Explore
Explore Validate
Validate Learn
Learn37-3400
antibody from Invitrogen Antibodies
Targeting: BTRC
beta-TrCP1, betaTrCP, bTrCP, bTrCP1, FBXW1A, Fwd1
 Western blot
Western blot ELISA
ELISA Immunocytochemistry
ImmunocytochemistryAntibody data
- Antibody Data
- Antigen structure
- References [21]
- Comments [0]
- Validations
- Immunocytochemistry [2]
- Flow cytometry [1]
- Other assay [25]
Submit
Validation data
Reference
Comment
Report error
- Product number
- 37-3400 - Provider product page

- Provider
- Invitrogen Antibodies
- Product name
- BTRC Monoclonal Antibody (1B1D2)
- Antibody type
- Monoclonal
- Antigen
- Recombinant full-length protein
- Reactivity
- Human, Rat
- Host
- Mouse
- Isotype
- IgG
- Antibody clone number
- 1B1D2
- Vial size
- 100 μg
- Concentration
- 0.5 mg/mL
- Storage
- -20°C
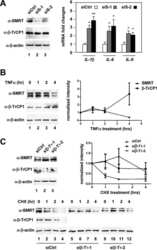
Submitted references Long noncoding RNA ASB16-AS1 inhibits adrenocortical carcinoma cell growth by promoting ubiquitination of RNA-binding protein HuR.
ZAKα Recognizes Stalled Ribosomes through Partially Redundant Sensor Domains.
Long noncoding RNA OCC-1 suppresses cell growth through destabilizing HuR protein in colorectal cancer.
TIAM1 Antagonizes TAZ/YAP Both in the Destruction Complex in the Cytoplasm and in the Nucleus to Inhibit Invasion of Intestinal Epithelial Cells.
Interaction of tankyrase and peroxiredoxin II is indispensable for the survival of colorectal cancer cells.
Hippo signaling interactions with Wnt/β-catenin and Notch signaling repress liver tumorigenesis.
Substrate trapping proteomics reveals targets of the βTrCP2/FBXW11 ubiquitin ligase.
Growth-arrest-specific 7C protein inhibits tumor metastasis via the N-WASP/FAK/F-actin and hnRNP U/β-TrCP/β-catenin pathways in lung cancer.
PLK1 and β-TrCP-dependent ubiquitination and degradation of Rap1GAP controls cell proliferation.
RASSF1A inactivation unleashes a tumor suppressor/oncogene cascade with context-dependent consequences on cell cycle progression.
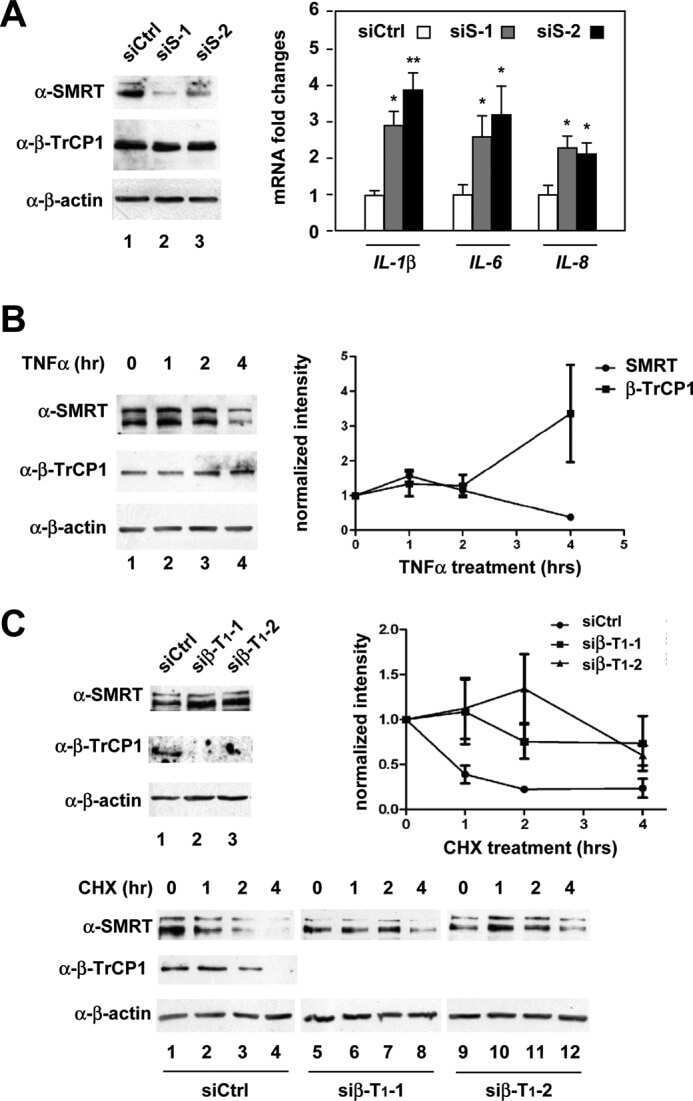
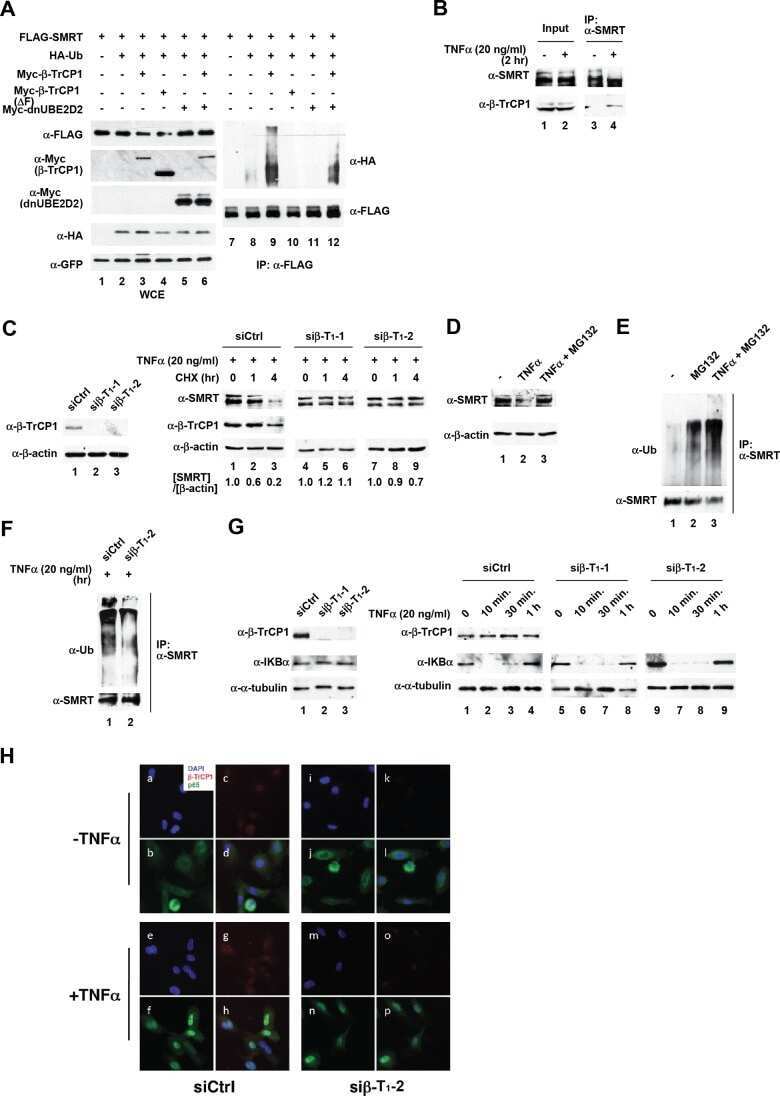
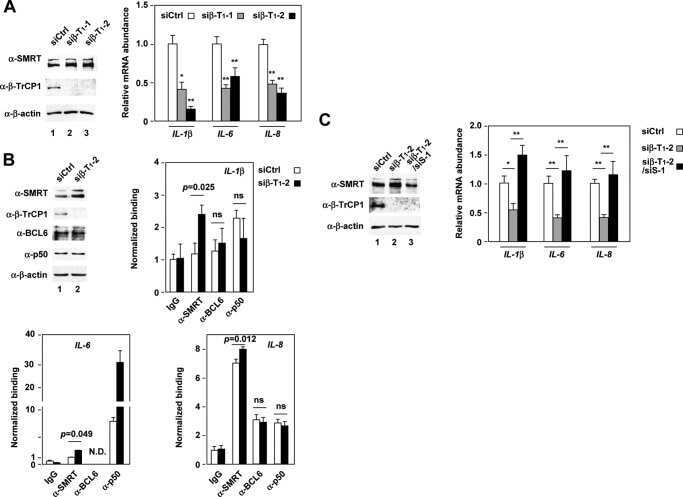
β-Transducin repeat-containing protein 1 (β-TrCP1)-mediated silencing mediator of retinoic acid and thyroid hormone receptor (SMRT) protein degradation promotes tumor necrosis factor α (TNFα)-induced inflammatory gene expression.
FAM123A binds to microtubules and inhibits the guanine nucleotide exchange factor ARHGEF2 to decrease actomyosin contractility.
mTOR drives its own activation via SCF(βTrCP)-dependent degradation of the mTOR inhibitor DEPTOR.
FOXO3a-Dependent Mechanism of E1A-Induced Chemosensitization.
SCF/{beta}-TrCP promotes glycogen synthase kinase 3-dependent degradation of the Nrf2 transcription factor in a Keap1-independent manner.
Preservations of nephrin and synaptopodin by recombinant hepatocyte growth factor in podocytes for the attenuations of foot process injury and albuminuria in nephritic mice.
βTrCP-dependent degradation of CDC25B phosphatase at the metaphase-anaphase transition is a pre-requisite for correct mitotic exit.
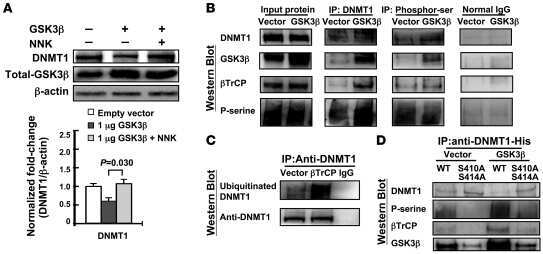
The tobacco-specific carcinogen NNK induces DNA methyltransferase 1 accumulation and tumor suppressor gene hypermethylation in mice and lung cancer patients.
SCFbeta-TRCP controls oncogenic transformation and neural differentiation through REST degradation.
TAZ promotes PC2 degradation through a SCFbeta-Trcp E3 ligase complex.
Destruction of Claspin by SCFbetaTrCP restrains Chk1 activation and facilitates recovery from genotoxic stress.
Long B, Yang X, Xu X, Li X, Xu X, Zhang X, Zhang S
Cell death & disease 2020 Nov 20;11(11):995
Cell death & disease 2020 Nov 20;11(11):995
ZAKα Recognizes Stalled Ribosomes through Partially Redundant Sensor Domains.
Vind AC, Snieckute G, Blasius M, Tiedje C, Krogh N, Bekker-Jensen DB, Andersen KL, Nordgaard C, Tollenaere MAX, Lund AH, Olsen JV, Nielsen H, Bekker-Jensen S
Molecular cell 2020 May 21;78(4):700-713.e7
Molecular cell 2020 May 21;78(4):700-713.e7
Long noncoding RNA OCC-1 suppresses cell growth through destabilizing HuR protein in colorectal cancer.
Lan Y, Xiao X, He Z, Luo Y, Wu C, Li L, Song X
Nucleic acids research 2018 Jun 20;46(11):5809-5821
Nucleic acids research 2018 Jun 20;46(11):5809-5821
TIAM1 Antagonizes TAZ/YAP Both in the Destruction Complex in the Cytoplasm and in the Nucleus to Inhibit Invasion of Intestinal Epithelial Cells.
Diamantopoulou Z, White G, Fadlullah MZH, Dreger M, Pickering K, Maltas J, Ashton G, MacLeod R, Baillie GS, Kouskoff V, Lacaud G, Murray GI, Sansom OJ, Hurlstone AFL, Malliri A
Cancer cell 2017 May 8;31(5):621-634.e6
Cancer cell 2017 May 8;31(5):621-634.e6
Interaction of tankyrase and peroxiredoxin II is indispensable for the survival of colorectal cancer cells.
Kang DH, Lee DJ, Lee S, Lee SY, Jun Y, Kim Y, Kim Y, Lee JS, Lee DK, Lee S, Jho EH, Yu DY, Kang SW
Nature communications 2017 Jun 28;8(1):40
Nature communications 2017 Jun 28;8(1):40
Hippo signaling interactions with Wnt/β-catenin and Notch signaling repress liver tumorigenesis.
Kim W, Khan SK, Gvozdenovic-Jeremic J, Kim Y, Dahlman J, Kim H, Park O, Ishitani T, Jho EH, Gao B, Yang Y
The Journal of clinical investigation 2017 Jan 3;127(1):137-152
The Journal of clinical investigation 2017 Jan 3;127(1):137-152
Substrate trapping proteomics reveals targets of the βTrCP2/FBXW11 ubiquitin ligase.
Kim TY, Siesser PF, Rossman KL, Goldfarb D, Mackinnon K, Yan F, Yi X, MacCoss MJ, Moon RT, Der CJ, Major MB
Molecular and cellular biology 2015 Jan;35(1):167-81
Molecular and cellular biology 2015 Jan;35(1):167-81
Growth-arrest-specific 7C protein inhibits tumor metastasis via the N-WASP/FAK/F-actin and hnRNP U/β-TrCP/β-catenin pathways in lung cancer.
Tseng RC, Chang JW, Mao JS, Tsai CD, Wu PC, Lin CJ, Lu YL, Liao SY, Cheng HC, Hsu HS, Wang YC
Oncotarget 2015 Dec 29;6(42):44207-21
Oncotarget 2015 Dec 29;6(42):44207-21
PLK1 and β-TrCP-dependent ubiquitination and degradation of Rap1GAP controls cell proliferation.
Wang D, Zhang P, Gao K, Tang Y, Jin X, Zhang Y, Yi Q, Wang C, Yu L
PloS one 2014;9(10):e110296
PloS one 2014;9(10):e110296
RASSF1A inactivation unleashes a tumor suppressor/oncogene cascade with context-dependent consequences on cell cycle progression.
Ram RR, Mendiratta S, Bodemann BO, Torres MJ, Eskiocak U, White MA
Molecular and cellular biology 2014 Jun;34(12):2350-8
Molecular and cellular biology 2014 Jun;34(12):2350-8
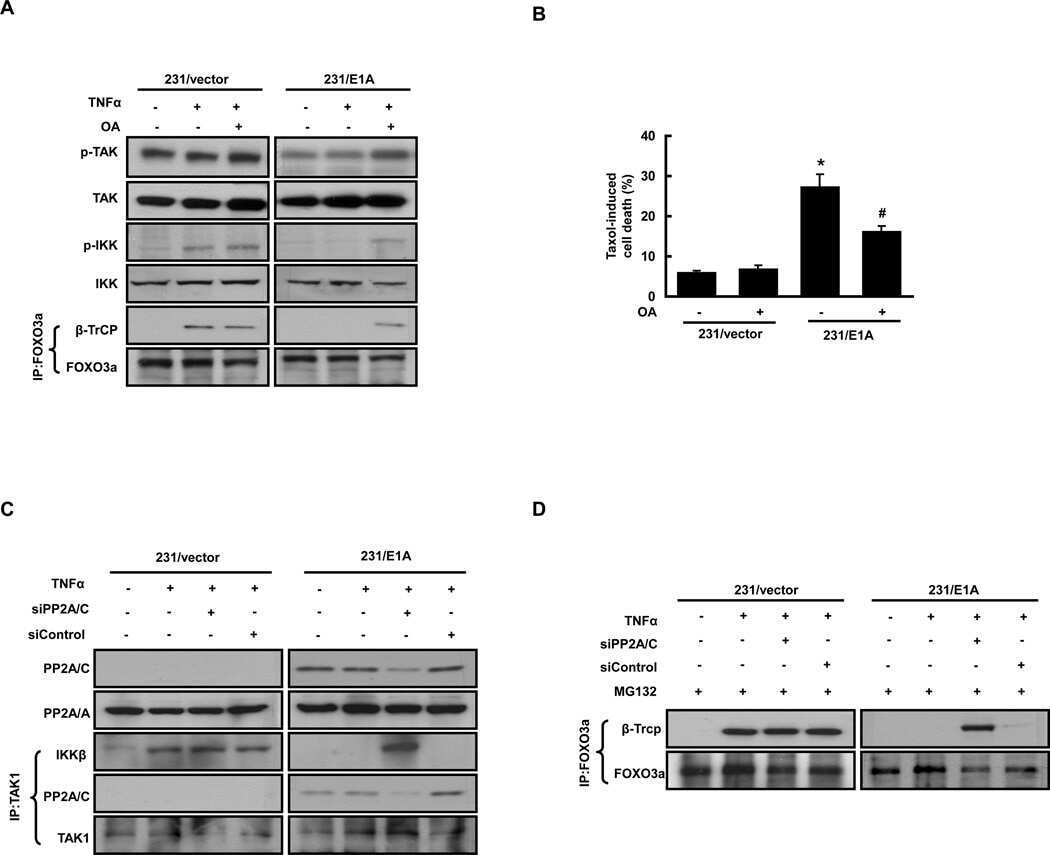
β-Transducin repeat-containing protein 1 (β-TrCP1)-mediated silencing mediator of retinoic acid and thyroid hormone receptor (SMRT) protein degradation promotes tumor necrosis factor α (TNFα)-induced inflammatory gene expression.
Hsu KS, Kao HY
The Journal of biological chemistry 2013 Aug 30;288(35):25375-25386
The Journal of biological chemistry 2013 Aug 30;288(35):25375-25386
FAM123A binds to microtubules and inhibits the guanine nucleotide exchange factor ARHGEF2 to decrease actomyosin contractility.
Siesser PF, Motolese M, Walker MP, Goldfarb D, Gewain K, Yan F, Kulikauskas RM, Chien AJ, Wordeman L, Major MB
Science signaling 2012 Sep 4;5(240):ra64
Science signaling 2012 Sep 4;5(240):ra64
mTOR drives its own activation via SCF(βTrCP)-dependent degradation of the mTOR inhibitor DEPTOR.
Gao D, Inuzuka H, Tan MK, Fukushima H, Locasale JW, Liu P, Wan L, Zhai B, Chin YR, Shaik S, Lyssiotis CA, Gygi SP, Toker A, Cantley LC, Asara JM, Harper JW, Wei W
Molecular cell 2011 Oct 21;44(2):290-303
Molecular cell 2011 Oct 21;44(2):290-303
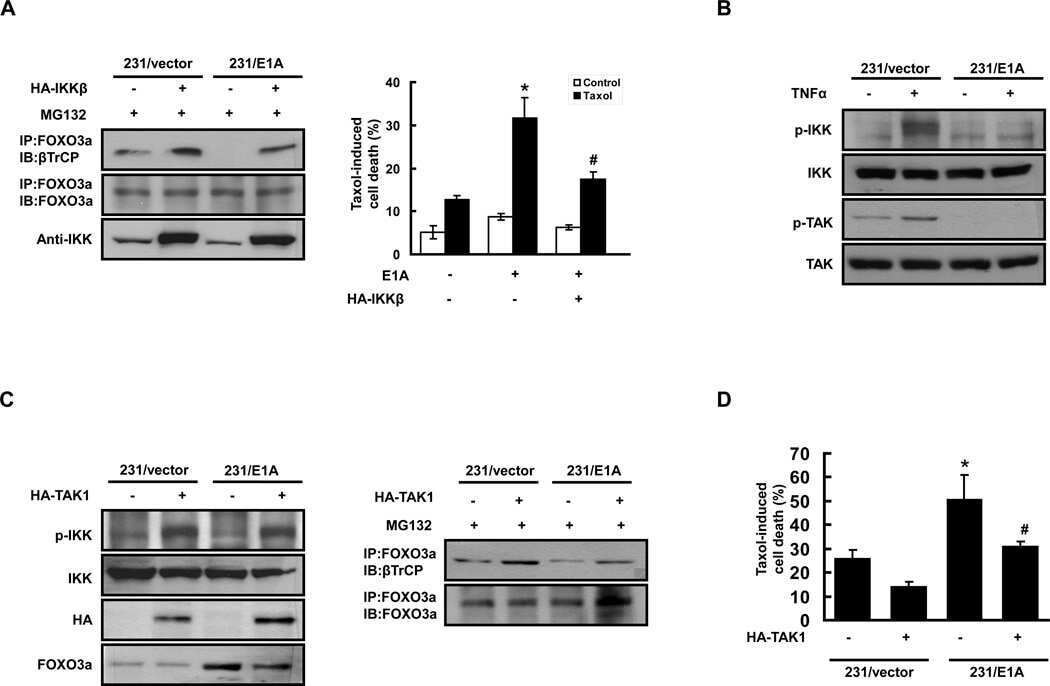
FOXO3a-Dependent Mechanism of E1A-Induced Chemosensitization.
Su JL, Cheng X, Yamaguchi H, Chang YW, Hou CF, Lee DF, Ko HW, Hua KT, Wang YN, Hsiao M, Chen PB, Hsu JM, Bast RC Jr, Hortobagyi GN, Hung MC
Cancer research 2011 Nov 1;71(21):6878-87
Cancer research 2011 Nov 1;71(21):6878-87
SCF/{beta}-TrCP promotes glycogen synthase kinase 3-dependent degradation of the Nrf2 transcription factor in a Keap1-independent manner.
Rada P, Rojo AI, Chowdhry S, McMahon M, Hayes JD, Cuadrado A
Molecular and cellular biology 2011 Mar;31(6):1121-33
Molecular and cellular biology 2011 Mar;31(6):1121-33
Preservations of nephrin and synaptopodin by recombinant hepatocyte growth factor in podocytes for the attenuations of foot process injury and albuminuria in nephritic mice.
Kato T, Mizuno S, Nakamura T
Nephrology (Carlton, Vic.) 2011 Mar;16(3):310-8
Nephrology (Carlton, Vic.) 2011 Mar;16(3):310-8
βTrCP-dependent degradation of CDC25B phosphatase at the metaphase-anaphase transition is a pre-requisite for correct mitotic exit.
Thomas Y, Coux O, Baldin V
Cell cycle (Georgetown, Tex.) 2010 Nov 1;9(21):4338-50
Cell cycle (Georgetown, Tex.) 2010 Nov 1;9(21):4338-50
The tobacco-specific carcinogen NNK induces DNA methyltransferase 1 accumulation and tumor suppressor gene hypermethylation in mice and lung cancer patients.
Lin RK, Hsieh YS, Lin P, Hsu HS, Chen CY, Tang YA, Lee CF, Wang YC
The Journal of clinical investigation 2010 Feb;120(2):521-32
The Journal of clinical investigation 2010 Feb;120(2):521-32
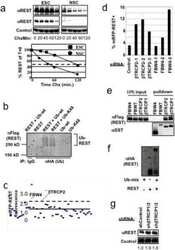
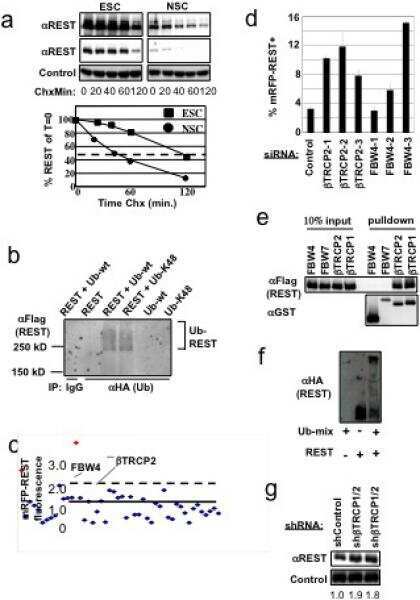
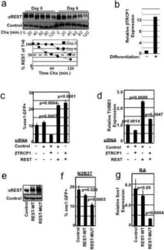
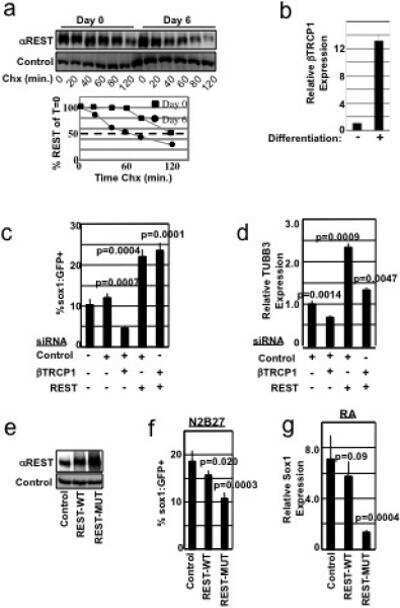
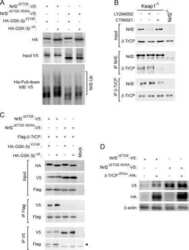
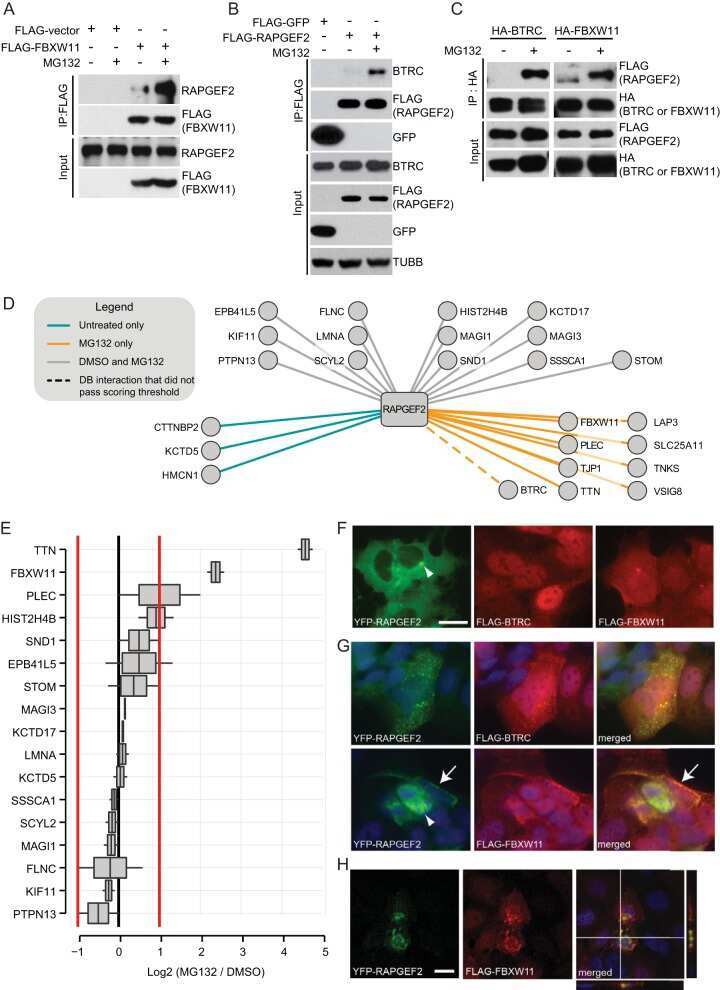
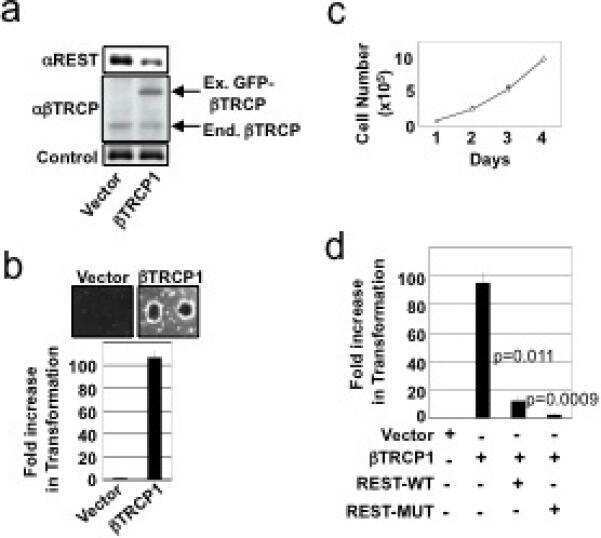
SCFbeta-TRCP controls oncogenic transformation and neural differentiation through REST degradation.
Westbrook TF, Hu G, Ang XL, Mulligan P, Pavlova NN, Liang A, Leng Y, Maehr R, Shi Y, Harper JW, Elledge SJ
Nature 2008 Mar 20;452(7185):370-4
Nature 2008 Mar 20;452(7185):370-4
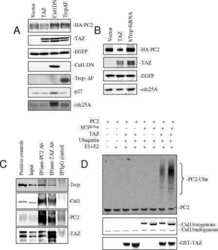
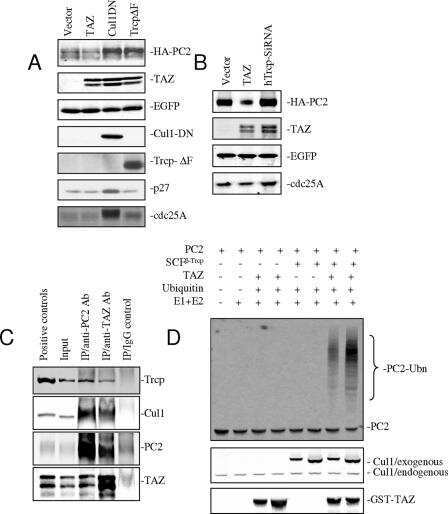
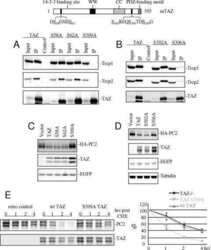
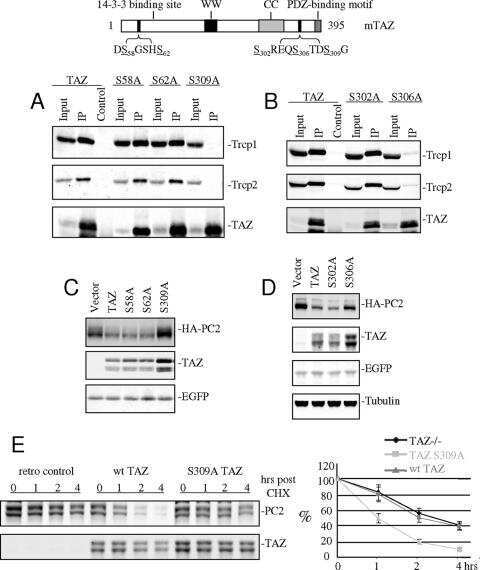
TAZ promotes PC2 degradation through a SCFbeta-Trcp E3 ligase complex.
Tian Y, Kolb R, Hong JH, Carroll J, Li D, You J, Bronson R, Yaffe MB, Zhou J, Benjamin T
Molecular and cellular biology 2007 Sep;27(18):6383-95
Molecular and cellular biology 2007 Sep;27(18):6383-95
Destruction of Claspin by SCFbetaTrCP restrains Chk1 activation and facilitates recovery from genotoxic stress.
Mailand N, Bekker-Jensen S, Bartek J, Lukas J
Molecular cell 2006 Aug 4;23(3):307-18
Molecular cell 2006 Aug 4;23(3):307-18
No comments: Submit comment
Supportive validation
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
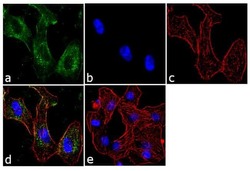
- Experimental details
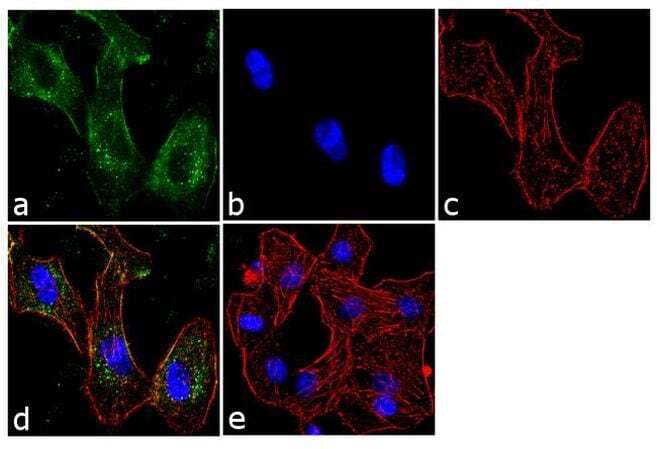
- Immunofluorescent analysis of BTRC was performed using 70% confluent log phase A549 cells. The cells were fixed with 4% paraformaldehyde for 10 minutes, permeabilized with 0.1% Triton™ X-100 for 10 minutes, and blocked with 1% BSA for 1 hour at room temperature. The cells were labeled with BTRC (1B1D2) Mouse Monoclonal Antibody (Product # 37-3400) at 2 µg/mL in 0.1% BSA and incubated for 3 hours at room temperature and then labeled with Goat anti-Mouse IgG (H+L) Superclonal™ Secondary Antibody, Alexa Fluor® 488 conjugate (Product # A28175) a dilution of 1:2000 for 45 minutes at room temperature (Panel a: green). Nuclei (Panel b: blue) were stained with SlowFade® Gold Antifade Mountant with DAPI (Product # S36938). F-actin (Panel c: red) was stained with Alexa Fluor® 555 Rhodamine Phalloidin (Product # R415, 1:300). Panel d represents the merged image showing cytoplasmic localization. Panel e shows the no primary antibody control. The images were captured at 60X magnification.
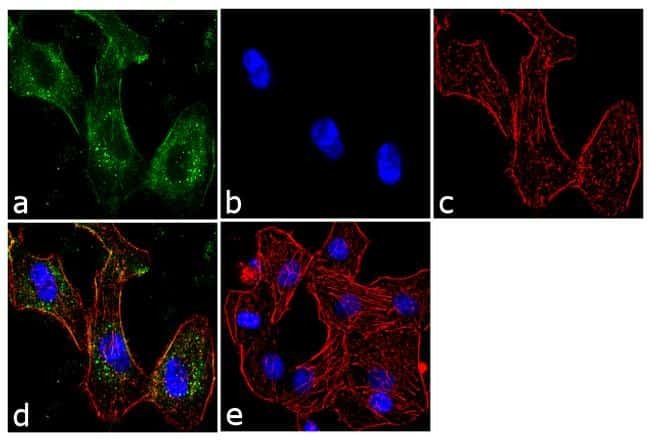
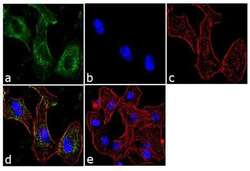
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- Immunofluorescent analysis of BTRC was performed using 70% confluent log phase A549 cells. The cells were fixed with 4% paraformaldehyde for 10 minutes, permeabilized with 0.1% Triton™ X-100 for 10 minutes, and blocked with 1% BSA for 1 hour at room temperature. The cells were labeled with BTRC (1B1D2) Mouse Monoclonal Antibody (Product # 37-3400) at 2 µg/mL in 0.1% BSA and incubated for 3 hours at room temperature and then labeled with Goat anti-Mouse IgG (H+L) Superclonal™ Secondary Antibody, Alexa Fluor® 488 conjugate (Product # A28175) a dilution of 1:2000 for 45 minutes at room temperature (Panel a: green). Nuclei (Panel b: blue) were stained with SlowFade® Gold Antifade Mountant with DAPI (Product # S36938). F-actin (Panel c: red) was stained with Alexa Fluor® 555 Rhodamine Phalloidin (Product # R415, 1:300). Panel d represents the merged image showing cytoplasmic localization. Panel e shows the no primary antibody control. The images were captured at 60X magnification.
Supportive validation
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
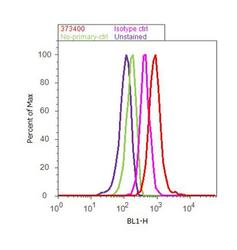
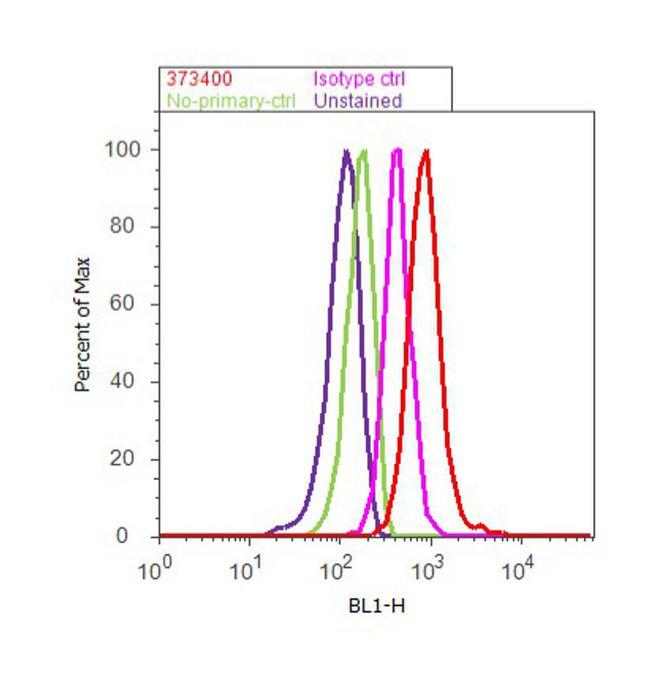
- Flow cytometry analysis of BTRC was done on A549 cells. Cells were fixed with 70% ethanol for 10 minutes, permeabilized with 0.25% Triton™ X-100 for 20 minutes, and blocked with 5% BSA for 30 minutes at room temperature. Cells were labeled with BTRC Mouse Monoclonal Antibody (37-3400, red histogram) or with mouse isotype control (pink histogram) at 3-5 ug/million cells in 2.5% BSA. After incubation at room temperature for 2 hours, the cells were labeled with Alexa Fluor® 488 Rabbit Anti-Mouse Secondary Antibody (A11059) at a dilution of 1:400 for 30 minutes at room temperature. The representative 10, 000 cells were acquired and analyzed for each sample using an Attune® Acoustic Focusing Cytometer. The purple histogram represents unstained control cells and the green histogram represents no-primary-antibody control.
Supportive validation
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
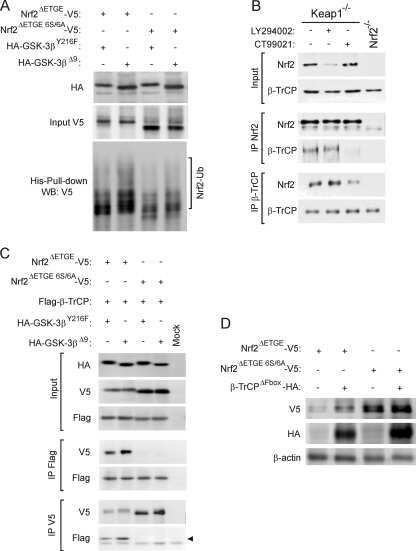
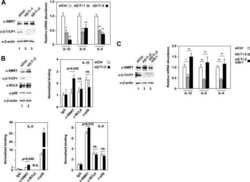
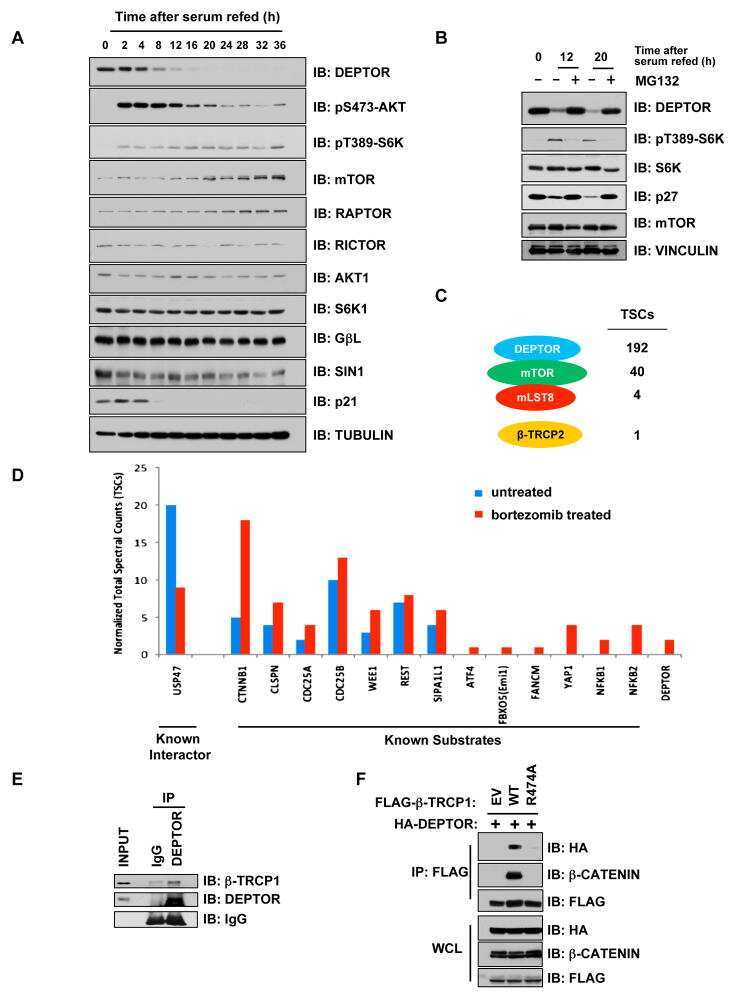
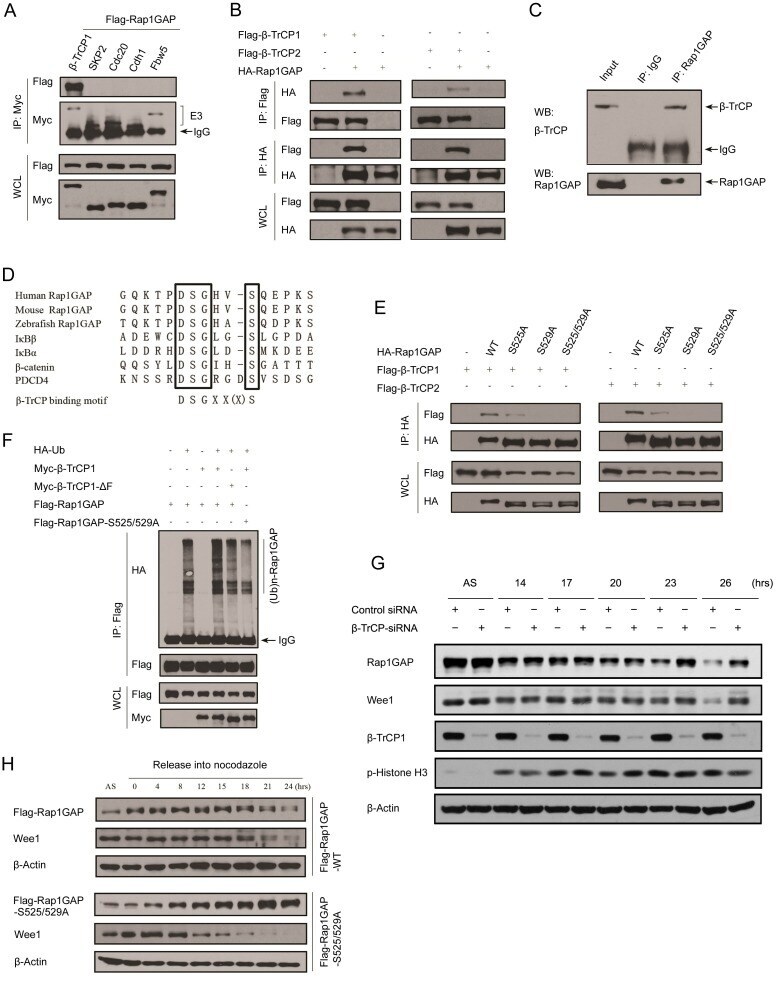
- Figure 2 Rap1GAP is ubiquitinated and degraded in a beta-TrCP dependent manner. (A) 293T cells were co-transfected with Flag-Rap1GAP and the indicated Myc-tagged E3 ligases constructs (beta-TrCP1, SKP2, Cdc20, Cdh1 and Fbw5). At 24 h after transfection, cells were harvested and lysed. Cell lysates were subjected to immunoprecipitation with anti-Myc antibody. The immunoprecipitates were immunoblotted with anti-Flag and anti-Myc antibodies, respectively. (B) 293T cells were co-transfected with HA-Rap1GAP and Flag- beta-TrCP1/2 constructs. Cell lysates were prepared and subjected to immunoprecipitation with anti-HA or anti-Flag antibodies, respectively. (C) Endogenous Rap1GAP interacts with endogenous beta-TrCP in HeLa cells. At the 4 h before harvesting, cells were treated with the proteasome inhibitor, MG132. HeLa cell lysates were incubated with protein A/G sepharose conjugated with either control IgG or Rap1GAP antibody. The immunoprecipitates were immunoblotted with Rap1GAP or beta-TrCP antibodies, respectively. (D) Alignment of amino acids corresponding to the DSGxxS sequence with Rap1GAP1 orthologs and other beta-TrCP substrates. (E) 293T cells were co-transfected with Flag-beta-TrCP1/2 and wild type HA-Rap1GAP or SA mutant (S525A, S529A and S525/529A) constructs. At 24 h after transfection, cells were harvested and lysed. Cell lysates were subjected to immunoprecipitation with anti-HA antibody. The immunoprecipitates were immunoblotted with anti-Flag and anti-HA antibodi
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
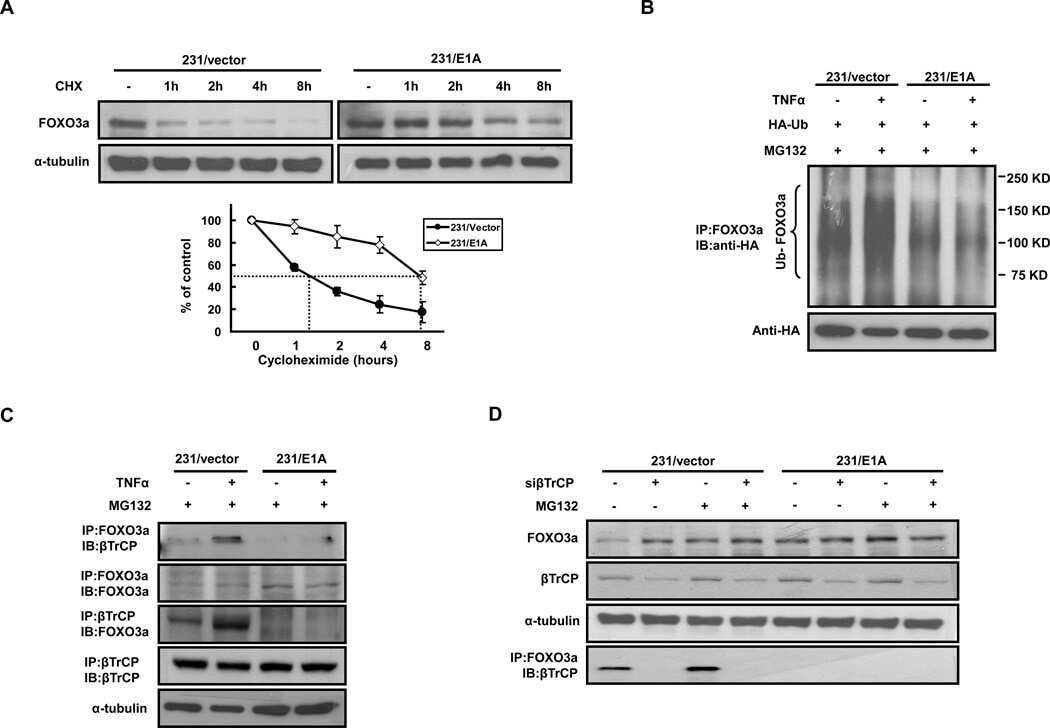
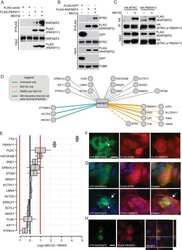
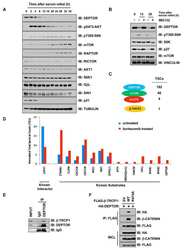
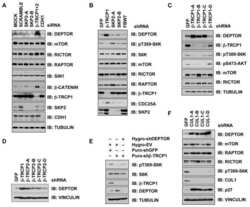
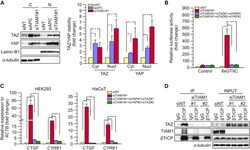
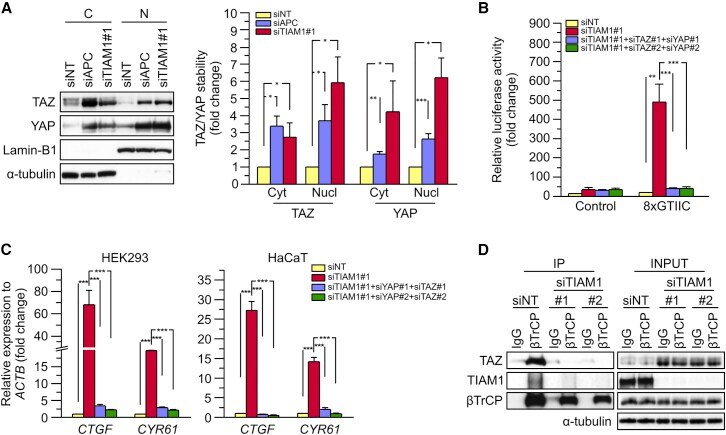
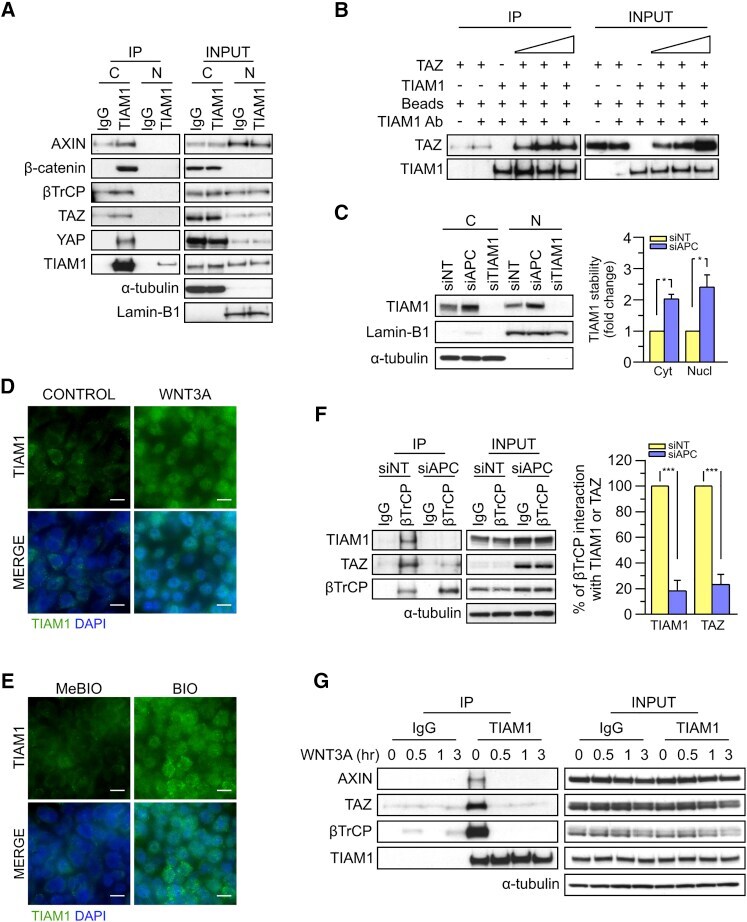
- Figure 4 TIAM1 Regulates TAZ Stability and Nuclear Translocation (A) Western blot analyses of cytoplasmic (C) and nuclear (N) fractions from confluent HEK293 cells treated with APC or TIAM1#1 siRNAs. Graph shows mean TAZ and YAP levels normalized to cytoplasmic (alpha-tubulin) or nuclear (Lamin-B1) markers and then to negative control siRNA (siNT). (B) Luciferase assay for 8xGTIIC-Lux or control reporter indicating TAZ/YAP-dependent transcriptional activity in confluent HEK293 cells transfected with TIAM1#1 siRNA, either alone or with two different pairs of TAZ/YAP siRNAs as indicated. Data are normalized to a Renilla reporter and to negative control siRNA (siNT). (C) qPCR for TAZ/YAP target genes CTGF and CYR61 normalized to ACTB expression in confluent HEK293 and HaCaT cells transfected with TIAM1#1 siRNA, either alone or with two different pairs of TAZ/YAP siRNAs as indicated. Data are normalized to negative control siRNA (siNT). (D) Western blot analyses of endogenous TAZ and TIAM1 co-immunoprecipitating (IP) with endogenous betaTrCP from confluent HEK293 cells treated with TIAM1#1 or TIAM1#2 siRNA. Immunoglobulin G (IgG) was used as a control antibody for immunoprecipitation. Figure is representative of three independent experiments. Data shown in graphs are presented as mean +- SEM with siNT set as 1 (unpaired t test for three independent experiments: * p < 0.05, ** p < 0.01, *** p < 0.001). See also Figure S4 .
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
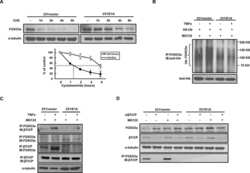
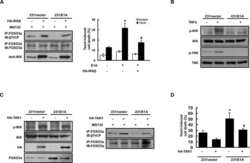
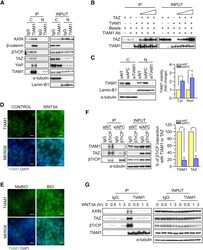
- Figure 5 TIAM1 is Regulated by the Canonical WNT Pathway (A) Western blot analyses of endogenous proteins co-immunoprecipitating with endogenous TIAM1 from cytoplasmic (C) and nuclear (N) fractions of confluent HEK293 cells. IgG was used as a control antibody for immunoprecipitation. Figure is representative of three independent experiments. (B) Western blot analyses of increasing amounts of recombinant TAZ immunoprecipitated with recombinant TIAM1. Beads alone, no recombinant TIAM1, or no recombinant TAZ were used as negative controls. Figure is representative of three independent experiments. (C) Western blot analyses of cytoplasmic (C) and nuclear (N) fractions from confluent HEK293 cells transfected with APC siRNA. Lysates from TIAM1-depleted cells (TIAM1#1 siRNA) were used as specificity control for the TIAM1 antibody. Graph shows mean TIAM1 levels normalized to cytoplasmic (alpha-tubulin) or nuclear (Lamin-B1) markers and then to negative control siRNA (siNT). (D and E) Representative confocal images of TIAM1 localization in confluent RKO cells treated with control or WNT3A-conditioned medium (D) or control (MeBIO) or GSK3a inhibitor (BIO) (E). Images are representative of three independent experiments. Scale bars, 10 mum. (F) Western blot analyses of endogenous TIAM1 and TAZ co-immunoprecipitating (IP) with endogenous betaTrCP from confluent HEK293 cells treated with APC siRNA. IgG was used as a control antibody for immunoprecipitation. Graph shows average betaTrCP/TIA
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
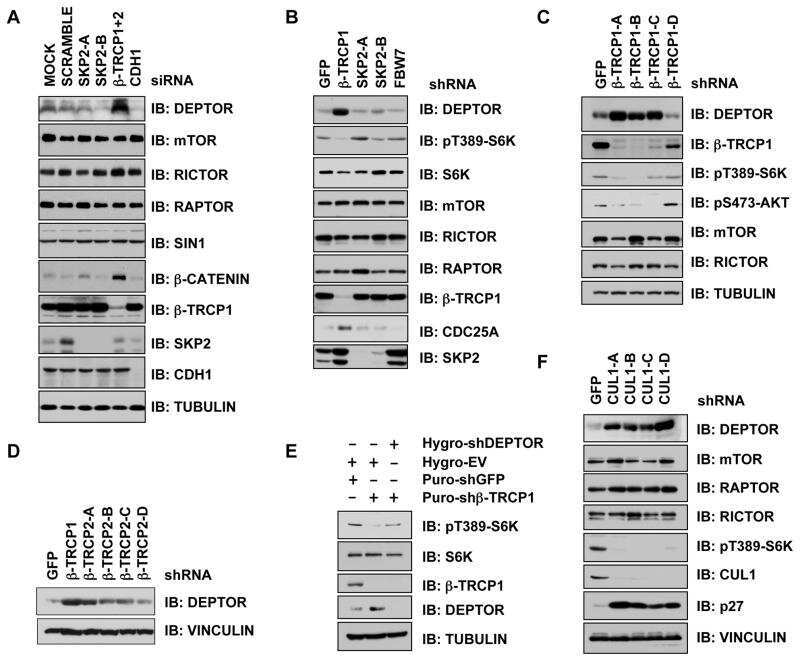
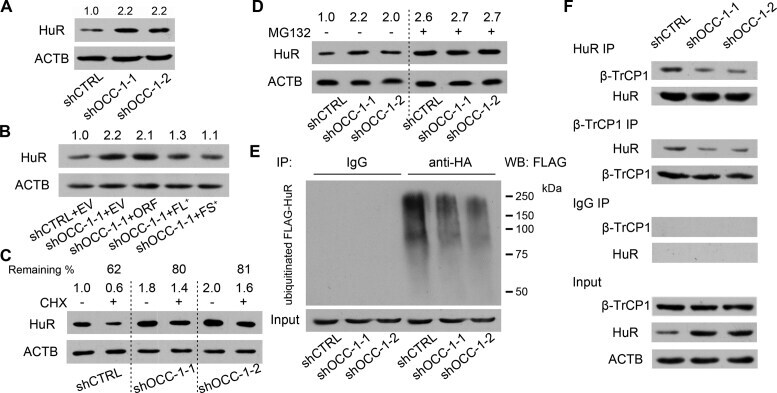
- Figure 6. OCC-1 promotes the ubiquitination and degradation of HuR by enhancing its binding to the ubiquitin E3 ligase beta-TrCP1. ( A ) Western blot analysis showed that knockdown of OCC-1 led to the increase of HuR in Caco-2 cells. ( B ) Overxpression of OCC-1 FL + and FS + RNA but not the ORF reduced HuR level in shOCC-1-1 Caco-2 cells. EV, empty vector. ( C ) Cells were treated with CHX to inhibit protein synthesis and the remaining of HuR was measured by western blot in Caco-2 cells after OCC-1 knockdown. The percent of remaining HuR in the CHX treated cells (+) relative to the control cells treated with DMSO (-) was indicated. ( D ) Western blot analysis of HuR in the OCC-1 knockdown cells treated with MG132 (+) or DMSO (-). The density of protein bands was measured by Image J software and the relative level of HuR protein was calculated after normalizing to ACTB protein. ( E ) The OCC-1 knockdown cells were co-transfected with a plasmid expressing a HA-tagged ubiquitin (HA-Ub) and a plasmid expressing a FLAG-tagged HuR (FLAG-HuR). After MG132 treatment, cell lysates were prepared and subjected to IP using an anti-HA antibody. The ubiquitinated FLAG-HuR was further detected by western blot using an anti-FLAG antibody. ( F ) Co-IP experiments showed that the interaction between HuR and the ubiquitin E3 ligase beta-TrCP1 was attenuated after OCC-1 knockdown in Caco-2 cells.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
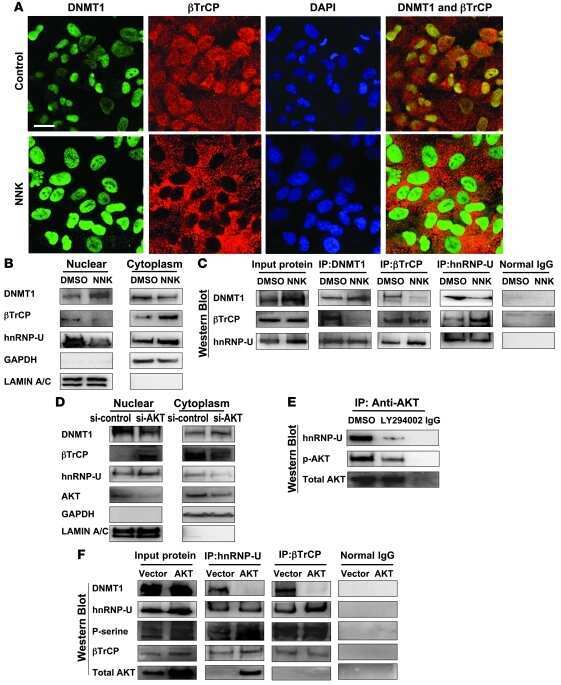
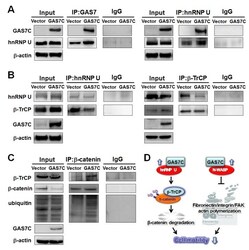
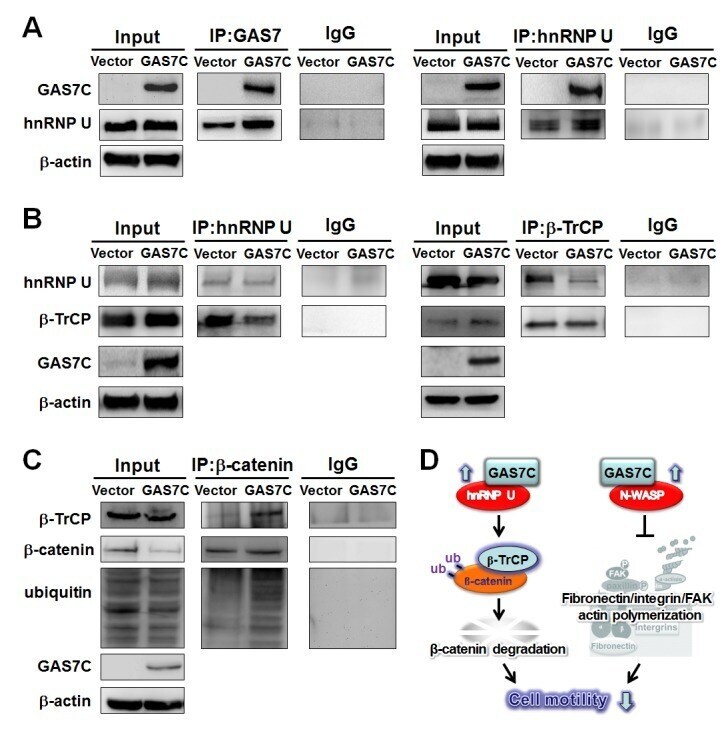
- Figure 5 GAS7C protein is associated with hnRNP U, which is involved in the beta-TrCP-mediated beta-catenin degradation pathway A. Protein lysates of A549 cells expressing control or GAS7C vector underwent IP with anti-GAS7 (left) or anti-hnRNP U (right) antibody followed by Western blot analysis. Normal IgG served as a negative control. GAS7C and hnRNP-U were found in the same protein complex. B. Reduced protein-protein interactions between hnRNP U and beta-TrCP in cells overexpressing GAS7C were confirmed by IP-Western blotting. C. The interaction between beta-catenin and beta-TrCP was increased along with an enhancement of ubiquitinated beta-catenin level upon GAS7C overexpression. D. Schematic presentation using a lung model of how GAS7C inhibits cell mobility by interacting with hnRNP U to promote beta-catenin degradation by beta-TrCP (left) and by interacting with N-WASP to suppress fibronectin/integrin/FAK/actin polymerization (right).
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
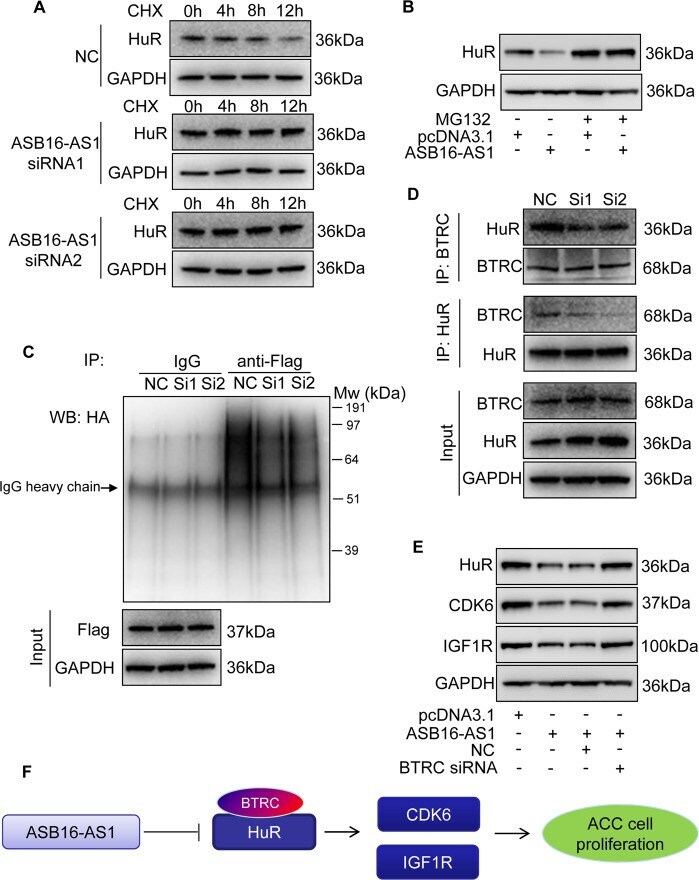
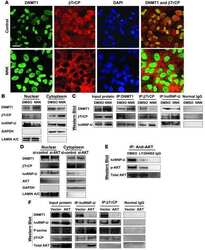
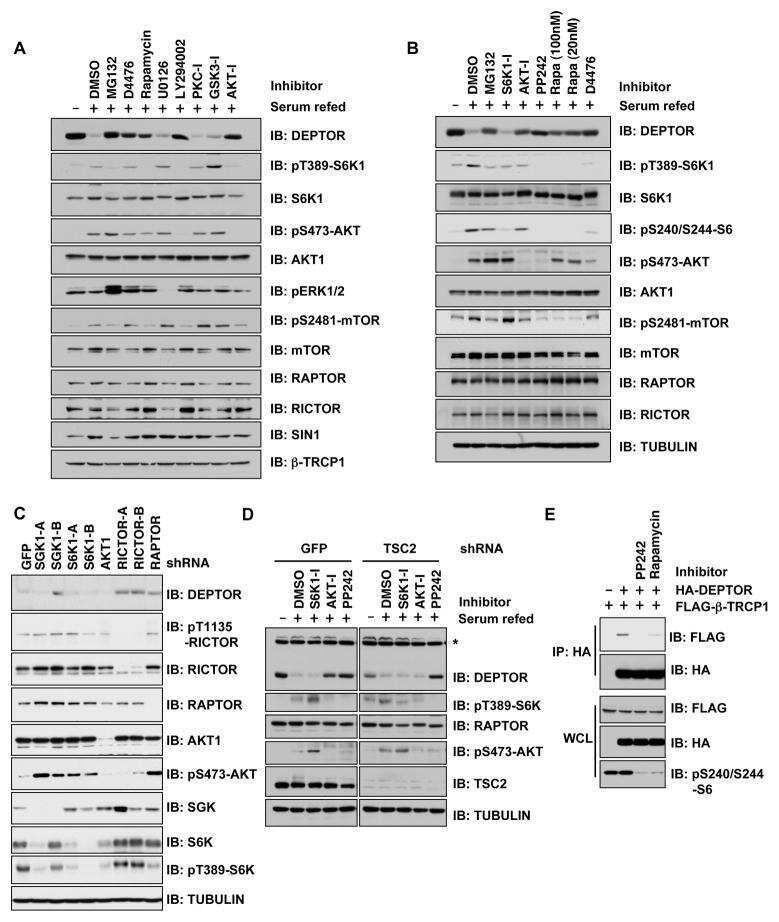
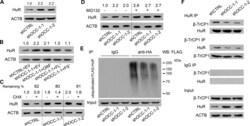
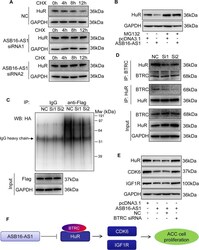
- Fig. 7 ASB16-AS1 post-translationally represses HuR expression through BTRC-mediated ubiquitination. a Inhibition of ASB16-AS1 expression attenuates HuR protein degradation. SW-13 cells were transfected with ASB16-AS1 siRNA1 or siRNA2 or its negative control (NC) and then treated with 50 mug/ml cycloheximide (CHX) for the indicated period of time. HuR protein levels were detected by western blot. b ASB16-AS1 promotes HuR degradation via proteasomal degradation. SW-13 cells were transfected with a plasmid encoding ASB16-AS1 or pcDNA3.1 control and cells were treated with 20 muM MG132 for 24 h. HuR protein levels were detected by western blot. c ASB16-AS1 promotes HuR protein ubiquitination. SW-13 cells were co-transfected with HA-Ub, Flag-HuR, ASB16-AS1 siRNA1 (Si1), or ASB16-AS1 siRNA2 (Si2) or its negative control (NC), the cells were then treated with 20 muM MG132 for 24 h. The cells were then lysed for immunoprecipitation. Immunoprecipitation was performed using anti-Flag antibody. IgG served as negative control. Western blot was used to detect ubiquitinated HuR protein using anti-HA antibody. d Inhibition of ASB16-AS1 expression attenuates the interaction between HuR and the ubiquitin E3 ligase BTRC. SW-13 cells were transfected with ASB16-AS1 siRNA1 (Si1) or ASB16-AS1 siRNA2 (Si2), immunoprecipitation was performed using either anti-HuR or anti-BTRC antibody, western blot was performed to detect the association between HuR and BTRC protein. e Knockdown of BTRC abolished