
Spatial relationships of connexin36, connexin57 and zonula occludens-1 in the outer plexiform layer of mouse retina.
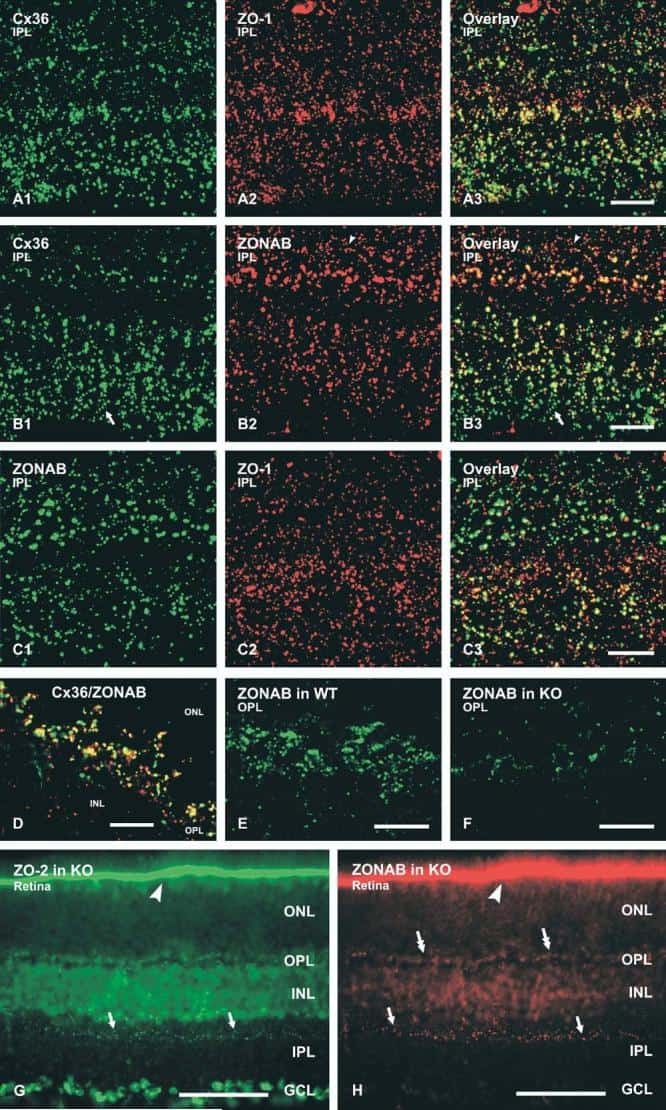
Association of connexin36 and zonula occludens-1 with zonula occludens-2 and the transcription factor zonula occludens-1-associated nucleic acid-binding protein at neuronal gap junctions in rodent retina.
Ultrastructural localization of connexins (Cx36, Cx43, Cx45), glutamate receptors and aquaporin-4 in rodent olfactory mucosa, olfactory nerve and olfactory bulb.
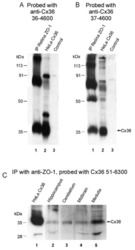
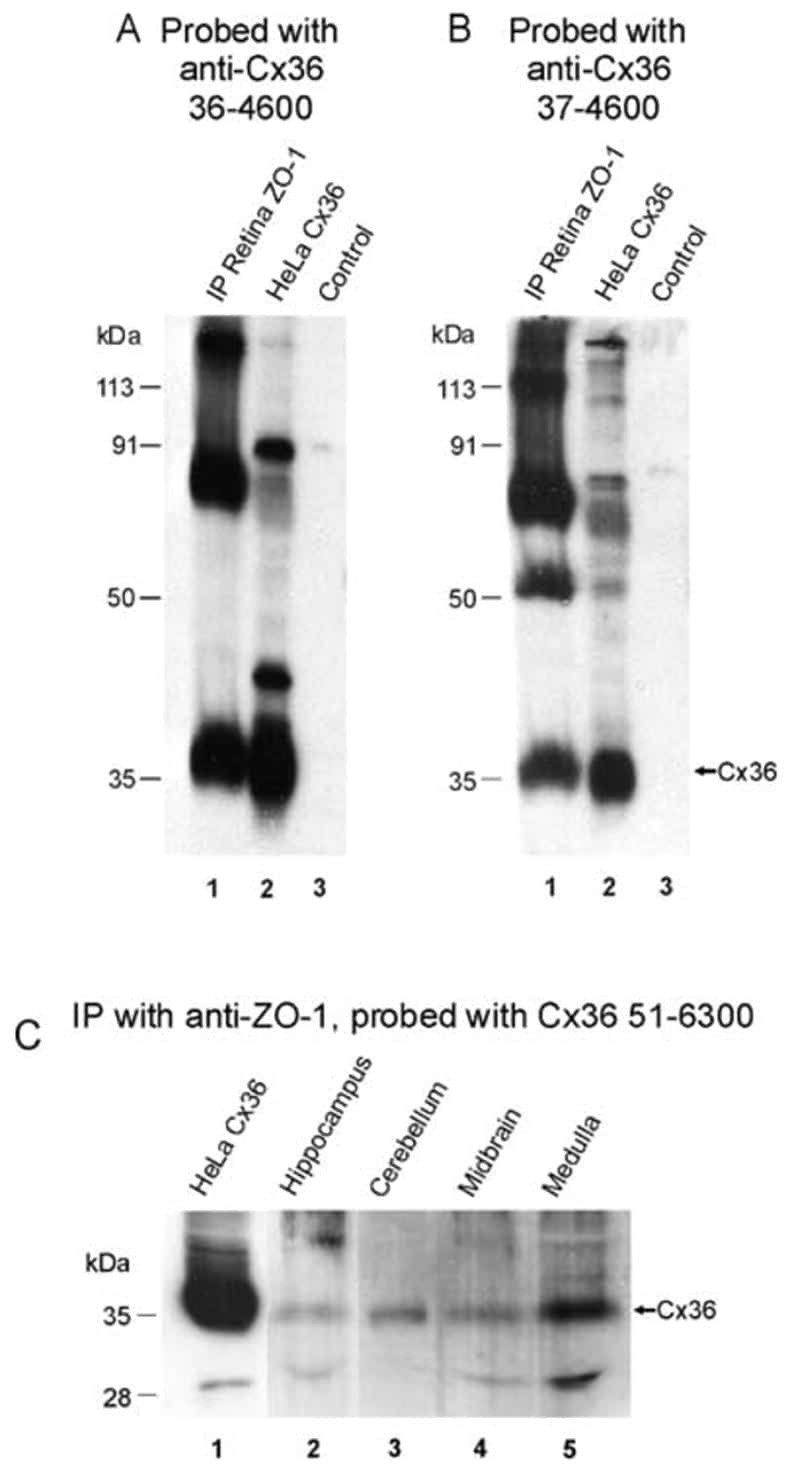
Association of connexin36 with zonula occludens-1 in HeLa cells, betaTC-3 cells, pancreas, and adrenal gland.
Neuronal connexin36 association with zonula occludens-1 protein (ZO-1) in mouse brain and interaction with the first PDZ domain of ZO-1.
Chen AM, Azar SS, Harris A, Brecha NC, Pérez de Sevilla Müller L
Frontiers in neuroanatomy 2021;15:629244
Frontiers in neuroanatomy 2021;15:629244
Synaptic protein interaction networks encode experience by assuming stimulus-specific and brain-region-specific states.
Lautz JD, Tsegay KB, Zhu Z, Gniffke EP, Welsh JP, Smith SEP
Cell reports 2021 Nov 30;37(9):110076
Cell reports 2021 Nov 30;37(9):110076
Amacrine Cells Forming Gap Junctions With Intrinsically Photosensitive Retinal Ganglion Cells: ipRGC Types, Neuromodulator Contents, and Connexin Isoform.
Harrison KR, Chervenak AP, Resnick SM, Reifler AN, Wong KY
Investigative ophthalmology & visual science 2021 Jan 4;62(1):10
Investigative ophthalmology & visual science 2021 Jan 4;62(1):10
Phosphorylation of Connexin36 near the C-terminus switches binding affinities for PDZ-domain and 14-3-3 proteins in vitro.
Tetenborg S, Wang HY, Nemitz L, Depping A, Espejo AB, Aseervatham J, Bedford MT, Janssen-Bienhold U, O'Brien J, Dedek K
Scientific reports 2020 Oct 27;10(1):18378
Scientific reports 2020 Oct 27;10(1):18378
Effects of Cone Connexin-36 Disruption on Light Adaptation and Circadian Regulation of the Photopic ERG.
Zhang S, Lyuboslavsky P, Dixon JA, Chrenek MA, Sellers JT, Hamm JM, Ribelayga CP, Zhang Z, Le YZ, Iuvone PM
Investigative ophthalmology & visual science 2020 Jun 3;61(6):24
Investigative ophthalmology & visual science 2020 Jun 3;61(6):24
Gap Junctions in A8 Amacrine Cells Are Made of Connexin36 but Are Differently Regulated Than Gap Junctions in AII Amacrine Cells.
Yadav SC, Tetenborg S, Dedek K
Frontiers in molecular neuroscience 2019;12:99
Frontiers in molecular neuroscience 2019;12:99
Extrasynaptic NMDA Receptors on Rod Pathway Amacrine Cells: Molecular Composition, Activation, and Signaling.
Veruki ML, Zhou Y, Castilho Á, Morgans CW, Hartveit E
The Journal of neuroscience : the official journal of the Society for Neuroscience 2019 Jan 23;39(4):627-650
The Journal of neuroscience : the official journal of the Society for Neuroscience 2019 Jan 23;39(4):627-650
Downregulation of Neuronal and Dendritic Connexin36-Made Electrical Synapses Without Glutamatergic Axon Terminals in Spinal Anterior Horn Cells From the Early Stage of Amyotrophic Lateral Sclerosis.
Kobayakawa Y, Masaki K, Yamasaki R, Shiraishi W, Hayashida S, Hayashi S, Okamoto K, Matsushita T, Kira JI
Frontiers in neuroscience 2018;12:894
Frontiers in neuroscience 2018;12:894
Differential Distribution of Retinal Ca(2+)/Calmodulin-Dependent Kinase II (CaMKII) Isoforms Indicates CaMKII-β and -δ as Specific Elements of Electrical Synapses Made of Connexin36 (Cx36).
Tetenborg S, Yadav SC, Hormuzdi SG, Monyer H, Janssen-Bienhold U, Dedek K
Frontiers in molecular neuroscience 2017;10:425
Frontiers in molecular neuroscience 2017;10:425
Connexin30.2: In Vitro Interaction with Connexin36 in HeLa Cells and Expression in AII Amacrine Cells and Intrinsically Photosensitive Ganglion Cells in the Mouse Retina.
Meyer A, Tetenborg S, Greb H, Segelken J, Dorgau B, Weiler R, Hormuzdi SG, Janssen-Bienhold U, Dedek K
Frontiers in molecular neuroscience 2016;9:36
Frontiers in molecular neuroscience 2016;9:36
Expression of connexin 26 and connexin 43 is reduced in Hirschsprung's disease.
Coyle D, Doyle B, Murphy JM, O'Donnell AM, Gillick J, Puri P
The Journal of surgical research 2016 Nov;206(1):242-251
The Journal of surgical research 2016 Nov;206(1):242-251
Expression and Localization of Connexins in the Outer Retina of the Mouse.
Bolte P, Herrling R, Dorgau B, Schultz K, Feigenspan A, Weiler R, Dedek K, Janssen-Bienhold U
Journal of molecular neuroscience : MN 2016 Feb;58(2):178-92
Journal of molecular neuroscience : MN 2016 Feb;58(2):178-92
Abundance of gap junctions at glutamatergic mixed synapses in adult Mosquitofish spinal cord neurons.
Serrano-Velez JL, Rodriguez-Alvarado M, Torres-Vazquez II, Fraser SE, Yasumura T, Vanderpool KG, Rash JE, Rosa-Molinar E
Frontiers in neural circuits 2014;8:66
Frontiers in neural circuits 2014;8:66
Maternal treatment with glucocorticoids modulates gap junction protein expression in the ovine fetal brain.
Sadowska GB, Stonestreet BS
Neuroscience 2014 Sep 5;275:248-58
Neuroscience 2014 Sep 5;275:248-58
Re-evaluation of connexins associated with motoneurons in rodent spinal cord, sexually dimorphic motor nuclei and trigeminal motor nucleus.
Bautista W, Rash JE, Vanderpool KG, Yasumura T, Nagy JI
The European journal of neuroscience 2014 Mar;39(5):757-70
The European journal of neuroscience 2014 Mar;39(5):757-70
Molecular and functional asymmetry at a vertebrate electrical synapse.
Rash JE, Curti S, Vanderpool KG, Kamasawa N, Nannapaneni S, Palacios-Prado N, Flores CE, Yasumura T, O'Brien J, Lynn BD, Bukauskas FF, Nagy JI, Pereda AE
Neuron 2013 Sep 4;79(5):957-69
Neuron 2013 Sep 4;79(5):957-69
Diurnal and circadian regulation of connexin 36 transcript and protein in the mammalian retina.
Katti C, Butler R, Sekaran S
Investigative ophthalmology & visual science 2013 Jan 28;54(1):821-9
Investigative ophthalmology & visual science 2013 Jan 28;54(1):821-9
The rod pathway of the microbat retina has bistratified rod bipolar cells and tristratified AII amacrine cells.
Müller B, Butz E, Peichl L, Haverkamp S
The Journal of neuroscience : the official journal of the Society for Neuroscience 2013 Jan 16;33(3):1014-23
The Journal of neuroscience : the official journal of the Society for Neuroscience 2013 Jan 16;33(3):1014-23
Connexins and steroidogenesis in mouse Leydig cells.
Li D, Sekhon P, Barr KJ, Márquez-Rosado L, Lampe PD, Kidder GM
Canadian journal of physiology and pharmacology 2013 Feb;91(2):157-64
Canadian journal of physiology and pharmacology 2013 Feb;91(2):157-64
Mixed Electrical-Chemical Synapses in Adult Rat Hippocampus are Primarily Glutamatergic and Coupled by Connexin-36.
Hamzei-Sichani F, Davidson KG, Yasumura T, Janssen WG, Wearne SL, Hof PR, Traub RD, Gutiérrez R, Ottersen OP, Rash JE
Frontiers in neuroanatomy 2012;6:13
Frontiers in neuroanatomy 2012;6:13
Connexin and AMPA receptor expression changes over time in the rat olfactory bulb.
Corthell JT, Fadool DA, Trombley PQ
Neuroscience 2012 Oct 11;222:38-48
Neuroscience 2012 Oct 11;222:38-48
Subcellular distribution of connexin45 in OFF bipolar cells of the mouse retina.
Hilgen G, von Maltzahn J, Willecke K, Weiler R, Dedek K
The Journal of comparative neurology 2011 Feb 15;519(3):433-50
The Journal of comparative neurology 2011 Feb 15;519(3):433-50
Modafinil normalized hyperreflexia after spinal transection in adult rats.
Yates CC, Charlesworth A, Reese NB, Ishida K, Skinner RD, Garcia-Rill E
Spinal cord 2009 Jun;47(6):481-5
Spinal cord 2009 Jun;47(6):481-5
Unique expression of connexins in the human cochlea.
Liu W, Boström M, Kinnefors A, Rask-Andersen H
Hearing research 2009 Apr;250(1-2):55-62
Hearing research 2009 Apr;250(1-2):55-62
Connexin45-containing neuronal gap junctions in rodent retina also contain connexin36 in both apposing hemiplaques, forming bihomotypic gap junctions, with scaffolding contributed by zonula occludens-1.
Li X, Kamasawa N, Ciolofan C, Olson CO, Lu S, Davidson KG, Yasumura T, Shigemoto R, Rash JE, Nagy JI
The Journal of neuroscience : the official journal of the Society for Neuroscience 2008 Sep 24;28(39):9769-89
The Journal of neuroscience : the official journal of the Society for Neuroscience 2008 Sep 24;28(39):9769-89
The effects of passive exercise therapy initiated prior to or after the development of hyperreflexia following spinal transection.
Yates CC, Charlesworth A, Reese NB, Skinner RD, Garcia-Rill E
Experimental neurology 2008 Oct;213(2):405-9
Experimental neurology 2008 Oct;213(2):405-9
Neuronal connexin expression in the cochlear nucleus of big brown bats.
Horowitz SS, Stamper SA, Simmons JA
Brain research 2008 Mar 4;1197:76-84
Brain research 2008 Mar 4;1197:76-84
The onset of hyperreflexia in the rat following complete spinal cord transection.
Yates C, Charlesworth A, Allen SR, Reese NB, Skinner RD, Garcia-Rill E
Spinal cord 2008 Dec;46(12):798-803
Spinal cord 2008 Dec;46(12):798-803
Identification of connexin36 in gap junctions between neurons in rodent locus coeruleus.
Rash JE, Olson CO, Davidson KG, Yasumura T, Kamasawa N, Nagy JI
Neuroscience 2007 Jul 29;147(4):938-56
Neuroscience 2007 Jul 29;147(4):938-56
Spatial relationships of connexin36, connexin57 and zonula occludens-1 in the outer plexiform layer of mouse retina.
Ciolofan C, Lynn BD, Wellershaus K, Willecke K, Nagy JI
Neuroscience 2007 Aug 24;148(2):473-88
Neuroscience 2007 Aug 24;148(2):473-88
Association of connexin36 and zonula occludens-1 with zonula occludens-2 and the transcription factor zonula occludens-1-associated nucleic acid-binding protein at neuronal gap junctions in rodent retina.
Ciolofan C, Li XB, Olson C, Kamasawa N, Gebhardt BR, Yasumura T, Morita M, Rash JE, Nagy JI
Neuroscience 2006 Jun 30;140(2):433-51
Neuroscience 2006 Jun 30;140(2):433-51
Ultrastructural localization of connexins (Cx36, Cx43, Cx45), glutamate receptors and aquaporin-4 in rodent olfactory mucosa, olfactory nerve and olfactory bulb.
Rash JE, Davidson KG, Kamasawa N, Yasumura T, Kamasawa M, Zhang C, Michaels R, Restrepo D, Ottersen OP, Olson CO, Nagy JI
Journal of neurocytology 2005 Sep;34(3-5):307-41
Journal of neurocytology 2005 Sep;34(3-5):307-41
Association of connexin36 with zonula occludens-1 in HeLa cells, betaTC-3 cells, pancreas, and adrenal gland.
Li X, Olson C, Lu S, Nagy JI
Histochemistry and cell biology 2004 Nov;122(5):485-98
Histochemistry and cell biology 2004 Nov;122(5):485-98
Neuronal connexin36 association with zonula occludens-1 protein (ZO-1) in mouse brain and interaction with the first PDZ domain of ZO-1.
Li X, Olson C, Lu S, Kamasawa N, Yasumura T, Rash JE, Nagy JI
The European journal of neuroscience 2004 Apr;19(8):2132-46
The European journal of neuroscience 2004 Apr;19(8):2132-46
No comments: Submit comment
Supportive validation
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
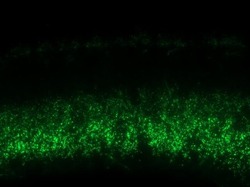
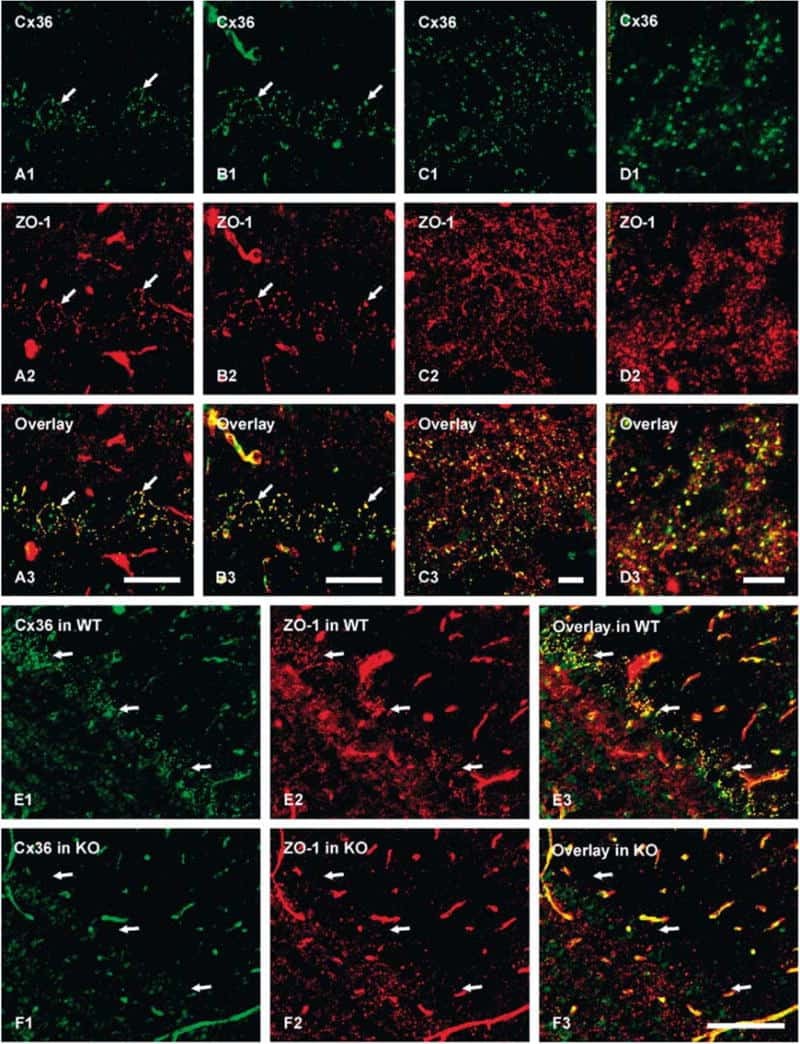
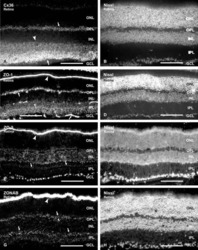
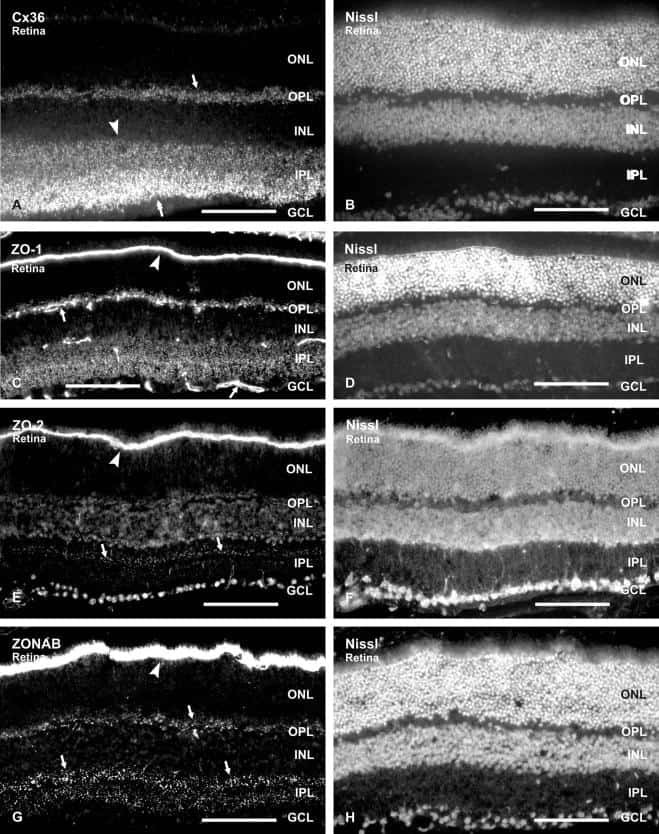
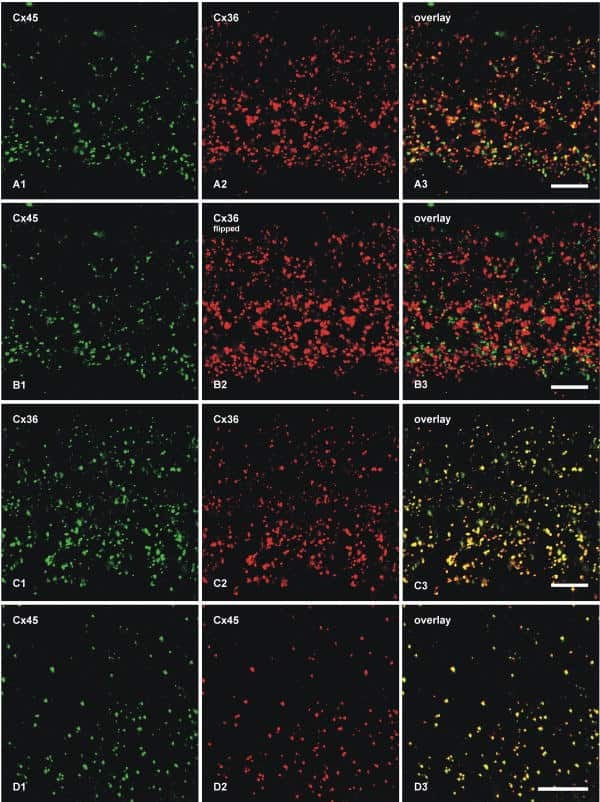
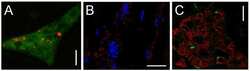
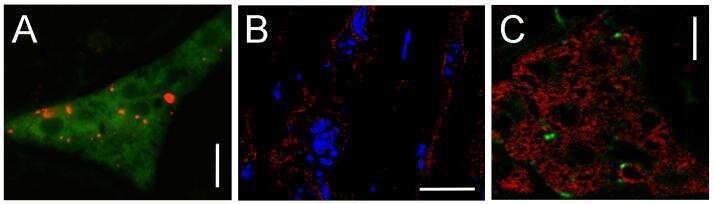
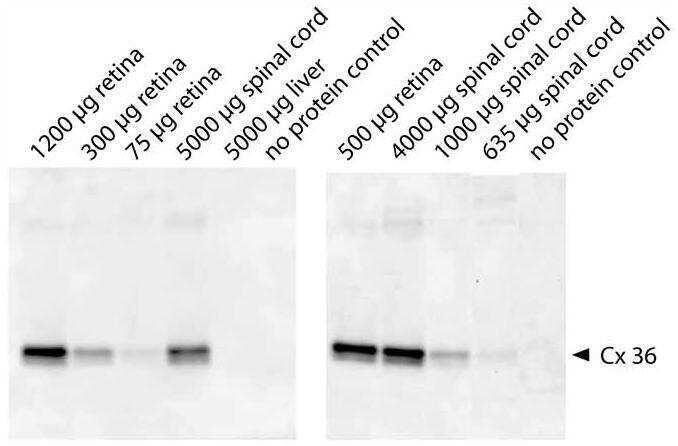
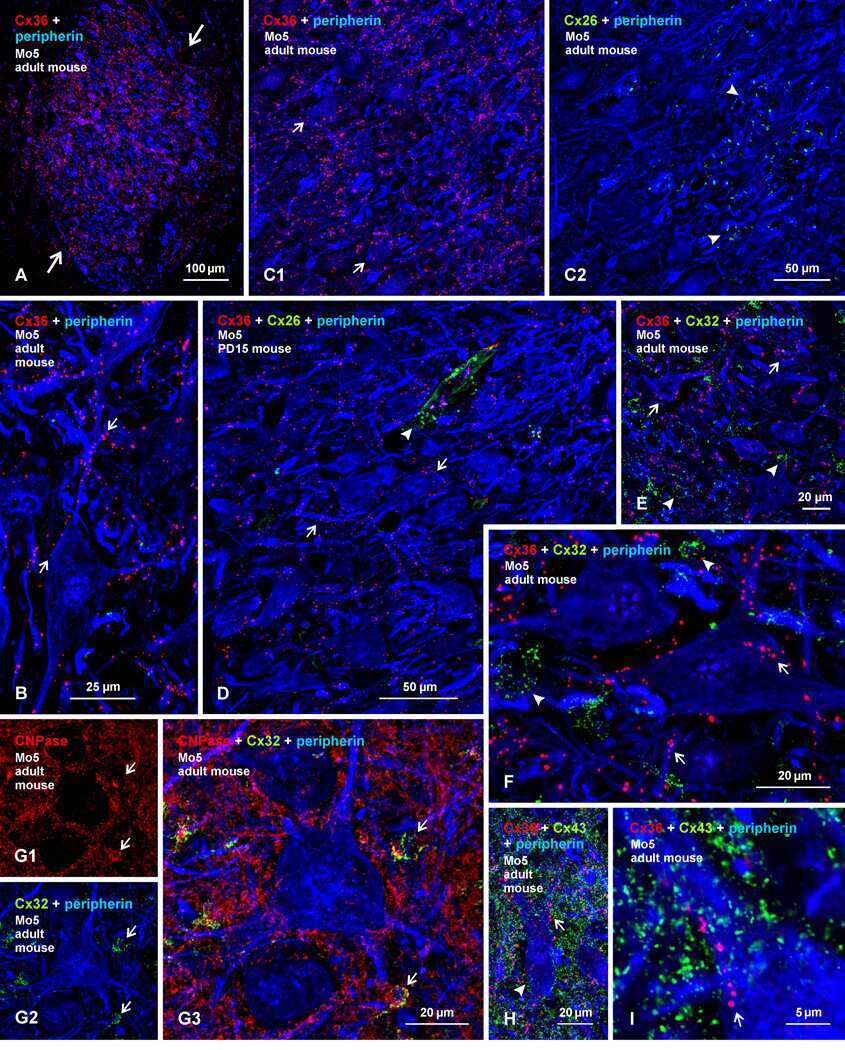
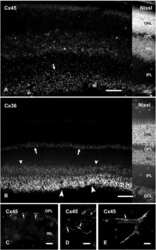
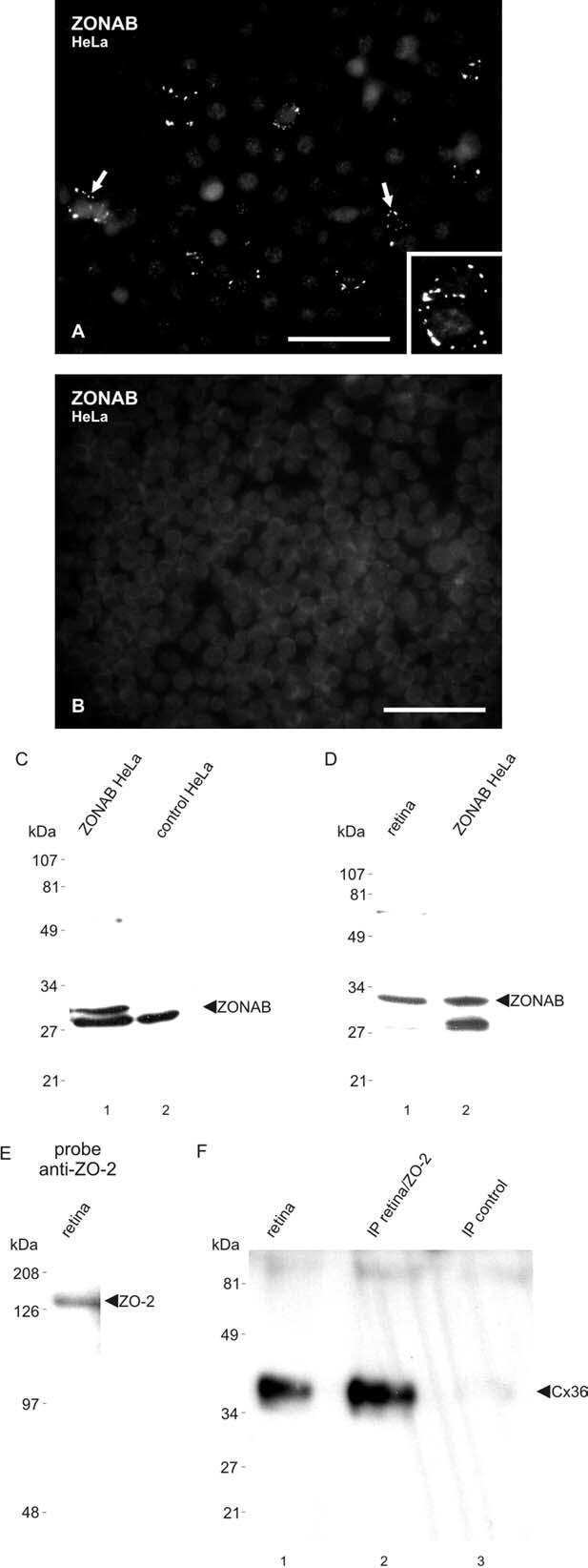
- Immunofluorescence was performed on sections of adult mouse retina. Tissue sections were probed overnight at 4°C with mouse monoclonal anti-Cx36 antibody (Product # 37-4600, 2 µg/mL), and detected with donkey anti-mouse Alexa Fluor® 488 conjugate. Images were taken on a Zeiss Z2 scanning microscope at 40X magnification, and demonstrate antibody labeling for Cx36 localized at gap junctions between neurons in the inner plexiform layer of mouse retina. Image courtesy of Dr. James Nagy, University of Manitoba, Winnipeg, Canada.
Supportive validation
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
- NULL

- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
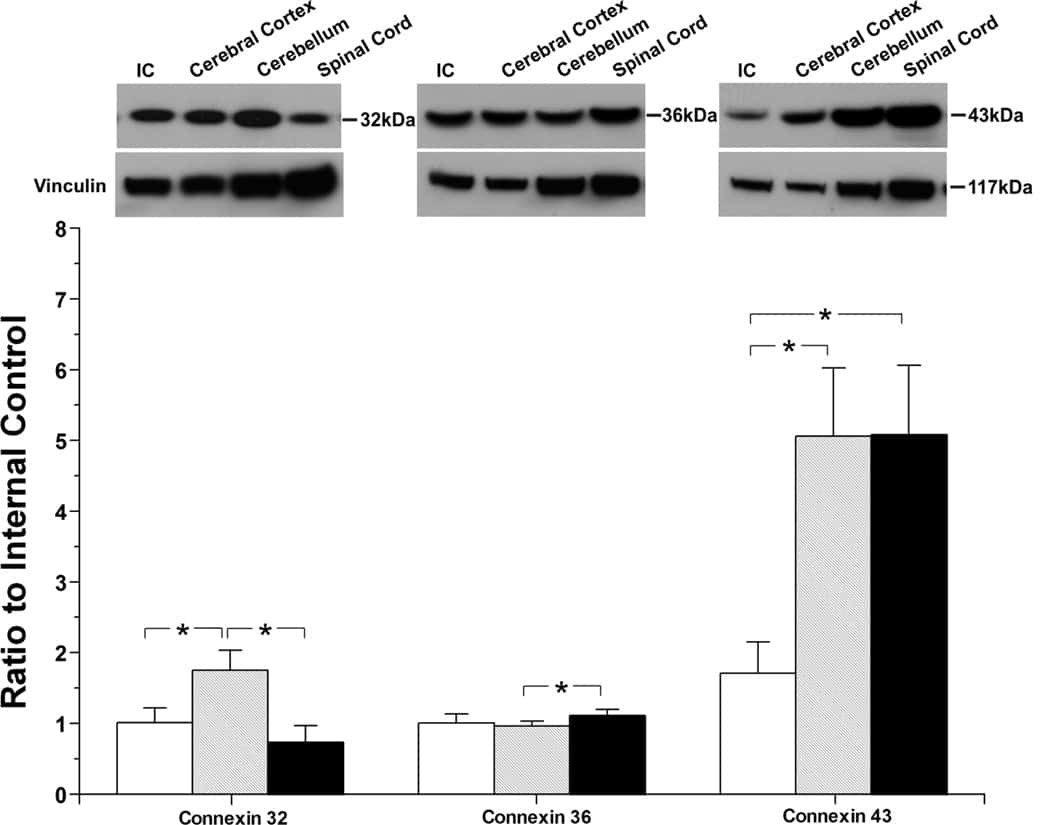
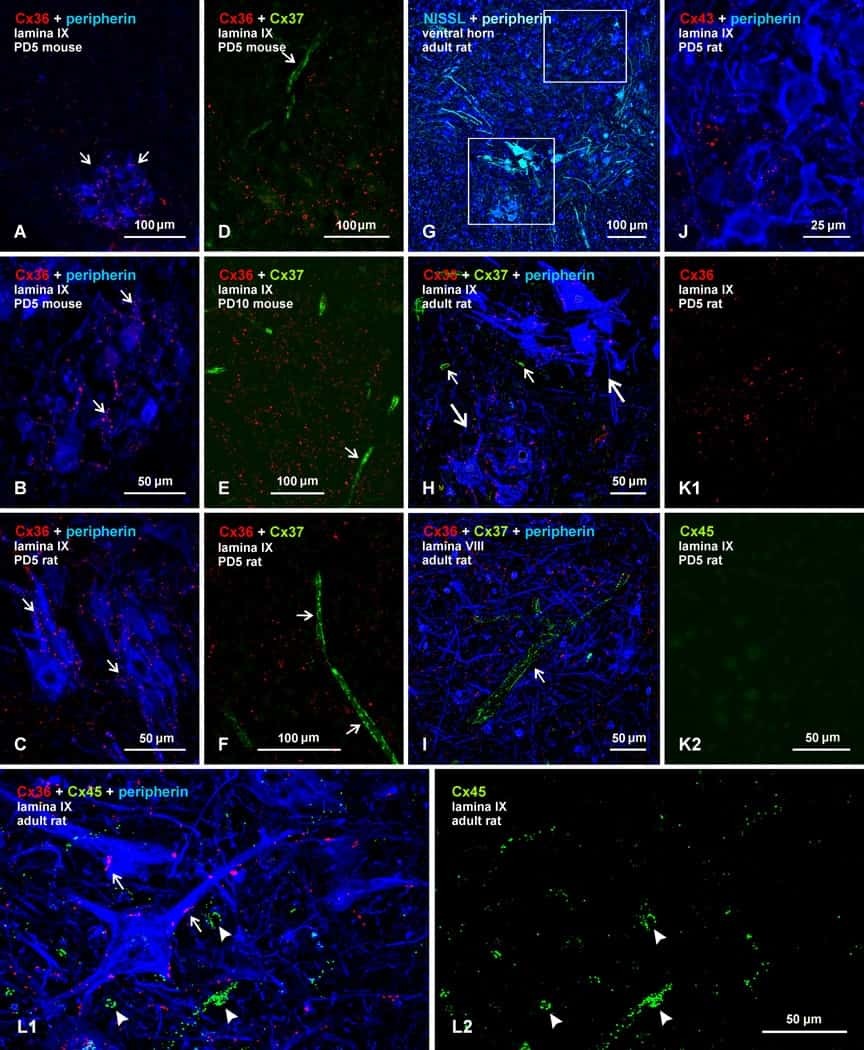
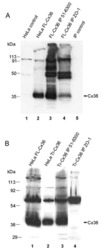
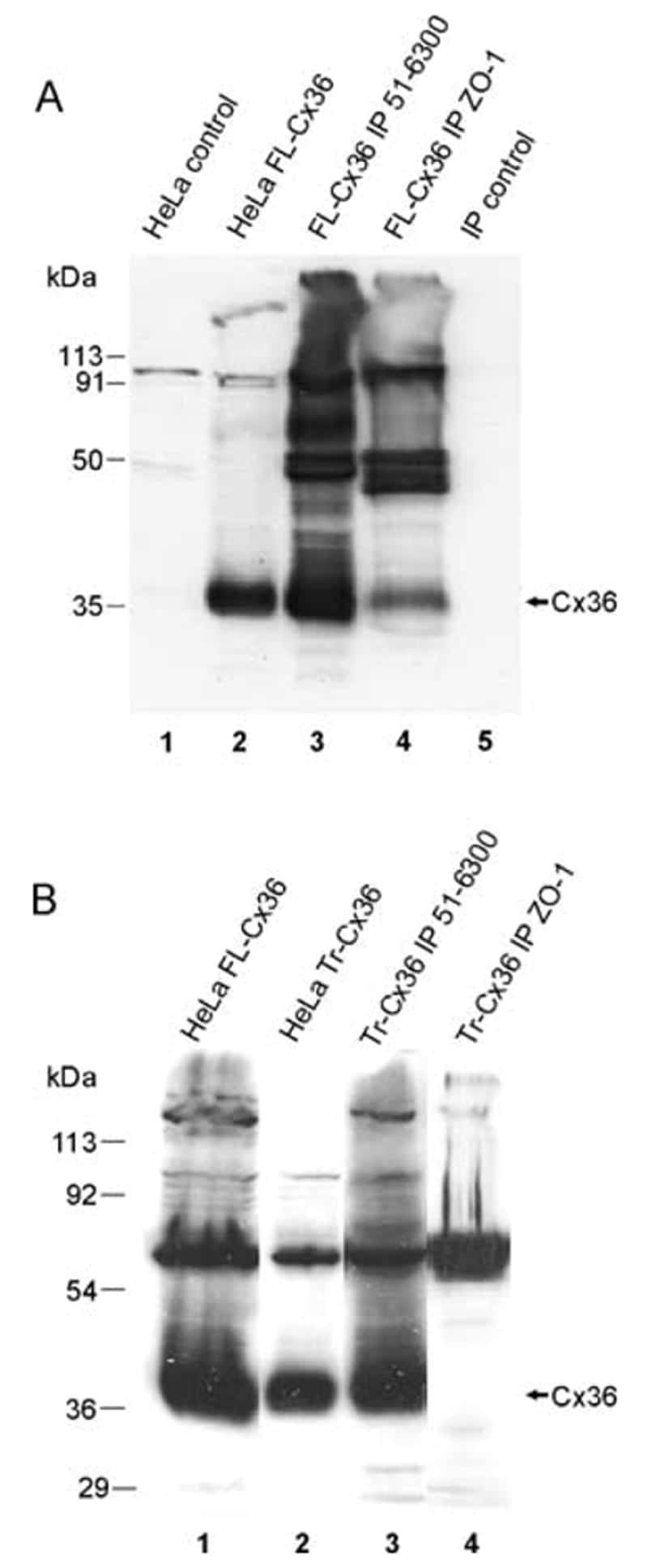
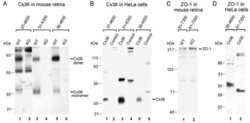
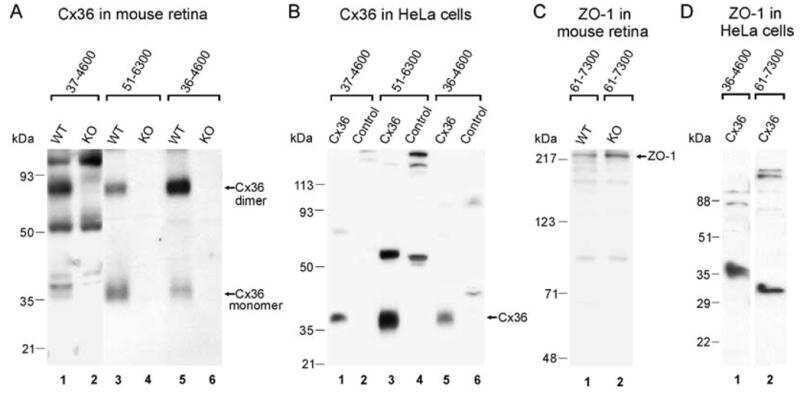
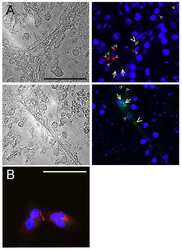
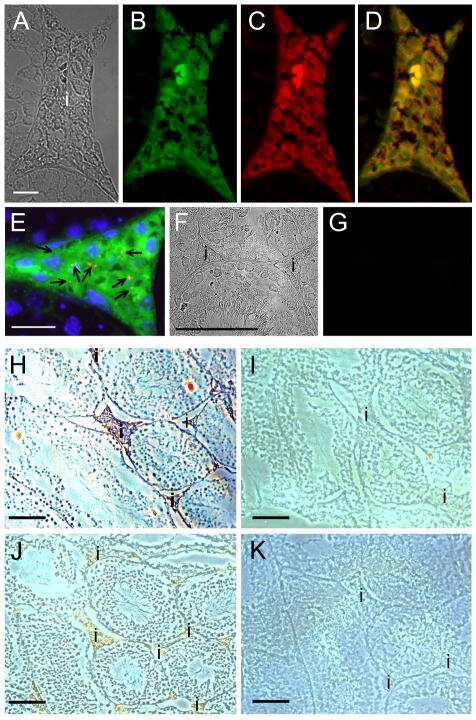
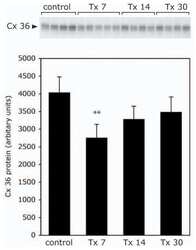
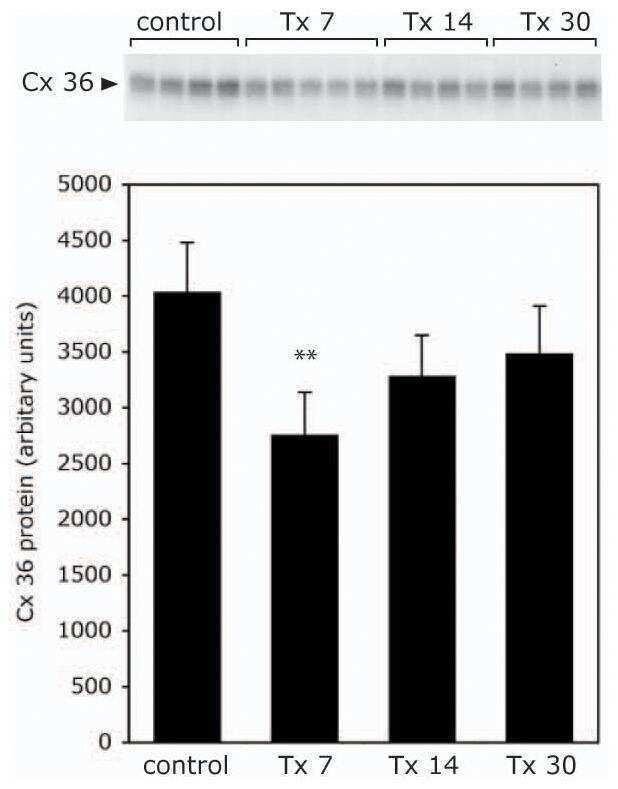
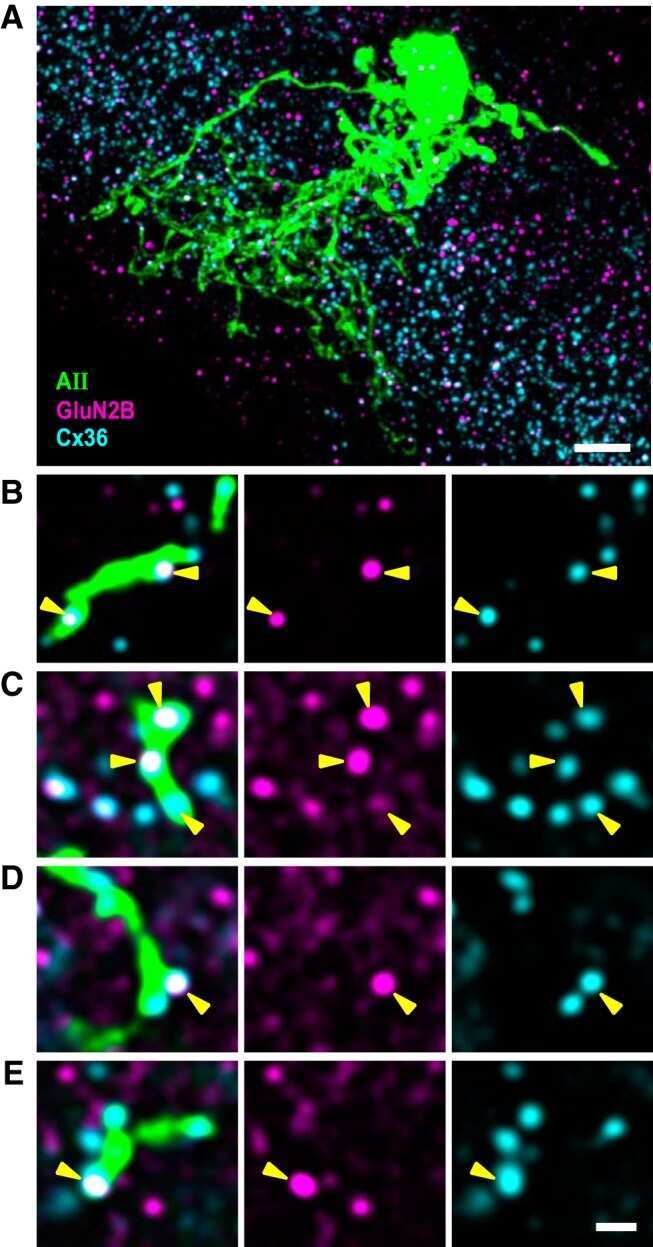
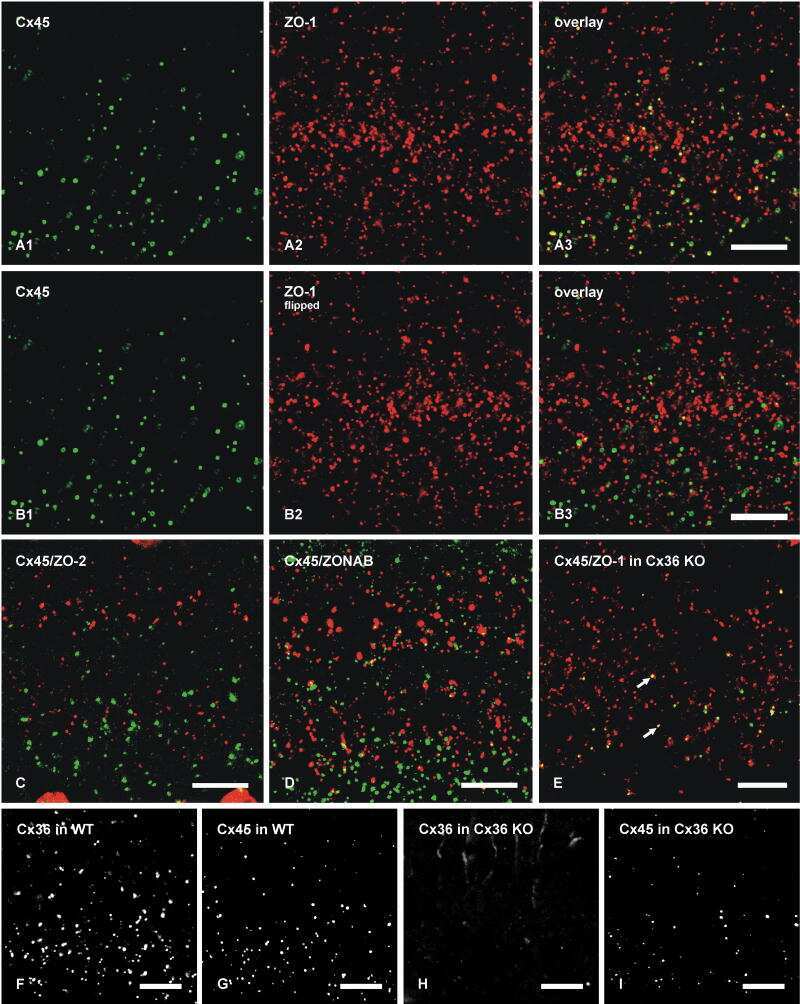
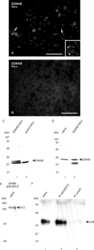
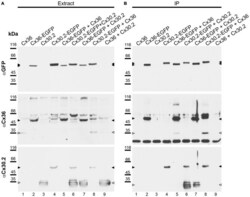
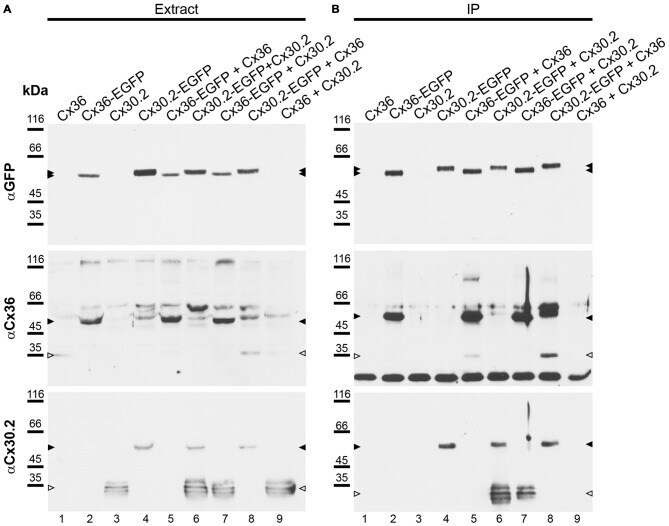
- Figure 4 Newly generated anti-Cx30.2 antibodies (rabbit) are specific and reveal interaction of Cx30.2 and Cx36. (A) Western blot analysis of HeLa cell extracts transfected with different combinations of Cx30.2 and Cx36 and their enhanced green fluorescent protein (EGFP)-tagged variants. Blots were probed with anti-GFP, anti-Cx36 and newly generated anti-Cx30.2 antibodies from rabbit. Anti-Cx36 and anti-Cx30.2 antibodies recognize the respective connexin ( A,B , open arrowheads) and its fusion variant ( A,B , filled arrowheads). Please note that several bands become visible for untagged Cx30.2, pointing to different phosphorylation states (also see Supplementary Figure 1). These bands are not visible for the fusion protein. The Cx36 signal (lane 5) is extremely weak and nearly not visible but becomes apparent after immunoprecipitation (IP; B , lane 5). (B) IP using the same HeLa cell transfections as in (A) . Single or double transfections with untagged connexins did not show any bands in IP, as expected (lanes 1, 3, 9). Most importantly, when cells expressed Cx36-EGFP and untagged Cx30.2, Cx30.2 was detected in the IP (lane 7, lowest blot, open arrowhead); similarly, when cells expressed the Cx30.2-EGFP fusion protein together with Cx36, a band appeared for the native Cx36 (lane 8, middle blot, open arrowhead). These results suggest interaction of Cx30.2 and Cx36 in HeLa cells. Please note that the anti-Cx36 antibody produced several unspecific bands. The strong band at 25 k
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
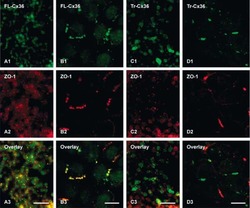
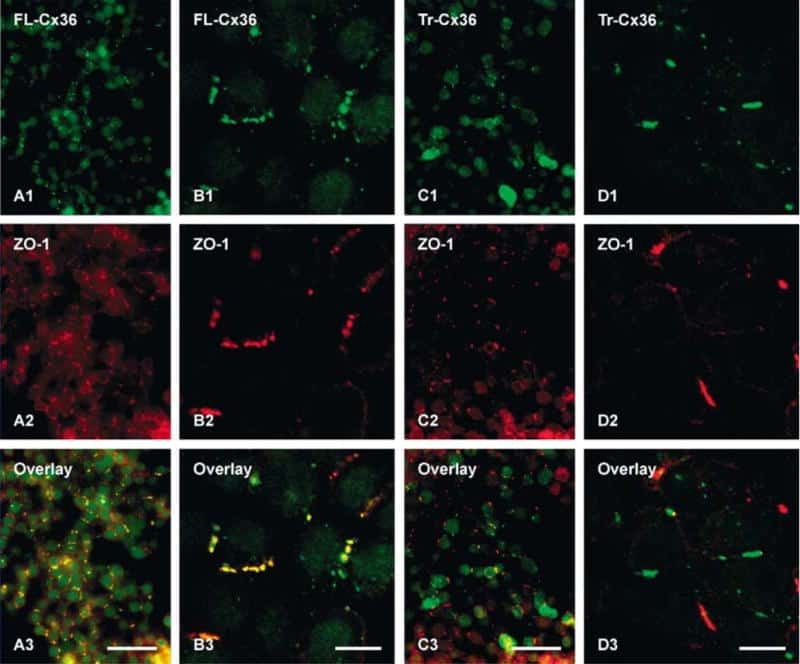
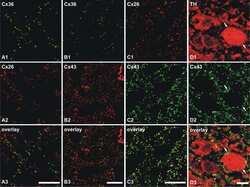
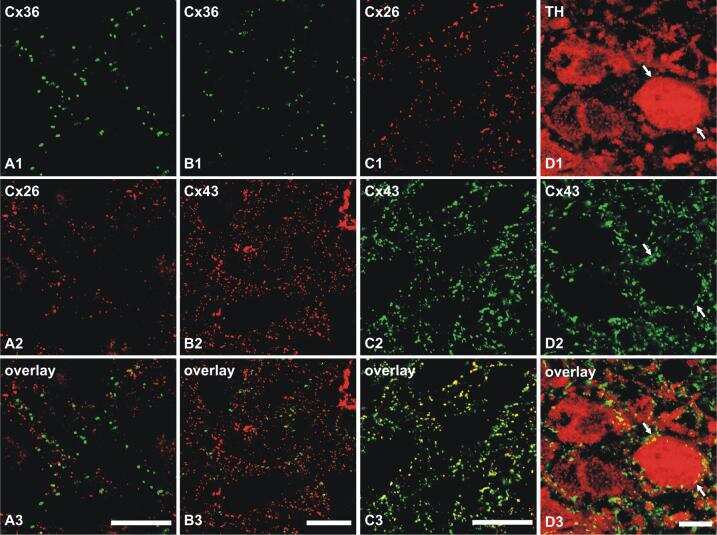
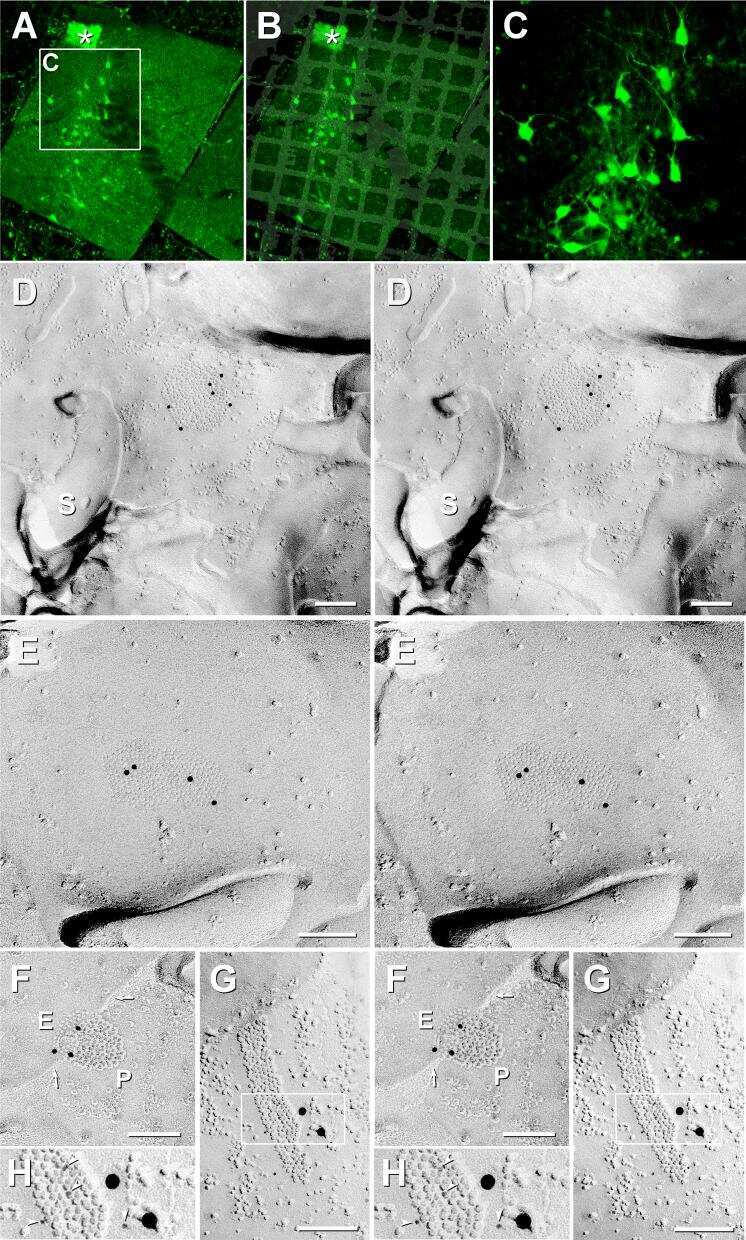
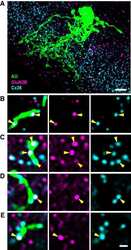
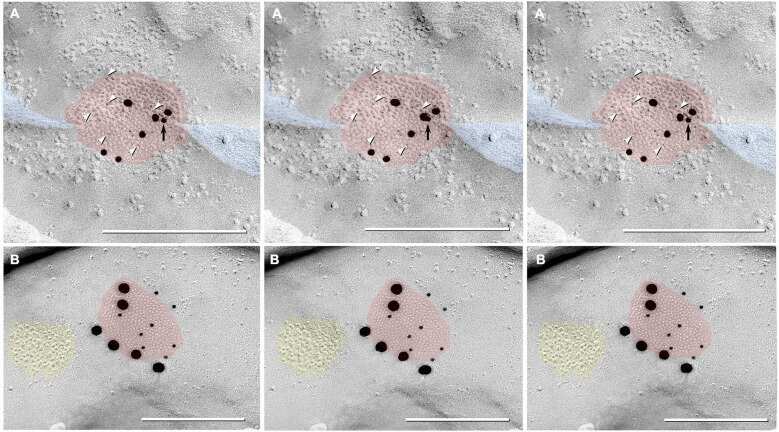
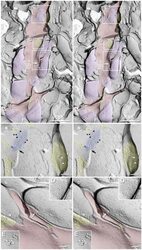
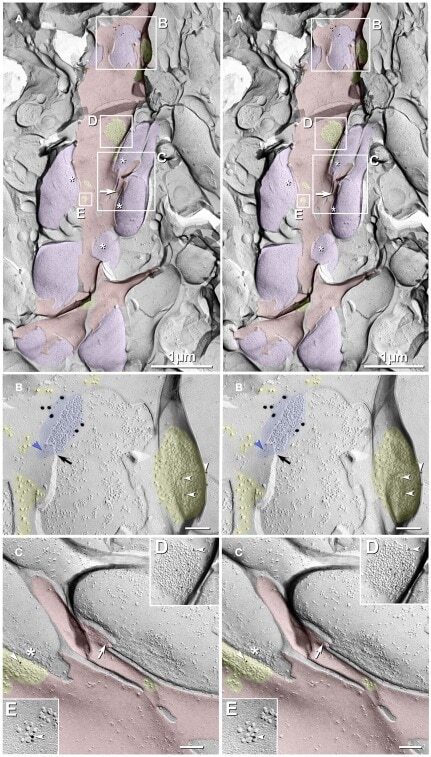
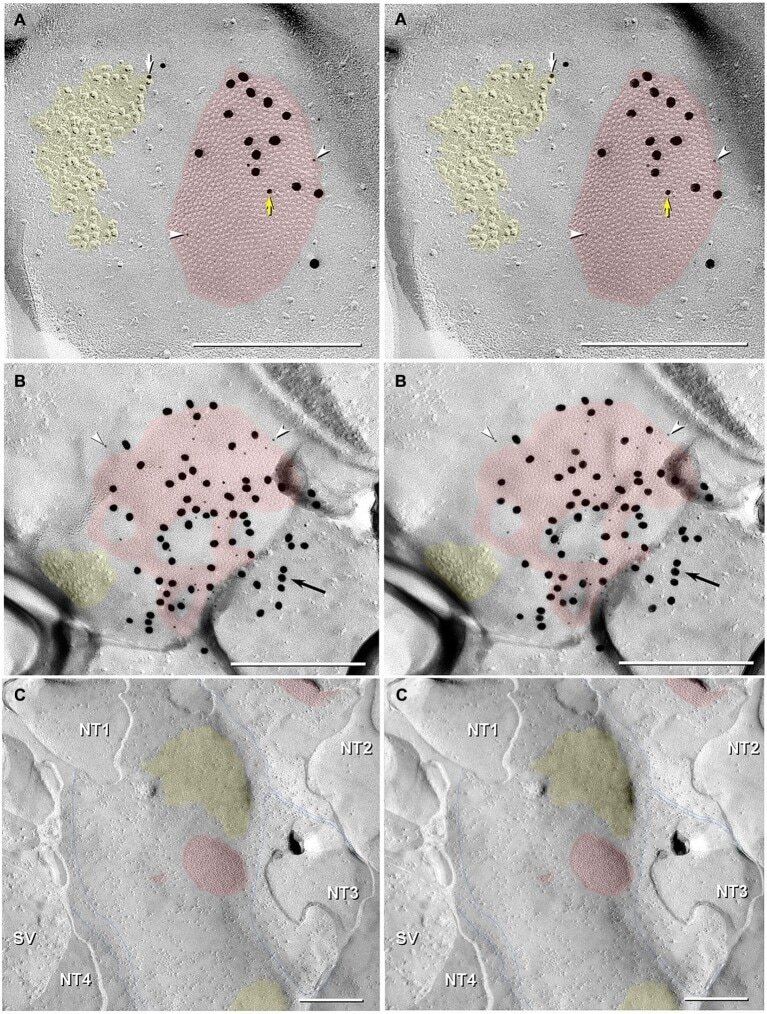
- Figure 3 Freeze-fracture replica immunogold labeling images of a spiny dendrite in stratum oriens , with gap junction double-labeled for Cx36 (6 and 18-nm gold beads ; within blue overlay ) and cluster of 10-nm E-face IMPs labeled for NR1 glutamate receptor subunits ( 12-nm gold beads within yellow overlays ) . (A) Low magnification stereoscopic images of one of three gap junctions labeled for Cx36 found in a 20 x 20 mum area of stratum oriens . A narrow branched spine [ arrow ; shown at higher magnification as (C) ] is consistent with identification of this neuronal plasma membrane E-face as either a portion of a CA3pyr dendrite (Chicurel and Harris, 1992 ) or of a spiny interneuron. A second branched spine is at the bottom ( red overlay ), separated from the co-planar dendritic shaft by ca. 20 nm, potentially indicating that the two membrane areas were continuous before fracturing. (B) Higher magnification stereoscopic image from (A) showing a medium-diameter gap junction ( blue overlay ) that is postsynaptic to a glutamatergic nerve terminal, identified by the presence of a shared PSD that is immunogold-labeled for NMDA-R1 by five 12-nm gold beads ( arrowheads ). Tilting, rotation, and stereoscopic analysis was required to clearly discern the PSD and gold labels beneath the almost vertical aspect of the replicated PSD in (A) . Note narrowing of the extracellular space ( black arrow ) at edge of gap junction. Air-drying of the high-relief replicas caused compression and art
- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
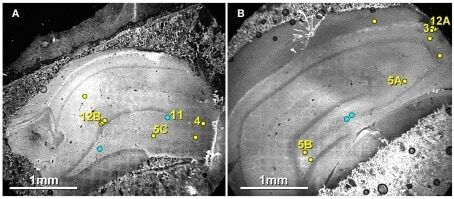
- Figure 6 Confocal maps showing exact locations of 20 Cx36-immunogold-labeled gap junctions found in adult rat hippocampus . (A,B) The labeled gap junctions shown in Figures 3 , 4 , 5 A-C, 11 , and 12 A,B are indicated by numbered yellow dots . Exact locations of seven other Cx36-labeled gap junctions ( unnumbered yellow dots ) found in these same two replicas are also indicated. In addition, the approximate locations of four other Cx36-labeled gap junctions found in three other replicas are indicated ( unnumbered blue dots ). Note the faint image of the gold ""index""TEM grid beneath and bonded to the tissue slice. Scale bars are 1 mm.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
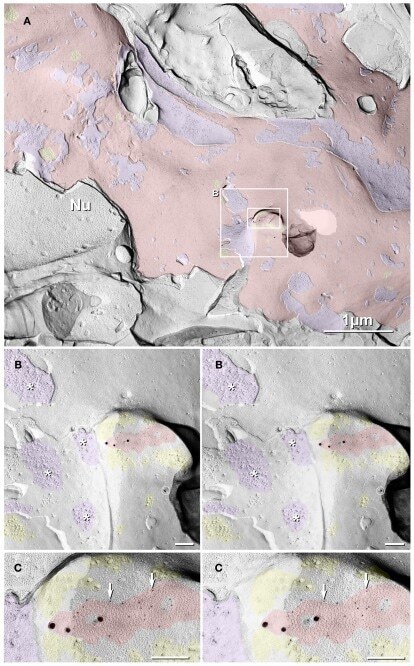
- Figure 10 Dentate granule cell somatic and proximal dendrite plasma membrane E-face, with immunogold-labeled reticular gap junction . Large area of exposed neuronal plasma membrane ( red overlay ; ca. 1/4 of the 10 mum x 10 mum area is shown) is abutted by multiple large-diameter glutamatergic axon terminals ( purple overlays ). This replica was double-labeled for Cx35 + Cx36. Boxed areas are enlarged as stereoscopic images. (B) At intermediate magnification, active zones (*) are visible in the axon terminal P-face, and clusters of 10-nm E-face IMPs are seen in the subjacent granule cell proximal dendrite E-face, demonstrating two primary components defining multiple chemical synapses. In the center of the image is a two-lobed ""mushroom"" spine that, fortuitously, was shadowed deeply [enlarged as (C) ]. Several active zones (* and purple overlays ), one with a closely-associated PSD ( yellow overlay ), are indicated. A single gold bead above the replica, and therefore representing ""definitive noise,"" is indicated by "" ."" (C) At high-magnification, a medium-size (>150 connexons) ""reticular"" gap junction ( red overlay ) was faintly resolved deep in the spine head, but was easily detected because it is double-labeled by three 18-nm gold beads attached to rabbit polyclonal antibody for Cx35 (fish ortholog of Cx36; O'Brien et al., 2004 , 2006 ) and by 18 6-nm gold beads ( white arrows ) that label mouse monoclonal antibody for Cx36. Glutamate-receptor-like E-face IMPs
- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
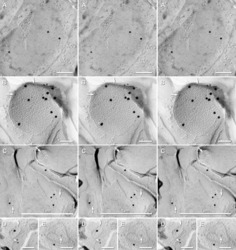
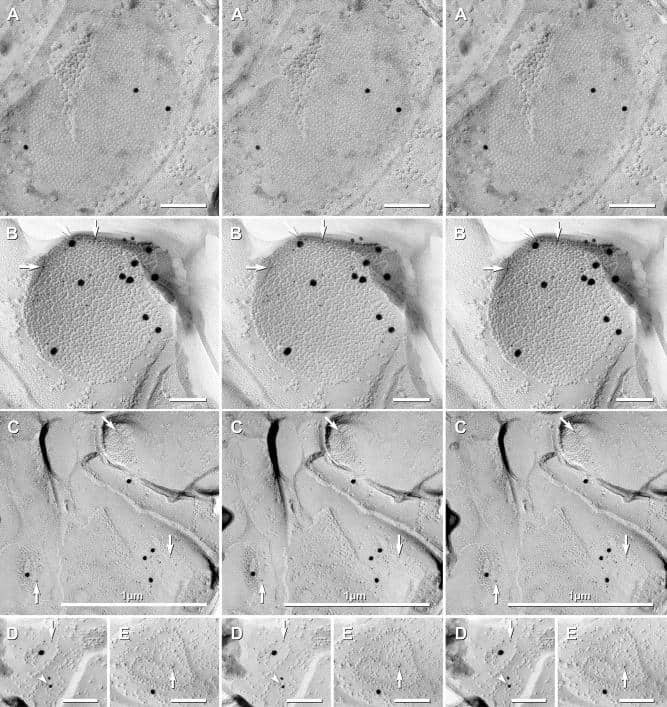
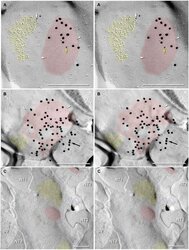
- Figure 11 Images of freeze-fracture replica of immunogold labeled dendritic gap junctions in hippocampus . (A) Large reticular gap junction found between dendritic shafts of two probable interneurons in stratum oriens . It is labeled for Cx36 by ca. 88 6-nm gold beads and 26 18-nm gold beads. (B) Image of three Cx36-immunogold-labeled gap junctions between unidentified neuronal processes (axon terminals or dendrites). A nearby GABAergic synapse ( green overlay ) is on an unidentified cell process. The adjacent neuronal P-face ( purple overlay ) has the high density of IMPs associated with MF axon varicosities. Green overlay = GABA receptor 9-nm P-face IMPs. One of the gap junctions ( box ) is enlarged as an inset ( bottom center ). AZ, active zone of putative GABAergic synapse. Scale bars are 0.1 mum, unless otherwise indicated.

- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- Figure 7 Stereoscopic images of large plaque and reticular gap junctions in adult male Mosquitofish. (A) Large plaque gap junction ( red overlay ) on a neuronal postsynaptic E-face labeled with antibodies against mouse/human Cx36 (36-4600, 6-nm [ white arrowheads ] and much more electron-dense 18-nm gold beads) and for GluR2 AMPA receptors (12-nm gold beads, white arrow ) ( Yellow arrow indicates either a 12-nm gold bead as ""noise"" on the Cx36-labeled gap junction or an anomalously small ""18-nm"" gold bead for Cx36). (B) High-magnification stereoscopic image of E-face of a neuronal reticular gap junction ( red overlay ) labeled for Cx35 (P-Ser276) by 6-nm ( white arrowheads ) and 18-nm gold beads. The black arrow points to ""cryptic"" labeling of connexins in an extended portion of the gap junction beneath the cross-fractured neuronal cytoplasm. Immediately adjacent to the gap junction is a cluster of E-face IMPs ( yellow overlay , not labeled) similar to other immunogold-labeled glutamate receptor channels. (C) P-face image showing three of more than a dozen unlabeled postsynaptic hemiplaques ( red overlays ) in the same replica as Figures 4 , 5B , where presynaptic hemiplaques are heavily labeled for Cx35 (P-Ser276). This absence of labeling in C is consistent with our demonstration in goldfish giant club ending/Mauthner cell mixed synapses that Cx35 is exclusively presynaptic and that Cx34.7 is exclusively postsynaptic (Rash et al., 2013 ). The postsynaptic P-face revea
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
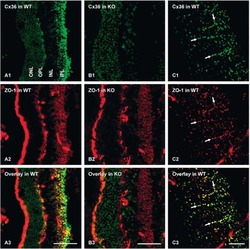
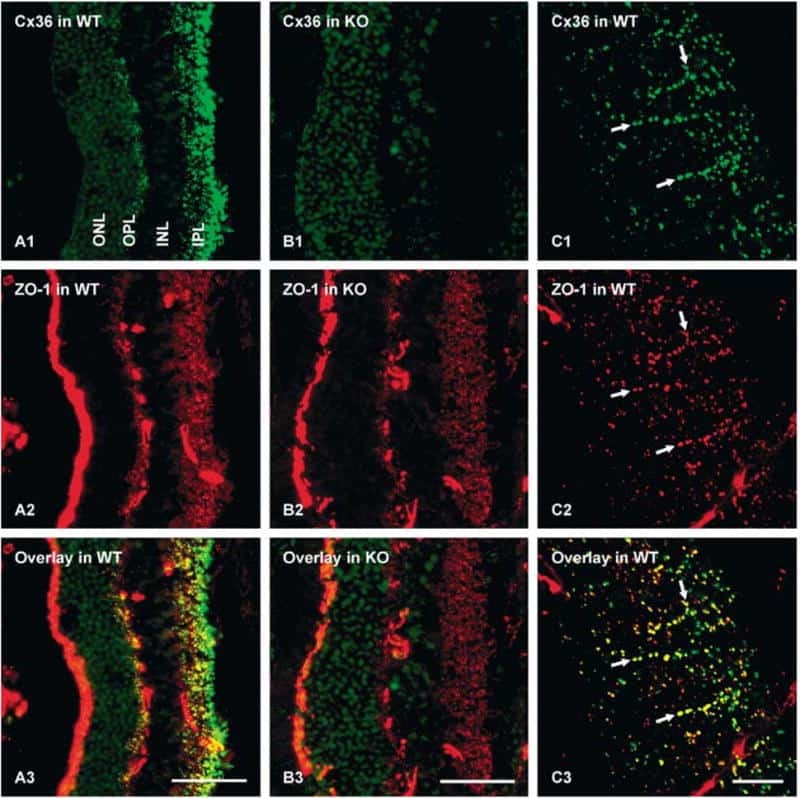
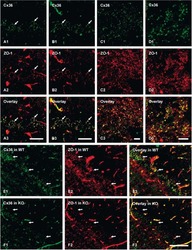
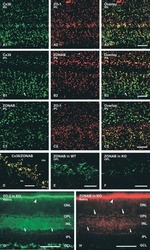
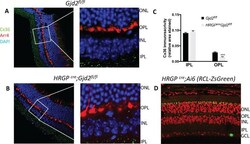
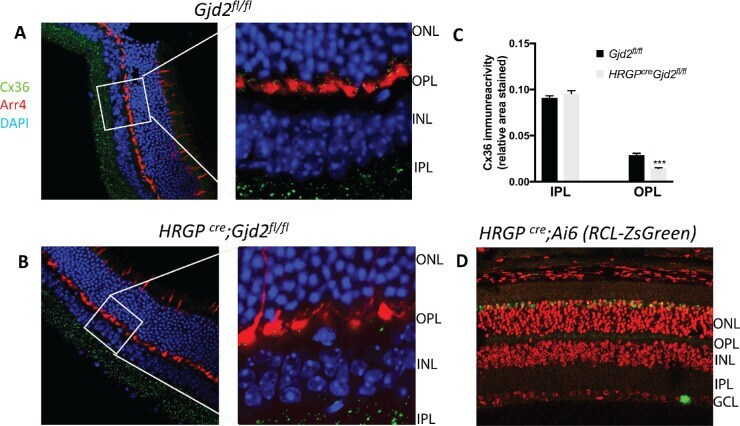
- Figure 1. Immunofluorescence staining of retinas showed selective disruption of Cx36 expression in cone photoreceptors in HRGP cre Gjd2 fl/fl . ( A, B ) Cx36 immunoreactive plaques were observed in the IPL and OPL of Gjd2 fl/fl and HRGP cre Gjd2 fl/fl mice (magnification: left 60x, right 240x); see black and white images in Supplementary Figure S1 . ( C ) Quantification of Cx36 immunoreactive plaques in retinas of HRGP cre Gjd2 fl/fl mice was lower compared to the littermate Gjd2 fl/fl control mice (*** P < 0.001, sample size: 6-7 mice / genotype). ( D ) Fluorescence was observed almost exclusively in cone photoreceptors in Ai6 (RCL-ZsGreen) Cre reporter mice. Retinal layers: ONL, outer nuclear layer; OPL, outer plexiform layer; INL, inner nuclear layer; IPL, inner plexiform layer; GCL, ganglion cell layer.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
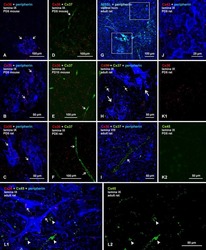
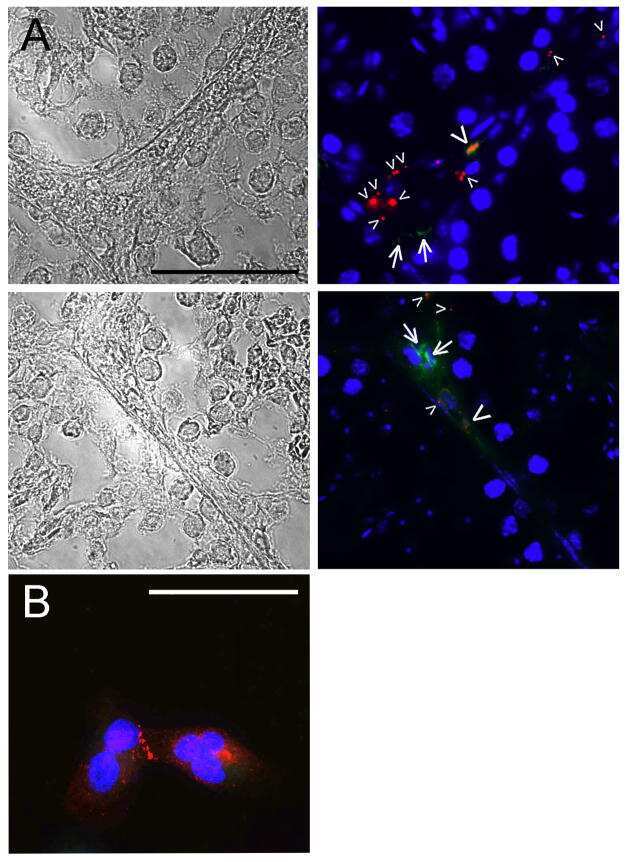
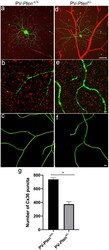
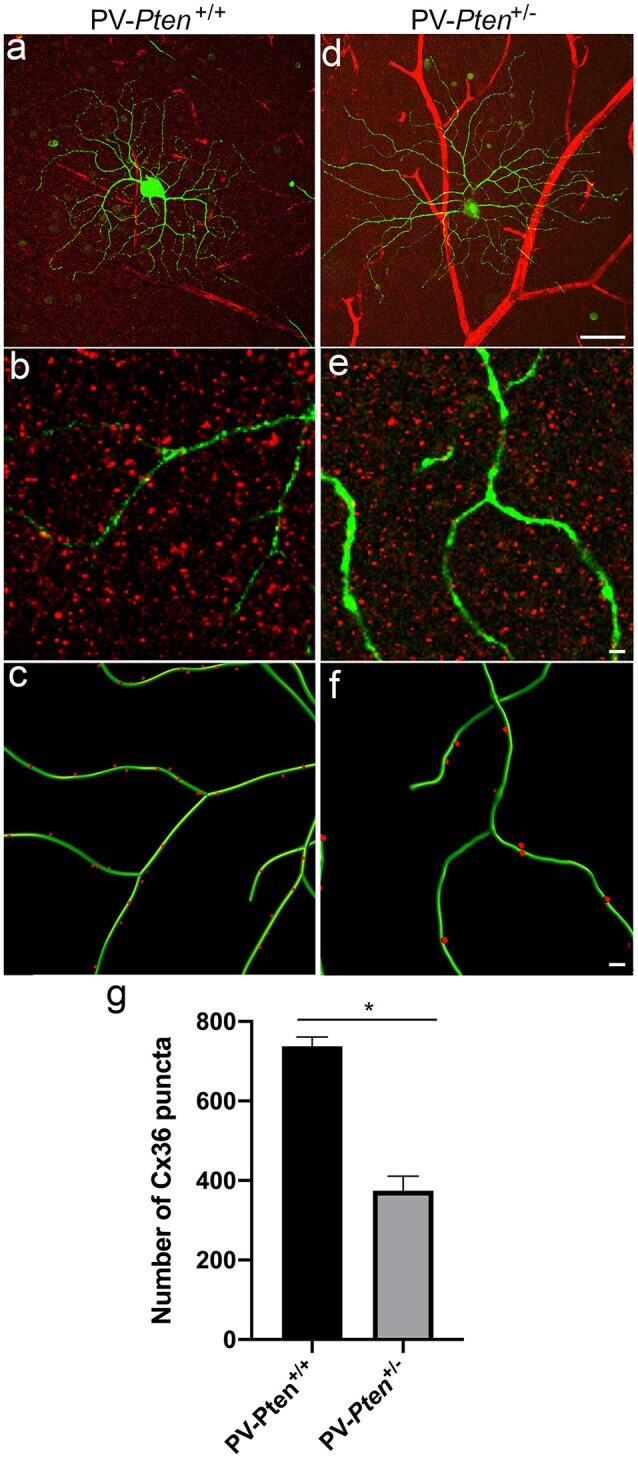
- Figure 8 Cx36 expression in OFF alpha-RGCs in PV- Pten +/+ and PV- Pten +/- retinas. (A) A maximum projection of a Neurobiotin-filled OFF alpha-RGC (green) stained with Cx36 antibodies (red) in a PV- Pten +/+ retina at 1.0 mm from the optic nerve head. (B) A single optical section of the Neurobiotin-filled OFF alpha-RGC (green) stained with Cx36 antibodies (red) in PV- Pten +/+ retina. (C) Imaris reconstruction of panel (B) . Cx36 puncta closer than 0.3 mum to the dendrites are represented in red. (D) A maximum projection of a Neurobiotin-filled OFF alpha-RGC (green) stained with Cx36 antibodies (red) in PV- Pten +/- retina at 1.0 mm from the optic nerve head. (E) A single optical section of the Neurobiotin-filled OFF alpha-RGC (green) stained with Cx36 antibodies (red) in PV- Pten +/- retina. (F) Imaris reconstruction of panel (E) . Cx36 puncta closer than 0.3 mum to the dendrites are represented in red. (G) Histogram indicating the mean (+- SEM) of Cx36 puncta in OFF alpha-RGCs ( n = 3 cells from PV- Pten +/+ retinas, n = 5 cells from PV- Pten +/- retinas, * P < 0.05, Mann-Whitney test). Scale bar in (A,D) = 50 mum. Scale bar in (B,C,E,F) = 2 mum.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
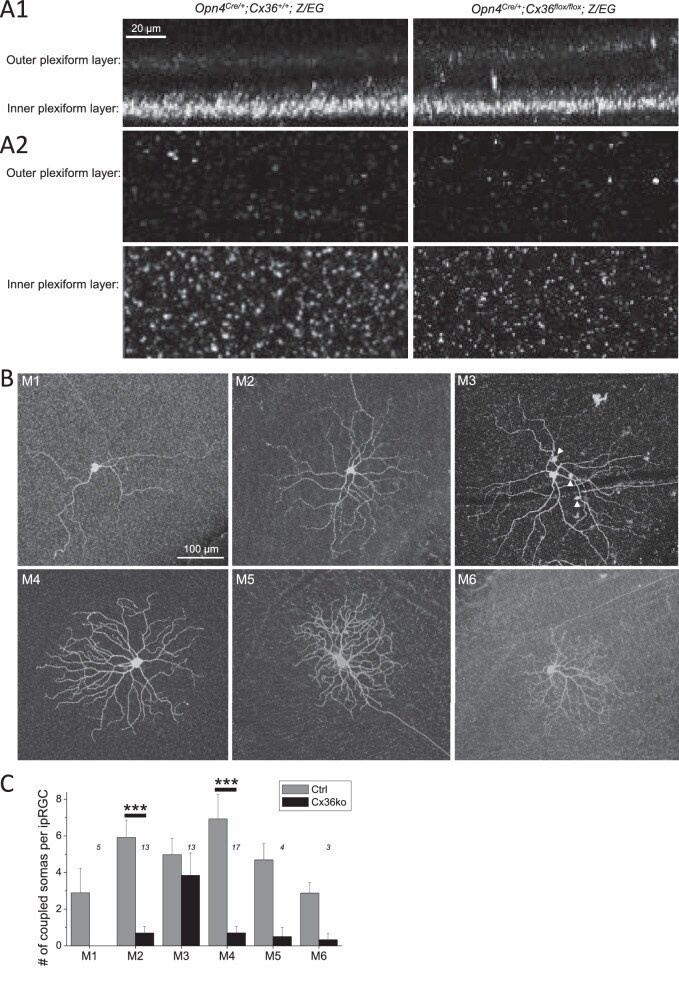
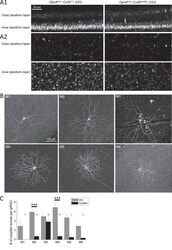
- Figure 5. ipRGC-amacrine coupling is mediated in part by Cx36. ( A ) Cx36 immunostaining confirms nonglobal Cx36 knockout in Opn4 Cre/+ ; Cx36 flox/ flox ; Z/EG retinas. A1 : Cx36 immunostaining was imaged confocally in whole-mount retinas, and the z-stacks were rotated 90deg to show these orthogonal views of Opn4 Cre/+ ; Cx36 +/+ ; Z/EG ( left ) and Opn4 Cre/+ ; Cx36 flox/ flox ; Z/EG ( right ) retinas. A2 : Representative whole-mount images at focal planes within the outer ( top ) and inner ( bottom ) plexiform layers. ( B ) Neurobiotin staining patterns of six representative Opn4 Cre/+ ; Cx36 flox/ flox ; Z/EG ipRGCs. Arrowheads mark Neurobiotin-filled somas within the M3 ipRGC's dendritic field. ( C ) Population-averaged numbers of somas coupled to each Opn4 Cre/+ ; Cx36 flox/ flox ; Z/EG ipRGC of every type, including uncoupled ipRGCs ( black columns ). The number above each column is the number of ipRGCs analyzed for that ipRGC type. The Opn4 Cre/+ ; Z/EG control data ( gray columns ) have been replotted from Figure 1 C. *** P < 0.001.