 Explore
Explore Validate
Validate Learn
Learn14-5983-82
antibody from Invitrogen Antibodies
Targeting: CD274
B7-H, B7-H1, B7H1, PD-L1, PDCD1LG1, PDL1
 Western blot
Western blot Immunohistochemistry
ImmunohistochemistryAntibody data
- Antibody Data
- Antigen structure
- References [102]
- Comments [0]
- Validations
- Western blot [1]
- Immunocytochemistry [1]
- Flow cytometry [1]
- Other assay [89]
Submit
Validation data
Reference
Comment
Report error
- Product number
- 14-5983-82 - Provider product page

- Provider
- Invitrogen Antibodies
- Product name
- CD274 (PD-L1, B7-H1) Monoclonal Antibody (MIH1), eBioscience™
- Antibody type
- Monoclonal
- Antigen
- Other
- Description
- Description: The MIH1 monoclonal antibody reacts with human B7-H1, also known as PD-L1. B7-H1, a member of the B7 family, has a predicted molecular weight of approximately 40 kDa and belongs to the Ig superfamily. B7-H1 is expressed on a majority of leukocytes. B7-H1 is a ligand for PD-1. Interaction of PD-1 with either PD-L1 (B7-H1) or PD-L2 (B7-DC) results in inhibition of T and B cell responses. MIH1 is reported to be a blocking antibody. Applications Reported: The MIH1 antibody has been reported for use in flow cytometric analysis, and immunohistochemical staining of frozen tissue sections. It has also been reported in blocking in in vitro functional assays. (Please use Functional Grade purified MIH1, Product # 16-5983, in functional assays). Applications Tested: The MIH1 antibody has been tested by flow cytometric analysis of normal human peripheral blood cells. This can be used at less than or equal to 1 µg per test. A test is defined as the amount (µg) of antibody that will stain a cell sample in a final volume of 100 µL. Cell number should be determined empirically but can range from 10^5 to 10^8 cells/test. It is recommended that the antibody be carefully titrated for optimal performance in the assay of interest. Purity: Greater than 90%, as determined by SDS-PAGE. Aggregation: Less than 10%, as determined by HPLC. Filtration: 0.2 µm post-manufacturing filtered.
- Reactivity
- Human
- Host
- Mouse
- Isotype
- IgG
- Antibody clone number
- MIH1
- Vial size
- 100 µg
- Concentration
- 0.5 mg/mL
- Storage
- 4° C
Submitted references Active PD-L1 incorporation within HIV virions functionally impairs T follicular helper cells.
Deciphering the role of miR-187-3p/LRFN1 axis in modulating progression, aerobic glycolysis and immune microenvironment of clear cell renal cell carcinoma.
PD-L1 CAR effector cells induce self-amplifying cytotoxic effects against target cells.
Different Induction of PD-L1 (CD274) and PD-1 (CD279) Expression in THP-1-Differentiated Types 1 and 2 Macrophages.
Overexpression of PD-L1 is an Independent Predictor for Recurrence in HCC Patients Who Receive Sorafenib Treatment After Surgical Resection.
Microbe-Mediated Activation of Toll-like Receptor 2 Drives PDL1 Expression in HNSCC.
Concomitant High Apoptosis Inhibitor of Macrophage (AIM) and Low Prostate-Specific Antigen (PSA) Indicates Activated T Cell-Mediated Anticancer Immunity, Enhance Sensitivity to Pembrolizumab, and Elicit Good Prognosis in Prostate Cancer.
PD-1 independent of PD-L1 ligation promotes glioblastoma growth through the NFκB pathway.
Protocol for high-throughput compound screening using flow cytometry in THP-1 cells.
NEK2 inhibition triggers anti-pancreatic cancer immunity by targeting PD-L1.
Ruxolitinib reverses checkpoint inhibition by reducing programmed cell death ligand-1 (PD-L1) expression and increases anti-tumour effects of T cells in multiple myeloma.
IL-27 Mediates PD-L1 Expression and Release by Human Mesothelioma Cells.
β-Catenin induces transcriptional expression of PD-L1 to promote glioblastoma immune evasion.
TASL is the SLC15A4-associated adaptor for IRF5 activation by TLR7-9.
Cancer cell-intrinsic expression of MHC II in lung cancer cell lines is actively restricted by MEK/ERK signaling and epigenetic mechanisms.
Protein kinase D3 regulates the expression of the immunosuppressive protein, PD‑L1, through STAT1/STAT3 signaling.
TLR9 Mediated Tumor-Stroma Interactions in Human Papilloma Virus (HPV)-Positive Head and Neck Squamous Cell Carcinoma Up-Regulate PD-L1 and PD-L2.
PD-L1 checkpoint blockade delivered by retroviral replicating vector confers anti-tumor efficacy in murine tumor models.
Phospho‑STAT1 expression as a potential biomarker for anti‑PD‑1/anti‑PD‑L1 immunotherapy for breast cancer.
Low frequency, weak MCP-1 secretion and exhausted immune status of peripheral monocytes were associated with progression of severe enterovirus A71-infected hand, foot and mouth disease.
Quantitative Interactomics in Primary T Cells Provides a Rationale for Concomitant PD-1 and BTLA Coinhibitor Blockade in Cancer Immunotherapy.
High PD-L1 Expression Predicts for Worse Outcome of Leukemia Patients with Concomitant NPM1 and FLT3 Mutations.
PD-L1:CD80 Cis-Heterodimer Triggers the Co-stimulatory Receptor CD28 While Repressing the Inhibitory PD-1 and CTLA-4 Pathways.
Immune Heterogeneity Between Primary Tumors and Corresponding Metastatic Lesions and Response to Platinum Therapy in Primary Ovarian Cancer.
CA-170 - A Potent Small-Molecule PD-L1 Inhibitor or Not?
Suppression of Exosomal PD-L1 Induces Systemic Anti-tumor Immunity and Memory.
Exhaustion of the CD8(+) T Cell Compartment in Patients with Mutations in Phosphoinositide 3-Kinase Delta.
Disruption of PD-1 Enhanced the Anti-tumor Activity of Chimeric Antigen Receptor T Cells Against Hepatocellular Carcinoma.
Generation of Tumor-Reactive T Cells by Co-culture of Peripheral Blood Lymphocytes and Tumor Organoids.
Apigenin suppresses PD-L1 expression in melanoma and host dendritic cells to elicit synergistic therapeutic effects.
Specific expression of PD-L1 in RELA-fusion supratentorial ependymoma: Implications for PD-1-targeted therapy.
ATM‑JAK‑PD‑L1 signaling pathway inhibition decreases EMT and metastasis of androgen‑independent prostate cancer.
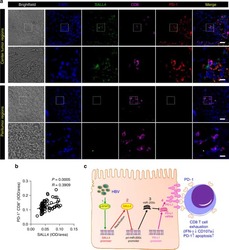
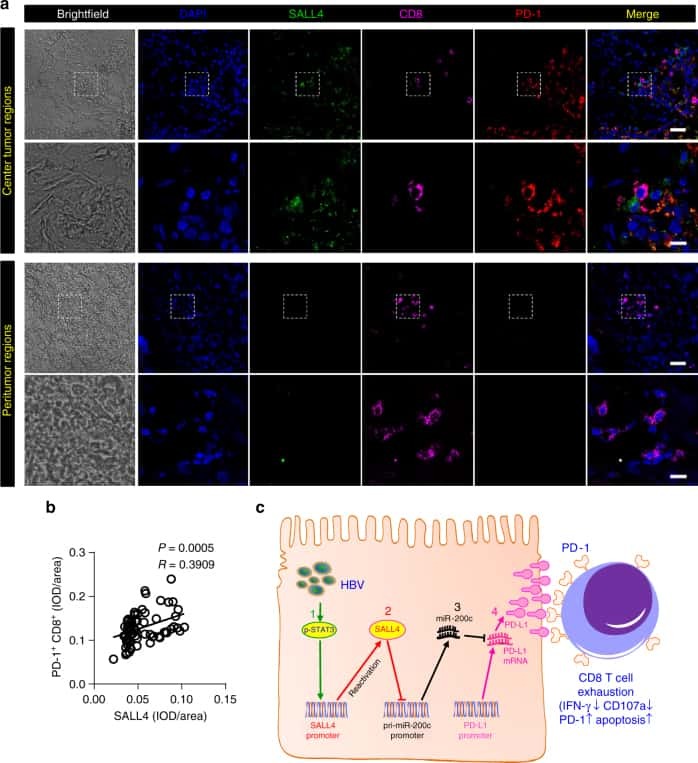
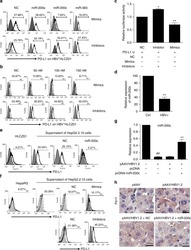
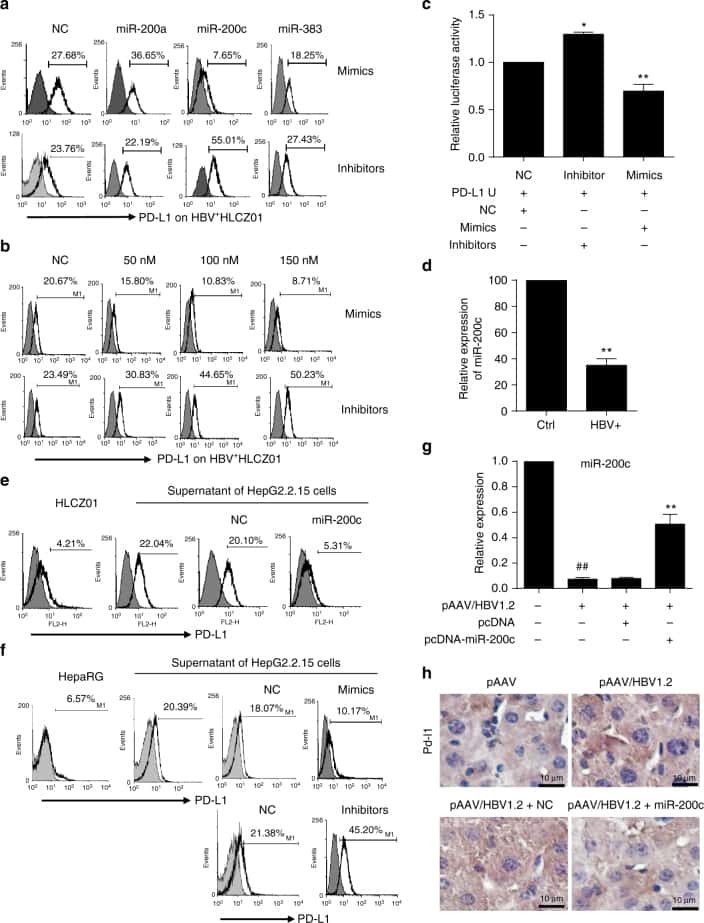
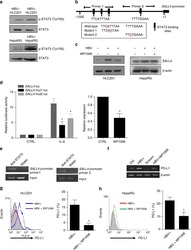
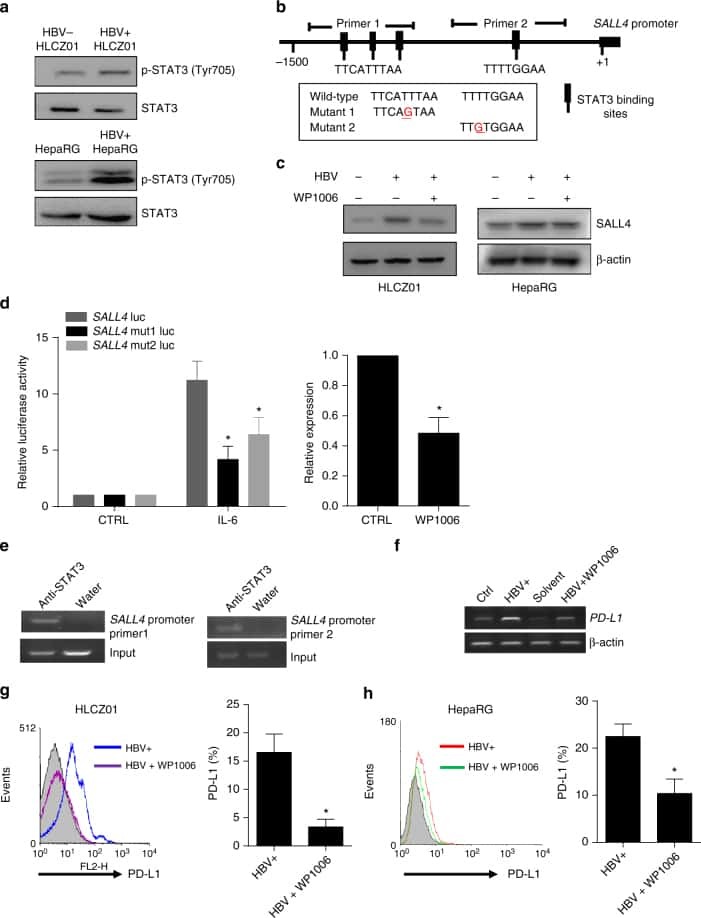
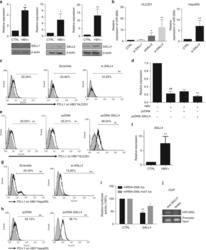
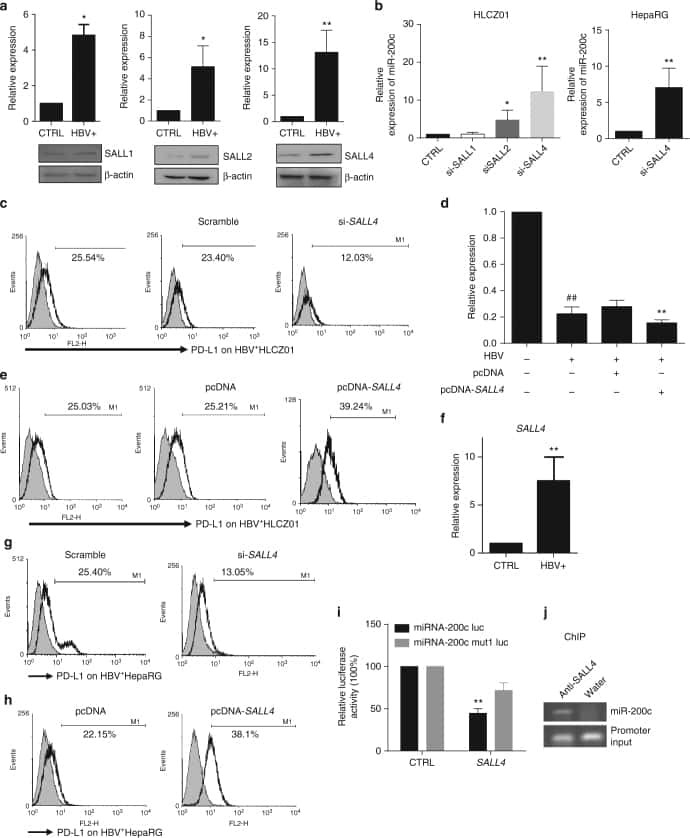
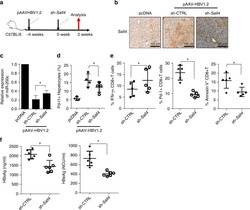
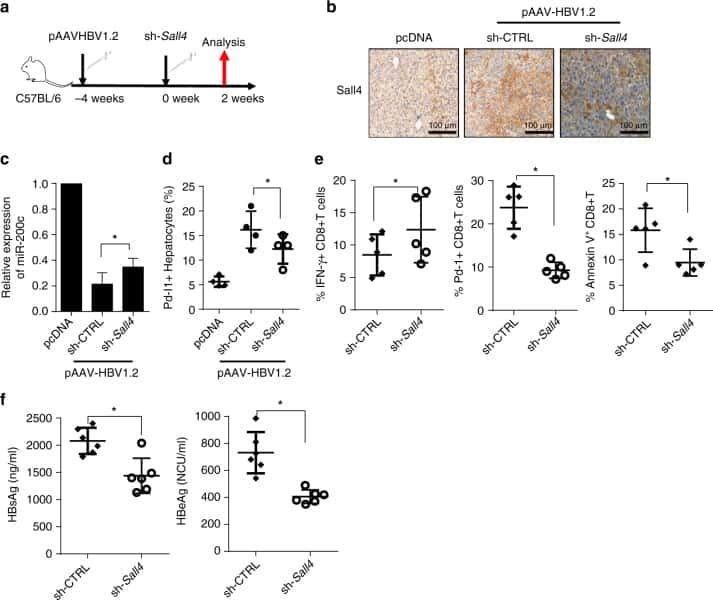
Oncofetal gene SALL4 reactivation by hepatitis B virus counteracts miR-200c in PD-L1-induced T cell exhaustion.
Dominant Role for Regulatory T Cells in Protecting Females Against Pulmonary Hypertension.
A versatile T cell-based assay to assess therapeutic antigen-specific PD-1-targeted approaches.
Antigen-Presenting Cell-Intrinsic PD-1 Neutralizes PD-L1 in cis to Attenuate PD-1 Signaling in T Cells.
PD-1 axis expression in musculoskeletal tumors and antitumor effect of nivolumab in osteosarcoma model of humanized mouse.
Similarities and differences between helminth parasites and cancer cell lines in shaping human monocytes: Insights into parallel mechanisms of immune evasion.
Amphotericin B suppresses M2 phenotypes and B7-H1 expression in macrophages to prevent Raji cell proliferation.
PD-L1 expression in medulloblastoma: an evaluation by subgroup.
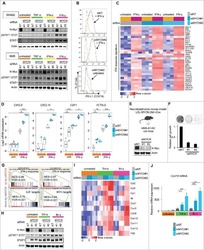
Amplification of N-Myc is associated with a T-cell-poor microenvironment in metastatic neuroblastoma restraining interferon pathway activity and chemokine expression.
Idelalisib and caffeine reduce suppression of T cell responses mediated by activated chronic lymphocytic leukemia cells.
Mesenchymal Stromal Cell Secretion of Programmed Death-1 Ligands Regulates T Cell Mediated Immunosuppression.
Structure-guided development of a high-affinity human Programmed Cell Death-1: Implications for tumor immunotherapy.
Retroviral Replicating Vector Delivery of miR-PDL1 Inhibits Immune Checkpoint PDL1 and Enhances Immune Responses In Vitro.
Characterization and Purification of Neoplastic Cells of Nodular Lymphocyte Predominant Hodgkin Lymphoma from Lymph Nodes by Flow Cytometry and Flow Cytometric Cell Sorting.
Oncogenic RAS Signaling Promotes Tumor Immunoresistance by Stabilizing PD-L1 mRNA.
Hormonal vitamin D up-regulates tissue-specific PD-L1 and PD-L2 surface glycoprotein expression in humans but not mice.
PD-L1/PD-1 expression and tumor-infiltrating lymphocytes in conjunctival melanoma.
PD-L1 expression in human cancers and its association with clinical outcomes.
Tolerogenic IDO(+) Dendritic Cells Are Induced by PD-1-Expressing Mast Cells.
Ly6C(hi) monocytes regulate T cell responses in viral hepatitis.
Regulatory IgDhi B Cells Suppress T Cell Function via IL-10 and PD-L1 during Progressive Visceral Leishmaniasis.
Expression of programmed death 1 ligand 1 on periodontal tissue cells as a possible protective feedback mechanism against periodontal tissue destruction.
CD28 family of receptors on T cells in chronic HBV infection: Expression characteristics, clinical significance and correlations with PD-1 blockade.
Psoriasis is characterized by deficient negative immune regulation compared to transient delayed-type hypersensitivity reactions.
Upregulation of Programmed Death-1 and Its Ligand in Cardiac Injury Models: Interaction with GADD153.
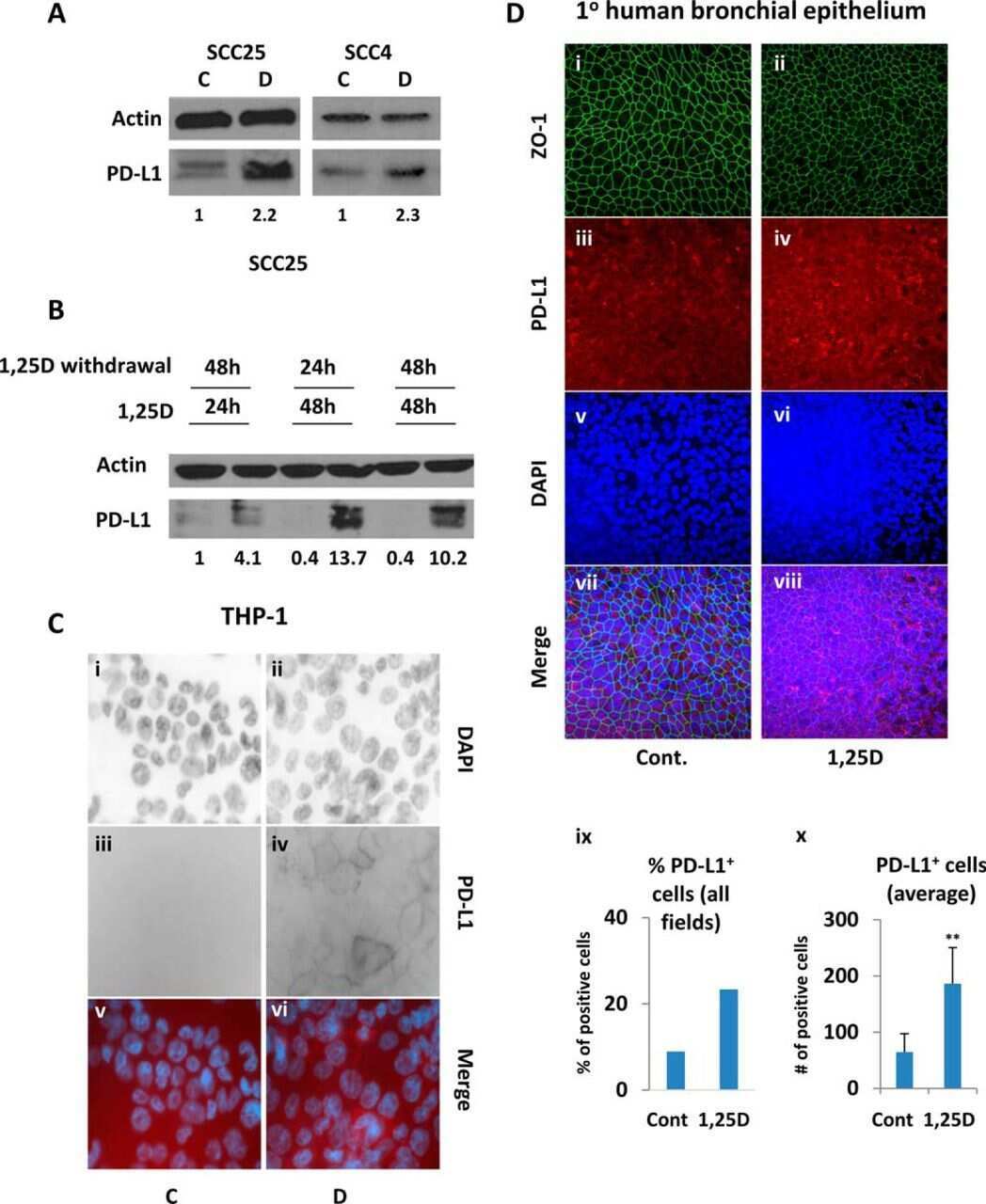
Interleukin-25 Mediates Transcriptional Control of PD-L1 via STAT3 in Multipotent Human Mesenchymal Stromal Cells (hMSCs) to Suppress Th17 Responses.
Programmed death ligand 1 on Burkholderia pseudomallei-infected human polymorphonuclear neutrophils impairs T cell functions.
Assays for predicting and monitoring responses to lung cancer immunotherapy.
Control of signaling-mediated clearance of apoptotic cells by the tumor suppressor p53.
Paucity of PD-L1 expression in prostate cancer: innate and adaptive immune resistance.
IL-27 induces the expression of IDO and PD-L1 in human cancer cells.
Inhibition of mitogen-activated protein kinase pathway can induce upregulation of human leukocyte antigen class I without PD-L1-upregulation in contrast to interferon-γ treatment.
Immune-escape markers in relation to clinical outcome of advanced melanoma patients following immunotherapy.
Galectin-9 and IL-21 mediate cross-regulation between Th17 and Treg cells during acute hepatitis C.
NF-κB plays a key role in inducing CD274 expression in human monocytes after lipopolysaccharide treatment.
Soluble CD80 restores T cell activation and overcomes tumor cell programmed death ligand 1-mediated immune suppression.
Alterations of immune response of Non-Small Cell Lung Cancer with Azacytidine.
Antagonist antibodies to PD-1 and B7-H1 (PD-L1) in the treatment of advanced human cancer.
Programmed death 1-mediated T cell exhaustion during visceral leishmaniasis impairs phagocyte function.
Inhibiting the programmed death 1 pathway rescues Mycobacterium tuberculosis-specific interferon γ-producing T cells from apoptosis in patients with pulmonary tuberculosis.
Foxp3+ regulatory T cells among tuberculosis patients: impact on prognosis and restoration of antigen specific IFN-γ producing T cells.
Tissue biomarkers in melanoma patients treated with TIL.
TNF-α and TGF-β counter-regulate PD-L1 expression on monocytes in systemic lupus erythematosus.
Upregulation of programmed death-1 on T cells and programmed death ligand-1 on monocytes in septic shock patients.
HIV-mediated phosphatidylinositol 3-kinase/serine-threonine kinase activation in APCs leads to programmed death-1 ligand upregulation and suppression of HIV-specific CD8 T cells.
Activated memory B cells may function as antigen-presenting cells in the joints of children with juvenile idiopathic arthritis.
Tumor cell programmed death ligand 1-mediated T cell suppression is overcome by coexpression of CD80.
Immunostaining of PD-1/PD-Ls in liver tissues of patients with hepatitis and hepatocellular carcinoma.
Heat shock protein 27 differentiates tolerogenic macrophages that may support human breast cancer progression.
Ex vivo expanded hematopoietic stem cells overcome the MHC barrier in allogeneic transplantation.
Doxorubicin downregulates cell surface B7-H1 expression and upregulates its nuclear expression in breast cancer cells: role of B7-H1 as an anti-apoptotic molecule.
The PD-1/PD-L1 axis modulates the natural killer cell versus multiple myeloma effect: a therapeutic target for CT-011, a novel monoclonal anti-PD-1 antibody.
Mechanisms of immune suppression for CD8+ T cells by human corneal endothelial cells via membrane-bound TGFbeta.
Role played by the programmed death-1-programmed death ligand pathway during innate immunity against Mycobacterium tuberculosis.
Tumor-infiltrating NY-ESO-1-specific CD8+ T cells are negatively regulated by LAG-3 and PD-1 in human ovarian cancer.
B7-H1 (PD-L1, CD274) suppresses host immunity in T-cell lymphoproliferative disorders.
Costimulatory pathways in multiple sclerosis: distinctive expression of PD-1 and PD-L1 in patients with different patterns of disease.
PD-1 is a regulator of NY-ESO-1-specific CD8+ T cell expansion in melanoma patients.
Activated monocytes in peritumoral stroma of hepatocellular carcinoma foster immune privilege and disease progression through PD-L1.
Human corneal endothelial cells expressing programmed death-ligand 1 (PD-L1) suppress PD-1+ T helper 1 cells by a contact-dependent mechanism.
PD-1/PD-L1 expression in human T-cell leukemia virus type 1 carriers and adult T-cell leukemia/lymphoma patients.
MicroRNA-513 regulates B7-H1 translation and is involved in IFN-gamma-induced B7-H1 expression in cholangiocytes.
Interaction of human PD-L1 and B7-1.
Expression of B7-H1 in inflammatory renal tubular epithelial cells.
The B7-H1 (PD-L1) T lymphocyte-inhibitory molecule is expressed in breast cancer patients with infiltrating ductal carcinoma: correlation with important high-risk prognostic factors.
NK cells that are activated by CXCL10 can kill dormant tumor cells that resist CTL-mediated lysis and can express B7-H1 that stimulates T cells.
Clinical significance of programmed death-1 ligand-1 and programmed death-1 ligand-2 expression in human esophageal cancer.
Blockade of B7-H1 suppresses the development of chronic intestinal inflammation.
Engagement of the PD-1 immunoinhibitory receptor by a novel B7 family member leads to negative regulation of lymphocyte activation.
B7-H1, a third member of the B7 family, co-stimulates T-cell proliferation and interleukin-10 secretion.
Munoz O, Banga R, Schelling R, Procopio FA, Mastrangelo A, Nortier P, Ohmiti K, Daraspe J, Cavassini M, Fenwick C, Perez L, Perreau M
PLoS pathogens 2022 Jul;18(7):e1010673
PLoS pathogens 2022 Jul;18(7):e1010673
Deciphering the role of miR-187-3p/LRFN1 axis in modulating progression, aerobic glycolysis and immune microenvironment of clear cell renal cell carcinoma.
Xu W, Liu W, Anwaier A, Tian X, Su J, Shi G, Wei S, Qu Y, Zhang H, Ye D
Discover. Oncology 2022 Jul 7;13(1):59
Discover. Oncology 2022 Jul 7;13(1):59
PD-L1 CAR effector cells induce self-amplifying cytotoxic effects against target cells.
Bajor M, Graczyk-Jarzynka A, Marhelava K, Burdzinska A, Muchowicz A, Goral A, Zhylko A, Soroczynska K, Retecki K, Krawczyk M, Klopotowska M, Pilch Z, Paczek L, Malmberg KJ, Wälchli S, Winiarska M, Zagozdzon R
Journal for immunotherapy of cancer 2022 Jan;10(1)
Journal for immunotherapy of cancer 2022 Jan;10(1)
Different Induction of PD-L1 (CD274) and PD-1 (CD279) Expression in THP-1-Differentiated Types 1 and 2 Macrophages.
Lai CY, Tseng PC, Chen CL, Satria RD, Wang YT, Lin CF
Journal of inflammation research 2021;14:5241-5249
Journal of inflammation research 2021;14:5241-5249
Overexpression of PD-L1 is an Independent Predictor for Recurrence in HCC Patients Who Receive Sorafenib Treatment After Surgical Resection.
Tan Y, Xu Q, Wu Z, Zhang W, Li B, Zhang B, Xu X, Zhang B, Yan K, Song J, Lv T, Yang J, Jiang L, Shi Y, Yang J, Yan L
Frontiers in oncology 2021;11:783335
Frontiers in oncology 2021;11:783335
Microbe-Mediated Activation of Toll-like Receptor 2 Drives PDL1 Expression in HNSCC.
Mann JE, Ludwig ML, Kulkarni A, Scheftz EB, Murray IR, Zhai J, Gensterblum-Miller E, Jiang H, Brenner JC
Cancers 2021 Sep 24;13(19)
Cancers 2021 Sep 24;13(19)
Concomitant High Apoptosis Inhibitor of Macrophage (AIM) and Low Prostate-Specific Antigen (PSA) Indicates Activated T Cell-Mediated Anticancer Immunity, Enhance Sensitivity to Pembrolizumab, and Elicit Good Prognosis in Prostate Cancer.
Bamodu OA, Wang YH, Yeh CT, Ho CH, Chiang YT, Kao WT, Liu CH, Wu CC
Biomedicines 2021 Sep 15;9(9)
Biomedicines 2021 Sep 15;9(9)
PD-1 independent of PD-L1 ligation promotes glioblastoma growth through the NFκB pathway.
Mirzaei R, Gordon A, Zemp FJ, Kumar M, Sarkar S, Luchman HA, Bellail AC, Hao C, Mahoney DJ, Dunn JF, Bose P, Yong VW
Science advances 2021 Nov 5;7(45):eabh2148
Science advances 2021 Nov 5;7(45):eabh2148
Protocol for high-throughput compound screening using flow cytometry in THP-1 cells.
Spangenberg SH, Zavareh RB, Lairson LL
STAR protocols 2021 Jun 18;2(2):100400
STAR protocols 2021 Jun 18;2(2):100400
NEK2 inhibition triggers anti-pancreatic cancer immunity by targeting PD-L1.
Zhang X, Huang X, Xu J, Li E, Lao M, Tang T, Zhang G, Guo C, Zhang X, Chen W, Yadav DK, Bai X, Liang T
Nature communications 2021 Jul 27;12(1):4536
Nature communications 2021 Jul 27;12(1):4536
Ruxolitinib reverses checkpoint inhibition by reducing programmed cell death ligand-1 (PD-L1) expression and increases anti-tumour effects of T cells in multiple myeloma.
Chen H, Li M, Ng N, Yu E, Bujarski S, Yin Z, Wen M, Hekmati T, Field D, Wang J, Nassir I, Yu J, Huang J, Daniely D, Wang CS, Xu N, Spektor TM, Berenson JR
British journal of haematology 2021 Feb;192(3):568-576
British journal of haematology 2021 Feb;192(3):568-576
IL-27 Mediates PD-L1 Expression and Release by Human Mesothelioma Cells.
Carbotti G, Dozin B, Martini S, Giordano C, Scordamaglia F, Croce M, Filaci G, Ferrini S, Fabbi M
Cancers 2021 Aug 9;13(16)
Cancers 2021 Aug 9;13(16)
β-Catenin induces transcriptional expression of PD-L1 to promote glioblastoma immune evasion.
Du L, Lee JH, Jiang H, Wang C, Wang S, Zheng Z, Shao F, Xu D, Xia Y, Li J, Zheng Y, Qian X, Li X, Kim HR, Xing D, Liu P, Lu Z, Lyu J
The Journal of experimental medicine 2020 Nov 2;217(11)
The Journal of experimental medicine 2020 Nov 2;217(11)
TASL is the SLC15A4-associated adaptor for IRF5 activation by TLR7-9.
Heinz LX, Lee J, Kapoor U, Kartnig F, Sedlyarov V, Papakostas K, César-Razquin A, Essletzbichler P, Goldmann U, Stefanovic A, Bigenzahn JW, Scorzoni S, Pizzagalli MD, Bensimon A, Müller AC, King FJ, Li J, Girardi E, Mbow ML, Whitehurst CE, Rebsamen M, Superti-Furga G
Nature 2020 May;581(7808):316-322
Nature 2020 May;581(7808):316-322
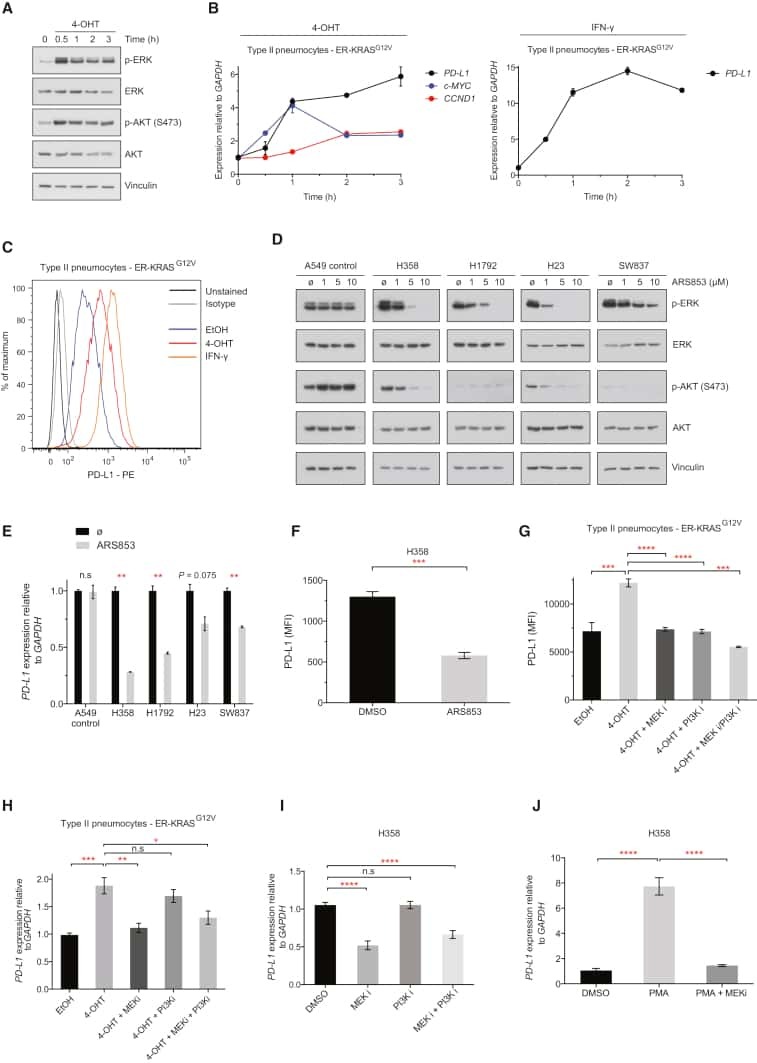
Cancer cell-intrinsic expression of MHC II in lung cancer cell lines is actively restricted by MEK/ERK signaling and epigenetic mechanisms.
Neuwelt AJ, Kimball AK, Johnson AM, Arnold BW, Bullock BL, Kaspar RE, Kleczko EK, Kwak JW, Wu MH, Heasley LE, Doebele RC, Li HY, Nemenoff RA, Clambey ET
Journal for immunotherapy of cancer 2020 Apr;8(1)
Journal for immunotherapy of cancer 2020 Apr;8(1)
Protein kinase D3 regulates the expression of the immunosuppressive protein, PD‑L1, through STAT1/STAT3 signaling.
Cui B, Chen J, Luo M, Wang L, Chen H, Kang Y, Wang J, Zhou X, Feng Y, Zhang P
International journal of oncology 2020 Apr;56(4):909-920
International journal of oncology 2020 Apr;56(4):909-920
TLR9 Mediated Tumor-Stroma Interactions in Human Papilloma Virus (HPV)-Positive Head and Neck Squamous Cell Carcinoma Up-Regulate PD-L1 and PD-L2.
Baruah P, Bullenkamp J, Wilson POG, Lee M, Kaski JC, Dumitriu IE
Frontiers in immunology 2019;10:1644
Frontiers in immunology 2019;10:1644
PD-L1 checkpoint blockade delivered by retroviral replicating vector confers anti-tumor efficacy in murine tumor models.
Mitchell LA, Yagiz K, Hofacre A, Viaud S, Munday AW, Espinoza FL, Mendoza D, Rodriguez-Aguirre ME, Bergqvist S, Haghighi A, Miner MV, Accomando WP, Burrascano C, Gammon D, Gruber HE, Jolly DJ, Lin AH
Oncotarget 2019 Mar 19;10(23):2252-2269
Oncotarget 2019 Mar 19;10(23):2252-2269
Phospho‑STAT1 expression as a potential biomarker for anti‑PD‑1/anti‑PD‑L1 immunotherapy for breast cancer.
Nakayama Y, Mimura K, Tamaki T, Shiraishi K, Kua LF, Koh V, Ohmori M, Kimura A, Inoue S, Okayama H, Suzuki Y, Nakazawa T, Ichikawa D, Kono K
International journal of oncology 2019 Jun;54(6):2030-2038
International journal of oncology 2019 Jun;54(6):2030-2038
Low frequency, weak MCP-1 secretion and exhausted immune status of peripheral monocytes were associated with progression of severe enterovirus A71-infected hand, foot and mouth disease.
Pei X, Fan X, Zhang H, Duan H, Xu C, Xie B, Wang L, Li X, Peng Y, Shen T
Clinical and experimental immunology 2019 Jun;196(3):353-363
Clinical and experimental immunology 2019 Jun;196(3):353-363
Quantitative Interactomics in Primary T Cells Provides a Rationale for Concomitant PD-1 and BTLA Coinhibitor Blockade in Cancer Immunotherapy.
Celis-Gutierrez J, Blattmann P, Zhai Y, Jarmuzynski N, Ruminski K, Grégoire C, Ounoughene Y, Fiore F, Aebersold R, Roncagalli R, Gstaiger M, Malissen B
Cell reports 2019 Jun 11;27(11):3315-3330.e7
Cell reports 2019 Jun 11;27(11):3315-3330.e7
High PD-L1 Expression Predicts for Worse Outcome of Leukemia Patients with Concomitant NPM1 and FLT3 Mutations.
Brodská B, Otevřelová P, Šálek C, Fuchs O, Gašová Z, Kuželová K
International journal of molecular sciences 2019 Jun 10;20(11)
International journal of molecular sciences 2019 Jun 10;20(11)
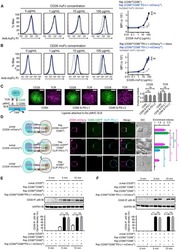
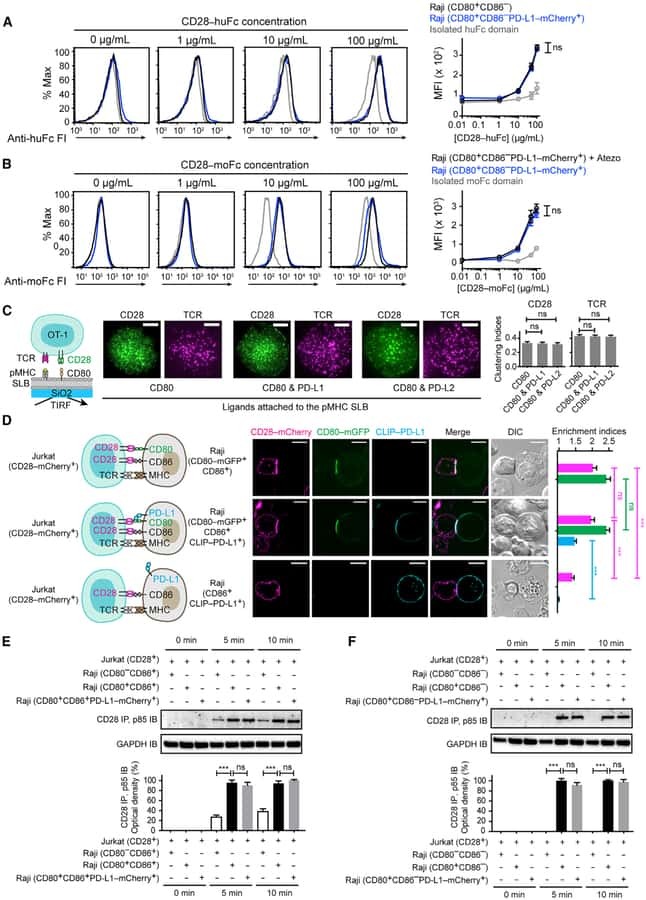
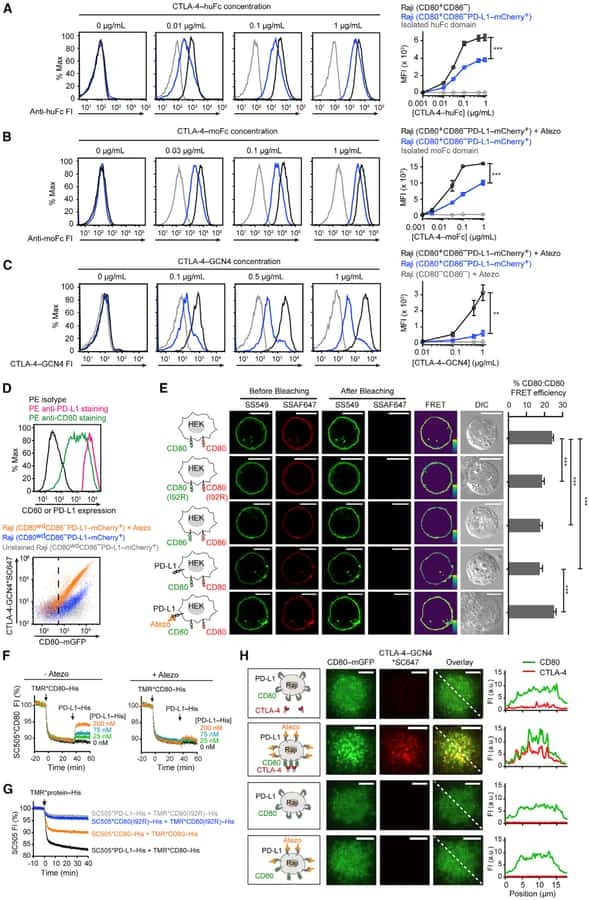
PD-L1:CD80 Cis-Heterodimer Triggers the Co-stimulatory Receptor CD28 While Repressing the Inhibitory PD-1 and CTLA-4 Pathways.
Zhao Y, Lee CK, Lin CH, Gassen RB, Xu X, Huang Z, Xiao C, Bonorino C, Lu LF, Bui JD, Hui E
Immunity 2019 Dec 17;51(6):1059-1073.e9
Immunity 2019 Dec 17;51(6):1059-1073.e9
Immune Heterogeneity Between Primary Tumors and Corresponding Metastatic Lesions and Response to Platinum Therapy in Primary Ovarian Cancer.
Dötzer K, Schlüter F, Schoenberg MB, Bazhin AV, von Koch FE, Schnelzer A, Anthuber S, Grab D, Czogalla B, Burges A, Werner J, Mahner S, Mayer B
Cancers 2019 Aug 26;11(9)
Cancers 2019 Aug 26;11(9)
CA-170 - A Potent Small-Molecule PD-L1 Inhibitor or Not?
Musielak B, Kocik J, Skalniak L, Magiera-Mularz K, Sala D, Czub M, Stec M, Siedlar M, Holak TA, Plewka J
Molecules (Basel, Switzerland) 2019 Aug 1;24(15)
Molecules (Basel, Switzerland) 2019 Aug 1;24(15)
Suppression of Exosomal PD-L1 Induces Systemic Anti-tumor Immunity and Memory.
Poggio M, Hu T, Pai CC, Chu B, Belair CD, Chang A, Montabana E, Lang UE, Fu Q, Fong L, Blelloch R
Cell 2019 Apr 4;177(2):414-427.e13
Cell 2019 Apr 4;177(2):414-427.e13
Exhaustion of the CD8(+) T Cell Compartment in Patients with Mutations in Phosphoinositide 3-Kinase Delta.
Wentink MWJ, Mueller YM, Dalm VASH, Driessen GJ, van Hagen PM, van Montfrans JM, van der Burg M, Katsikis PD
Frontiers in immunology 2018;9:446
Frontiers in immunology 2018;9:446
Disruption of PD-1 Enhanced the Anti-tumor Activity of Chimeric Antigen Receptor T Cells Against Hepatocellular Carcinoma.
Guo X, Jiang H, Shi B, Zhou M, Zhang H, Shi Z, Du G, Luo H, Wu X, Wang Y, Sun R, Li Z
Frontiers in pharmacology 2018;9:1118
Frontiers in pharmacology 2018;9:1118
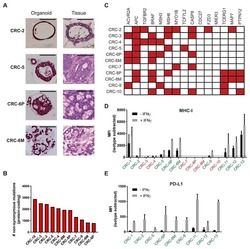
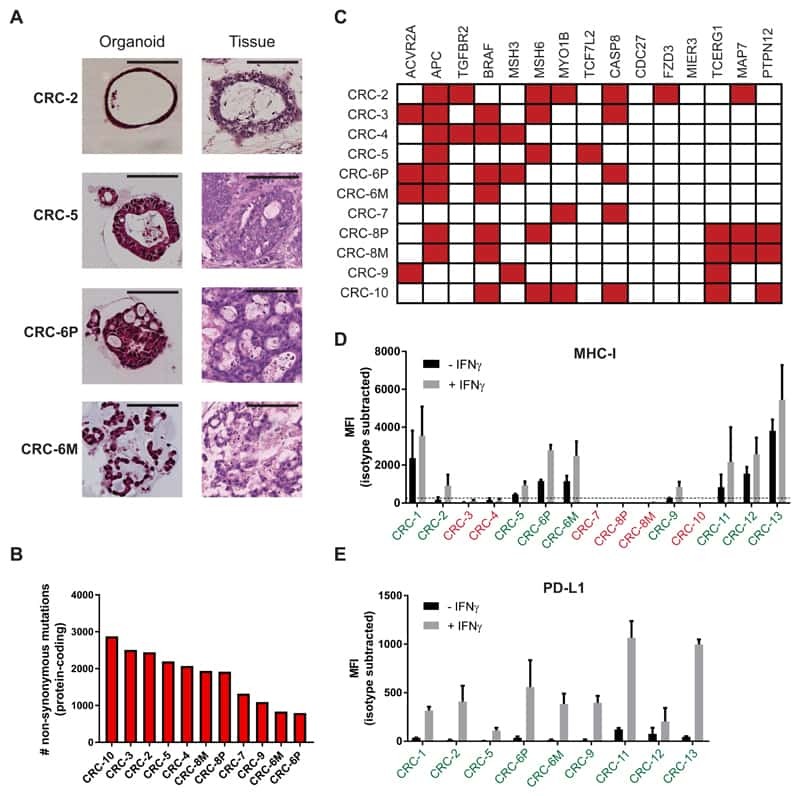
Generation of Tumor-Reactive T Cells by Co-culture of Peripheral Blood Lymphocytes and Tumor Organoids.
Dijkstra KK, Cattaneo CM, Weeber F, Chalabi M, van de Haar J, Fanchi LF, Slagter M, van der Velden DL, Kaing S, Kelderman S, van Rooij N, van Leerdam ME, Depla A, Smit EF, Hartemink KJ, de Groot R, Wolkers MC, Sachs N, Snaebjornsson P, Monkhorst K, Haanen J, Clevers H, Schumacher TN, Voest EE
Cell 2018 Sep 6;174(6):1586-1598.e12
Cell 2018 Sep 6;174(6):1586-1598.e12
Apigenin suppresses PD-L1 expression in melanoma and host dendritic cells to elicit synergistic therapeutic effects.
Xu L, Zhang Y, Tian K, Chen X, Zhang R, Mu X, Wu Y, Wang D, Wang S, Liu F, Wang T, Zhang J, Liu S, Zhang Y, Tu C, Liu H
Journal of experimental & clinical cancer research : CR 2018 Oct 29;37(1):261
Journal of experimental & clinical cancer research : CR 2018 Oct 29;37(1):261
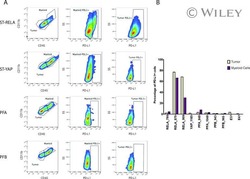
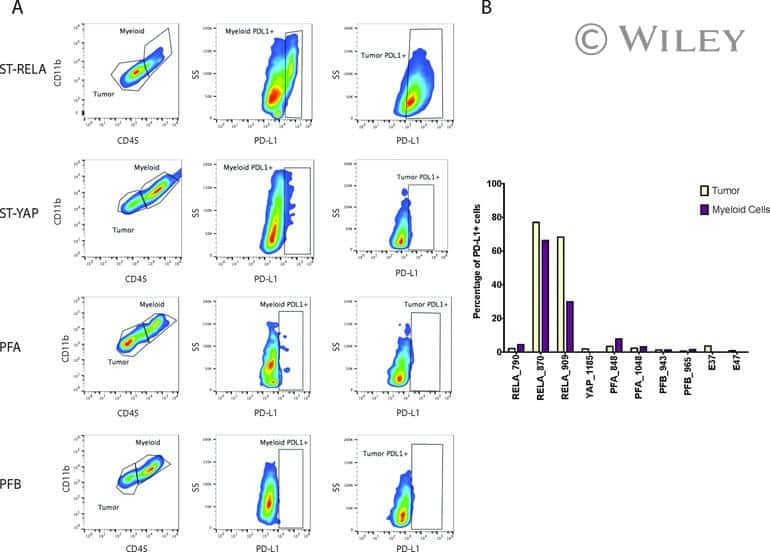
Specific expression of PD-L1 in RELA-fusion supratentorial ependymoma: Implications for PD-1-targeted therapy.
Witt DA, Donson AM, Amani V, Moreira DC, Sanford B, Hoffman LM, Handler MH, Levy JMM, Jones KL, Nellan A, Foreman NK, Griesinger AM
Pediatric blood & cancer 2018 May;65(5):e26960
Pediatric blood & cancer 2018 May;65(5):e26960
ATM‑JAK‑PD‑L1 signaling pathway inhibition decreases EMT and metastasis of androgen‑independent prostate cancer.
Zhang L, Xu LJ, Zhu J, Li J, Xue BX, Gao J, Sun CY, Zang YC, Zhou YB, Yang DR, Shan YX
Molecular medicine reports 2018 May;17(5):7045-7054
Molecular medicine reports 2018 May;17(5):7045-7054
Oncofetal gene SALL4 reactivation by hepatitis B virus counteracts miR-200c in PD-L1-induced T cell exhaustion.
Sun C, Lan P, Han Q, Huang M, Zhang Z, Xu G, Song J, Wang J, Wei H, Zhang J, Sun R, Zhang C, Tian Z
Nature communications 2018 Mar 28;9(1):1241
Nature communications 2018 Mar 28;9(1):1241
Dominant Role for Regulatory T Cells in Protecting Females Against Pulmonary Hypertension.
Tamosiuniene R, Manouvakhova O, Mesange P, Saito T, Qian J, Sanyal M, Lin YC, Nguyen LP, Luria A, Tu AB, Sante JM, Rabinovitch M, Fitzgerald DJ, Graham BB, Habtezion A, Voelkel NF, Aurelian L, Nicolls MR
Circulation research 2018 Jun 8;122(12):1689-1702
Circulation research 2018 Jun 8;122(12):1689-1702
A versatile T cell-based assay to assess therapeutic antigen-specific PD-1-targeted approaches.
Versteven M, Van den Bergh JMJ, Broos K, Fujiki F, Campillo-Davo D, De Reu H, Morimoto S, Lecocq Q, Keyaerts M, Berneman Z, Sugiyama H, Van Tendeloo VFI, Breckpot K, Lion E
Oncotarget 2018 Jun 12;9(45):27797-27808
Oncotarget 2018 Jun 12;9(45):27797-27808
Antigen-Presenting Cell-Intrinsic PD-1 Neutralizes PD-L1 in cis to Attenuate PD-1 Signaling in T Cells.
Zhao Y, Harrison DL, Song Y, Ji J, Huang J, Hui E
Cell reports 2018 Jul 10;24(2):379-390.e6
Cell reports 2018 Jul 10;24(2):379-390.e6
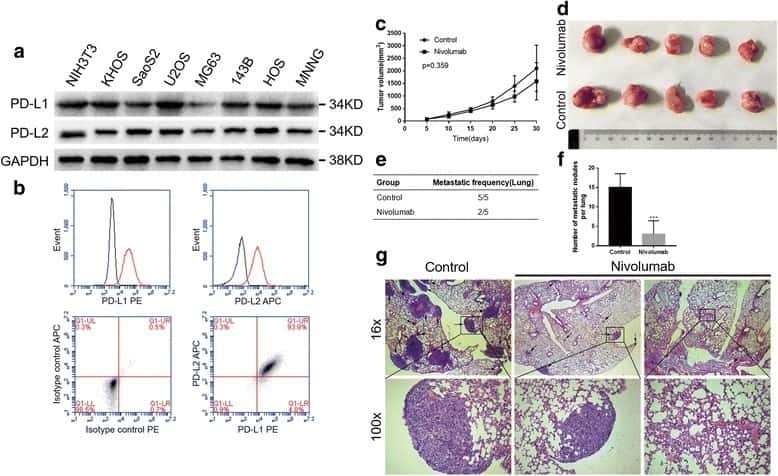
PD-1 axis expression in musculoskeletal tumors and antitumor effect of nivolumab in osteosarcoma model of humanized mouse.
Zheng B, Ren T, Huang Y, Sun K, Wang S, Bao X, Liu K, Guo W
Journal of hematology & oncology 2018 Feb 6;11(1):16
Journal of hematology & oncology 2018 Feb 6;11(1):16
Similarities and differences between helminth parasites and cancer cell lines in shaping human monocytes: Insights into parallel mechanisms of immune evasion.
Narasimhan PB, Akabas L, Tariq S, Huda N, Bennuru S, Sabzevari H, Hofmeister R, Nutman TB, Tolouei Semnani R
PLoS neglected tropical diseases 2018 Apr;12(4):e0006404
PLoS neglected tropical diseases 2018 Apr;12(4):e0006404
Amphotericin B suppresses M2 phenotypes and B7-H1 expression in macrophages to prevent Raji cell proliferation.
Zhang J, Cao D, Yu S, Chen L, Wei D, Shen C, Zhuang L, Wang Q, Xu X, Tong Y
BMC cancer 2018 Apr 26;18(1):467
BMC cancer 2018 Apr 26;18(1):467
PD-L1 expression in medulloblastoma: an evaluation by subgroup.
Martin AM, Nirschl CJ, Polanczyk MJ, Bell WR, Nirschl TR, Harris-Bookman S, Phallen J, Hicks J, Martinez D, Ogurtsova A, Xu H, Sullivan LM, Meeker AK, Raabe EH, Cohen KJ, Eberhart CG, Burger PC, Santi M, Taube JM, Pardoll DM, Drake CG, Lim M
Oncotarget 2018 Apr 10;9(27):19177-19191
Oncotarget 2018 Apr 10;9(27):19177-19191
Amplification of N-Myc is associated with a T-cell-poor microenvironment in metastatic neuroblastoma restraining interferon pathway activity and chemokine expression.
Layer JP, Kronmüller MT, Quast T, van den Boorn-Konijnenberg D, Effern M, Hinze D, Althoff K, Schramm A, Westermann F, Peifer M, Hartmann G, Tüting T, Kolanus W, Fischer M, Schulte J, Hölzel M
Oncoimmunology 2017;6(6):e1320626
Oncoimmunology 2017;6(6):e1320626
Idelalisib and caffeine reduce suppression of T cell responses mediated by activated chronic lymphocytic leukemia cells.
Hock BD, MacPherson SA, McKenzie JL
PloS one 2017;12(3):e0172858
PloS one 2017;12(3):e0172858
Mesenchymal Stromal Cell Secretion of Programmed Death-1 Ligands Regulates T Cell Mediated Immunosuppression.
Davies LC, Heldring N, Kadri N, Le Blanc K
Stem cells (Dayton, Ohio) 2017 Mar;35(3):766-776
Stem cells (Dayton, Ohio) 2017 Mar;35(3):766-776
Structure-guided development of a high-affinity human Programmed Cell Death-1: Implications for tumor immunotherapy.
Lázár-Molnár E, Scandiuzzi L, Basu I, Quinn T, Sylvestre E, Palmieri E, Ramagopal UA, Nathenson SG, Guha C, Almo SC
EBioMedicine 2017 Mar;17:30-44
EBioMedicine 2017 Mar;17:30-44
Retroviral Replicating Vector Delivery of miR-PDL1 Inhibits Immune Checkpoint PDL1 and Enhances Immune Responses In Vitro.
Lin AH, Twitty CG, Burnett R, Hofacre A, Mitchell LA, Espinoza FL, Gruber HE, Jolly DJ
Molecular therapy. Nucleic acids 2017 Mar 17;6:221-232
Molecular therapy. Nucleic acids 2017 Mar 17;6:221-232
Characterization and Purification of Neoplastic Cells of Nodular Lymphocyte Predominant Hodgkin Lymphoma from Lymph Nodes by Flow Cytometry and Flow Cytometric Cell Sorting.
Fromm JR, Thomas A, Wood BL
The American journal of pathology 2017 Feb;187(2):304-317
The American journal of pathology 2017 Feb;187(2):304-317
Oncogenic RAS Signaling Promotes Tumor Immunoresistance by Stabilizing PD-L1 mRNA.
Coelho MA, de Carné Trécesson S, Rana S, Zecchin D, Moore C, Molina-Arcas M, East P, Spencer-Dene B, Nye E, Barnouin K, Snijders AP, Lai WS, Blackshear PJ, Downward J
Immunity 2017 Dec 19;47(6):1083-1099.e6
Immunity 2017 Dec 19;47(6):1083-1099.e6
Hormonal vitamin D up-regulates tissue-specific PD-L1 and PD-L2 surface glycoprotein expression in humans but not mice.
Dimitrov V, Bouttier M, Boukhaled G, Salehi-Tabar R, Avramescu RG, Memari B, Hasaj B, Lukacs GL, Krawczyk CM, White JH
The Journal of biological chemistry 2017 Dec 15;292(50):20657-20668
The Journal of biological chemistry 2017 Dec 15;292(50):20657-20668
PD-L1/PD-1 expression and tumor-infiltrating lymphocytes in conjunctival melanoma.
Cao J, Brouwer NJ, Richards KE, Marinkovic M, van Duinen S, Hurkmans D, Verdegaal EME, Jordanova ES, Jager MJ
Oncotarget 2017 Aug 15;8(33):54722-54734
Oncotarget 2017 Aug 15;8(33):54722-54734
PD-L1 expression in human cancers and its association with clinical outcomes.
Wang X, Teng F, Kong L, Yu J
OncoTargets and therapy 2016;9:5023-39
OncoTargets and therapy 2016;9:5023-39
Tolerogenic IDO(+) Dendritic Cells Are Induced by PD-1-Expressing Mast Cells.
Rodrigues CP, Ferreira AC, Pinho MP, de Moraes CJ, Bergami-Santos PC, Barbuto JA
Frontiers in immunology 2016;7:9
Frontiers in immunology 2016;7:9
Ly6C(hi) monocytes regulate T cell responses in viral hepatitis.
Zhu J, Chen H, Huang X, Jiang S, Yang Y
JCI insight 2016 Oct 20;1(17):e89880
JCI insight 2016 Oct 20;1(17):e89880
Regulatory IgDhi B Cells Suppress T Cell Function via IL-10 and PD-L1 during Progressive Visceral Leishmaniasis.
Schaut RG, Lamb IM, Toepp AJ, Scott B, Mendes-Aguiar CO, Coutinho JF, Jeronimo SM, Wilson ME, Harty JT, Waldschmidt TJ, Petersen CA
Journal of immunology (Baltimore, Md. : 1950) 2016 May 15;196(10):4100-9
Journal of immunology (Baltimore, Md. : 1950) 2016 May 15;196(10):4100-9
Expression of programmed death 1 ligand 1 on periodontal tissue cells as a possible protective feedback mechanism against periodontal tissue destruction.
Zhang J, Wang CM, Zhang P, Wang X, Chen J, Yang J, Lu W, Zhou W, Yuan W, Feng Y
Molecular medicine reports 2016 Mar;13(3):2423-30
Molecular medicine reports 2016 Mar;13(3):2423-30
CD28 family of receptors on T cells in chronic HBV infection: Expression characteristics, clinical significance and correlations with PD-1 blockade.
Tang ZS, Hao YH, Zhang EJ, Xu CL, Zhou Y, Zheng X, Yang DL
Molecular medicine reports 2016 Aug;14(2):1107-16
Molecular medicine reports 2016 Aug;14(2):1107-16
Psoriasis is characterized by deficient negative immune regulation compared to transient delayed-type hypersensitivity reactions.
Gulati N, Suárez-Fariñas M, Correa da Rosa J, Krueger JG
F1000Research 2015;4:149
F1000Research 2015;4:149
Upregulation of Programmed Death-1 and Its Ligand in Cardiac Injury Models: Interaction with GADD153.
Baban B, Liu JY, Qin X, Weintraub NL, Mozaffari MS
PloS one 2015;10(4):e0124059
PloS one 2015;10(4):e0124059
Interleukin-25 Mediates Transcriptional Control of PD-L1 via STAT3 in Multipotent Human Mesenchymal Stromal Cells (hMSCs) to Suppress Th17 Responses.
Wang WB, Yen ML, Liu KJ, Hsu PJ, Lin MH, Chen PM, Sudhir PR, Chen CH, Chen CH, Sytwu HK, Yen BL
Stem cell reports 2015 Sep 8;5(3):392-404
Stem cell reports 2015 Sep 8;5(3):392-404
Programmed death ligand 1 on Burkholderia pseudomallei-infected human polymorphonuclear neutrophils impairs T cell functions.
Buddhisa S, Rinchai D, Ato M, Bancroft GJ, Lertmemongkolchai G
Journal of immunology (Baltimore, Md. : 1950) 2015 May 1;194(9):4413-21
Journal of immunology (Baltimore, Md. : 1950) 2015 May 1;194(9):4413-21
Assays for predicting and monitoring responses to lung cancer immunotherapy.
Teixidó C, Karachaliou N, González-Cao M, Morales-Espinosa D, Rosell R
Cancer biology & medicine 2015 Jun;12(2):87-95
Cancer biology & medicine 2015 Jun;12(2):87-95
Control of signaling-mediated clearance of apoptotic cells by the tumor suppressor p53.
Yoon KW, Byun S, Kwon E, Hwang SY, Chu K, Hiraki M, Jo SH, Weins A, Hakroush S, Cebulla A, Sykes DB, Greka A, Mundel P, Fisher DE, Mandinova A, Lee SW
Science (New York, N.Y.) 2015 Jul 31;349(6247):1261669
Science (New York, N.Y.) 2015 Jul 31;349(6247):1261669
Paucity of PD-L1 expression in prostate cancer: innate and adaptive immune resistance.
Martin AM, Nirschl TR, Nirschl CJ, Francica BJ, Kochel CM, van Bokhoven A, Meeker AK, Lucia MS, Anders RA, DeMarzo AM, Drake CG
Prostate cancer and prostatic diseases 2015 Dec;18(4):325-32
Prostate cancer and prostatic diseases 2015 Dec;18(4):325-32
IL-27 induces the expression of IDO and PD-L1 in human cancer cells.
Carbotti G, Barisione G, Airoldi I, Mezzanzanica D, Bagnoli M, Ferrero S, Petretto A, Fabbi M, Ferrini S
Oncotarget 2015 Dec 22;6(41):43267-80
Oncotarget 2015 Dec 22;6(41):43267-80
Inhibition of mitogen-activated protein kinase pathway can induce upregulation of human leukocyte antigen class I without PD-L1-upregulation in contrast to interferon-γ treatment.
Mimura K, Kua LF, Shiraishi K, Kee Siang L, Shabbir A, Komachi M, Suzuki Y, Nakano T, Yong WP, So J, Kono K
Cancer science 2014 Oct;105(10):1236-44
Cancer science 2014 Oct;105(10):1236-44
Immune-escape markers in relation to clinical outcome of advanced melanoma patients following immunotherapy.
Tjin EP, Krebbers G, Meijlink KJ, van de Kasteele W, Rosenberg EH, Sanders J, Nederlof PM, van de Wiel BA, Haanen JB, Melief CJ, Vyth-Dreese FA, Luiten RM
Cancer immunology research 2014 Jun;2(6):538-46
Cancer immunology research 2014 Jun;2(6):538-46
Galectin-9 and IL-21 mediate cross-regulation between Th17 and Treg cells during acute hepatitis C.
Kared H, Fabre T, Bédard N, Bruneau J, Shoukry NH
PLoS pathogens 2013;9(6):e1003422
PLoS pathogens 2013;9(6):e1003422
NF-κB plays a key role in inducing CD274 expression in human monocytes after lipopolysaccharide treatment.
Huang G, Wen Q, Zhao Y, Gao Q, Bai Y
PloS one 2013;8(4):e61602
PloS one 2013;8(4):e61602
Soluble CD80 restores T cell activation and overcomes tumor cell programmed death ligand 1-mediated immune suppression.
Haile ST, Dalal SP, Clements V, Tamada K, Ostrand-Rosenberg S
Journal of immunology (Baltimore, Md. : 1950) 2013 Sep 1;191(5):2829-36
Journal of immunology (Baltimore, Md. : 1950) 2013 Sep 1;191(5):2829-36
Alterations of immune response of Non-Small Cell Lung Cancer with Azacytidine.
Wrangle J, Wang W, Koch A, Easwaran H, Mohammad HP, Vendetti F, Vancriekinge W, Demeyer T, Du Z, Parsana P, Rodgers K, Yen RW, Zahnow CA, Taube JM, Brahmer JR, Tykodi SS, Easton K, Carvajal RD, Jones PA, Laird PW, Weisenberger DJ, Tsai S, Juergens RA, Topalian SL, Rudin CM, Brock MV, Pardoll D, Baylin SB
Oncotarget 2013 Nov;4(11):2067-79
Oncotarget 2013 Nov;4(11):2067-79
Antagonist antibodies to PD-1 and B7-H1 (PD-L1) in the treatment of advanced human cancer.
Sznol M, Chen L
Clinical cancer research : an official journal of the American Association for Cancer Research 2013 Mar 1;19(5):1021-34
Clinical cancer research : an official journal of the American Association for Cancer Research 2013 Mar 1;19(5):1021-34
Programmed death 1-mediated T cell exhaustion during visceral leishmaniasis impairs phagocyte function.
Esch KJ, Juelsgaard R, Martinez PA, Jones DE, Petersen CA
Journal of immunology (Baltimore, Md. : 1950) 2013 Dec 1;191(11):5542-50
Journal of immunology (Baltimore, Md. : 1950) 2013 Dec 1;191(11):5542-50
Inhibiting the programmed death 1 pathway rescues Mycobacterium tuberculosis-specific interferon γ-producing T cells from apoptosis in patients with pulmonary tuberculosis.
Singh A, Mohan A, Dey AB, Mitra DK
The Journal of infectious diseases 2013 Aug 15;208(4):603-15
The Journal of infectious diseases 2013 Aug 15;208(4):603-15
Foxp3+ regulatory T cells among tuberculosis patients: impact on prognosis and restoration of antigen specific IFN-γ producing T cells.
Singh A, Dey AB, Mohan A, Sharma PK, Mitra DK
PloS one 2012;7(9):e44728
PloS one 2012;7(9):e44728
Tissue biomarkers in melanoma patients treated with TIL.
Knol AC, Nguyen JM, Pandolfino MC, Quéreux G, Brocard A, Peuvrel L, Saint-Jean M, Saiagh S, Khammari A, Dréno B
PloS one 2012;7(12):e48729
PloS one 2012;7(12):e48729
TNF-α and TGF-β counter-regulate PD-L1 expression on monocytes in systemic lupus erythematosus.
Ou JN, Wiedeman AE, Stevens AM
Scientific reports 2012;2:295
Scientific reports 2012;2:295
Upregulation of programmed death-1 on T cells and programmed death ligand-1 on monocytes in septic shock patients.
Zhang Y, Li J, Lou J, Zhou Y, Bo L, Zhu J, Zhu K, Wan X, Cai Z, Deng X
Critical care (London, England) 2011;15(1):R70
Critical care (London, England) 2011;15(1):R70
HIV-mediated phosphatidylinositol 3-kinase/serine-threonine kinase activation in APCs leads to programmed death-1 ligand upregulation and suppression of HIV-specific CD8 T cells.
Muthumani K, Shedlock DJ, Choo DK, Fagone P, Kawalekar OU, Goodman J, Bian CB, Ramanathan AA, Atman P, Tebas P, Chattergoon MA, Choo AY, Weiner DB
Journal of immunology (Baltimore, Md. : 1950) 2011 Sep 15;187(6):2932-43
Journal of immunology (Baltimore, Md. : 1950) 2011 Sep 15;187(6):2932-43
Activated memory B cells may function as antigen-presenting cells in the joints of children with juvenile idiopathic arthritis.
Morbach H, Wiegering V, Richl P, Schwarz T, Suffa N, Eichhorn EM, Eyrich M, Girschick HJ
Arthritis and rheumatism 2011 Nov;63(11):3458-66
Arthritis and rheumatism 2011 Nov;63(11):3458-66
Tumor cell programmed death ligand 1-mediated T cell suppression is overcome by coexpression of CD80.
Haile ST, Bosch JJ, Agu NI, Zeender AM, Somasundaram P, Srivastava MK, Britting S, Wolf JB, Ksander BR, Ostrand-Rosenberg S
Journal of immunology (Baltimore, Md. : 1950) 2011 Jun 15;186(12):6822-9
Journal of immunology (Baltimore, Md. : 1950) 2011 Jun 15;186(12):6822-9
Immunostaining of PD-1/PD-Ls in liver tissues of patients with hepatitis and hepatocellular carcinoma.
Wang BJ, Bao JJ, Wang JZ, Wang Y, Jiang M, Xing MY, Zhang WG, Qi JY, Roggendorf M, Lu MJ, Yang DL
World journal of gastroenterology 2011 Jul 28;17(28):3322-9
World journal of gastroenterology 2011 Jul 28;17(28):3322-9
Heat shock protein 27 differentiates tolerogenic macrophages that may support human breast cancer progression.
Banerjee S, Lin CF, Skinner KA, Schiffhauer LM, Peacock J, Hicks DG, Redmond EM, Morrow D, Huston A, Shayne M, Langstein HN, Miller-Graziano CL, Strickland J, O'Donoghue L, De AK
Cancer research 2011 Jan 15;71(2):318-27
Cancer research 2011 Jan 15;71(2):318-27
Ex vivo expanded hematopoietic stem cells overcome the MHC barrier in allogeneic transplantation.
Zheng J, Umikawa M, Zhang S, Huynh H, Silvany R, Chen BP, Chen L, Zhang CC
Cell stem cell 2011 Aug 5;9(2):119-30
Cell stem cell 2011 Aug 5;9(2):119-30
Doxorubicin downregulates cell surface B7-H1 expression and upregulates its nuclear expression in breast cancer cells: role of B7-H1 as an anti-apoptotic molecule.
Ghebeh H, Lehe C, Barhoush E, Al-Romaih K, Tulbah A, Al-Alwan M, Hendrayani SF, Manogaran P, Alaiya A, Al-Tweigeri T, Aboussekhra A, Dermime S
Breast cancer research : BCR 2010;12(4):R48
Breast cancer research : BCR 2010;12(4):R48
The PD-1/PD-L1 axis modulates the natural killer cell versus multiple myeloma effect: a therapeutic target for CT-011, a novel monoclonal anti-PD-1 antibody.
Benson DM Jr, Bakan CE, Mishra A, Hofmeister CC, Efebera Y, Becknell B, Baiocchi RA, Zhang J, Yu J, Smith MK, Greenfield CN, Porcu P, Devine SM, Rotem-Yehudar R, Lozanski G, Byrd JC, Caligiuri MA
Blood 2010 Sep 30;116(13):2286-94
Blood 2010 Sep 30;116(13):2286-94
Mechanisms of immune suppression for CD8+ T cells by human corneal endothelial cells via membrane-bound TGFbeta.
Yamada Y, Sugita S, Horie S, Yamagami S, Mochizuki M
Investigative ophthalmology & visual science 2010 May;51(5):2548-57
Investigative ophthalmology & visual science 2010 May;51(5):2548-57
Role played by the programmed death-1-programmed death ligand pathway during innate immunity against Mycobacterium tuberculosis.
Alvarez IB, Pasquinelli V, Jurado JO, Abbate E, Musella RM, de la Barrera SS, García VE
The Journal of infectious diseases 2010 Aug 15;202(4):524-32
The Journal of infectious diseases 2010 Aug 15;202(4):524-32
Tumor-infiltrating NY-ESO-1-specific CD8+ T cells are negatively regulated by LAG-3 and PD-1 in human ovarian cancer.
Matsuzaki J, Gnjatic S, Mhawech-Fauceglia P, Beck A, Miller A, Tsuji T, Eppolito C, Qian F, Lele S, Shrikant P, Old LJ, Odunsi K
Proceedings of the National Academy of Sciences of the United States of America 2010 Apr 27;107(17):7875-80
Proceedings of the National Academy of Sciences of the United States of America 2010 Apr 27;107(17):7875-80
B7-H1 (PD-L1, CD274) suppresses host immunity in T-cell lymphoproliferative disorders.
Wilcox RA, Feldman AL, Wada DA, Yang ZZ, Comfere NI, Dong H, Kwon ED, Novak AJ, Markovic SN, Pittelkow MR, Witzig TE, Ansell SM
Blood 2009 Sep 3;114(10):2149-58
Blood 2009 Sep 3;114(10):2149-58
Costimulatory pathways in multiple sclerosis: distinctive expression of PD-1 and PD-L1 in patients with different patterns of disease.
Trabattoni D, Saresella M, Pacei M, Marventano I, Mendozzi L, Rovaris M, Caputo D, Borelli M, Clerici M
Journal of immunology (Baltimore, Md. : 1950) 2009 Oct 15;183(8):4984-93
Journal of immunology (Baltimore, Md. : 1950) 2009 Oct 15;183(8):4984-93
PD-1 is a regulator of NY-ESO-1-specific CD8+ T cell expansion in melanoma patients.
Fourcade J, Kudela P, Sun Z, Shen H, Land SR, Lenzner D, Guillaume P, Luescher IF, Sander C, Ferrone S, Kirkwood JM, Zarour HM
Journal of immunology (Baltimore, Md. : 1950) 2009 May 1;182(9):5240-9
Journal of immunology (Baltimore, Md. : 1950) 2009 May 1;182(9):5240-9
Activated monocytes in peritumoral stroma of hepatocellular carcinoma foster immune privilege and disease progression through PD-L1.
Kuang DM, Zhao Q, Peng C, Xu J, Zhang JP, Wu C, Zheng L
The Journal of experimental medicine 2009 Jun 8;206(6):1327-37
The Journal of experimental medicine 2009 Jun 8;206(6):1327-37
Human corneal endothelial cells expressing programmed death-ligand 1 (PD-L1) suppress PD-1+ T helper 1 cells by a contact-dependent mechanism.
Sugita S, Usui Y, Horie S, Futagami Y, Yamada Y, Ma J, Kezuka T, Hamada H, Usui T, Mochizuki M, Yamagami S
Investigative ophthalmology & visual science 2009 Jan;50(1):263-72
Investigative ophthalmology & visual science 2009 Jan;50(1):263-72
PD-1/PD-L1 expression in human T-cell leukemia virus type 1 carriers and adult T-cell leukemia/lymphoma patients.
Kozako T, Yoshimitsu M, Fujiwara H, Masamoto I, Horai S, White Y, Akimoto M, Suzuki S, Matsushita K, Uozumi K, Tei C, Arima N
Leukemia 2009 Feb;23(2):375-82
Leukemia 2009 Feb;23(2):375-82
MicroRNA-513 regulates B7-H1 translation and is involved in IFN-gamma-induced B7-H1 expression in cholangiocytes.
Gong AY, Zhou R, Hu G, Li X, Splinter PL, O'Hara SP, LaRusso NF, Soukup GA, Dong H, Chen XM
Journal of immunology (Baltimore, Md. : 1950) 2009 Feb 1;182(3):1325-33
Journal of immunology (Baltimore, Md. : 1950) 2009 Feb 1;182(3):1325-33
Interaction of human PD-L1 and B7-1.
Butte MJ, Peña-Cruz V, Kim MJ, Freeman GJ, Sharpe AH
Molecular immunology 2008 Aug;45(13):3567-72
Molecular immunology 2008 Aug;45(13):3567-72
Expression of B7-H1 in inflammatory renal tubular epithelial cells.
Chen Y, Zhang J, Li J, Zou L, Zhao T, Tang Y, Wu Y
Nephron. Experimental nephrology 2006;102(3-4):e81-92
Nephron. Experimental nephrology 2006;102(3-4):e81-92
The B7-H1 (PD-L1) T lymphocyte-inhibitory molecule is expressed in breast cancer patients with infiltrating ductal carcinoma: correlation with important high-risk prognostic factors.
Ghebeh H, Mohammed S, Al-Omair A, Qattan A, Lehe C, Al-Qudaihi G, Elkum N, Alshabanah M, Bin Amer S, Tulbah A, Ajarim D, Al-Tweigeri T, Dermime S
Neoplasia (New York, N.Y.) 2006 Mar;8(3):190-8
Neoplasia (New York, N.Y.) 2006 Mar;8(3):190-8
NK cells that are activated by CXCL10 can kill dormant tumor cells that resist CTL-mediated lysis and can express B7-H1 that stimulates T cells.
Saudemont A, Jouy N, Hetuin D, Quesnel B
Blood 2005 Mar 15;105(6):2428-35
Blood 2005 Mar 15;105(6):2428-35
Clinical significance of programmed death-1 ligand-1 and programmed death-1 ligand-2 expression in human esophageal cancer.
Ohigashi Y, Sho M, Yamada Y, Tsurui Y, Hamada K, Ikeda N, Mizuno T, Yoriki R, Kashizuka H, Yane K, Tsushima F, Otsuki N, Yagita H, Azuma M, Nakajima Y
Clinical cancer research : an official journal of the American Association for Cancer Research 2005 Apr 15;11(8):2947-53
Clinical cancer research : an official journal of the American Association for Cancer Research 2005 Apr 15;11(8):2947-53
Blockade of B7-H1 suppresses the development of chronic intestinal inflammation.
Kanai T, Totsuka T, Uraushihara K, Makita S, Nakamura T, Koganei K, Fukushima T, Akiba H, Yagita H, Okumura K, Machida U, Iwai H, Azuma M, Chen L, Watanabe M
Journal of immunology (Baltimore, Md. : 1950) 2003 Oct 15;171(8):4156-63
Journal of immunology (Baltimore, Md. : 1950) 2003 Oct 15;171(8):4156-63
Engagement of the PD-1 immunoinhibitory receptor by a novel B7 family member leads to negative regulation of lymphocyte activation.
Freeman GJ, Long AJ, Iwai Y, Bourque K, Chernova T, Nishimura H, Fitz LJ, Malenkovich N, Okazaki T, Byrne MC, Horton HF, Fouser L, Carter L, Ling V, Bowman MR, Carreno BM, Collins M, Wood CR, Honjo T
The Journal of experimental medicine 2000 Oct 2;192(7):1027-34
The Journal of experimental medicine 2000 Oct 2;192(7):1027-34
B7-H1, a third member of the B7 family, co-stimulates T-cell proliferation and interleukin-10 secretion.
Dong H, Zhu G, Tamada K, Chen L
Nature medicine 1999 Dec;5(12):1365-9
Nature medicine 1999 Dec;5(12):1365-9
No comments: Submit comment
Supportive validation
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
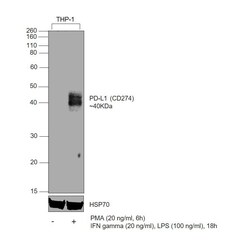
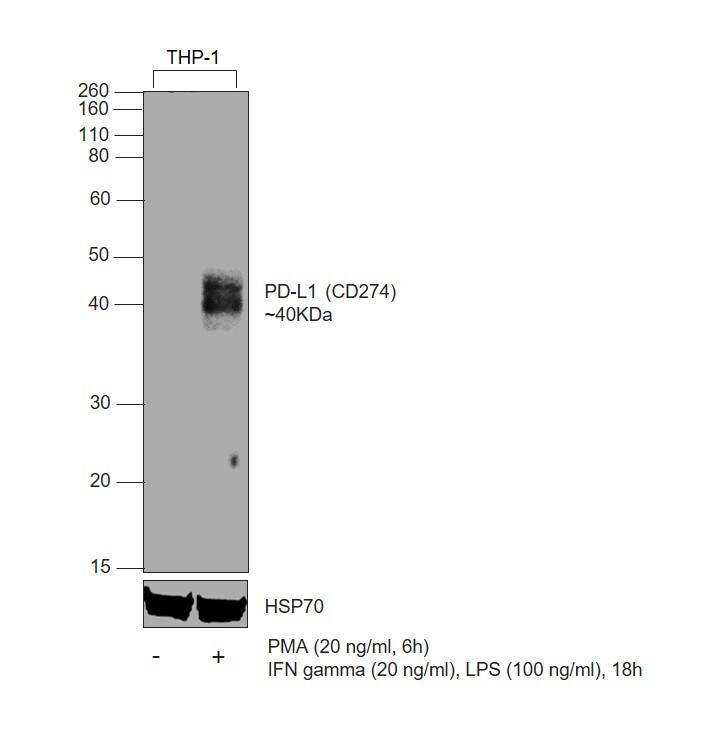
- Experimental details
- Western blot was performed using CD274 (PD-L1, B7-H1) Monoclonal Antibody (MIH1), eBioscience™ (Product # 14-5983-82) and a 40 kDa band corresponding to Programmed cell death 1 ligand 1 was observed across in treated THP-1 cells, but not in the untreated. Whole cell extracts (30 µg lysate) of THP-1 (Lane 1), THP-1 treated with PMA (200 ng/mL, 6 hours), IFN gamma (20 ng/mL) and LPS (100 ng/mL), 18hours (Lane 2) were electrophoresed using NuPAGE™ 10% Bis-Tris Protein Gel (Product # NP0302BOX), 12 well. Resolved proteins were then transferred onto a nitrocellulose membrane (Product # IB23002) by iBlot® 2 Dry Blotting System (Product # IB21001). The blot was probed with the primary antibody (1:500 dilution) and detected by chemiluminescence with Goat anti-Mouse IgG (H+L) Superclonal™ Recombinant Secondary Antibody, HRP (Product # A28177, 1:10,000) using the iBright™ FL1500 Imaging System (Product # A44115). Chemiluminescent detection was performed using SuperSignal™ West Atto Ultimate Sensitivity Substrate (Product # A38556).
Supportive validation
- Submitted by
- Invitrogen Antibodies (provider)
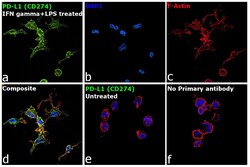
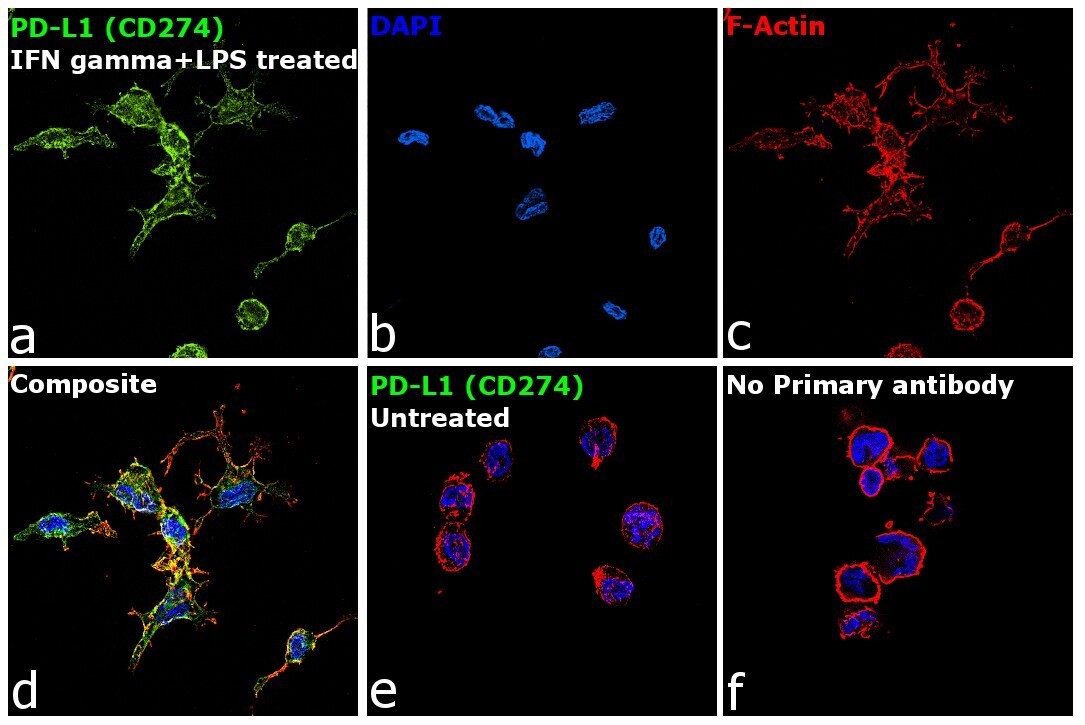
- Main image
- Experimental details
- Immunofluorescence analysis of Programmed cell death 1 ligand 1 was performed using 70% confluent log phase THP-1 cells treated with PMA (200 ng/mL for 6h) and further polarized into M1 macrophages using IFN gamma (20 ng/mL) and LPS (100 ng/mL) for 18h. The cells were fixed with 4% paraformaldehyde for 10 minutes, permeabilized with 0.1% Triton™ X-100 for 15 minutes and blocked with 2% BSA for 45 minutes at room temperature. The cells were labeled with CD274 (PD-L1, B7-H1) Monoclonal Antibody (MIH1), eBioscience™ (Product # 14-5983-82) at 1:100 dilution in 0.1% BSA, incubated at 4-degree Celsius overnight and then labeled with Donkey anti-Mouse IgG (H+L) Highly Cross-Adsorbed Secondary Antibody, Alexa Fluor Plus 488 (Product # A32766), (1:2000 dilution), for 45 minutes at room temperature (Panel a: Green). Nuclei (Panel b: Blue) were stained with ProLong™ Diamond Antifade Mountant with DAPI (Product # P36962). F-actin (Panel c: Red) was stained with Rhodamine Phalloidin (Product # R415, 1:300 dilution). Panel d represents the merged image showing membranous localization. Panel e represents untreated cells. Panel f represents control cells with no primary antibody to assess the background. The images were captured at 60X magnification.
Supportive validation
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
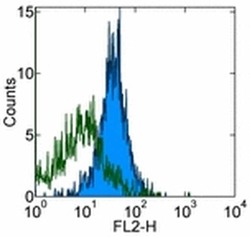
- Experimental details
- Staining of normal human peripheral blood cells with 0.5 µg of Mouse IgG1 K Isotype Control Purified (Product # 14-4714-82) (open histogram) or 0.5 µg of Anti-Human CD274 (B7-H1) Purified (filled histogram) followed by Anti-Mouse IgG Biotin (Product # 13-4013-85)and Streptavidin PE (Product # 12-4317-87).Cells in the lymphocyte gate were used for analysis.
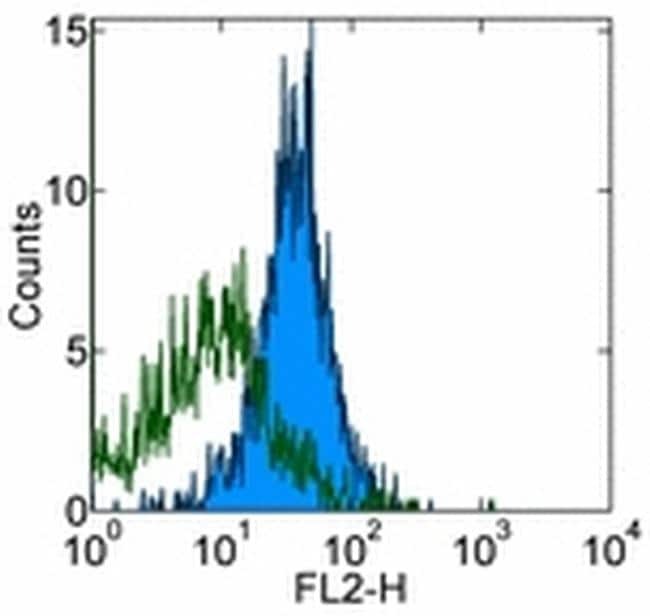
Supportive validation
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
- NULL

- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
- NULL

- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
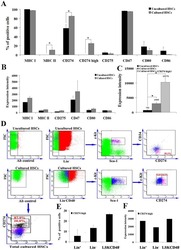
- Experimental details
- NULL
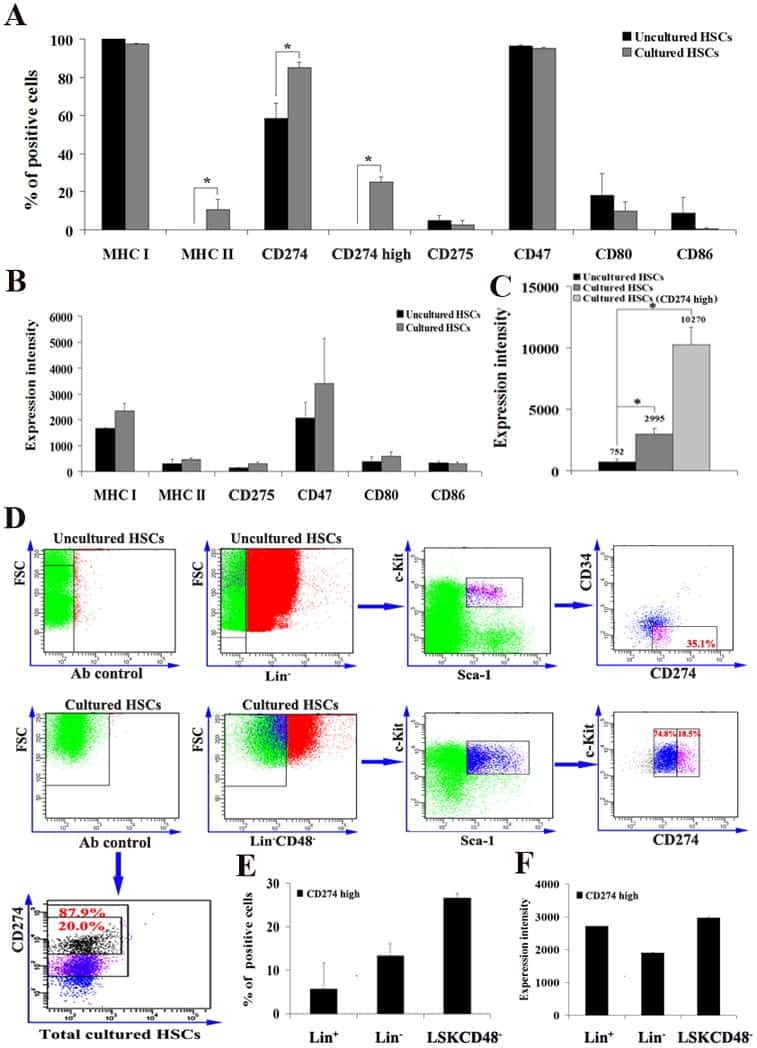
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
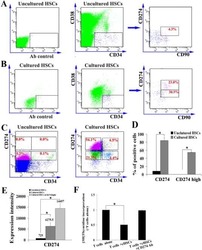
- Experimental details
- NULL
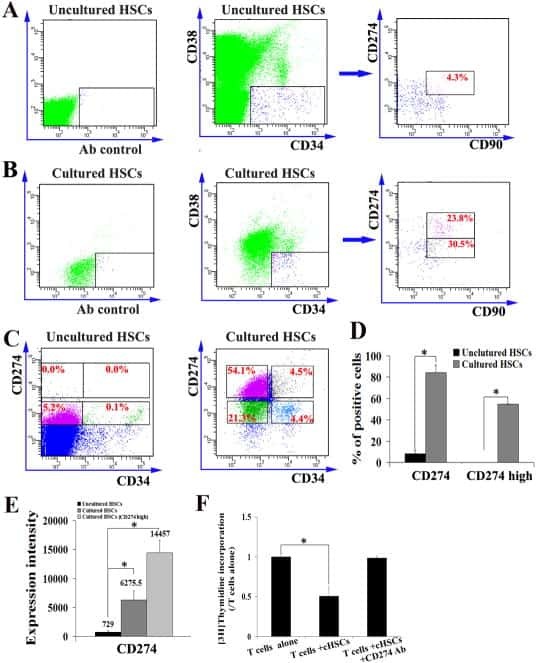
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
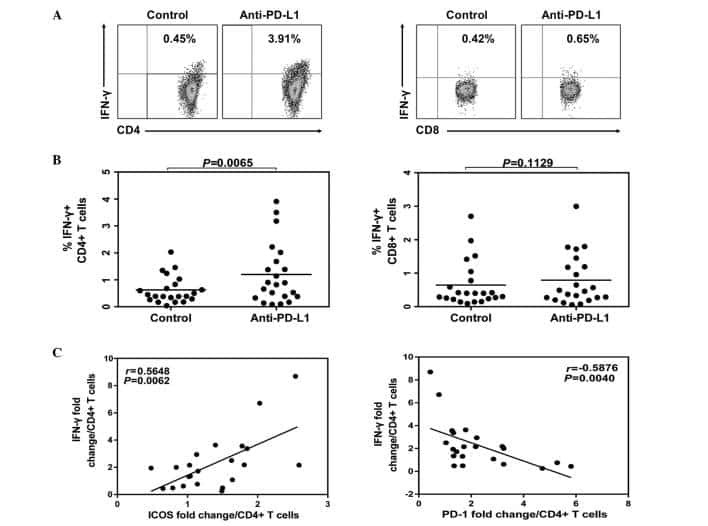
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
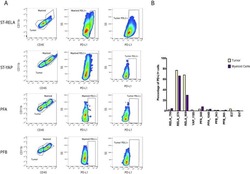
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
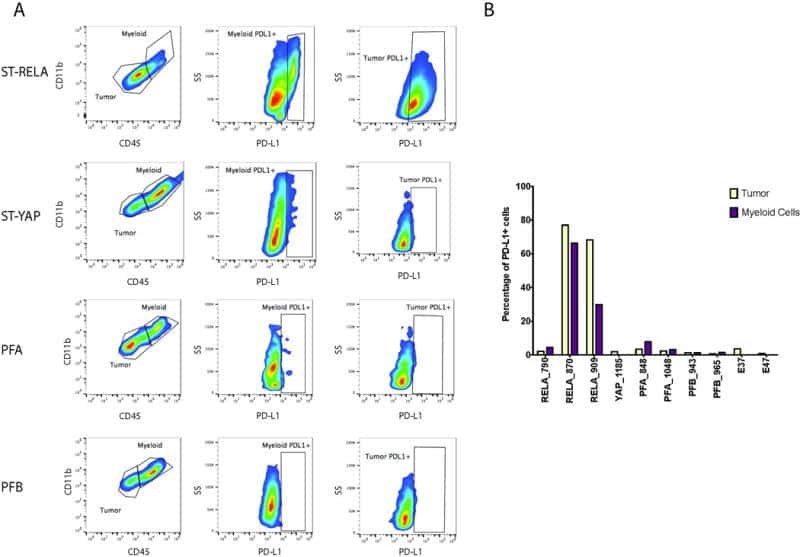
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
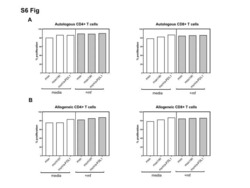
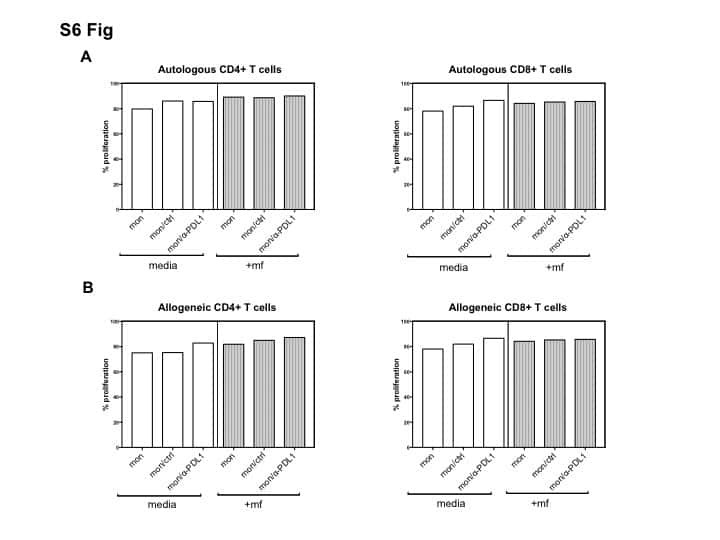
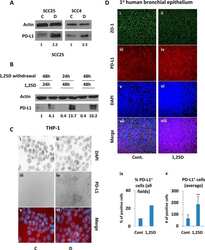
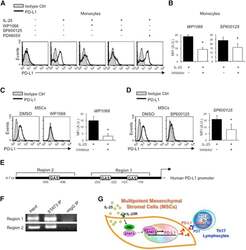
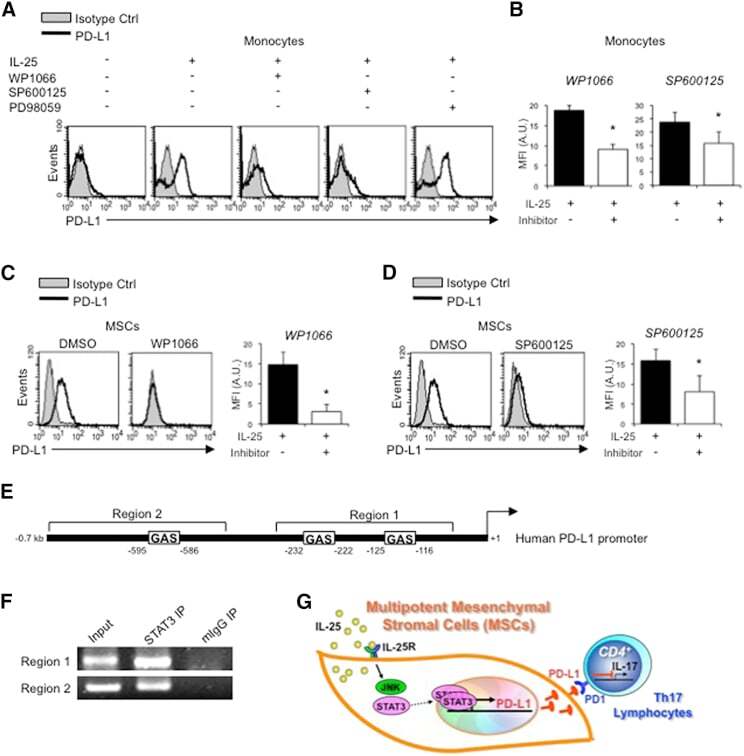
- Figure 6 IL-25-Mediated PD-L1 Expression in Human Monocytes and hMSCs Is Mediated through JNK and STAT3, with STAT3 Involved in Transcriptional Control of PD-L1 (A) Human PBLs were pretreated with inhibitors of STAT3 (WP1066; 2.5 muM), JNK (SP600125; 25 muM), or MEK1 (PD98059; 20 muM) prior to 100 ng/ml rhIL-25 for 18 hr, with subsequent flow cytometric analysis for PD-L1 surface expression on monocytes, gated using FSC and SSC. Filled histograms represent isotype control; unfilled histograms represent PD-L1 antibody staining. (B-D) Pooled data (three donors) are shown (B) with bars representing MFI. hMSCs were treated with inhibitors of STAT3 (C; WP1066, 2.5 muM) and JNK (D; SP600125, 25 muM) for 6 hr, and subsequently assessed by flow cytometric analysis for PD-L1 surface expression. Pooled data (all three donors) for each respective inhibitor are provided (left charts) with bars representing MFI. (E) Putative GAS elements (STAT-binding sites) in the proximal promoter region of human PD-L1 gene (700 bp region upstream from the transcription start site), as determined with TFSearch web-based software. (F) Binding of STAT3 or IgG (negative control) in hMSCs was analyzed by chromatin immunoprecipitation (ChIP) with promoter-specific primers for region 1 and region 2. The input samples (positive control) represent 1% starting chromatin. (G) Schematic shows a model of hMSC-mediated suppression of Th17 responses involving the IL-25/STAT3/PD-L1 axis.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
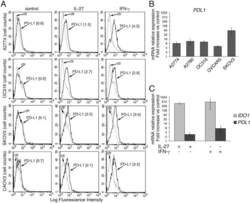
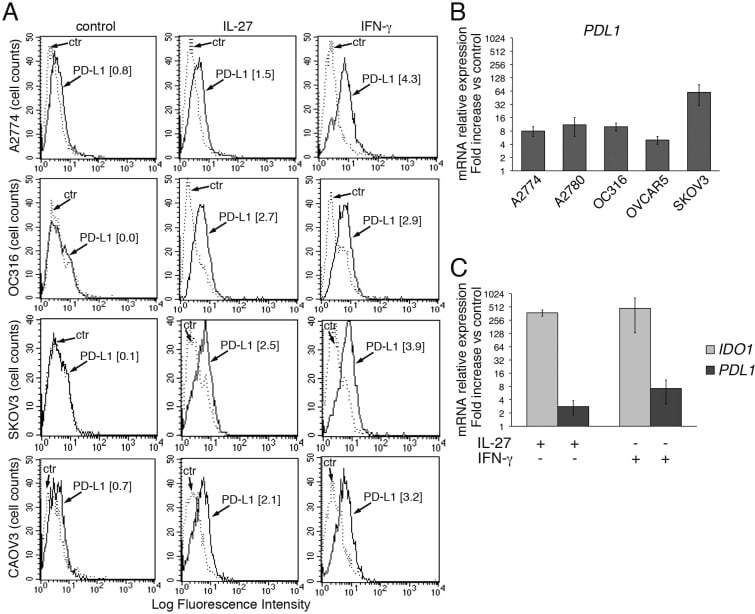
- Figure 2 IL-27 increases PD-L1 surface protein and mRNA expression in EOC cells in vitro A. FACS analysis of surface PD-L1 expression in four EOC cell lines, cultured in the presence of medium (control), IL-27 or IFN-gamma. Dotted lines are isotype-matched unrelated Ig staining controls. Numbers in brackets are Median Fluorescence Intensity (MFI) values calculated as median PD-L1 minus median Ig control. Data are representative of two independent experiments showing similar results. B. QRT-PCR analysis of PDL1 mRNA expression in five IL-27-stimulated EOC cells relative to untreated cells. Data are the mean (+-SD) of three independent experiments. C. Comparative analysis of IDO1 and PDL1 mRNA up-regulation by IL-27 or IFN-gamma in a representative EOC cell line (CAOV3). Data are the mean of two independent replicates and are expressed as DeltaDeltaCT-fold change. Error bars represent the minimum and maximum.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
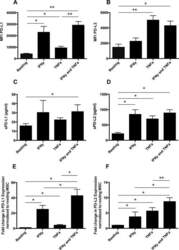
- Figure 1 Mesenchymal stromal cell (MSC) cell surface expression and secretion of PD-L1 and PD-L2 are potentiated by pro-inflammatory cytokines, IFNgamma and TNFalpha. MSCs ( n = 4) were exposed to 100 U/ml IFNgamma and 10 ng/ml TNFalpha for 3 days in culture. Cell surface expression (MFI) of (A) PD-L1 and (B) PD-L2 was assessed by flow cytometry. Secretion of (C) soluble (s)PD-L1 and (D) sPD-L2 within the conditioned media of stimulated cells was assessed by ELISA. Bar charts indicate mean +- SEM. Transcriptional regulation of (E) PD-L1 and (F) PD-L2 were assessed by qRT-PCR. mRNA data are expressed as fold change compared to unstimulated, resting MSCs +- SEM. *, p < .05; **, p < .01. Abbreviations: IFNgamma, Interferon gamma; MFI, mean fluorescence intensity; PD-L1 and PD-L2, programmed death 1 ligands 1 and 2; TNFalpha, tumor necrosis factor alpha.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
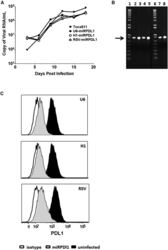
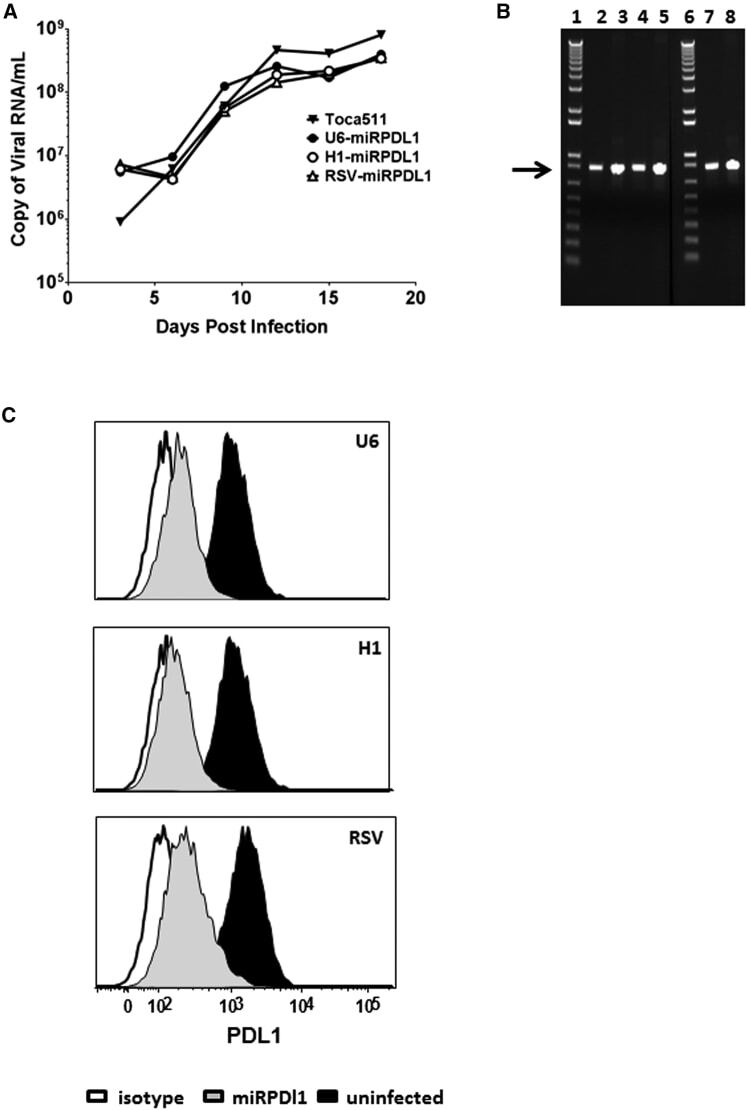
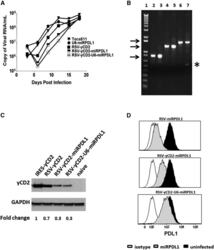
- Figure 4 RRV-RSV-miRPDL1 and RRV-H1-miRPDL1 Exhibit Comparable PDL1 Downregulation Activity as RRV-miRPDL1 (A) Replication kinetics of RRV-RSV-miRPDL1 and RRV-H1-miRPDL1. The viral genome in the supernatants of infected LN-18 cells (MOI of 0.1) at indicated time points were quantified by qRT-PCR using primer set targeted to the env region ( Figure 1 ). RRV-yCD2 and RRV-miRPDL1 (indicated as U6-miRPDL1 in the graph) were included as positive controls. (B) Vector stability of RRV-RSV-miRPDL1 and RRV-H1-miRPDL1 in LN-18 cells was analyzed by endpoint PCR at 14 and 30 days post infection. Lanes 1 and 6: DNA molecular marker (1 Kb Plus marker, Invitrogen); lanes 2, 4, and 7 are positive controls using the corresponding plasmid DNA as the templates; lane 3: RRV-H1-miRPDL1; lane 5: RRV-RSV-miRPDL1; and lane 8: RRV-miRPDL1. The arrow indicates the expected size of the PCR products. (C) LN-18 cells infected with RRV-miRPDL1 (U6), RRV-H1-miRPDL1 (H1), and RRV-RSV-miRPDL1 (RSV) were stained for PDL1 cell surface expression with PDL1 antibody and analyzed by flow cytometry.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
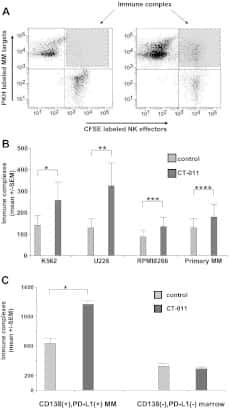
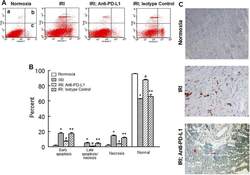
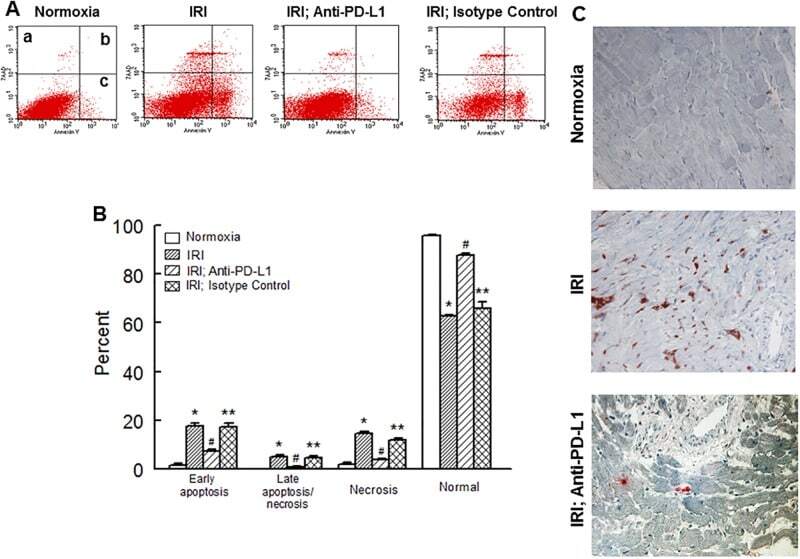
- Fig 6 Representative dot matrices for apoptotic and necrotic cell death of experimental groups are shown under panel A while bar graphs (panel B) show percent of each type of damaged/dead or normal cells for each group. Data are means +- SEM; n = 6 hearts/group for normoxic control, IRI and IRI; Anti-PD-L1 and n = 4 hearts for IRI; isotype matching control. Also shown are representative caspase 3 immunostaining for experimental groups (panel C). 400x. 7AAD: 7-Amino Actinomycin D. a: necrotic; b: apoptotic/necrotic; c: early apoptotic. IRI: Ischemia Reperfusion Injury. * p
- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
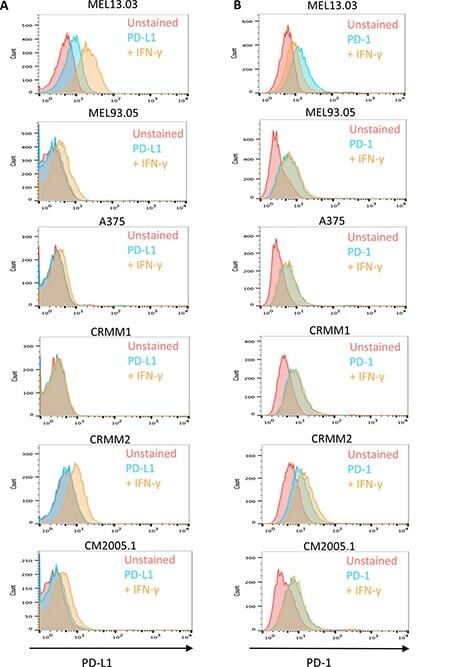
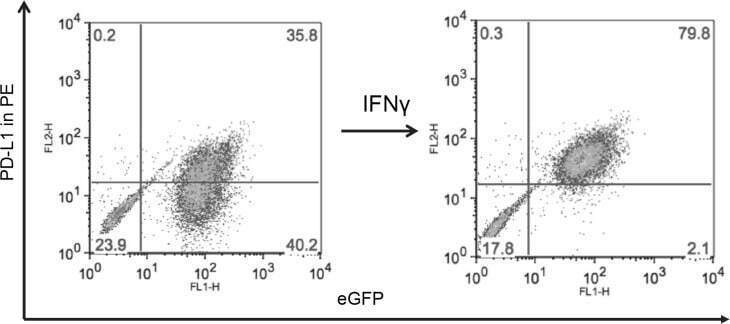
- Figure 6 Cutaneous (MEL13.03, MEL93.05 and A375) and conjunctival melanoma (CRMM1, CRMM2 and CM2005.1) cell lines express various levels of PD-L1 and PD-1 MEL13.03 is the positive control cell line for both PD-L1 and PD-1. Representative histograms show ( A ) PD-L1 and PD-1 ( B ) expression in cell lines with or without IFN-gamma (100 IU/ml) exposure for 48 h. Pink, blue and brown shaded histograms represent unstained, PD-L1 (PD-1) staining, and the effect of IFN-gamma stimulation on PD-L1 and PD-1, respectively.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
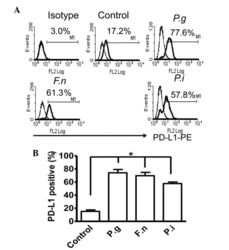
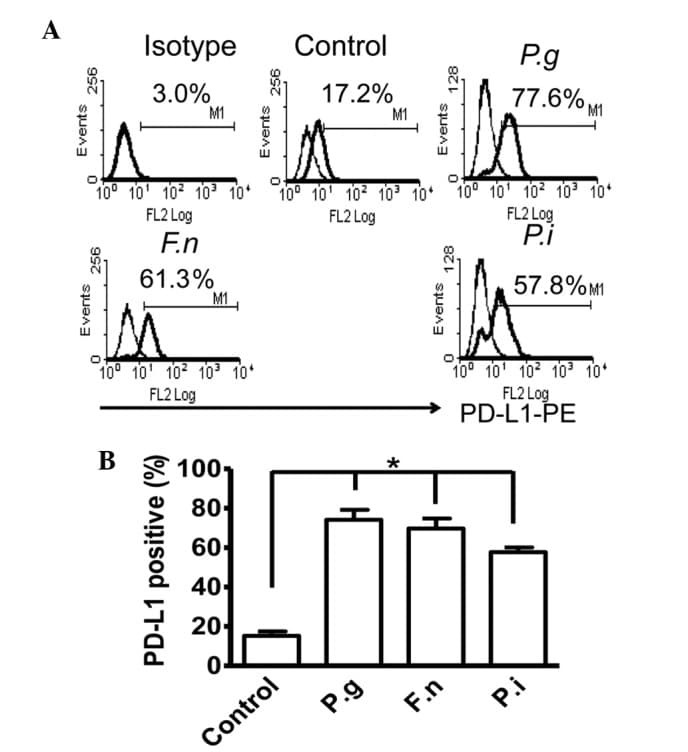
- Experimental details
- Figure 2 Common periodontal pathogens induce the expression of PD-L1 on PDLCs. (A) Flow cytometry histogram overlays of PDLCs co-cultured with P.g , F.n and P.i . (B) Comparison of expression levels of PD-L1 induced by P.g, F.n and P.i. Data are expressed as the mean +- standard error of the mean of three independent experiments. All three strains significantly increased the expression of PD-L1 on the PDLCs, * P
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
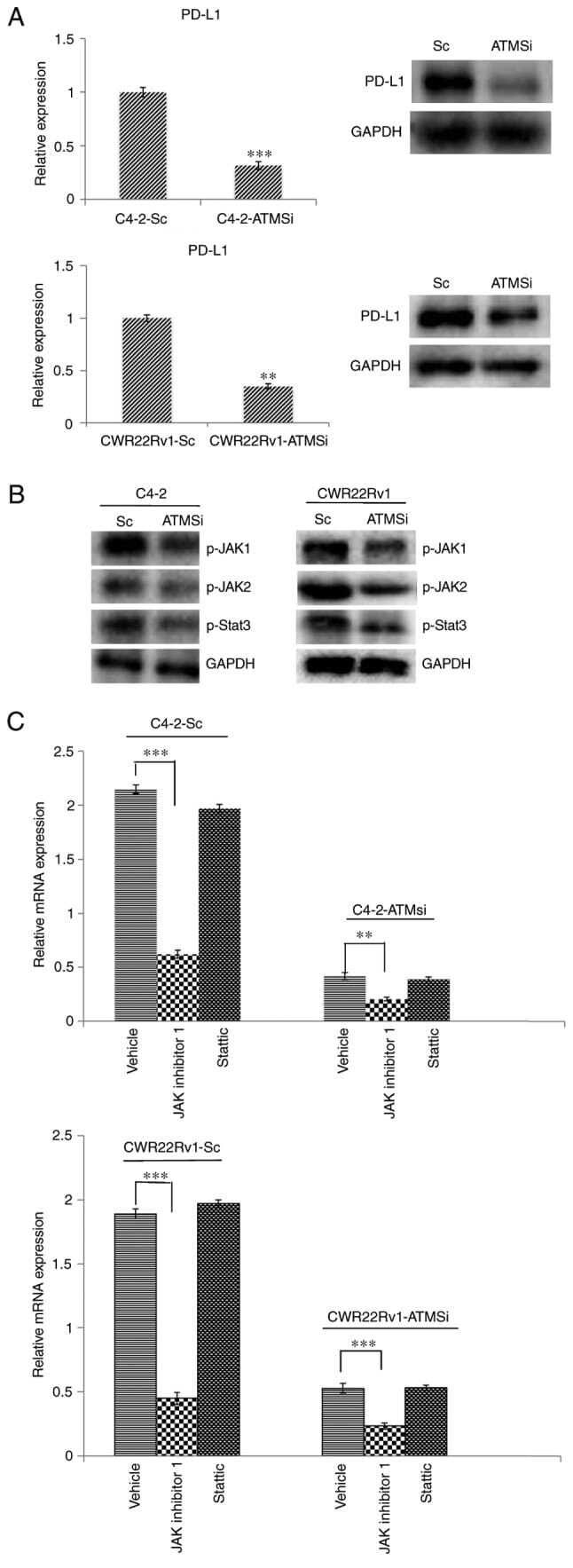
- Experimental details
- Figure 4. Levels of PD-L1, p-JAK1, p-JAK2 and p-STAT3 are suppressed in C4-2-ATMSi and CWR22Rv1-ATMSi cells compared with in the control cells, and JAK inhibitor 1 significantly suppresses the expression of PD-L1 in ATM knockout groups and control groups. (A) A significant decrease in PD-L1 expression was revealed in experimental groups by reverse transcription-quantitative polymerase chain reaction and western blotting. (B) Decreased levels of p-JAK1, p-JAK2 and p-STAT3 were revealed in the experimental groups by western blotting. (C) JAK inhibitor 1 and Stattic were used to treat all cell groups. Downregulation of JAK significantly reduced PD-L1 expression, whereas Stattic had no significant effect on PD-L1 expression. **P
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
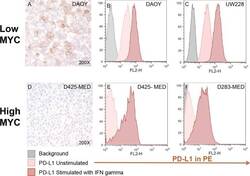
- Figure 4 Medulloblastoma cell lines express PD-L1 in a subgroup dependent manner PD-L1 expression in low and high MYC MB cell lines. Paraffin embedded cell line pellets stained for PD-L1 in DAOY ( A ) and D425-MED ( D ). Histograms of PD-L1 expression by flow cytometry using phycoerythrin (PE) conjugated MIH1 clone of PD-L1 (EBioscience) with and without IFN-gamma stimulation in DAOY ( B ), UW228 ( C ), D425-MED ( E ), and D283-MED ( F ).
- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
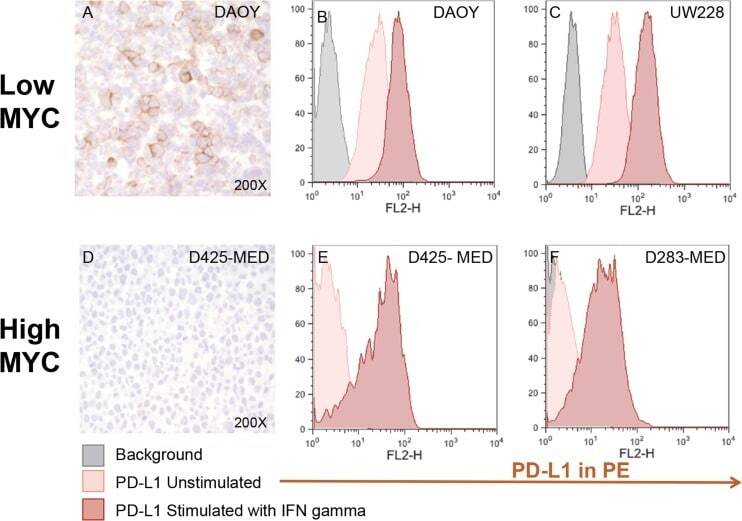
- Figure 5 MYC overexpression in DAOY (YM21) does not alter PD-L1 expression Dot Plots of PD-L1 expression by flow cytometry using PE-conjugated MIH1 clone of PD-L1 (EBioscience) in YM21 construct made by overexpressing MYC in DAOY via stable lentiviral transfection.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
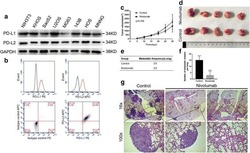
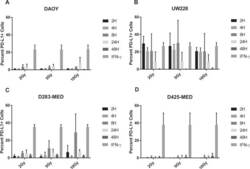
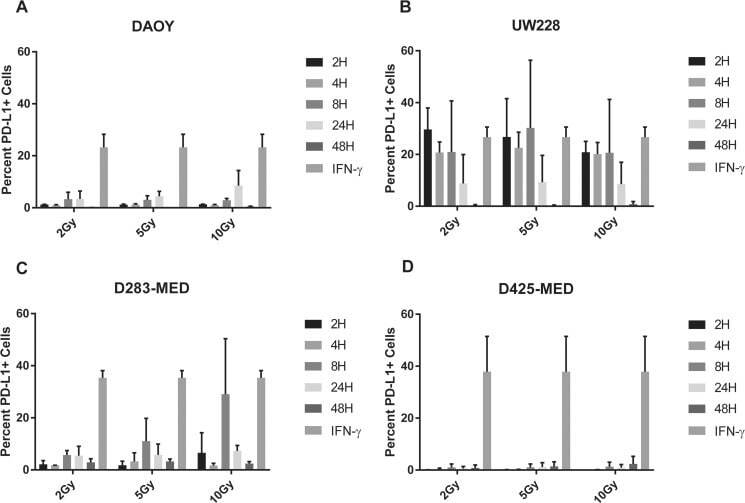
- Figure 6 Radiation induces PD-L1 in most medulloblastoma cell lines Bar graph depicts percent PD-L1 positive cells above baseline as determined by flow cytometry using PE-conjugated MIH1 clone of PD-L1 (EBioscience) in DAOY ( A ), UW228 ( B ), D283-MED ( C ), and D425-MED ( D ) after irradiation with 2, 5, or 10 Gy at 2, 4, 8, 24, and 48 hours. IFN-gamma values were obtained without radiation. Each bar graph depicts findings from 2 different experiments where all data points were repeated. Error bars represent standard deviation of the mean. Levels of PD-L1 that were comparable to those induced by IFN-gamma included UW228 hours 2-24 at 2, 5 and 10 Gy and D283-MED at hour 8, 10 Gy. All other values were significantly less than those induced by IFN-gamma as determined by 2-way ANOVA corrected for multiple comparisons. Full statistical analysis available in Supplementary Table 1-1-1-4 .
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
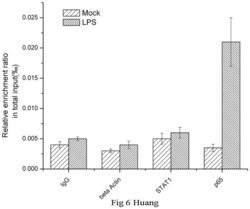
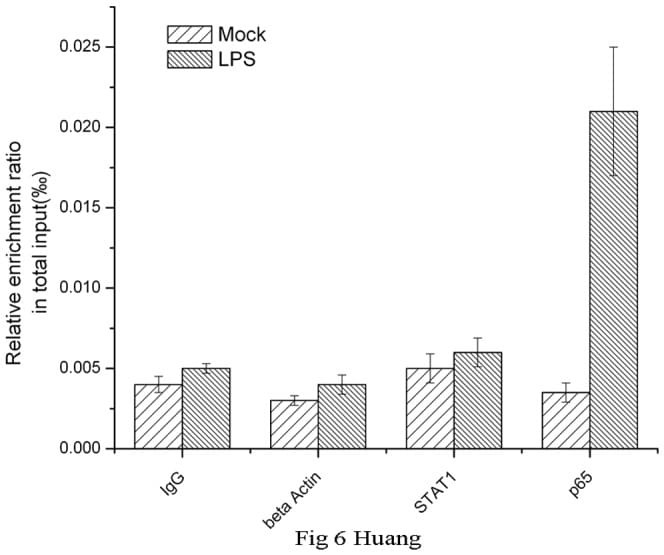
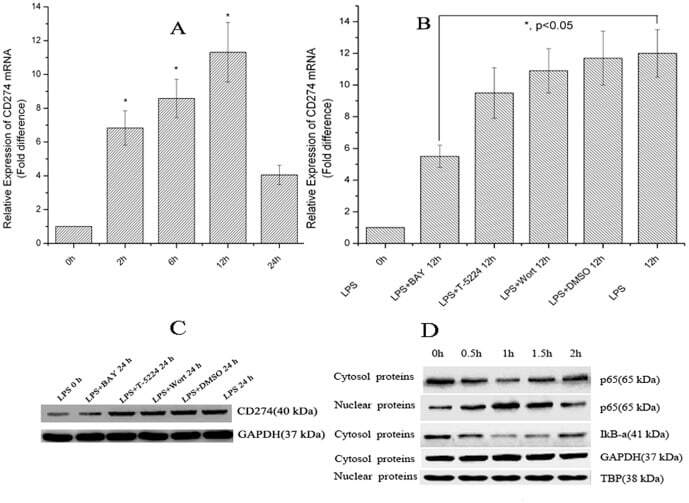
- Figure 6 Chromatin immunoprecipitation (ChIP) assays of the CD274 promoter in primary human monocytes. Cells were treated with or without LPS for 1 h. ChIP assays were carried out using an anti-p65 antibody. IgG, anti-beta-actin and anti-STAT1 antibodies were used as negative controls. Relative enrichment of each transcription factor-bound DNA was detected by qPCR using ChIP primers. All the results were normalized to input DNA.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
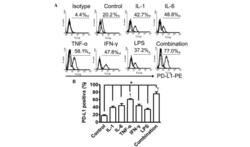
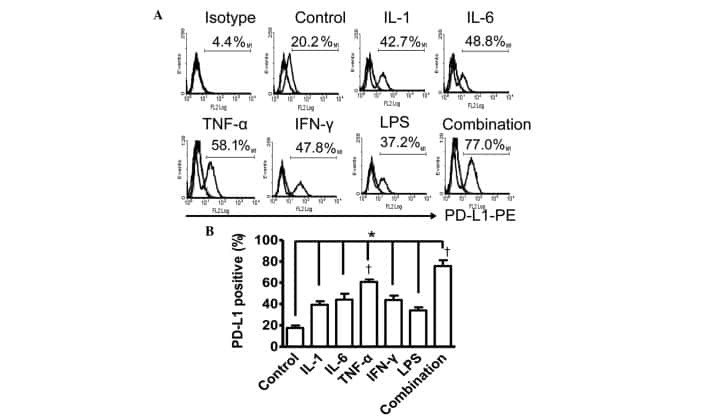
- Figure 1 LPS treatment regulates CD274 mRNA levels mainly via the NF-kappaB signaling pathway in primary human monocytes. CD274 mRNA levels were determined by quantitative real-time PCR and are shown as fold changes in arbitrary units. (A) LPS treatment regulates CD274 mRNA levels (n = 3, repeated three times, *P
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
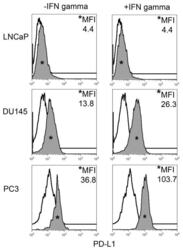
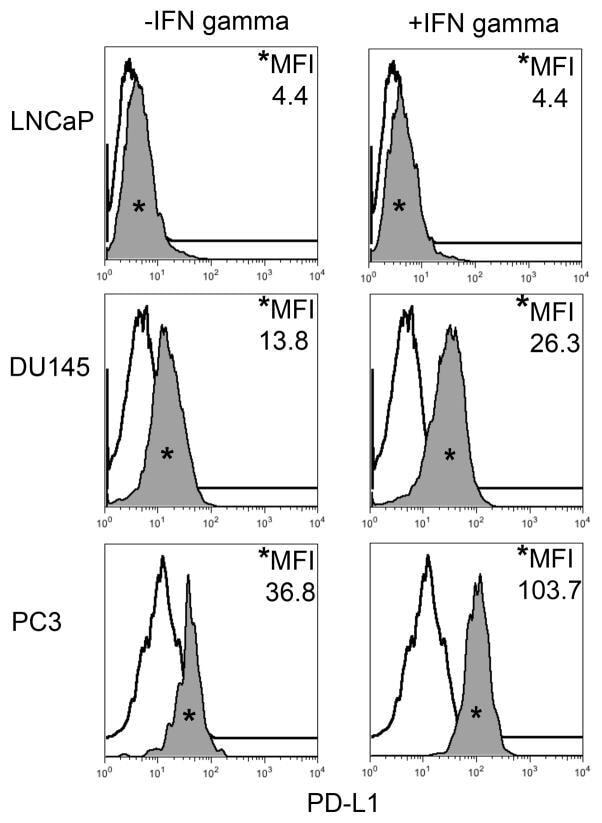
- FIGURE 1 Human Prostate Cancer Cell Lines Display Varying Expression of PD-L1 in Response to IFN-gamma Histograms representing PD-L1 surface expression as detected by flow cytometry in human prostate cancer cell lines with and without exposure to IFN-gamma. Open histograms represent cells stained with an isotype IgG1 antibody tagged with a matching PE fluorochrome. Shaded histograms represent PD-L1. Quantitative measurements reflect mean fluorescence index. LNCaP does not express PD-L1 at rest or in response to IFN-gamma. DU145 expresses PD-L1 at rest and has little response to IFN-gamma indicating an innate immune resistance phenotype. PC3 expresses PD-L1 at rest but robustly up-regulates this expression in response to IFN-gamma displaying a more classic adaptive immune resistance phenotype.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
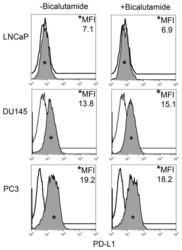
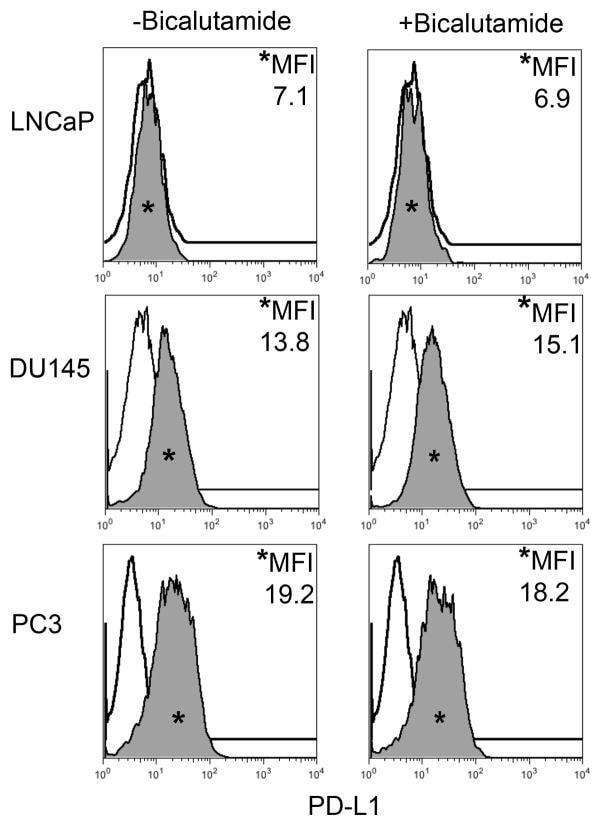
- FIGURE 2 Human Prostate Cancer Cell Lines Display No Change in PD-L1 Expression in Response to Bicalutamide Histograms representing PD-L1 surface expression as detected by flow cytometry in human prostate cancer cell lines with and without exposure to bicalutamide. Open histograms represent cells stained with an isotype IgG1 antibody tagged with a matching PE fluorochrome. Shaded histograms represent PD-L1. Quantitative measurements reflect mean fluorescence index. LNCaP, DU145, and PC3 displayed no changes in PD-L1 expression upon exposure to bicalutamide.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
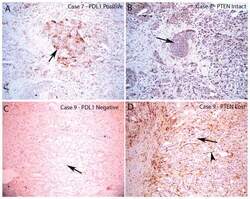
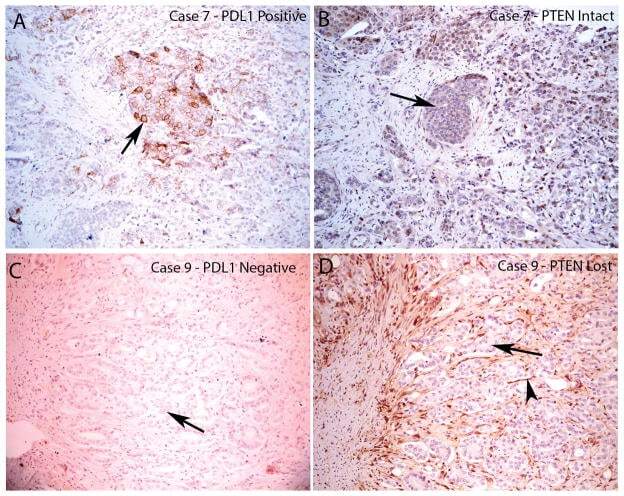
- FIGURE 4 Loss of PTEN Does Not Correlate with PD-L1 Expression in Resected Primary Prostate Carcinomas PD-L1 and PTEN staining in tumor lesions from two different patients. A. Case number 8 showing focally positive PD-L1 area. Arrow indicates positive tumor cell staining which is predominantly on the plasma membrane. B, similar region from adjacent section of case number 8 showing intact PTEN staining in all tumor cells (arrow shows a group of tumor cells with intact PTEN staining). C, case number 9 showing negative staining for PDL1 in all tumor cells (arrow shows tumor cells without PD-L1 staining). D, adjacent region showing PTEN loss in nearly all of the tumor cells (arrow shows tumor cells with negative PTEN staining; arrowhead shows stromal element with strongly positive PTEN staining). All images taken at 100 X original magnification.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
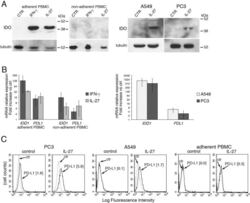
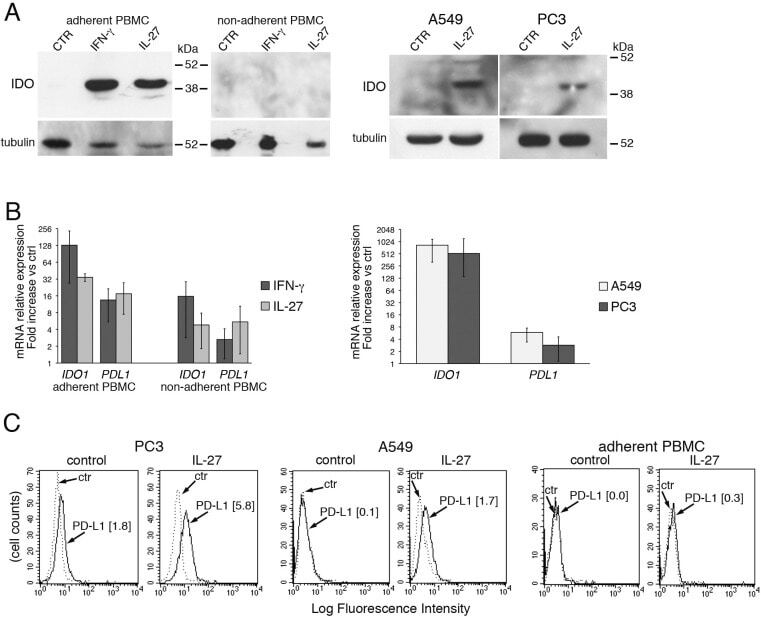
- Figure 7 IL-27 induces PD-L1 and/or IDO expression in human PC3 prostate and A549 lung cancer cells and adherent PBMC A. Western blot analysis of IDO expression in human adherent PBMC, non-adherent PBMC and in PC3 and A549 cells treated with the indicated cytokines or medium only for 48 hours. alpha-tubulin is used as loading control. Similar results were observed in two additional experiments. B. QRT-PCR analysis of IDO1 and PDL1 mRNA expression in cytokine-stimulated adherent or non-adherent PBMC, PC3 and A549 cells relative to untreated cells. Data are expressed as DeltaDeltaCT-fold change. Mean values of three independent experiments. Error bars represent SD. C. FACS analysis of surface PD-L1 in IL-27-treated or untreated PC3 and A549 cell lines and adherent PBMC. Dotted lines are isotype-matched unrelated Ig staining controls. Similar results were obtained in five different experiments (PC3: MFI 5.5 +- 2.7 vs 1.5 +- 0.6, mean +- SD in IL-27-treated vs un-stimulated cells, P = 0.03 by paired Student's t test; A549: MFI 2 +- 1 vs 0.43 +- 0.4, P = 0.01).
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
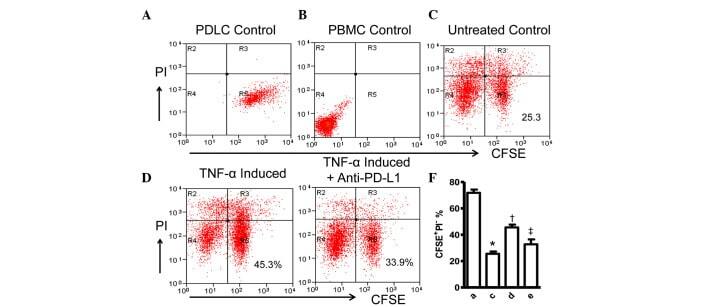
- Figure 4 Expression of PD-L1 on PDLCs improves survival of PDLCs. Flow cytometry histrograms of (A) PDLCs, (B) PHA-activated PBMCs, (C) PDLCs co-cultured with activated PBMCs, (D) PDLCs pretreated with TNF-alpha and co-cultured with activated PBMCs, and (E) PDLCs pretreated with TNF-alpha, and incubated with activated PBMCs and anti-PD-L1 antibodies. (F) Comparison of PDLC survival, according to the percentages of CFSE + /PI - cells. a, c, d and e represent the PDLC control, untreated control, TNF-alpha induced and TNF-alpha induced+anti-PD-L1 groups, respectively. Data are expressed as the mean +- standard error of the mean of three independent experiments. Co-culturing the activated PMBCs with untreated PDLCs resulted in a significant decrease in viable PDLCs ( * P
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
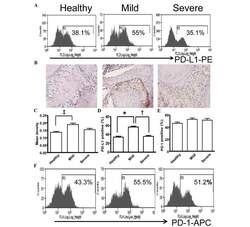
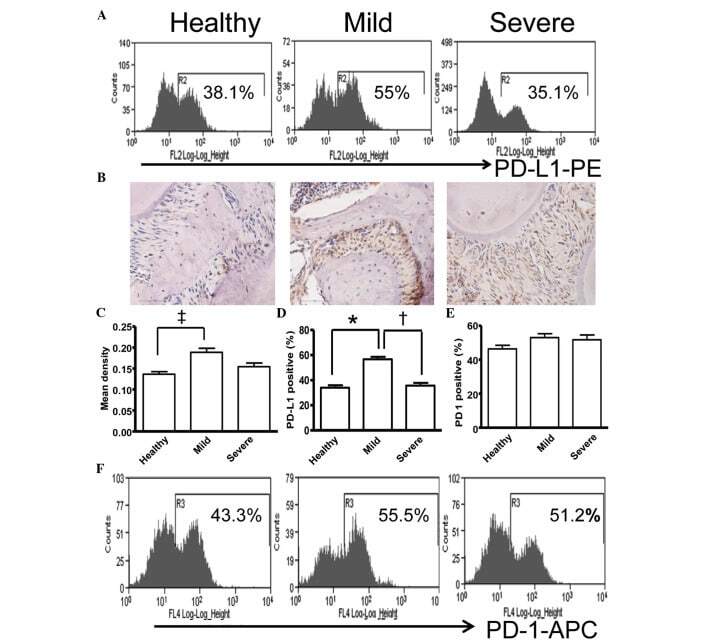
- Figure 5 Expression of PD-L1 is correlated with the severity of periodontitis in the experimental periodontitis model. (A) Flow cytometry histograms of the expression of PD-L1 on the surface of periodontal tissue cells from healthy mice, and mice with mild periodontitis and severe periodontitis. (B) Expression of PD-L1 in periodontal tissues from the three groups, detected by immunohistochemical staining (magnification, x400). (C) Comparison of the expression of PDL-1 in periodontal tissues between the three groups ( ++ P
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
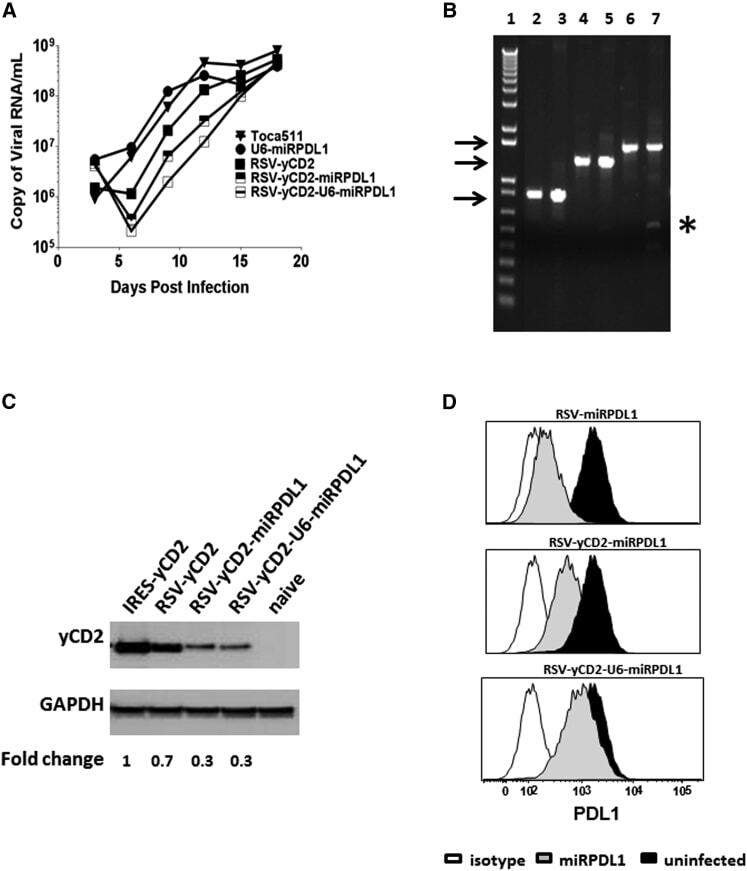
- Figure 5 RRV-RSV-yCD2-miRPDL1 and RRV-RSV-yCD2-U6-miRPDL1 Express yCD2 Protein and Exhibit PDL1 Downregulation Activity (A) Replication kinetics of RRV-RSV-yCD2-miRPDL1 and RRV-RSV-yCD2-U6-miRPDL1. The viral genome in the supernatants of infected LN-18 cells (MOI of 0.1) at indicated time points were quantified by qRT-PCR using primer set targeted to the env region ( Figure 1 ). A paired t test was performed and showed no statistically significant difference in replication kinetics between RRV-RSV-yCD2-miRPDL1 versus RRV-RSV-yCD2 (p = 0.0649) and RRV-RSV-yCD2-U6-miRPDL1 (p = 0.0801). RRV-yCD2, RRV-RSV-yCD2, and RRV-miRPDL1 (indicated as U6-miRPDL1 in the graph) were included as positive controls. (B) Vector stability of RRV-RSV-yCD2-miRPDL1 and RRV-RSV-yCD2-U6-miRPDL1 in LN-18 cells was analyzed by endpoint PCR at 14 days post infection. Lane 1: DNA molecular marker (1 Kb Plus marker, Invitrogen); lanes 2, 4, and 6 are positive controls using the corresponding plasmid DNA as the templates; lane 3: RRV-RSV-miRPDL1; lane 5: RRV-RSV-yCD2-miRPDL1; and lane 7: RRV-RSV-yCD2-U6-miRPDL1. The arrows indicate the expected size of the PCR products (844 bp for RRV-RSV-miRPDL1; 1,326 bp for RRV-RSV-yCD2-miRPDL1; and 1,591 bp for RRV-RSV-yCD2-U6-miRPDL1). (C) yCD2 protein expression in LN-18 cell infected with RRV-yCD2, RRV-RSV-yCD2, RRV-RSV-yCD2-miRPDL1, RRV-RSV-yCD2-U6-miRPDL1, and naive cells. GAPDH is included as loading control. The numbers shown on the bottom of the immunoblot indica
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
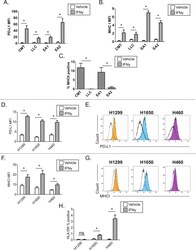
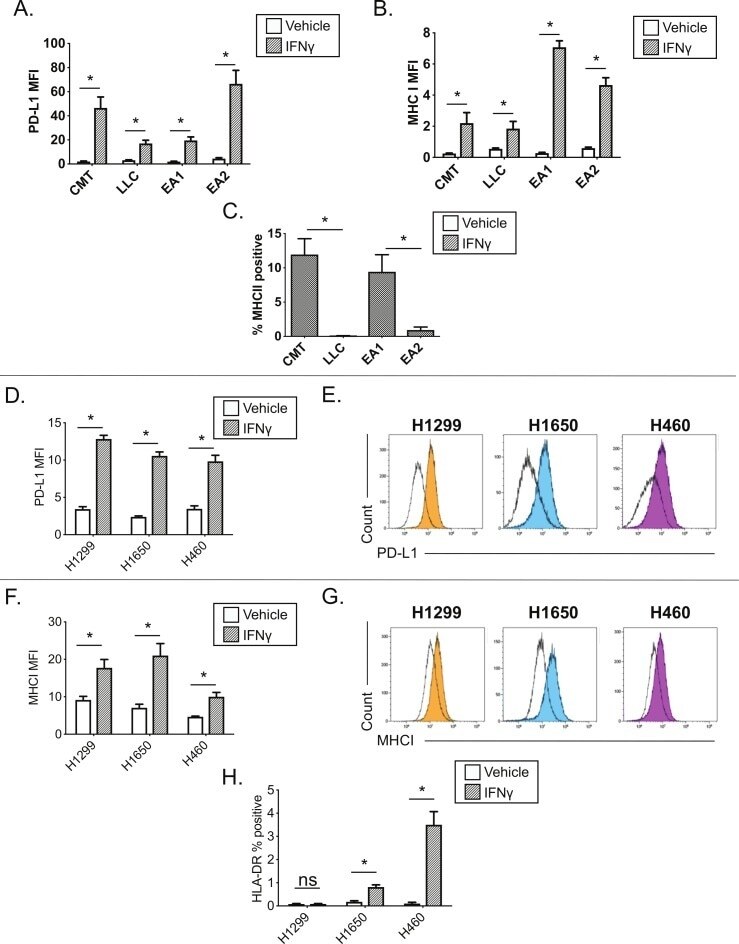
- Figure 2 Interferon gamma responsiveness in mouse and human NSCLC cell lines. In A-C, cultured cell lines (CMT167, LLC, EA1, EA2) were treated with vehicle or 100 ng/mL IFNgamma for 48 hours, and analyzed by flow cytometry for cell surface expression of (A) PD-L1, (B) MHC I, or (C) MHC II. Data for PD-L1 and MHC I are defined by median fluorescent intensity (MFI) of all cells, given the uniform induction of these proteins ( online supplementary figure 1 ). MHC II expression is defined as the frequency of positive cells, based on MHC II induction in a subset of cells ( online supplementary figure 2 ). Statistical analysis in (A, B) defined between vehicle and IFNgamma treated samples using an unpaired T-test (statistical significance identified by *p
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
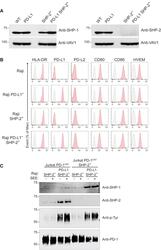
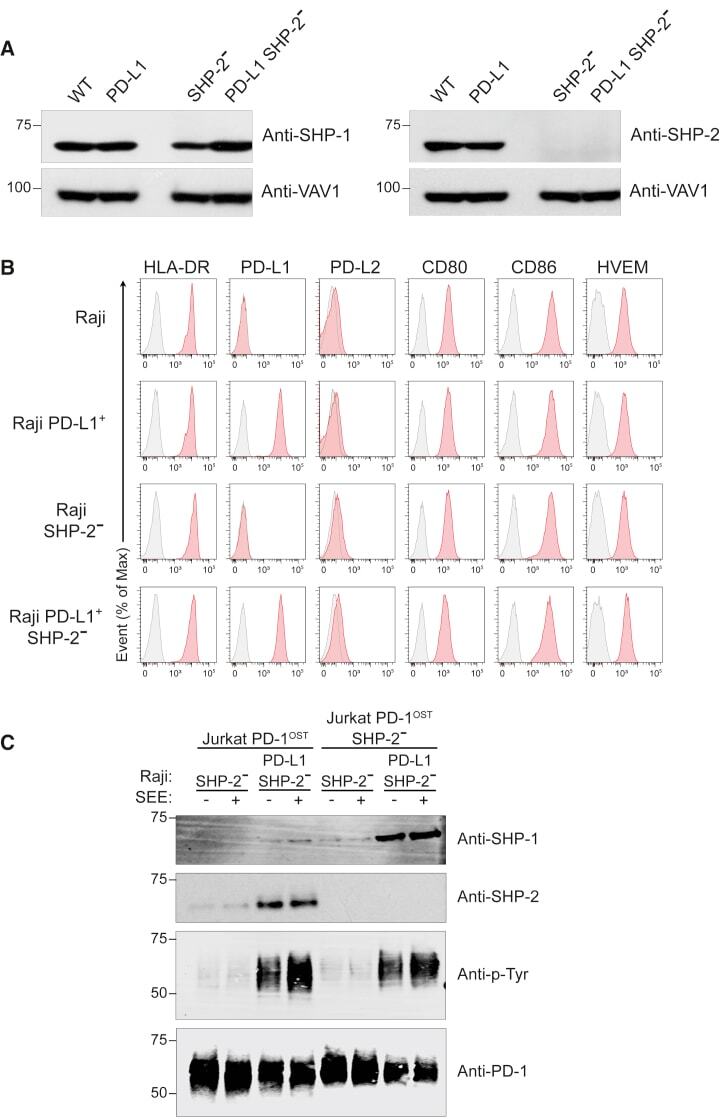
- Figure 6 SHP-1 Can Replace SHP-2 for PD-1 Coinhibition in Jurkat T Cells (A) Immunoblot analysis of equal amounts of proteins from total lysates of Raji (WT), Raji PD-L1, Raji SHP-2 - , and Raji PD-L1 SHP-2 - cells probed with antibodies to SHP-1 (right panel), SHP-2 (left panel), or VAV1 (loading control). Left margin, molecular size in kilodaltons. Data are representative of two independent experiments. (B) Raji, Raji PD-L1, Raji SHP-2 - , and Raji PD-L1 SHP-2 - cells were analyzed using flow cytometry for expression of HLA-DR, PD-L1, PD-L2, CD80, CD86, and HVEM. Gray shaded curves correspond to isotype-matched control antibody (negative control), and data are representative of two independent experiments. (C) Jurkat-PD-1 OST and Jurkat-PD-1 OST SHP-2 - cells were stimulated with Raji SHP-2 - cells or Raji PD-L1 SHP-2 - cells that have been preincubated in the absence (-) or presence (+) of SEE and lysed for 2 min after the initial contact. Immunoblot analysis of equal amounts (90%) of lysates from the specified conditions subjected to affinity purification (AP) on Strep-Tactin-Sepharose beads, followed by elution of proteins with D-biotin, and probed with antibody to anti-SHP-1, anti-SHP-2, and phosphorylated proteins (Anti-p-Tyr). Also shown is immunoblot analysis of equal amounts (10%) of total lysates of the specified cells probed with anti-PD-1 antibody (loading control). Left margin, molecular size in kilodaltons (kDa). Data are representative of two independent exper
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
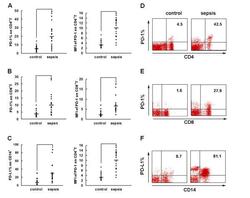
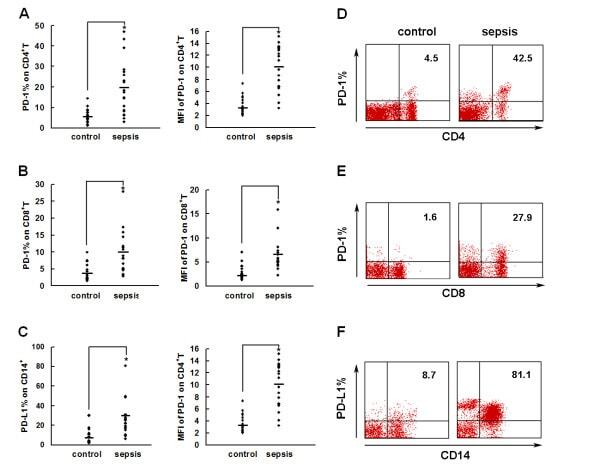
- Figure 2 PD-1 and PD-L1 were upregulated on T cells and monocytes in septic shock patients . Blood samples were obtained from 19 septic shock patients and 22 healthy controls and were stained for programmed death-1 (PD-1) and programmed death ligand-1 (PD-L1) gated on CD4 + T cells, CD8 + T cells, and CD14 + monocytes. (a) to (c) Percentage of PD-1 expression on (a) CD4 + T cells and (b) CD8 + T cells, and (c) percentage of PD-L1 expression on CD14 + monocytes. Each dot represents one individual. Data are mean +- standard error of the mean (SEM) of three independent experiments. ** P < 0.01 compared with healthy controls. (d) to (f) Mean fluorescence intensity (relative fluorescence units) of PD-1 expression on (d) CD4 + T cells, (e) PD-1 expression on CD8 + T cells, and (f) PD-L1 expression on CD14 + monocytes Each dot represents one individual. Data are mean +- SEM of three independent experiments. * P < 0.05 compared with healthy controls. (g) Representative PD-1 expression levels on CD4 + T cells and CD8 + T cells, and PD-L1 expression on CD14 + monocytes. Values in the upper-right quadrant indicate the percentage of cells that express PD-1 or PD-L1.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
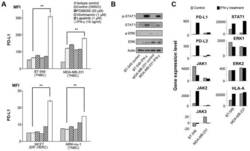
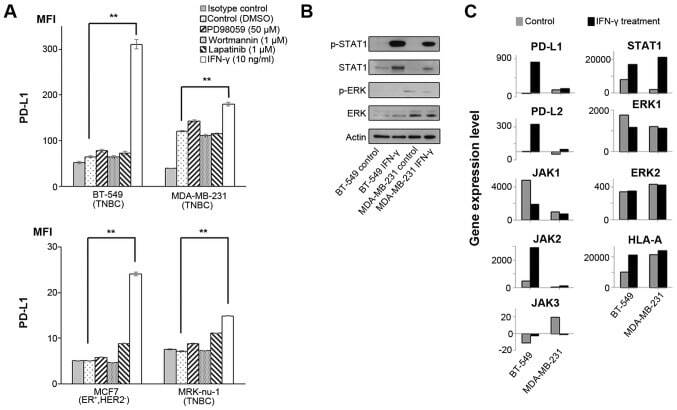
- Figure 1 Effects of IFN-gamma on PD-L1 and signaling pathways. (A) PD-L1 expression was measured by flow cytometry in the cell lines at 48 h following treatment with DMSO, which was used as a vehicle control, 50 u M PD98059 (MAPK inhibitor), 1 u M wortmannin (PI3K-AKT inhibitor), 1 u M lapatinib (combined epidermal growth factor receptor/human epidermal growth factor receptor 2 tyrosine kinase inhibitor), and 10 ng/ml IFN-gamma. Error bars represent the means +- SEM. ** P
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
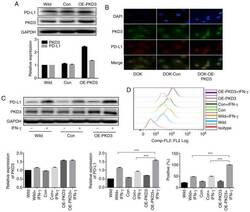
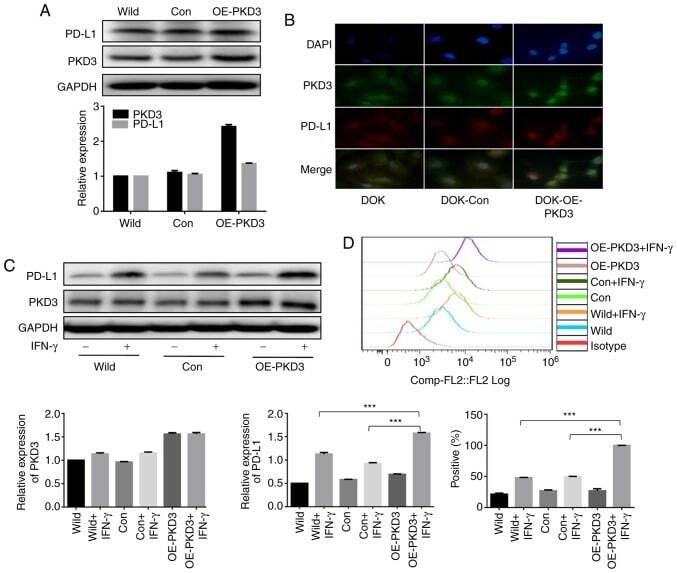
- Figure 3 PKD3 overexpression enhances IFN-gamma-induced PD-L1 expression in DOK cells. (A and B) The levels of PKD3 and PD-L1 protein were examined by western blot analysis and immunofluorescence. The PKD3 overexpression plasmid was transfected into DOK cells using Lipofectamine 2000. (C and D) Cells transfected with the PKD3 overexpression plasmid and cells transfected with control shRNA were cultured in the presence or absence of IFN-gamma (20 ng/ml) for 24 h. The expression levels of PKD3 and PD-L1 were determined by western blot analysis. In addition, the cell surface expression of PD-L1 was analyzed by flow cytometry. Data are presented as the means +- SD (n=3). *** P
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- Figure 3 Example flow cytometry plots demonstrating gating for PD-L1 positive cells THP-1 cells were (A and C) treated with IFN-gamma (50 ng/mL, approximate EC20 of PD-L1 activation), (B) left untreated, or (D) treated with IFN-gamma and a compound which we have identified that increases PD-L1 expression (unpublished data). Cells represented in (A) were not stained with the PE conjugated anti-PD-L1 antibody, whereas (B, C and D) were. All four groups of cells were otherwise stained (including fixable viability dye) and analyzed by flow cytometry as described in this protocol. All plots depict viable cells, gated as seen in Figure 2 .
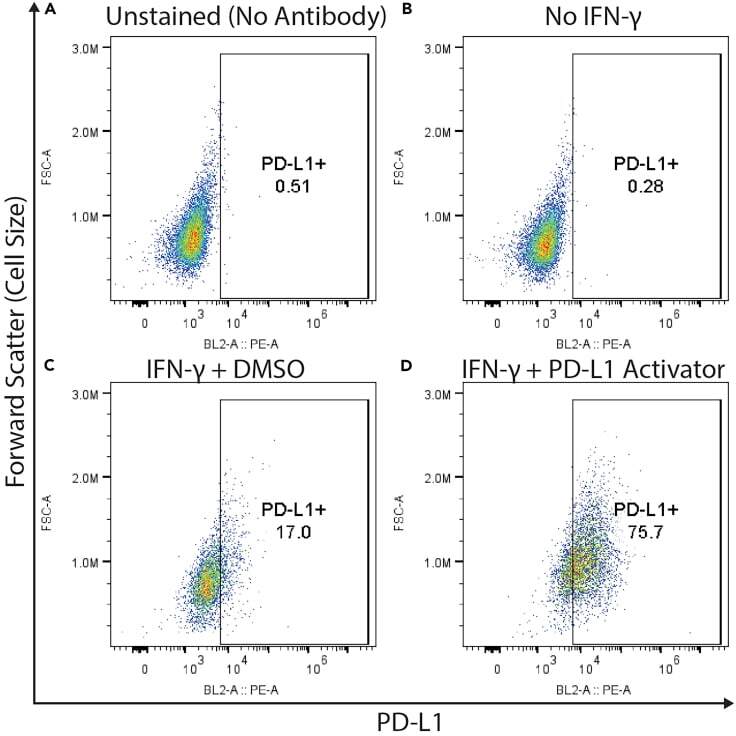
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
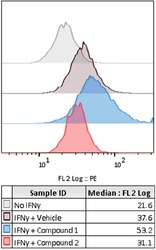
- Experimental details
- Figure 5 Representative control and compound treated wells Flow cytometry histogram plots showing PD-L1 expression (PE fluorescence signal) for wells treated with either No IFN-gamma, IFN-gamma and vehicle, or IFN-gamma and one of two compounds from a screening library which were identified as modulating PD-L1 expression.
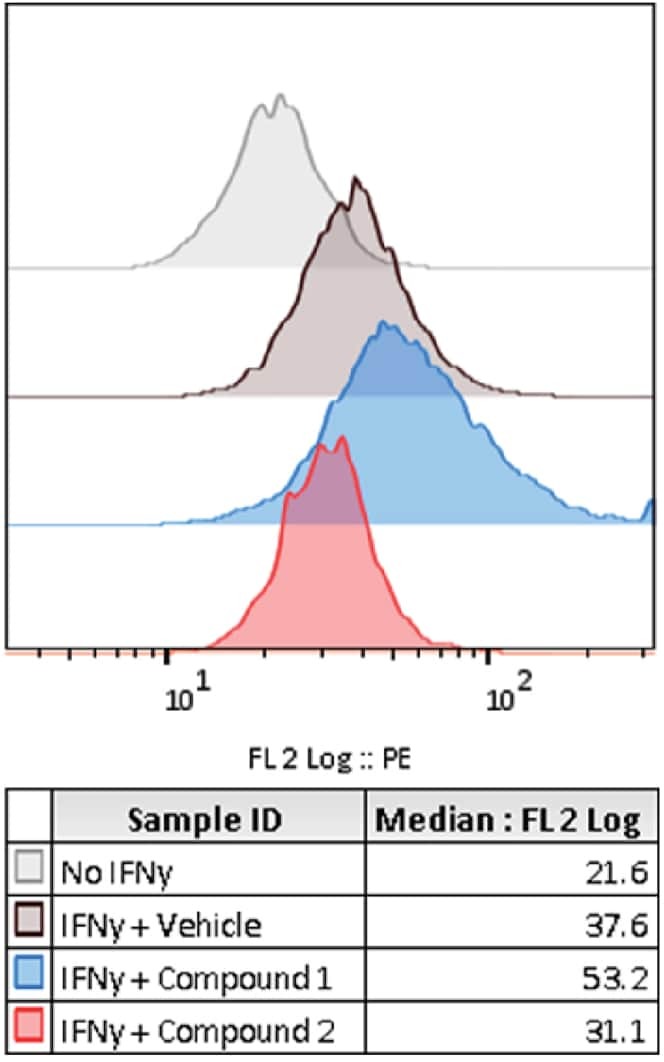
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
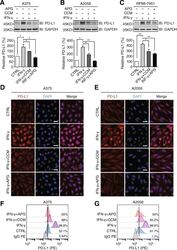
- Experimental details
- Fig. 3 Apigenin and curcumin suppress IFN-gamma-induced PD-L1 expression in human melanoma cells. a - c indicated melanoma cells were pretreated with DMSO, curcumin (25 muM), or apigenin (30 muM) for 4 h and then treated with IFN-gamma (10 ng/ml) for 24 h. Cells were lysed and PD-L1 expression was detected by Western blotting using a PD-L1 antibody. GAPDH was used as a loading control. Column charts below show quantitation data of relative PD-L1 expression. Error bars represent the mean +- S.D. ( n = 3, * P < 0.05 and ** P < 0.01). d and e cells were treated as described above and processed for immunofluorescence analysis. Representative micrographs show PD-L1 staining from different groups with same intensity settings on a fluorescent microscope (Olympus, Japan). DAPI stains nucleus. Scale bar = 25 mum. f and g , A375 and A2058 cells were treated as described above, and cell surface PD-L1 expression was determined by flow cytometry
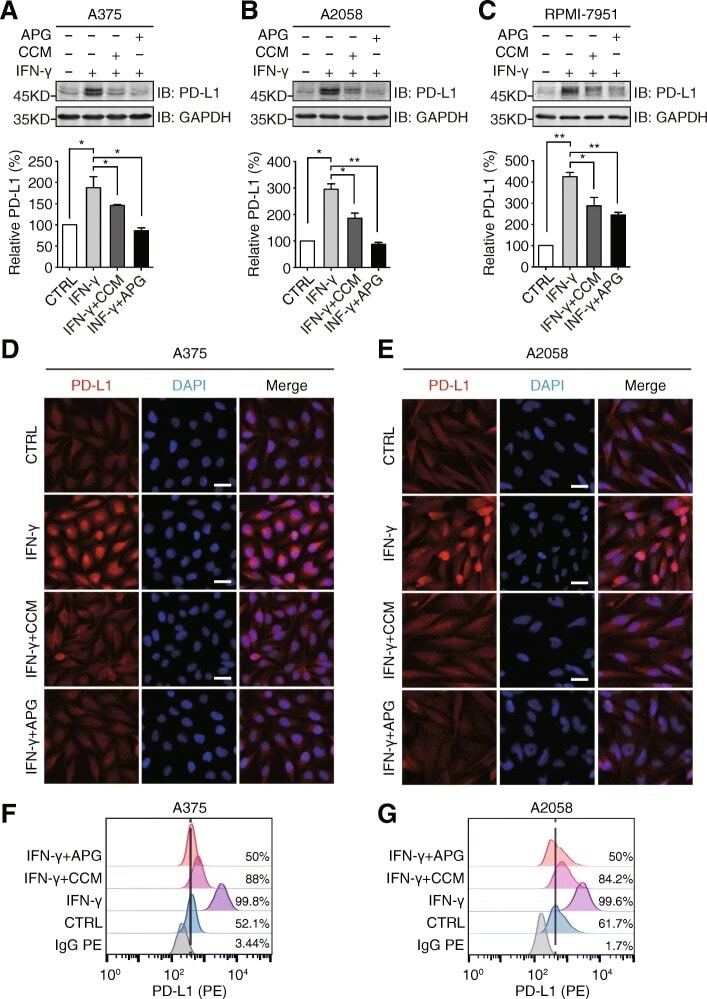
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
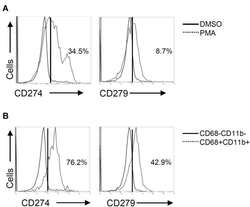
- Experimental details
- Figure 3 Phorbol 12-myristate 13-acetate (PMA) treatment induces PD-L1 (CD274) and PD-1 (CD279) expression in THP-1-differentiated macrophages. According to the experimental design shown in Figure 1A , immunostaining followed by flow cytometric histogram and dot-plot analysis showed the expression of PD-L1 (CD274) and PD-1 (CD279) in PMA-treated THP-1 cells ( A ) without or ( B ) with the gating of CD68 - CD11b - and CD68 + CD11b + cells. Representative data were selectively obtained from three individual experiments, and the percentage of positive cells is shown.
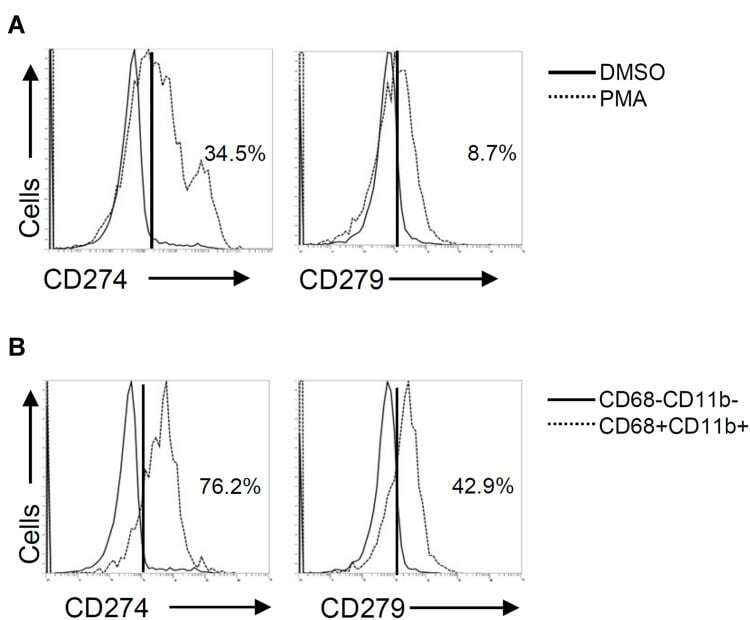
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- Figure 5 Different expression of PD-L1 (CD274) and PD-1 (CD279) in THP-1-differentiated macrophages. ( A ) In PMA-stimulated THP-1-differentiated M 0 macrophages, cells were then treated with LPS (1 mug/mL)/IFN-gamma (10 ng/mL) and IL-4 (25 ng/mL)/IL-13 (25 ng/mL) for polarization of M 1 and M 2 , respectively, in the absence and presence of neutralizing antibodies (5 mug/mL) against CD274 (alpha-PD-L1) and CD279 (alpha-PD-1) according to the experimental design. ( B ) For immunostaining, cells were stained with CD80 and CD209 for dissecting M 1 and M 2 , respectively. Immunostaining followed by flow cytometric histogram analysis showed the expression of CD274 and CD279 in these cells. ( C ) Furthermore, the expression of CD80 and CD209 in M 1 and M 2 without or with the blockade of CD274 and CD279 were shown. For all flow cytometric analysis, representative data were selectively obtained from three individual experiments, and the percentage of positive cells is shown. THP-1 (black); THP-1 M 0 (red); THP-1 M 1 (blue); THP-1 M 2 (green).
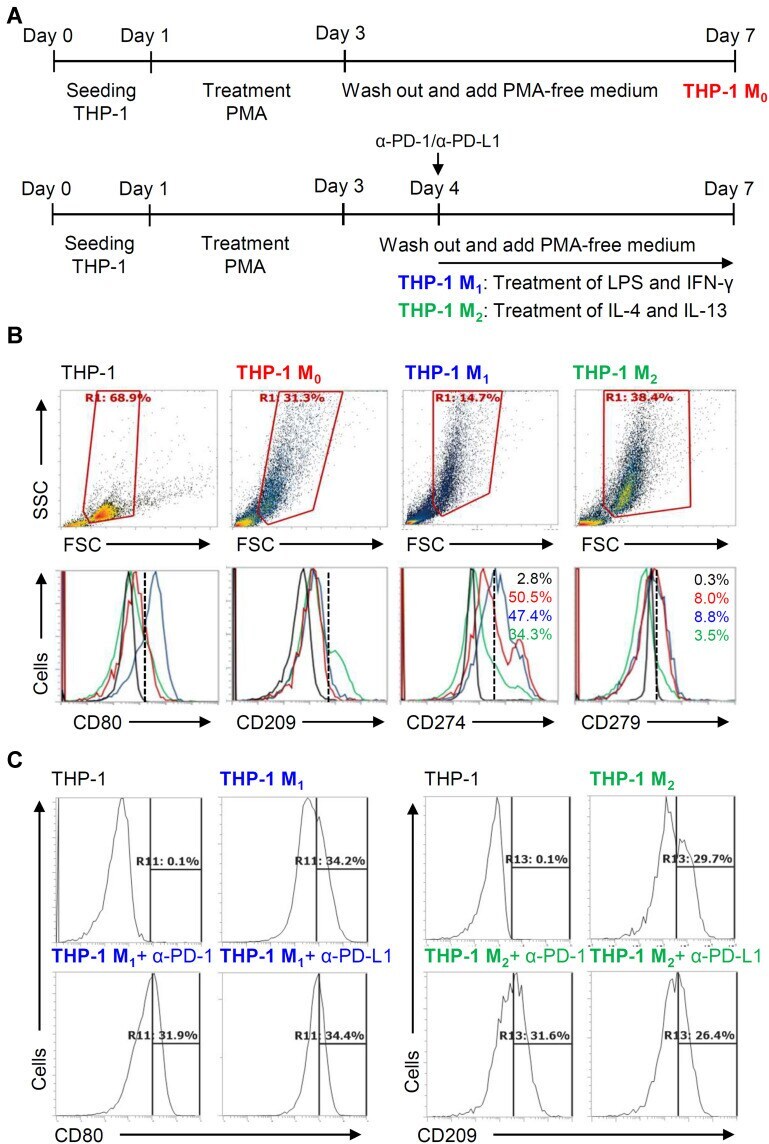
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- Figure 2 Correlation of PD-L1 cell surface amount with the transcript levels. The amount of PD-L1 on acute myeloid leukemia (AML) blast surface was determined using flow cytometry and expressed as the percentage of positive cells (left column) or as the mean fluorescence intensity (MFI) of the blast population (right column). The relative levels of v1 and v2 transcripts were measured by PCR and normalized to GAPDH.
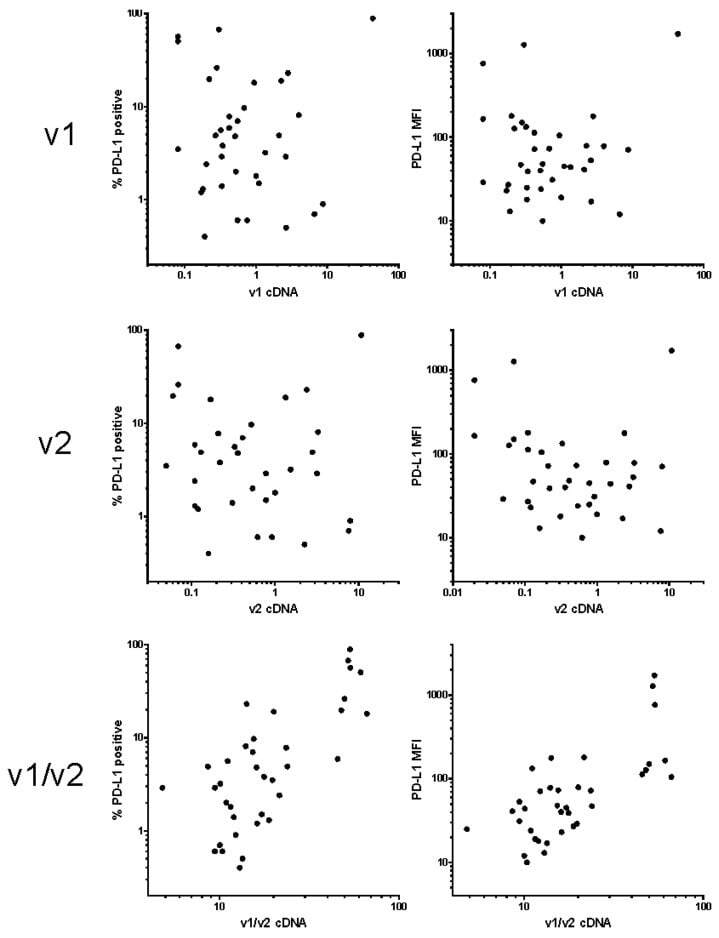
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
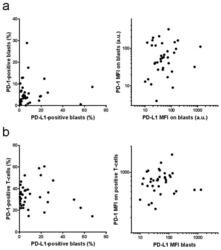
- Experimental details
- Figure 5 Correlation of PD-L1 and PD-1 expression. PD-L1 expression on AML blasts and PD-1 expression on AML blasts ( a ) or on the autologous T-cells ( b ) was determined by flow cytometry. Left: positive cell fractions, right: mean fluorescence intensity (MFI).
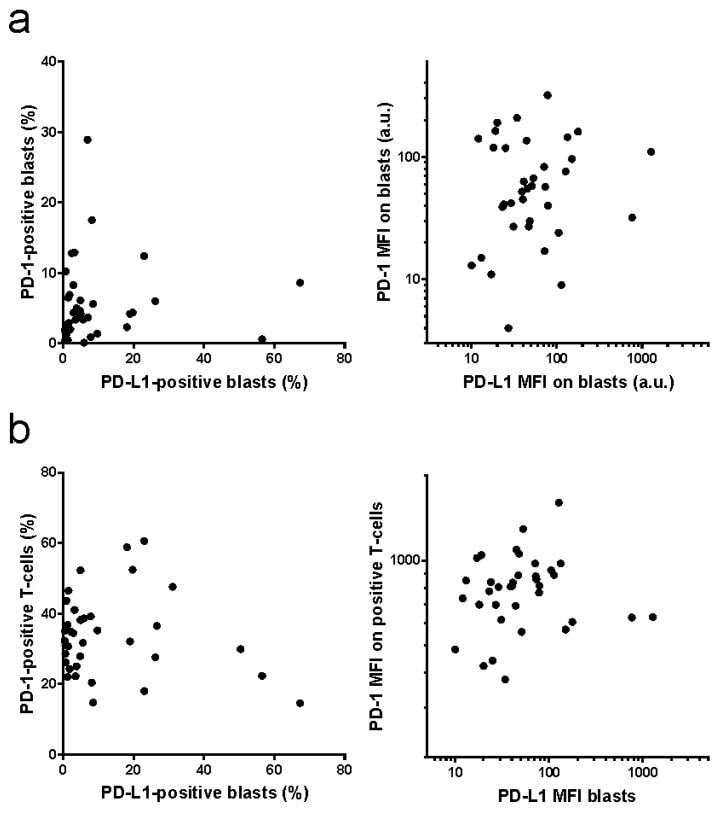
- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
- 1 Fig Expression of programmed cell death ligand-1 (PD-L1) is higher in patients with active multiple myeloma (MM) as compared to those with complete remission (CR). ( A ) Quantified polymerase chain reaction analysis of PD-L1 gene expression and ( B ) flow cytometric analysis of PD-L1 protein expression in bone marrow mononuclear cells (BMMCs) from patients with active MM. PD, progressive disease.
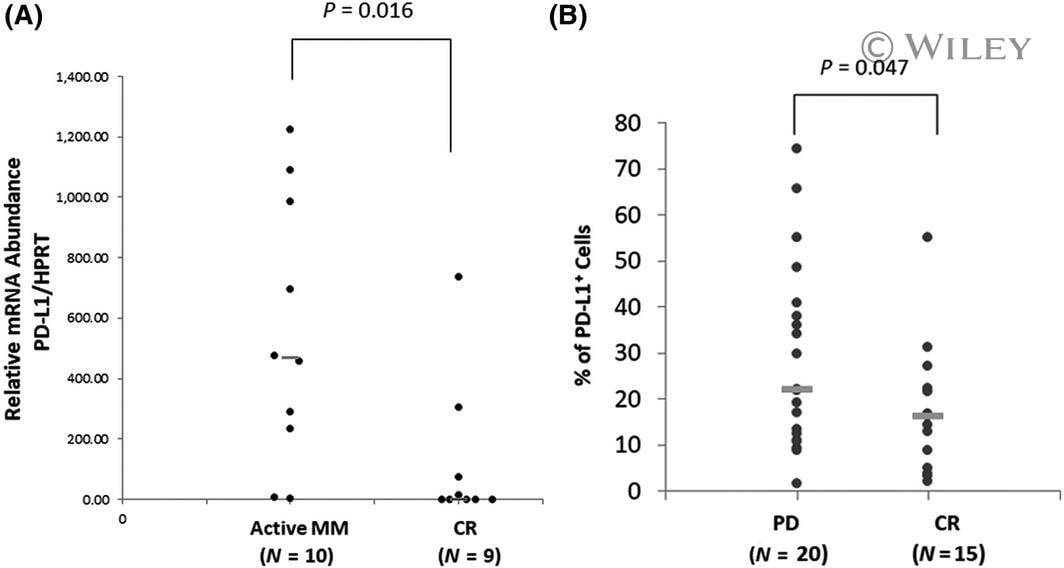
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
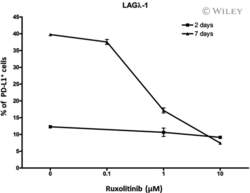
- Experimental details
- 5 Fig Ruxolitinib (RUX) reduces programmed cell death ligand-1 (PD-L1) expression in human multiple myeloma (MM) LAGlamda-1 cells. Flow cytometric analysis of percentage of PD-L1-postive cells in human MM xenograft LAGlamda-1 tumour cells treated with increasing concentrations of RUX for 2 or 7 days. All experiments were n = 3 (technical replicates).
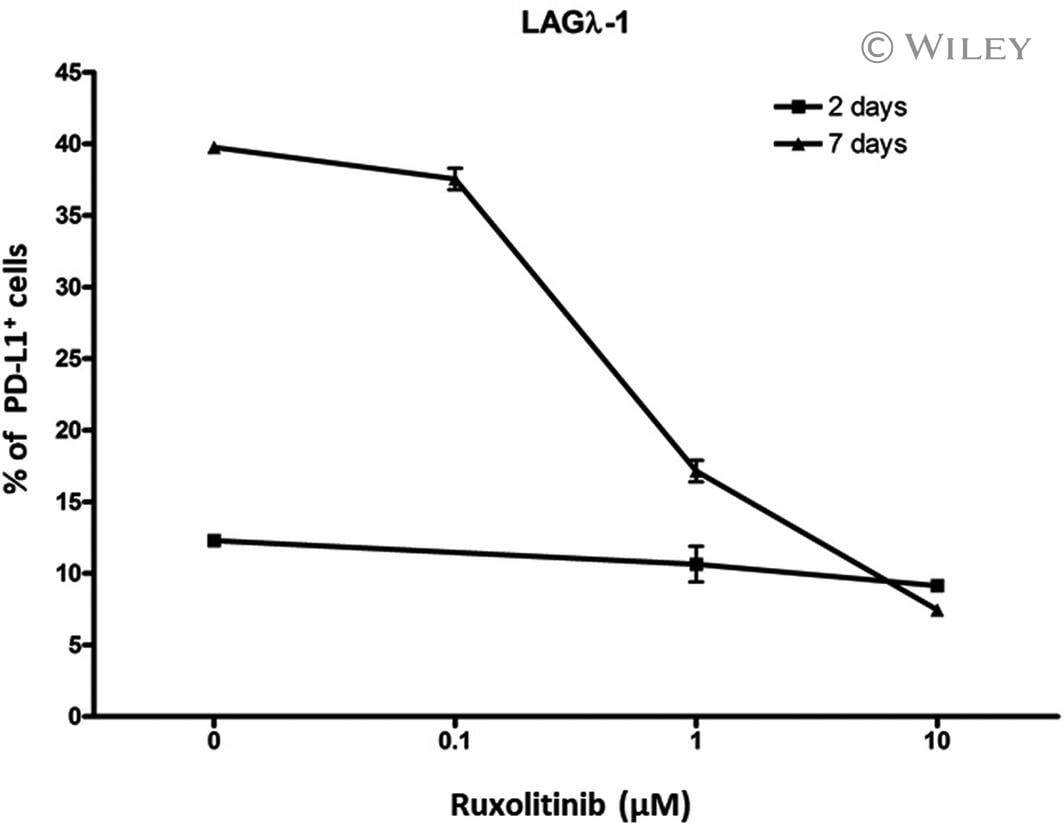
- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
- Figure 1 Azacytidine alters gene expression in NSCLC cell lines for multiple immune related pathways (A) Top panel: Gene Set Enrichment Analysis (GSEA) for pathways up-regulated by azacytidine. Normalized enrichment scores are plotted as a heat map. Bottom panel: boxplot showing degree of demethylation in each cell line, as measured by the difference in beta values between the AZA and mock-treated cells immediately after drug withdrawal and 7 days later. (B) FACS analysis shows increased level of cell surface PD-L1 after AZA treatment by day 10 in NSCLC lines H838 and H1299. (C) to (J) AZA-mediated expression changes at day 10 in key genes from pathways outlined in (A). Y axis = Ratio of expression values (log2) of AZA -treated vs. mock-treated cells; X-axis = gene names.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- Figure 3 TCR + PD-1 + 2D3 cells as a model assay for evaluation of involvement of PD-1 signaling in cell-mediated antigen-specific T-cell activation ( A - C ) WT1 (A, B) and gp100 (A, C) specific T-cell activation expressed as percentage viable CD8 + eGFP + PD-1 - and PD-1 + 2D3 cells (+- SEM) after 24 hours co-culture with different PD-L1 + stimulator cells. Neutralizing antibody against PD-1 (alphaPD-1; A, B; 15 ug/mL in WT1 model (A, B), 5 ug/mL in gp100 model (A)) or PD-L1 (alphaPD-L1; C) was added to cells 1 hour prior to co-culture to verify PD-1-mediated signaling, where indicated. (A) PD-1-dependent stimulating capacity of two differently generated peptide-pulsed mature monocyte-derived dendritic cells (WT1 (4 DC donors tested in two independent experiments); gp100 (4 DC donors in four independent experiments)). (B)Impact of induced PD-L1 expression on peptide-pulsed THP-1 leukemic cells on WT1-specific T-cell activation (6 replicate experiments). (C) gp100-specific PD-1 + T cell-activating capacity of peptide-pulsed wild-type or stably transduced PD-L1 + MCF-7 breast carcinoma cells (4 replicate experiments). Data information: in A, the horizontal line represents the median percentage eGFP expression ( n = 4). * P
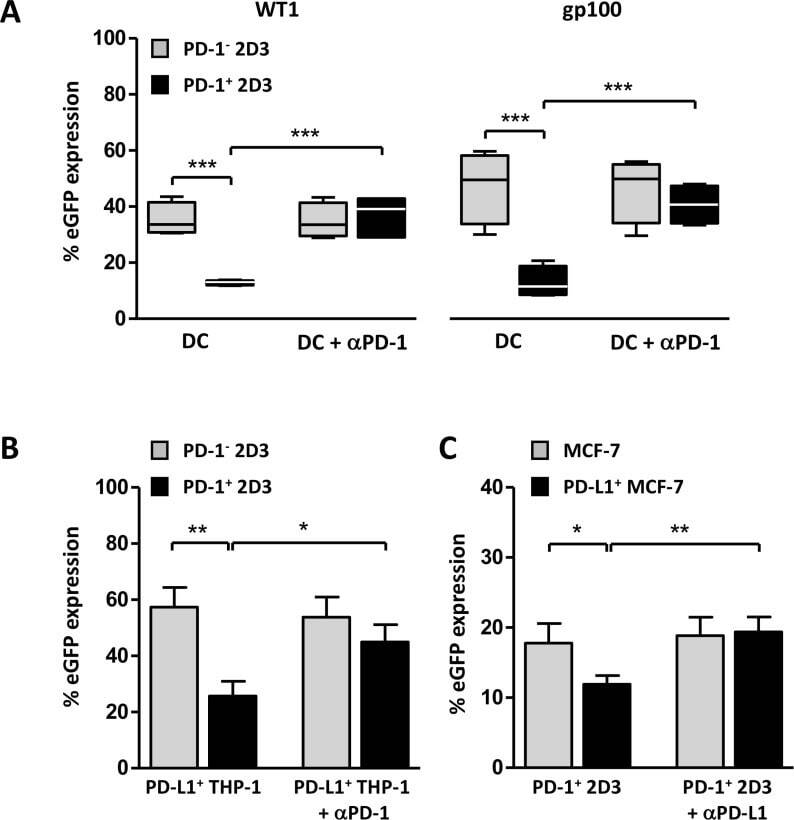
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
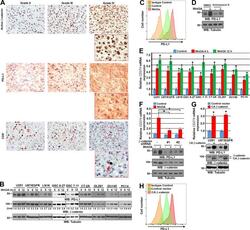
- Experimental details
- Figure 1. Wnt-induced beta-catenin activation results in PD-L1 up-regulation in tumor cells. (B, D, F, and G) Immunoblotting analyses were performed with the indicated antibodies. (E-G) Data represent the means +- SD of three independent experiments. (A) IHC staining of 20 diffuse astrocytoma (grade II), 20 anaplastic astrocytoma (grade III), and 50 GBM (grade IV) specimens was performed with the indicated antibodies. Representative images of IHC staining from the specimens are shown. Scale bar, 50 um. Red arrows point to CD8 + cells. (B) The indicated tumor cells were serum starved for 12 h and then stimulated with Wnt3A (20 ng/ml) for the indicated periods of time. Immunoblotting analyses were performed. (C) U87/EGFR cells were treated with or without Wnt3A (20 ng/ml) for 12 h. A cell surface analysis of PD-L1 protein was performed using a flow cytometer. (D) Serum-starved U87/EGFR cells were pretreated with or without actinomycin D (1 ug/ml) for 2 h and then stimulated with or without Wnt3A (20 ng/ml) for 12 h. (E) The indicated tumor cells were serum starved for 12 h and then stimulated with or without Wnt3A (20 ng/ml) for the indicated periods of time. Real-time PCR analyses were performed. *, P < 0.0001, on the basis of Student's t test. (F) U87/EGFR cells with stable expression of beta-catenin shRNA or a control shRNA were treated with or without Wnt3A (20 ug/ml) for 12 h. A real-time PCR analysis (top panel) and immunoblotting analyses (bottom panel) were performed. b
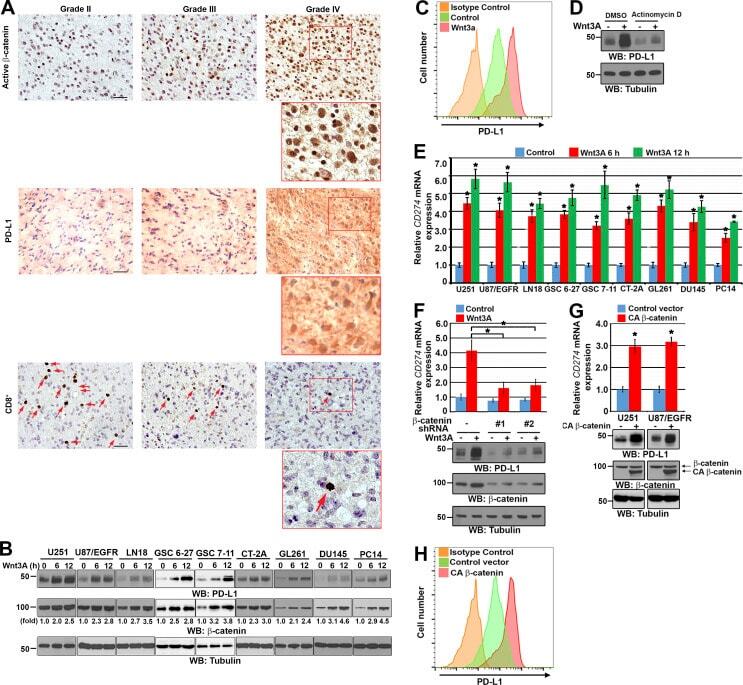
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- Extended Data Figure 5 TASL mirrors SLC15A4 requirement for TLR7/8 activation. ( a ) Fraction of R848-induced genes affected by SLC15A4 and TASL knockout, related to Fig. 2b . ( b ) Upset plot representing number of R848-induced genes commonly affected by indicated sgRNAs, related to Fig. 2b . ( c ) TASL gene expression levels in indicated THP1 cells, related to Fig. 2b . Bar graphs shown mean (n=3 biological replicates), error bars show 95 % confidence interval of mean. ( d ) Flow cytometry of PD-L1 surface expression in indicated unstimulated (ns) or R848-stimulated (5 mug/ml, 24h) THP1 cells. ( e ) Immunoblots of indicated THP1 DUAL cells. Lysates treated with PNGase F as indicated. ( f-i,k ) Indicated THP1 DUAL cells were (co-)treated for 24 hours with R848 (5 mug/ml), CL075 (5 mug/ml), ssRNA40/LyoVec(tm) (5 mug/ml) or inactive control ssRNA41/LyoVec(tm) (5 mug/ml), C12-iE-DAP (5 mug/ml), MDP (10 mug/ml), murabutide (10 mug/ml), Pam3CSK4 (0.1 mug/ml), Flagellin (0.1 mug/ml), cGAMP (3 mug/ml) or interferon beta (20 ng/ml). ( h ) CRISPR/Cas9 editing efficiency (%) estimated by TIDE. ( j ) Indicated THP1 DUAL cells were primed or not with interferon gamma (0.1 mug/ml) for 24h, washed and stimulated or not with MDP (10 mug/ml, 24h). ( f-k ) Supernatants were analyzed for ISRE and NF-kappaB reporter activity. Mean +- s.d. (n=3 biological replicates). ( l ) Relative mRNA expression of SLC15A4 , TASL or MYD88 in siRNA-transfected CD14 + monocytes in comparison to control (si CTR
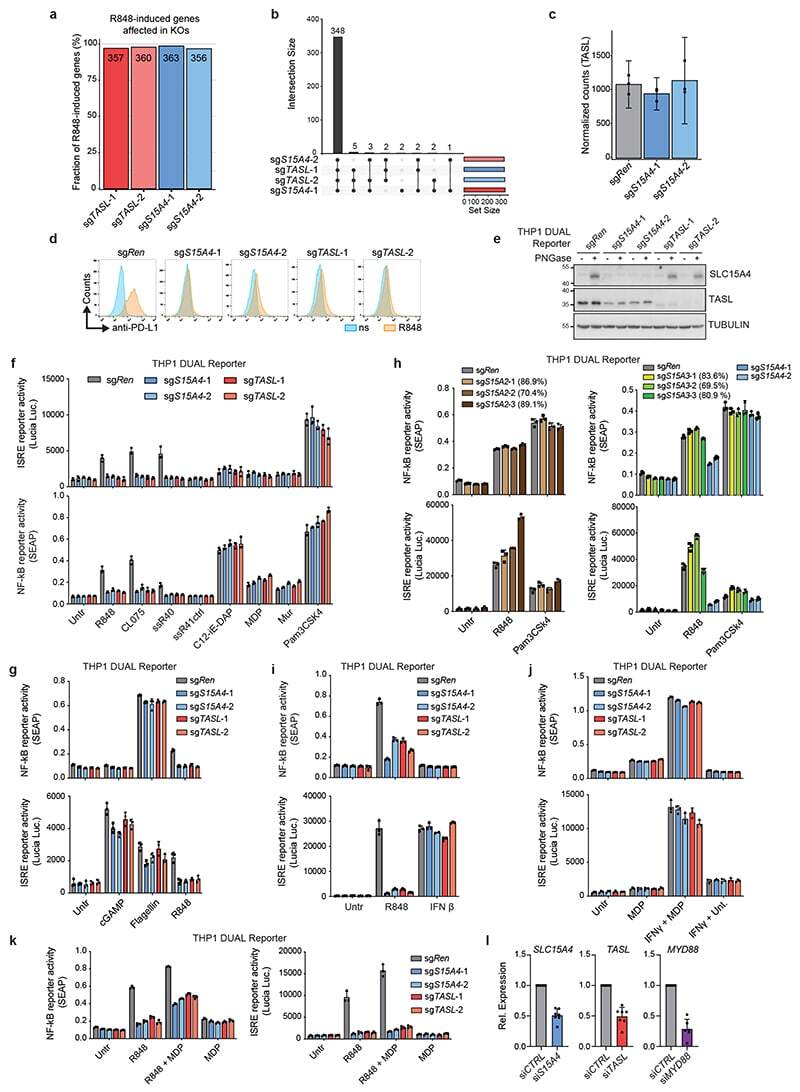
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
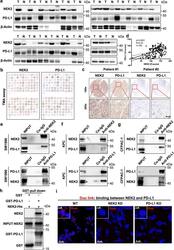
- Experimental details
- Fig. 4 NEK2 positively correlates and interacts with PD-L1 in pancreatic cancer. a Western blot analysis of NEK2 and PD-L1 in clinical pancreatic tissue samples from patients ( n = 20) (N: Normal pancreatic tissue; T: Pancreatic tumor tissue). b - d Representative images and statistical results of IHC staining of NEK2 and PD-L1 in a tissue microarray ( n = 156). e - g Cell lysates from SW1990, KPC, and CFPAC-1 separately analyzed by IP and Western blotting using the antibodies indicated. Representative image is shown n = 3 independent experiments. h GST-pull down assay of NEK2-His and GST-PD-L1 protein. Representative image is shown n = 3 independent experiments. i Representative images of individual immunofluorescence staining of NEK2 and PD-L1 interaction in KPC cells by Duolink assay. The red dots (NEK2/PD-L1 interaction) indicate their interaction. Representative image is shown n = 3 independent experiments. The Spearman correlations and p -values by Spearman's test are indicated in d .
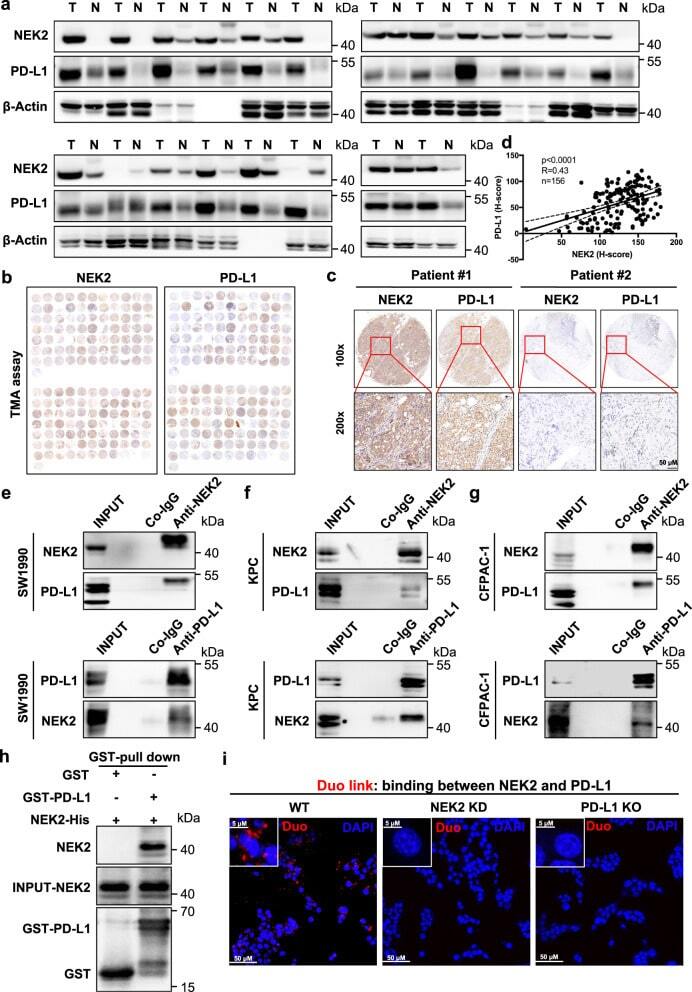
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
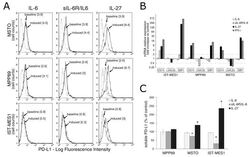
- Experimental details
- Figure 2 IL-27 upregulates PD-L1 molecule expression and release by MM cells. ( A ) Flow cytometry analysis of membrane PD-L1 expression in MSTO, MPP89, and IST-MES1 MM cell lines cultured with medium alone (baseline), IL-6, sIL-6R/IL-6 chimera, or IL-27 (induced). Dotted line shows isotype-matched Ig control. Numbers in brackets represent Median Fluorescence Intensity (MFI) values calculated as median anti-PD-L1 mAb minus median Ig control. Data are representative of two independent experiments yielding similar results. ( B ) qRT-PCR analysis of CD274 (PD-L1) mRNA expression in the same three MM cell lines stimulated with IL-6, sIL-6R/IL-6 chimera, IL-27, or IFN-gamma relative to untreated cells. Data are calculated with the DeltaDeltaCT method and expressed as fold change versus untreated control. Error bars represent SD of triplicates. LGALS3 (Galectin 3) and GBP1 (Guanylate Binding Protein 1) mRNA levels are shown, respectively, as negative and positive controls of IL-27 activity. ( C ) Evaluation by ELISA of soluble (s)PD-L1 release in the conditioned media of the indicated MM cells treated with IL-6, sIL-6R/IL-6 chimera, or IL-27. Data are expressed as a percent of untreated control and are the mean of two independent experiments, run in duplicates. Error bars represent the minimum and maximum values (* p < 0.001, Student's t -test versus untreated control).
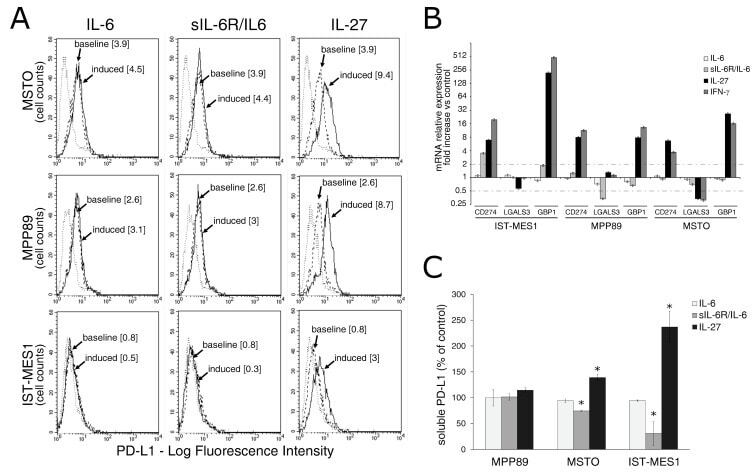
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
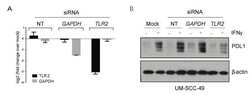
- Experimental details
- Figure 3 TLR2 depletion attenuates IFNgamma-mediated upregulation of PDL1 in UM-SCC-49 cells. ( A ) UM-SCC-49 cells were treated with siRNA as indicated, and knockdown of TLR2 and GAPDH RNA expression was tested by qPCR. ( B ) Cells were transfected with non-targeting (NT), GAPDH, or TLR2 siRNA for 24 h were then treated -/+ 10 ng/mL IFNgamma for 48 h, and protein expression was assessed by immunoblot as indicated. The uncropped Western blots have been shown in Figure S3 .
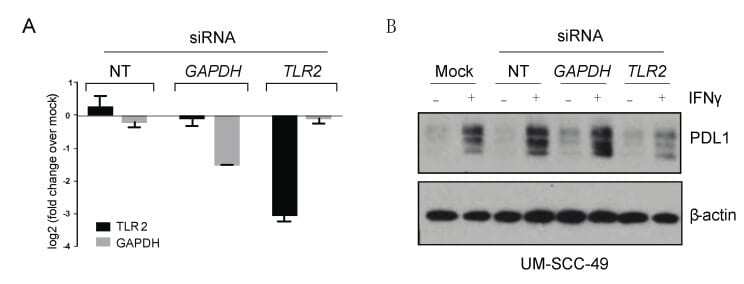
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- Figure 7 AIM binds directly to PSA, and the ectopic (re)expression of AIM enhances the anticancer therapeutic effect of Pembrolizumab against PCa cells. 2D visualization of the auto-generated ( A ) CD5L-KLK3 interactome showing complicity in the immune landscape, macrophages, EMT and CSC regulation, ( B ) CD5L interactome, or KLK3 interactome. ( C ) Three-dimensional visualization of the direct complex formation by AIM/CD5L (surface model) and PSA/KLK3 (ribbon model). ( D ) Line graph showing the effect of CD5L_OE with or without 5 muM Pembrolizumab on PC3 cell proliferation. ( E ) Representative photomicrographs ( upper ) and histograms ( lower ) showing the effect of CD5L_OE with or without 5 muM Pembrolizumab on PC3 cell invasion. ( F ) Representative photomicrographs ( upper ) and quantitative chart ( lower ) showing the effect of CD5L_OE with or without 5 muM Pembrolizumab on the tumorsphere formation capability of PC3 cells. ( G ) Representative IHC photomicrographs ( upper ) and histograms ( lower ) of the immunoreactivity of AIM, PSA, and PDL1 in pembro_responders or pembro_non-responders from the TMU-SHH cohort. ( H ) Histograms showing the differential CD8 + T-cell count/field in pembro_responders or pembro_non-responders from the TMU-SHH cohort. WT, wild type; OE, overexpression; * p < 0.05; ** p < 0.01; *** p < 0.001.
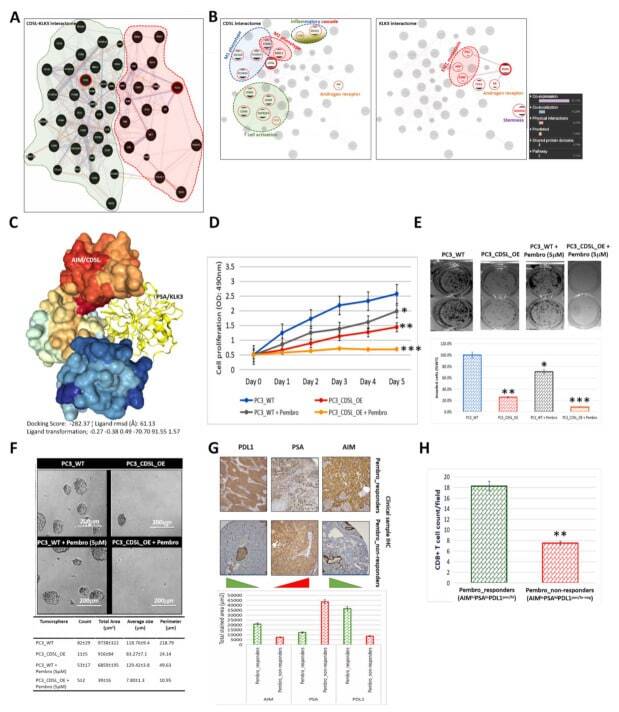
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
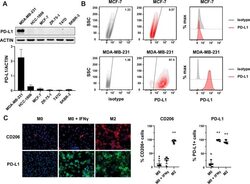
- Experimental details
- Figure 1 Expression of PD-L1 in breast cancer cell lines and macrophages. (A) The representative western blot analysis of PD-L1 expression in triple-negative (MDA-MB-231, HCC-1806), ER-positive (MCF-7, ZR-75-1, T47D), and HER-2-positive (SKBR-3) breast cancer cell lines (upper panel). beta-actin was used as a loading control. The experiment was repeated three times. Bands were quantified by densitometry; the signal for PD-L1 band was normalized to the corresponding actin band (lower panel). (B) Representative density plots and histogram overlays illustrating PD-L1 expression (red) against a background from isotype control (gray) for MCF-7 (upper panel) and MDA-MB-231 (lower panel) breast cancer cell lines using flow cytometry. The staining was performed using an anti-PD-L1 antibody (cat. no. 12-5983-42, eBioscience, clone MIH1, dilution 1:100). Numbers on the density plots indicate the percentage of PD-L1-positive cells. The experiment was repeated at least three times. (C) PD-L1 expression in macrophage subpopulations (M0, M0+IFNgamma, M2) detected by immunocytohistochemistry assay using Cytation 1 Cell Imaging Multi-Mode Reader (BioTek, Agilent). PD-L1 positively stained cells were detected using an anti-PD-L1 antibody (clone MIH1, cat. no. 14-5983-82, eBioscience, dilution 1:100) and are marked in green; red shows CD206-positive cells (cat. no. AF2534, R&D Systems, dilution 1:100). The signal was developed using AF488-conjugated or AF647-conjugated secondary antibody, resp
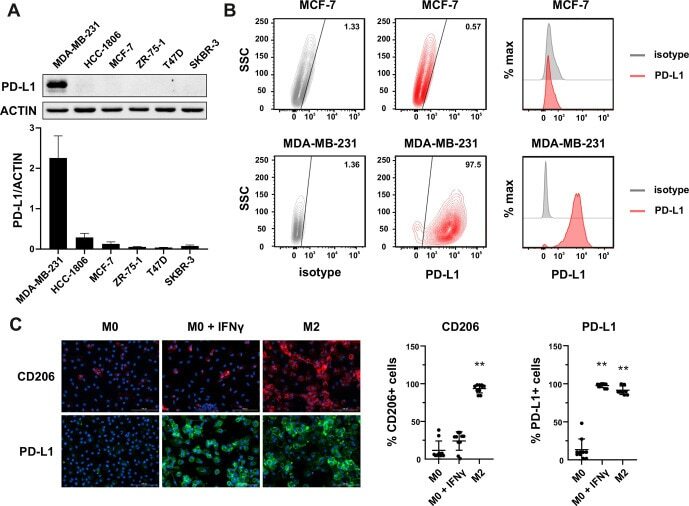
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
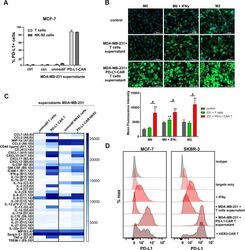
- Experimental details
- Figure 5 Induction of PD-L1 expression on the target cells. (A) PD-L1 expression induced by supernatant from activated CAR T or CAR-NK-92 cells on cancer cells was assessed by flow cytometry. The control (only medium) and conditioned supernatants from the 24 hours coincubation cultures of control (unmodified) or PD-L1-CAR T/NK-92 cells with the target MDA-MB-231 cells were transferred onto the culture of MCF-7 and incubated for 48 hours. Next, PD-L1 surface presence was assessed using anti-PD-L1 antibody (clone MIH1) by flow cytometry. The experiment was repeated three times. (B) Representative images of different subpopulations of macrophages (M0, M0+IFNgamma, M2) stained for PD-L1 assessed by immunocytochemistry assay using Cytation 1 Cell Imaging Multi-Mode Reader (BioTek, Agilent). Macrophages were incubated with 10 ng/mL IFNgamma or IL-4 and IL-10 (for M2) for 5 days before staining (every 2-day fresh portion of cytokines was added). The control (only medium) and conditioned supernatants from the 24 hours coincubation cultures of control (unmodified) or PD-L1-CAR T cells with the target MDA-MB-231 cells were transferred onto different subpopulations of macrophages and incubated for 48 hours. Next, PD-L1 surface presence was assessed using anti-PD-L1 antibody (clone MIH1, cat. no. 14-5983-82, eBioscience, diluted 1:100). The signal was developed using AF488-conjugated secondary antibody (green), and nuclei were counterstained with DAPI (blue), scale bar: 100 um. The backg
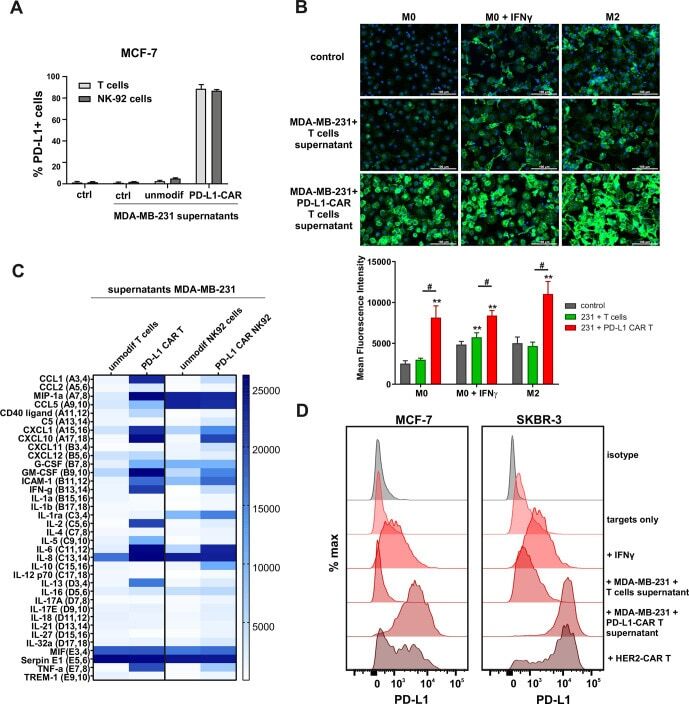
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- Figure 6 Expression of PD-L1 and PD-L1-CAR T mediated cytotoxicity in non-malignant cells. (A) IFNgamma induced expression of PD-L1 on HEK293T cell line derived from human embryonic kidney cells and non-malignant cells (HMEC, MCF-10A, and bone marrow-derived mesenchymal stem cells (BM-MSC)) assessed by flow cytometry as presented on the left panel. PD-L1 staining was performed using anti-PD-L1 antibody (clone MIH1). The representative western blot analysis of PD-L1 expression in human embryonic kidney HEK293T cells and non-malignant mammary epithelial HMEC and MCF10A cells, and BM-MSC (right panel). beta-actin was used as a loading control. The experiment was repeated three times. (B) PD-L1 expression induced on MCF-10A cells by activated CAR T cells (left panel) and RTCA-monitored cytotoxic activity of PD-L1 CAR T cells toward MCF-10A cells (right panel). The control (only medium) and conditioned supernatants from the 24 hours coincubation cultures in the presence of control (unmodified) T cells or PD-L1-CAR T cells with the target MDA-MB-231 cells were transferred onto the cultures of MCF-10A cell line and incubated for 48 hours. Next, PD-L1 surface presence was assessed by flow cytometry using anti-PD-L1 antibody (clone MIH1). Cytotoxic activity of PD-L1-CAR T cells against MCF-10A non-malignant cell line was measured by impedance analysis at the E:T ratios of 1:1 and 2:1. Samples were internally normalized for the cell index value measured before PD-L1-CAR T cells additio
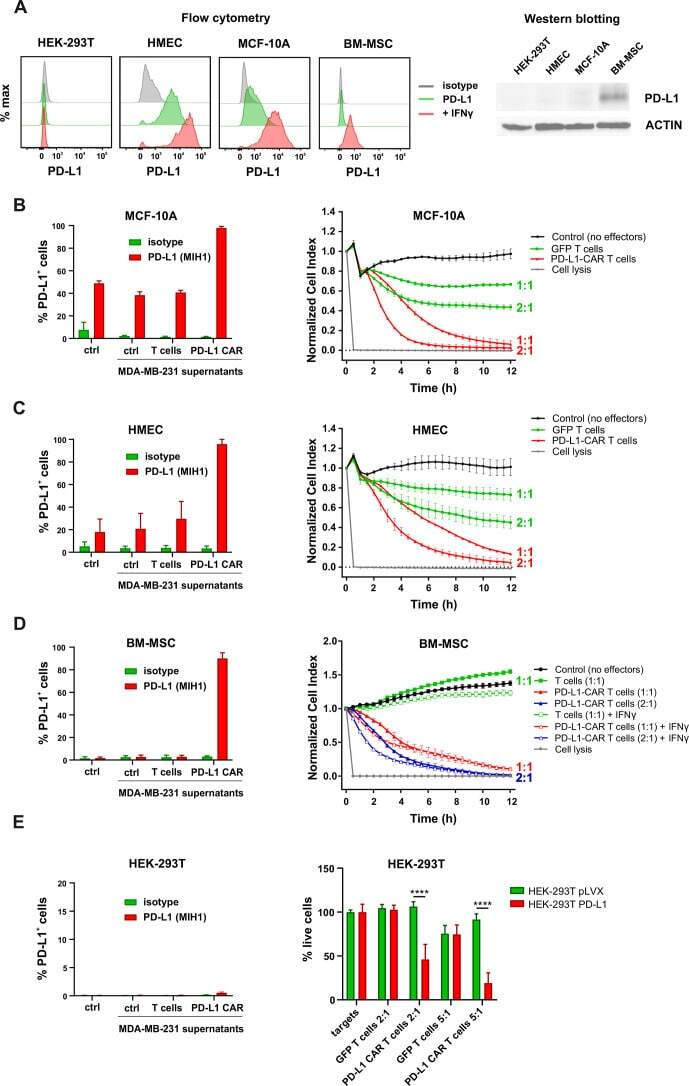
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- 10.1371/journal.pone.0124059.g007 Fig 7 Panels A-E show representative histograms for proliferative pattern of T lymphocytes (i.e., CD71+). Panel A shows T lymphocytes alone while panel B shows T lymphocytes co-cultured with antigen presenting cells (APCs). On the other hand, panel C shows the pattern for co-culturing of T lymphocytes with normoxic cardiac cells (CCs) while panels D and E show patterns for T lymphocyte co-cultured with ischemic-reperfused CCs in the absence and presence of the PD-L1 blocking antibody, respectively. Panel F shows the percent of proliferating T lymphocytes under each condition; data are average of the triplicate samples for each heart (n = 3 hearts/group/condition). IRI: Ischemia Reperfusion Injury. * p
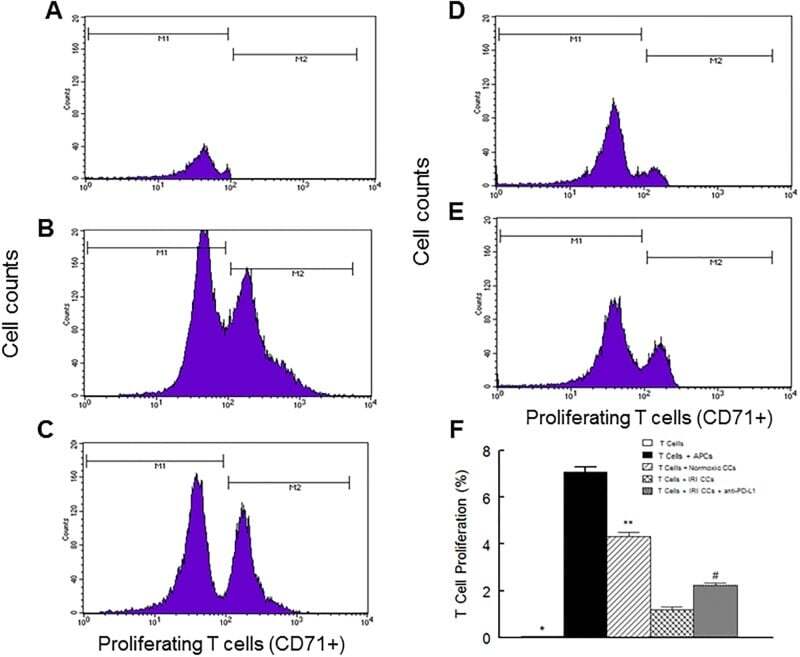
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- Figure 5 IL-25 Induces PD-L1 Surface Expression on hMSCs and Human Monocytes (A) PD-L1 in siCtrl MSCs (left) and siPD-L1 MSCs (right) was analyzed by surface staining. (B) Freshly isolated human PBLs were co-cultured without (left) or with siCtrl MSCs (middle) or siPD-L1 MSCs (right) for 3 days, followed by PMA/ionomycin stimulation for 6 hr. IL-17A production in CD3 + T cells was assessed by intracellular staining. Representative data are shown with numbers in the top right quadrants representing the percentages of IL-17A-producing CD3 + T cells. (C) Pooled data from PBLs (n = 4) and two hMSC donors (donors A and B) are shown. (D) Folds of reversed phenotypes of siIL-25 and siPD-L1 are shown. (E) PD-L1 expression on siCtrl hMSCs (left) and siIL-25 hMSCs (right) was assessed by cell surface staining. Filled histograms represent isotype control; unfilled histograms represent PD-L1 antibody staining. (F) Pooled data of PD-L1 expression (indicated by fold change in MFI) on siIL-25 hMSCs and siPD-L1 hMSCs (all three donors) are shown. PD-L1 expression levels were compared between hMSCs silenced for the target gene (IL-25 or PD-L1) and the respective siCtrl. (G) hMSCs were treated with the indicated doses of rhIL-25 for 18 hr and assessed for cell surface PD-L1 expression by cell surface staining. Pooled data (all three donors) are shown in chart to the right with bars representing MFI. (H) Human PBLs were treated with the indicated doses of rhIL-25 for 18 hr and assessed for cell
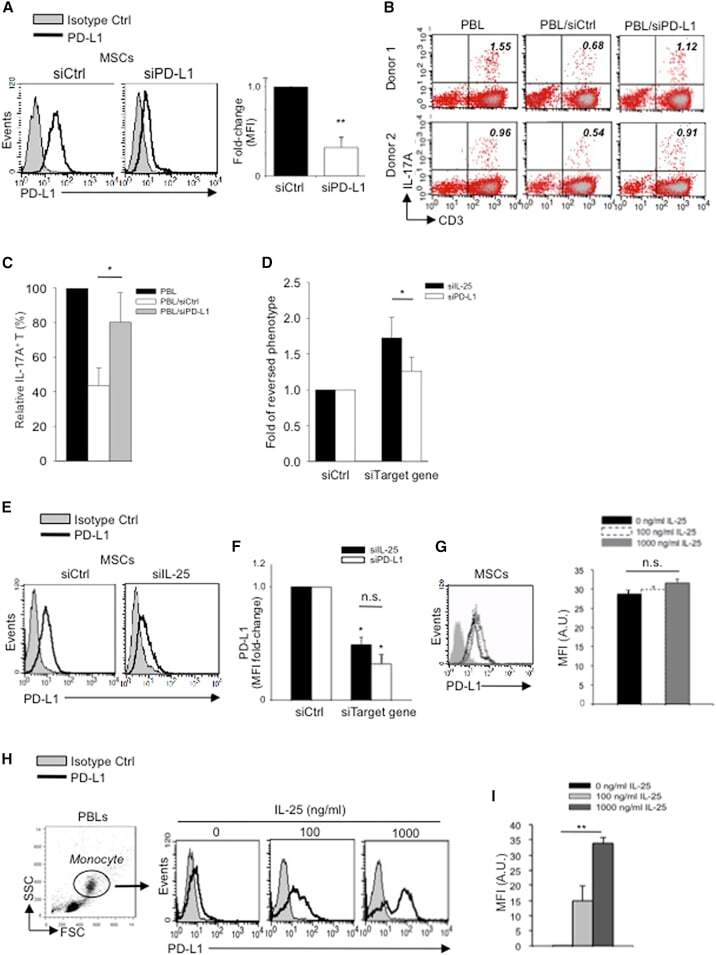
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- Figure 7 Blockade of APC-Intrinsic PD-1 Enhances the Synaptic Enrichment of T Cell-Intrinsic PD-1 and Inhibits T Cell-Mediated Cytotoxicity (A) Left: cartoon showing a Jurkat cell expressing PD-1-mGFP (shown as PD-1 in green) conjugated with a Raji cell co-transduced with PD-L1-mCherry and unlabeled PD-1. Right: representative confocal images of the conjugate at the indicated channel acquired 2 min after cell-cell contact. Scale bars, 10 um. (B-D) Same as (A), except that Raji (PD-L1+/PD-1+) cells (B), Jurkat (PD-1-mGFP) cells (C), or both (D) were preincubated with pembrolizumab and washed extensively prior to conjugation. Scale bars, 10 um. (E) Bar graph comparing the interface enrichment indices (calculated as described in STAR Methods) of the four conditions shown in (A)-(D). Data are expressed as mean +- SEM; n = 40 cells from three independent experiments. (F) Cartoon illustrating the cytotoxicity assay, in which EL4 cells were used as the target for OT-I cytotoxic T cells. (G) Bar graph summarizing the OT-I cytotoxicity under indicated conditions, with PD-1 blockade antibody J43 or isotype antibody (Iso) preincubated with neither cell, only EL4, only OT-I, or both cell types. See STAR Methods for details. Cytotoxicity was normalized to the no blockade antibody condition (i.e., EL4 and OT-I preincubated with the isotype control). n.s., not significantly different. Data are presented as mean +- SEM from three independent replicates. (H) Left: FACS histograms showing PD-1
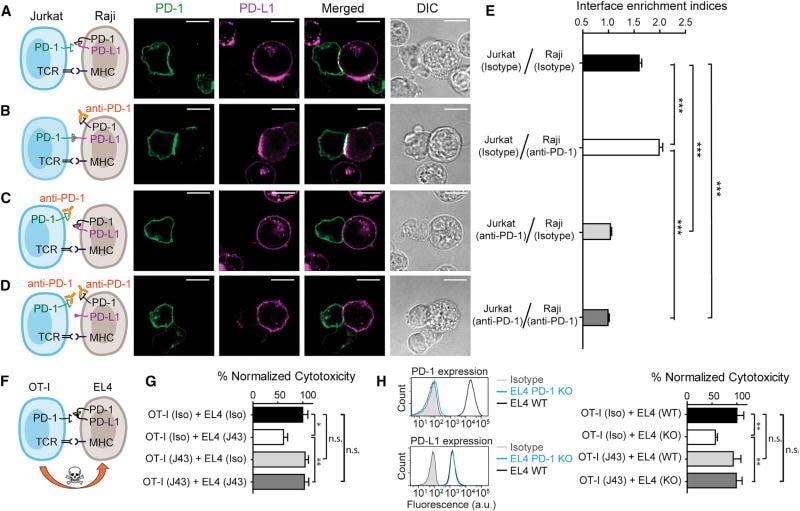
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
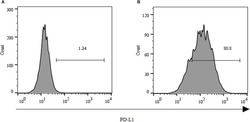
- Experimental details
- FIGURE 2 Expression of PD-L1 on human HCC PLC/PRF/5 cells. (A) Human HCC PLC/PRF/5 cells were cultured alone in the absence of GPC3-CAR T cells in RPMI 1640 medium containing 10% FBS. (B) Human HCC PLC/PRF/5 cells were cocultured with GPC3-CAR T cells at an effector:Target ratio of 1:1 for 18 h in RPMI 1640 medium containing 10% FBS. PD-L1 was determined by flow cytometry in the CD3-negative gate, and the fixable, viable stain 780 was used for discriminating live from dead cells.
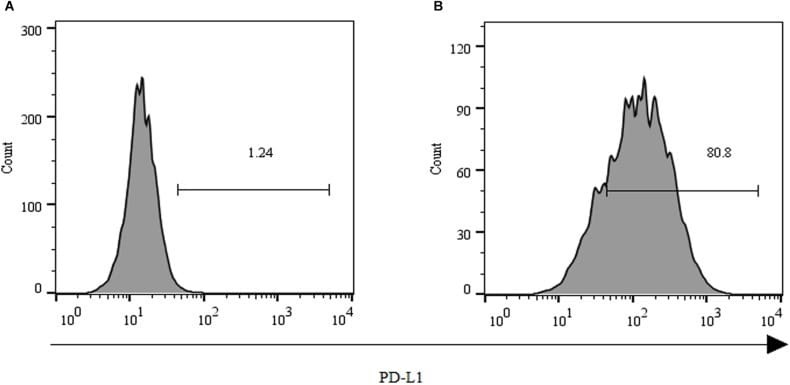
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- Figure 5 The activities of the tested molecules in the cell-based in vitro immune checkpoint blockade assay. ( A ) Western blot analysis of the expression of the human PD-L1 protein in the hPD-L1 aAPCs (artificial antigen-presenting cells, CHO-K1 TCRact/PD-L1) in comparison to the original CHO-K1 cells (CHO-K1 wt) and CHO-K1 overexpressing only the TCR Activator molecule (CHO-K1 TCRact). ( B ) Flow cytometry analysis of the expression of human PD-L1 on the surface of the hPD-L1 aAPCs. ( C ) Comparison of bioactivities of the known PD-L1-targeting molecules: nivolumab, peptide-57 , BMS-1166 , and the tested molecules, CA-170 and AUNP-12 in the hPD-1/hPD-L1 immune checkpoint assay.
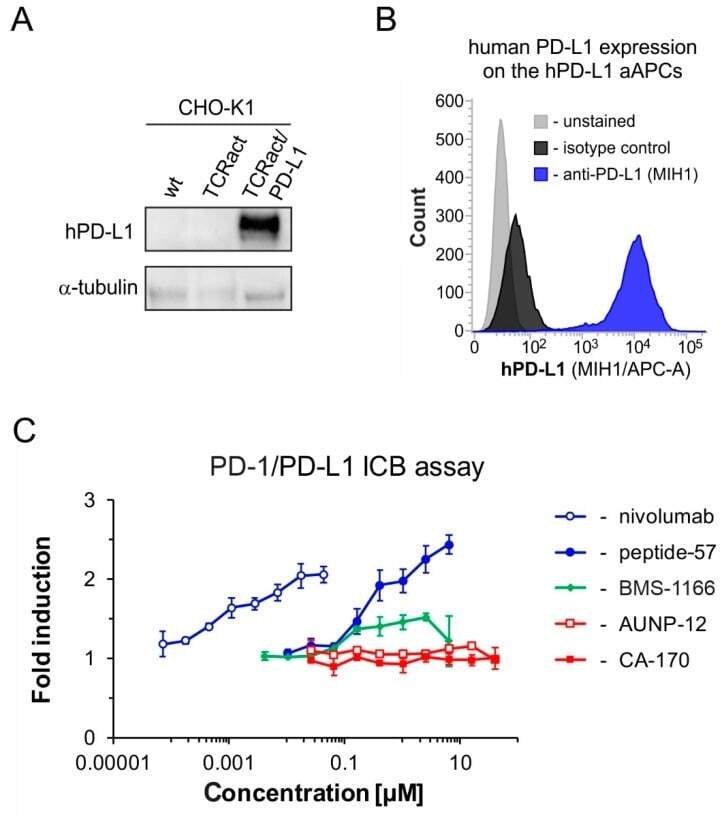
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
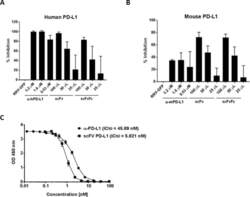
- Experimental details
- Figure 3 Detection of scFv PD-L1 binding to PD-L1 by competitive ELISA Wells in a 96-well microtiter plate were coated with ( A ) recombinant human or ( B ) mouse PD-L1-Fc followed by co-incubation with His-tagged recombinant PD-1-Fc in competition with 25, 50 or 100 uL of supernatant of undefined scFv PD-L1 (scFv) and scFvFc PD-L1 (scFvFc) protein concentration collected from CT26 cells maximally infected with RRV-scFv-PDL1 and RRV-scFvFc-PDL1, respectively. Anti-human or mouse PD-L1 antibody was included as positive control (indicated as alpha-hPD-L1 and alpha-mPD-L1). Anti- His tag antibody was used to detect bound His-tagged PD-1-Fc. Optical density was measured at 450 nm. The percentage of inhibition was calculated with respect to the supernatant from CT26 maximally infected with RRV-GFP (non-scFv-PD-L1) used in the competition. Error bars indicate the standard deviation of the dataset. ( C ) Wells were coated with recombinant human PDL1-Fc followed by co-incubation with His-tagged recombinant PD-1-Fc in competition with purified scFv PD-L1 or anti-human PD-L1 antibody range from 0.01-100 nM. IC50 values were determined using best-fit values from non-linear three-parameters logistic.
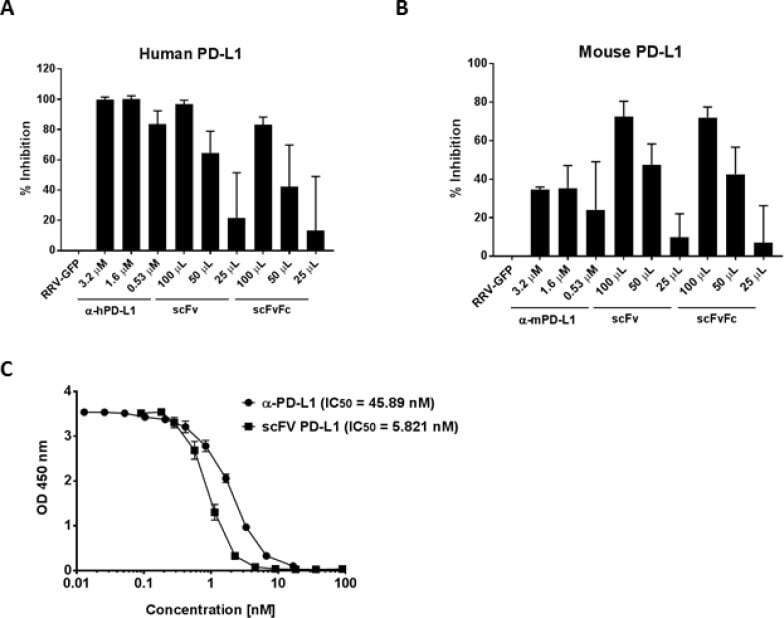
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
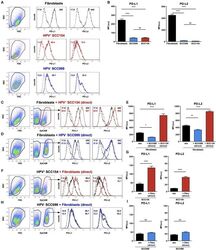
- Experimental details
- Figure 3 HPV-positive HNSCCs increase PD-L1 and PD-L2 on fibroblasts. PD-L1 and PD-L2 expression on primary BJ human fibroblasts, HPV-positive (SCC154), and HPV-negative (SCC099) HNSCC cell lines (HNSCCs) was detected by flow cytometry. Illustrative histograms show PD-L1 and PD-L2 expression on fibroblasts (black histograms), HPV-positive (red histograms), or HPV-negative (blue histograms) HNSCCs (A) . Graphs show PD-L1 and PD-L2 expression (mean +- SEM; n = 5) in fibroblasts, HPV-positive, and HPV-negative HNSCCs (B) . Fibroblasts were cultured alone or co-cultured in direct contact (direct) with HPV-positive (SCC154) or HPV-negative (SCC099) HNSCCs. Fibroblasts were identified in co-cultures by lack of EpCAM expression. Illustrative histograms show PD-L1 and PD-L2 expression on fibroblasts cultured alone (black histograms) or co-cultured directly with HPV-positive SCC154 ( C ; red histograms) or HPV-negative SCC099 ( D ; blue histograms). Graphs show PD-L1 and PD-L2 expression (mean +- SEM; n = 5) on fibroblasts cultured alone (w/o) or co-cultured directly with HNSCC cells (E) . HPV-positive (SCC154) or HPV-negative (SCC099) HNSCCs were cultured alone or co-cultured in direct contact (direct) with fibroblasts. HNSCCs were identified in co-cultures by EpCAM expression. Illustrative histograms show PD-L1 and PD-L2 expression on HPV-positive SCC154 cultured alone (black histograms) or co-cultured directly with fibroblasts ( F ; red histograms). Graphs show PD-L1 and PD-
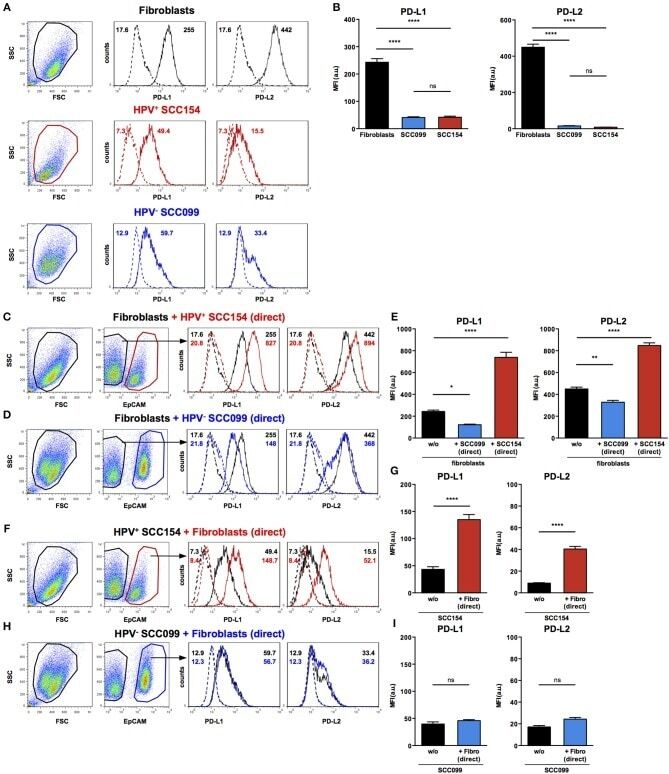
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
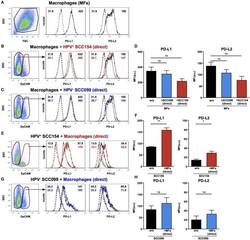
- Experimental details
- Figure 4 PD-L1 and PD-L2 expression in co-cultures of macrophages and HPV-positive or HPV-negative HNSCCs. Macrophages were cultured alone (w/o) or co-cultured in direct contact (direct) with HPV-positive (SCC154) or HPV-negative (SCC099) HNSCC cell lines (HNSCCs). Illustrative histograms show PD-L1 and PD-L2 expression on macrophages cultured alone ( A ; black histograms). Macrophages were identified in co-cultures by lack of EpCAM expression. Illustrative histograms show PD-L1 and PD-L2 expression on macrophages cultured alone (black histograms) or co-cultured directly with HPV-positive SCC154 ( B ; red histograms) or HPV-negative SCC099 ( C ; blue histograms). Graphs show PD-L1 and PD-L2 expression (mean +- SEM; n = 4) on macrophages cultured alone (w/o) or co-cultured directly with HNSCCs (D) . HPV-positive (SCC154) or HPV-negative (SCC099) HNSCCs were cultured alone or co-cultured in direct contact (direct) with macrophages. HNSCCs were identified in co-cultures by EpCAM expression. Illustrative histograms show PD-L1 and PD-L2 expression on HPV-positive SCC154 cultured alone (black histograms) or co-cultured directly with macrophages ( E ; red histograms). Graphs show PD-L1 and PD-L2 expression (mean +- SEM; n = 3) on HPV-positive SCC154 cultured alone or co-cultured directly with macrophages (F) . Illustrative histograms show PD-L1 and PD-L2 expression on HPV-negative SCC099 cultured alone (black histograms) or co-cultured directly with macrophages ( G ; blue hist
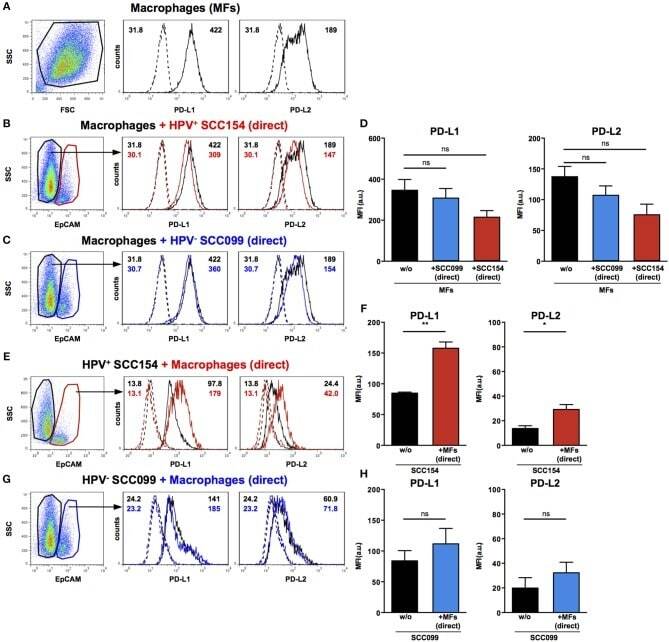
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
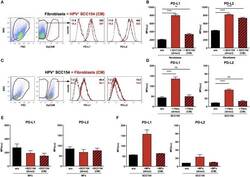
- Experimental details
- Figure 5 Conditioned medium from HPV-positive HNSCCs up-regulates PD-L1 and PD-L2 on fibroblasts. Fibroblasts were cultured alone or co-cultured in direct contact with HPV-positive SCC154 (direct) or with conditioned medium from HPV-positive SCC154 (CM). Illustrative histograms show PD-L1 and PD-L2 expression on fibroblasts cultured alone (black histograms) or co-cultured with conditioned medium from HPV-positive SCC154 ( A ; red histograms). Graphs show PD-L1 and PD-L2 expression (mean +- SEM; n = 13) on fibroblasts cultured alone (w/o), co-cultured directly with HPV-positive SCC154 (direct) or with conditioned medium from HPV-positive SCC154 (CM) (B) . HPV-positive (SCC154) HNSCCs were cultured alone or co-cultured in direct contact with fibroblasts (Fibro direct) or with conditioned medium from fibroblasts (Fibro CM). Illustrative histograms show PD-L1 and PD-L2 expression on HPV-positive SCC154 cultured alone (black histograms) or co-cultured with conditioned medium from fibroblasts ( C ; red histograms). Graphs show PD-L1 and PD-L2 expression (mean +- SEM; n = 4) on HPV-positive SCC154 cultured alone (w/o), co-cultured directly with fibroblasts (Fibro direct) or with conditioned medium from fibroblasts (Fibro CM) (D) . Macrophages were cultured alone (w/o) or co-cultured in direct contact with HPV-positive SCC154 (direct) or with conditioned medium from HPV-positive SCC154 (CM). Graphs show PD-L1 and PD-L2 expression (mean +- SEM; n = 3) on macrophages for the in
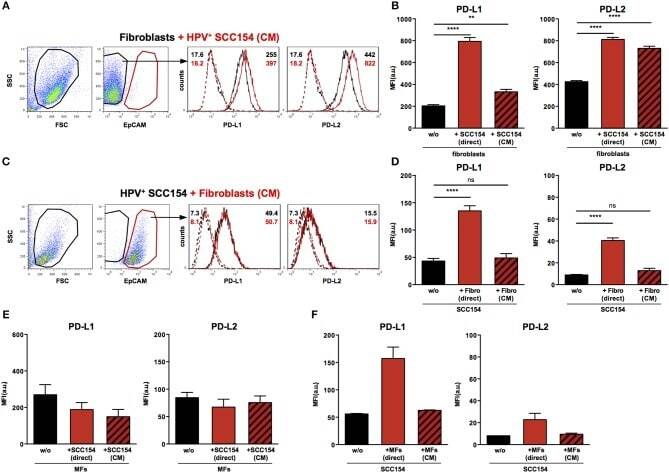
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- Figure 6 Blockade of IFN-gamma, TNF-alpha, or CD81 does not affect PD-L1 and PD-L2 up-regulation by HPV-positive HNSCCs. Fibroblasts were cultured alone (w/o) or co-cultured in direct contact with HPV-positive SCC154 (SCC154 direct) or with conditioned medium from HPV-positive SCC154 (SCC154 CM) as indicated. Graphs (A) show IFN-gamma and TNF-alpha levels in culture supernatants (mean +- SEM; n = 4). The dashed red line indicates the lowest value (15.6 pg/ml) of the dynamic range for the ELISA assays used. Neutralizing antibodies anti-IFN-gamma (B,C) , anti-TNF-alpha (D,E) , or anti-CD81 (F,G) were added to the cultures as indicated. Illustrative histograms show PD-L1 and PD-L2 expression on fibroblasts cultured alone (black histograms), co-cultured directly with HPV-positive SCC154 or with conditioned medium from HPV-positive SCC154 alone (red histograms) or in the presence of blocking antibodies (green histograms). Graphs show PD-L1 and PD-L2 expression (mean +- SEM; n = 3) on fibroblasts for the indicated treatments. ns, not significant (one-way ANOVA with Bonferroni correction for multiple comparisons) Numbers adjacent to plots represent MFI values; dashed histograms show control staining with isotype-matched antibodies. MFI, mean fluorescence intensity.
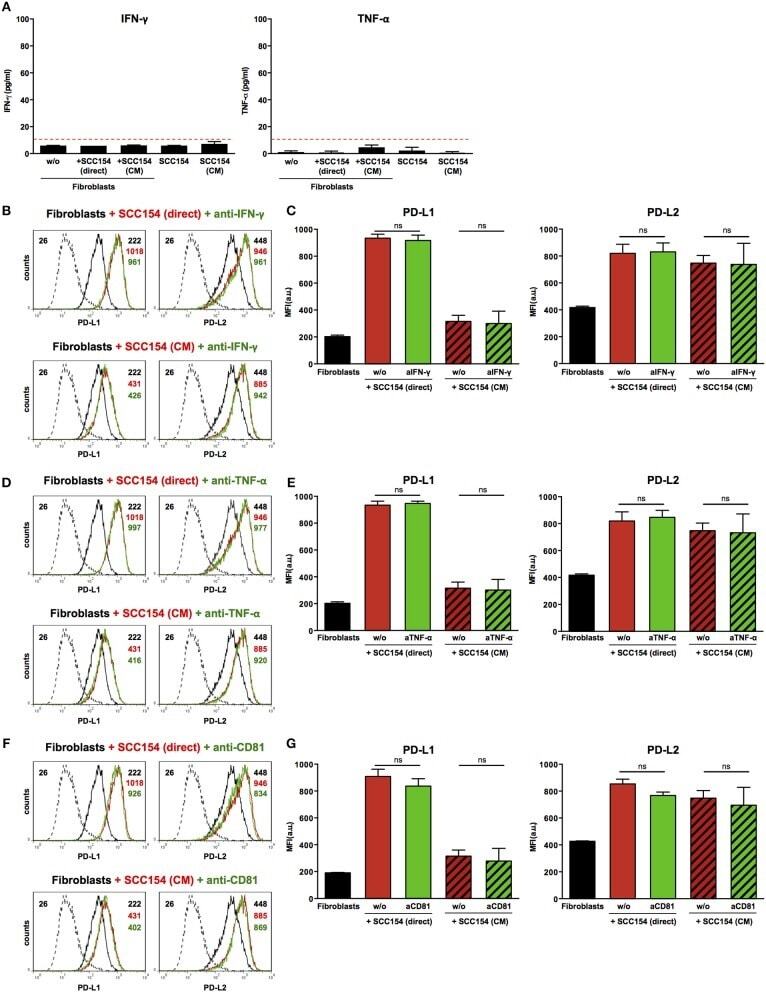
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- Figure 7 The TLR9 antagonists ODN TTAGGG and chloroquine inhibit PD-1 ligands up-regulation on fibroblasts co-cultured with HPV-positive HNSCCs. Fibroblasts were cultured alone (w/o) or co-cultured in direct contact with HPV-positive SCC154 (SCC154 direct) or with conditioned medium from HPV-positive SCC154 (SCC154 CM) in the presence or absence of the TLR9 antagonists ODN TTAGGG (ODN) or chloroquine (CHQ). Illustrative histograms show PD-L1 and PD-L2 expression on fibroblasts cultured alone (black histograms), co-cultured directly with HPV-positive SCC154 (red histograms) or co-cultured directly with HPV-positive SCC154 in the presence of ODN (A) or CHQ (H) (green histograms). Illustrative histograms show PD-L1 and PD-L2 expression on fibroblasts cultured alone (black histograms), cultured with conditioned medium from HPV-positive SCC154 (red histograms) or with conditioned medium from HPV-positive SCC154 in the presence of ODN (B) or CHQ (I) (green histograms). Graphs show PD-L1 and PD-L2 expression (mean +- SEM; n = 6) on fibroblasts for the indicated treatments (C,J) . Illustrative histograms show PD-L1 and PD-L2 expression on fibroblasts cultured alone (black histograms) or in the presence of ODN (D) or CHQ (K) (green histograms). Graphs show PD-L1 and PD-L2 expression (mean +- SEM; n = 4) on fibroblasts for the indicated treatments (E,L) . HPV-positive SCC154 were cultured alone or co-cultured in direct contact (direct) with fibroblasts in the presence
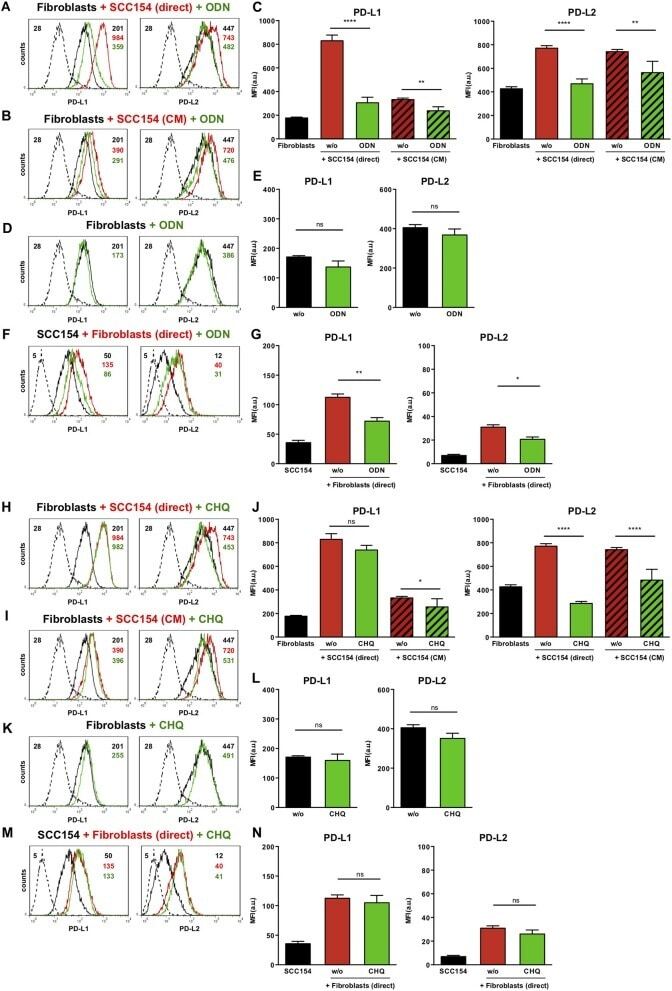
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- Figure 5. Exosomal PD-L1 Suppresses Tumor Progression in Syngeneic Colorectal Cancer Model (A) Western for PD-L1 in WT and Rab27a MC38 100k g extracellular fraction. (B) Flow cytometry for surface PD-L1 on MC38 cells. (C) Cell counts over time for Rab27a versus WT MC38 cells. n = 3. Error bars represent SD. (D) Cell counts over time for Pd-l1 versus WT MC38 cells. n = 3. Error bars represent SD. (E) Tumor growth over time following subcutaneous injection of 1 x 10 6 WT, Rab27a , or Pd-l1 MC38 cells into immunocompetent B6 mice. n = 5 for each genotype. Error bars represent SEM. MC38 WT versus MC38 Rab27a , p < 0.05. MC38 WT versus MC38 Pd-l1 , p < 0.05. MC38 WT versus MC38 Pd-l1 ; Rab27a , p < 0.05 (two-way ANOVA test). (F) Mouse survival curve following injection of cells like in (D). n = 10 for each genotype. MC38 WT versus MC38 Rab27a , p < 0.001. MC38 WT versus MC38 Pd-l1 , p < 0.001. MC38 WT versus MC38 Pd-l1 ; Rab27a , p < 0.001 (log rank test). (G) Survival curve for mice injected with WT, Rab27a , or Pd-l1 MC38 cells followed by treatment with either anti-PD-L1 or isotype control antibody. n = 5 for each condition. MC38 WT isotype versus MC38 WT anti-PD-L1, p < 0.01. MC38 Rab27a isotype versus MC38 Rab27a anti-PD-L1, p < 0.05. MC38 Pd-l1 isotype versus MC38 Rab27a anti-PD-L1, ns. (log rank test). See also Figure S7 .
