Explore
Explore Validate
Validate Learn
Learn44-696G
antibody from Invitrogen Antibodies
Targeting: PLCG1
NCKAP3, PLC-II, PLC1, PLC148, PLCgamma1
 Western blot
Western blot Immunocytochemistry
ImmunocytochemistryAntibody data
- Antibody Data
- Antigen structure
- References [18]
- Comments [0]
- Validations
- Immunocytochemistry [2]
- Other assay [10]
Submit
Validation data
Reference
Comment
Report error
- Product number
- 44-696G - Provider product page

- Provider
- Invitrogen Antibodies
- Product name
- Phospho-PLCG1 (Tyr783) Polyclonal Antibody
- Antibody type
- Polyclonal
- Antigen
- Synthetic peptide
- Reactivity
- Human, Mouse
- Host
- Rabbit
- Isotype
- IgG
- Vial size
- 100 μL
- Storage
- -20°C
Submitted references Paladin is a phosphoinositide phosphatase regulating endosomal VEGFR2 signalling and angiogenesis.
Downregulation of PUMA underlies resistance to FGFR1 inhibitors in the stem cell leukemia/lymphoma syndrome.
ADAP is an upstream regulator that precedes SLP-76 at sites of TCR engagement and stabilizes signaling microclusters.
THEMIS enhances TCR signaling and enables positive selection by selective inhibition of the phosphatase SHP-1.
RasGRP1 overexpression in T-ALL increases basal nucleotide exchange on Ras rendering the Ras/PI3K/Akt pathway responsive to protumorigenic cytokines.
LynA regulates an inflammation-sensitive signaling checkpoint in macrophages.
Small molecule inhibition of Csk alters affinity recognition by T cells.
Pak2 is required for actin cytoskeleton remodeling, TCR signaling, and normal thymocyte development and maturation.
Pak2 is required for actin cytoskeleton remodeling, TCR signaling, and normal thymocyte development and maturation.
Inhibition of the kinase Csk in thymocytes reveals a requirement for actin remodeling in the initiation of full TCR signaling.
Tetraspanin CD63 promotes vascular endothelial growth factor receptor 2-β1 integrin complex formation, thereby regulating activation and downstream signaling in endothelial cells in vitro and in vivo.
Quantitative differences in CD45 expression unmask functions for CD45 in B-cell development, tolerance, and survival.
PEST motif serine and tyrosine phosphorylation controls vascular endothelial growth factor receptor 2 stability and downregulation.
Vitamin D controls T cell antigen receptor signaling and activation of human T cells.
Paxillin-Y118 phosphorylation contributes to the control of Src-induced anchorage-independent growth by FAK and adhesion.
Stability of an autoinhibitory interface in the structure of the tyrosine kinase ZAP-70 impacts T cell receptor response.
Intramolecular regulatory switch in ZAP-70: analogy with receptor tyrosine kinases.
Functional tyrosine kinase inhibitor profiling: a generally applicable method points to a novel role of platelet-derived growth factor receptor-beta in tuberous sclerosis.
Nitzsche A, Pietilä R, Love DT, Testini C, Ninchoji T, Smith RO, Ekvärn E, Larsson J, Roche FP, Egaña I, Jauhiainen S, Berger P, Claesson-Welsh L, Hellström M
EMBO reports 2021 Feb 3;22(2):e50218
EMBO reports 2021 Feb 3;22(2):e50218
Downregulation of PUMA underlies resistance to FGFR1 inhibitors in the stem cell leukemia/lymphoma syndrome.
Liu Y, Cai B, Chong Y, Zhang H, Kemp CA, Lu S, Chang CS, Ren M, Cowell JK, Hu T
Cell death & disease 2020 Oct 20;11(10):884
Cell death & disease 2020 Oct 20;11(10):884
ADAP is an upstream regulator that precedes SLP-76 at sites of TCR engagement and stabilizes signaling microclusters.
Lewis JB, Scangarello FA, Murphy JM, Eidell KP, Sodipo MO, Ophir MJ, Sargeant R, Seminario MC, Bunnell SC
Journal of cell science 2018 Nov 8;131(21)
Journal of cell science 2018 Nov 8;131(21)
THEMIS enhances TCR signaling and enables positive selection by selective inhibition of the phosphatase SHP-1.
Choi S, Warzecha C, Zvezdova E, Lee J, Argenty J, Lesourne R, Aravind L, Love PE
Nature immunology 2017 Apr;18(4):433-441
Nature immunology 2017 Apr;18(4):433-441
RasGRP1 overexpression in T-ALL increases basal nucleotide exchange on Ras rendering the Ras/PI3K/Akt pathway responsive to protumorigenic cytokines.
Ksionda O, Melton AA, Bache J, Tenhagen M, Bakker J, Harvey R, Winter SS, Rubio I, Roose JP
Oncogene 2016 Jul 14;35(28):3658-68
Oncogene 2016 Jul 14;35(28):3658-68
LynA regulates an inflammation-sensitive signaling checkpoint in macrophages.
Freedman TS, Tan YX, Skrzypczynska KM, Manz BN, Sjaastad FV, Goodridge HS, Lowell CA, Weiss A
eLife 2015 Oct 30;4
eLife 2015 Oct 30;4
Small molecule inhibition of Csk alters affinity recognition by T cells.
Manz BN, Tan YX, Courtney AH, Rutaganira F, Palmer E, Shokat KM, Weiss A
eLife 2015 Aug 24;4
eLife 2015 Aug 24;4
Pak2 is required for actin cytoskeleton remodeling, TCR signaling, and normal thymocyte development and maturation.
Phee H, Au-Yeung BB, Pryshchep O, O'Hagan KL, Fairbairn SG, Radu M, Kosoff R, Mollenauer M, Cheng D, Chernoff J, Weiss A
eLife 2014 May 13;3:e02270
eLife 2014 May 13;3:e02270
Pak2 is required for actin cytoskeleton remodeling, TCR signaling, and normal thymocyte development and maturation.
Phee H, Au-Yeung BB, Pryshchep O, O'Hagan KL, Fairbairn SG, Radu M, Kosoff R, Mollenauer M, Cheng D, Chernoff J, Weiss A
eLife 2014 May 13;3:e02270
eLife 2014 May 13;3:e02270
Inhibition of the kinase Csk in thymocytes reveals a requirement for actin remodeling in the initiation of full TCR signaling.
Tan YX, Manz BN, Freedman TS, Zhang C, Shokat KM, Weiss A
Nature immunology 2014 Feb;15(2):186-94
Nature immunology 2014 Feb;15(2):186-94
Tetraspanin CD63 promotes vascular endothelial growth factor receptor 2-β1 integrin complex formation, thereby regulating activation and downstream signaling in endothelial cells in vitro and in vivo.
Tugues S, Honjo S, König C, Padhan N, Kroon J, Gualandi L, Li X, Barkefors I, Thijssen VL, Griffioen AW, Claesson-Welsh L
The Journal of biological chemistry 2013 Jun 28;288(26):19060-71
The Journal of biological chemistry 2013 Jun 28;288(26):19060-71
Quantitative differences in CD45 expression unmask functions for CD45 in B-cell development, tolerance, and survival.
Zikherman J, Doan K, Parameswaran R, Raschke W, Weiss A
Proceedings of the National Academy of Sciences of the United States of America 2012 Jan 3;109(1):E3-12
Proceedings of the National Academy of Sciences of the United States of America 2012 Jan 3;109(1):E3-12
PEST motif serine and tyrosine phosphorylation controls vascular endothelial growth factor receptor 2 stability and downregulation.
Meyer RD, Srinivasan S, Singh AJ, Mahoney JE, Gharahassanlou KR, Rahimi N
Molecular and cellular biology 2011 May;31(10):2010-25
Molecular and cellular biology 2011 May;31(10):2010-25
Vitamin D controls T cell antigen receptor signaling and activation of human T cells.
von Essen MR, Kongsbak M, Schjerling P, Olgaard K, Odum N, Geisler C
Nature immunology 2010 Apr;11(4):344-9
Nature immunology 2010 Apr;11(4):344-9
Paxillin-Y118 phosphorylation contributes to the control of Src-induced anchorage-independent growth by FAK and adhesion.
Sachdev S, Bu Y, Gelman IH
BMC cancer 2009 Jan 12;9:12
BMC cancer 2009 Jan 12;9:12
Stability of an autoinhibitory interface in the structure of the tyrosine kinase ZAP-70 impacts T cell receptor response.
Deindl S, Kadlecek TA, Cao X, Kuriyan J, Weiss A
Proceedings of the National Academy of Sciences of the United States of America 2009 Dec 8;106(49):20699-704
Proceedings of the National Academy of Sciences of the United States of America 2009 Dec 8;106(49):20699-704
Intramolecular regulatory switch in ZAP-70: analogy with receptor tyrosine kinases.
Brdicka T, Kadlecek TA, Roose JP, Pastuszak AW, Weiss A
Molecular and cellular biology 2005 Jun;25(12):4924-33
Molecular and cellular biology 2005 Jun;25(12):4924-33
Functional tyrosine kinase inhibitor profiling: a generally applicable method points to a novel role of platelet-derived growth factor receptor-beta in tuberous sclerosis.
Arbiser JL, Govindarajan B, Bai X, Onda H, Kazlauskas A, Lim SD, Amin MB, Claesson-Welsh L
The American journal of pathology 2002 Sep;161(3):781-6
The American journal of pathology 2002 Sep;161(3):781-6
No comments: Submit comment
Supportive validation
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
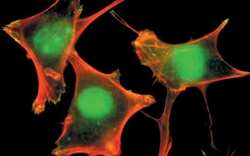
- Experimental details
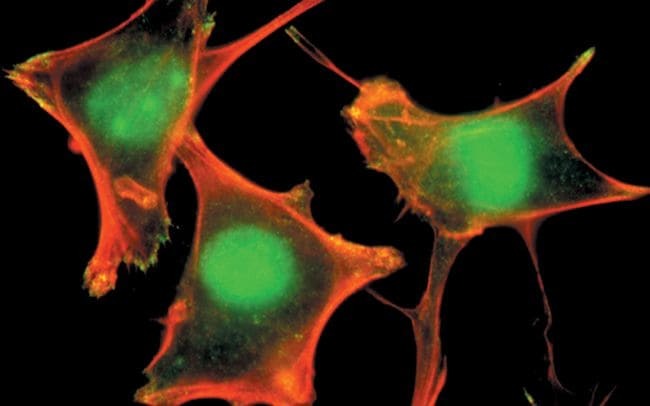
- PLC-1 (pY783) phosphospecific antibody, Colocalized detection of PLC-1 (pY783) (green) and actin stress fibers (red) at the sites of membrane ruffles and focal adhesions in fibroblasts.
- Submitted by
- Invitrogen Antibodies (provider)
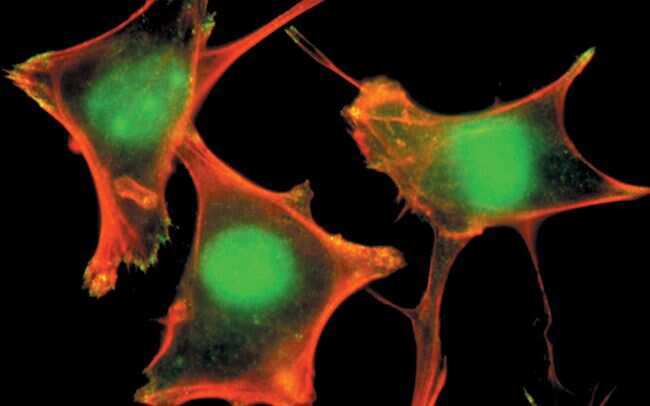
- Main image
- Experimental details
- PLC-1 (pY783) phosphospecific antibody, Colocalized detection of PLC-1 (pY783) (green) and actin stress fibers (red) at the sites of membrane ruffles and focal adhesions in fibroblasts.
Supportive validation
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
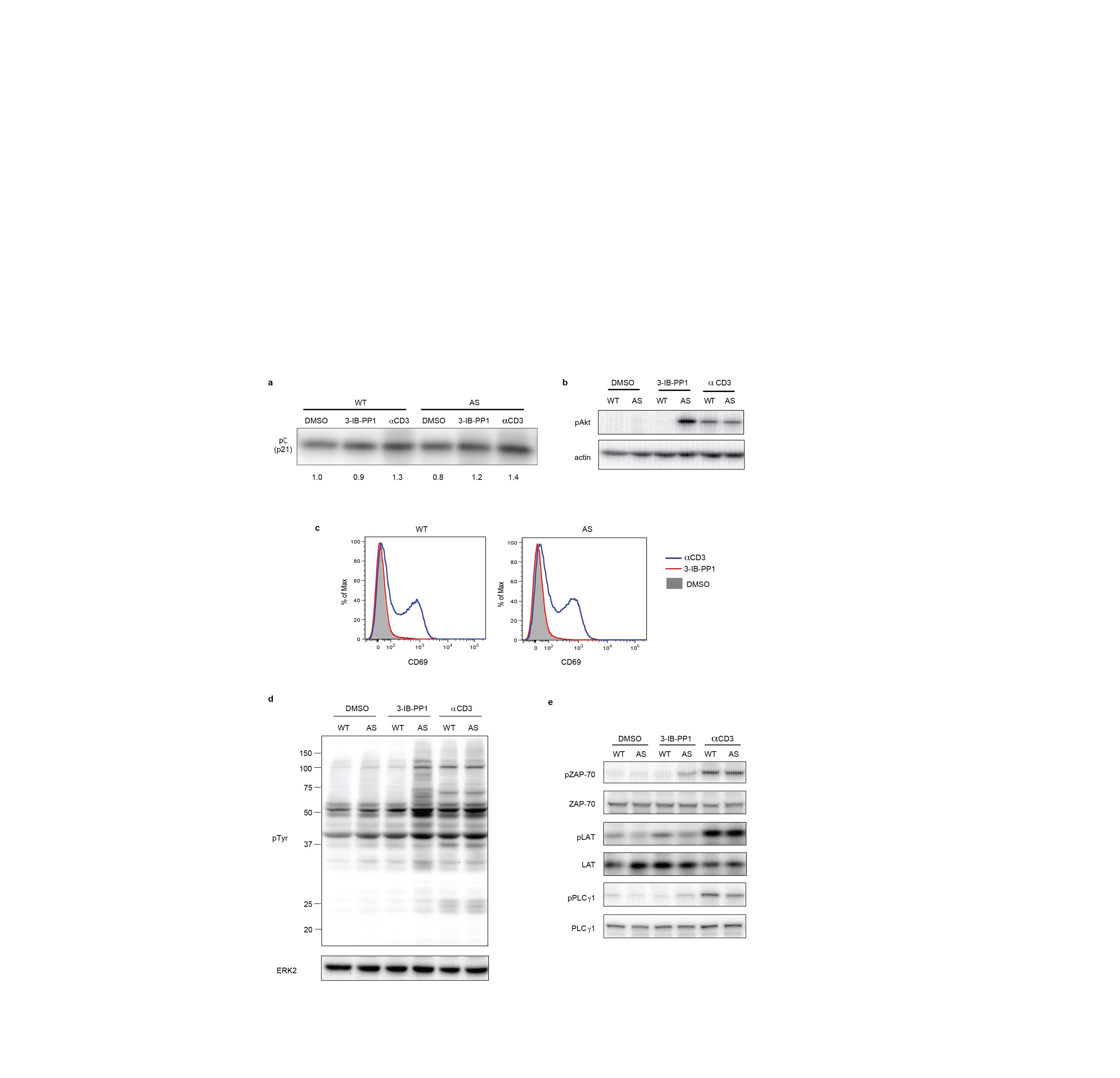
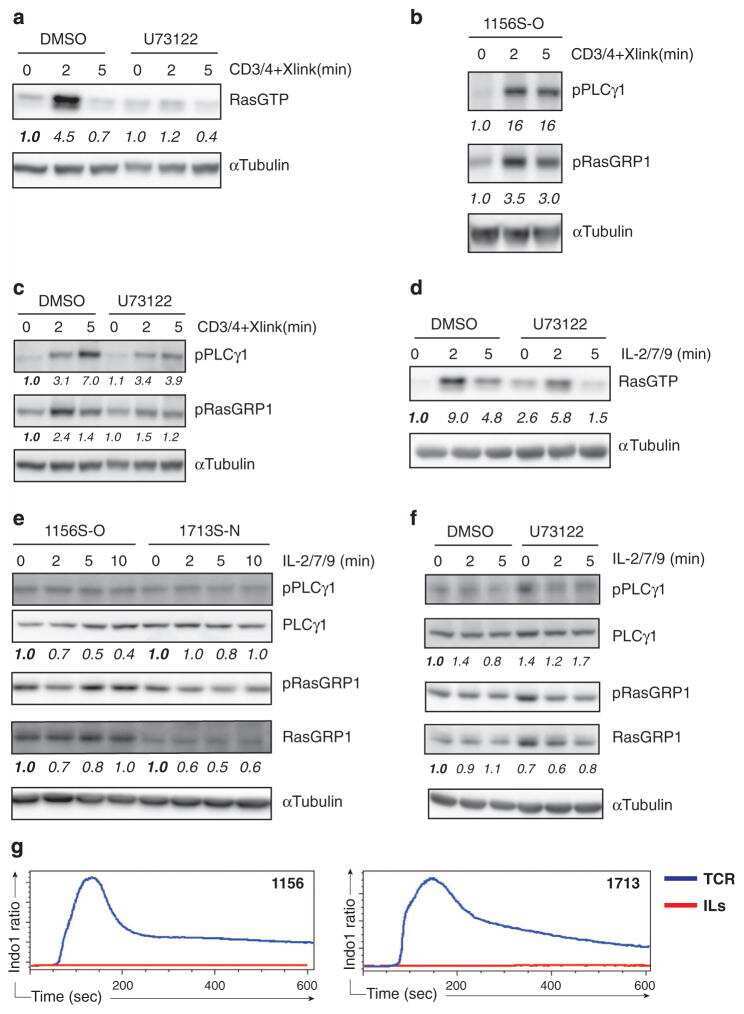
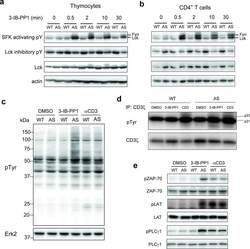
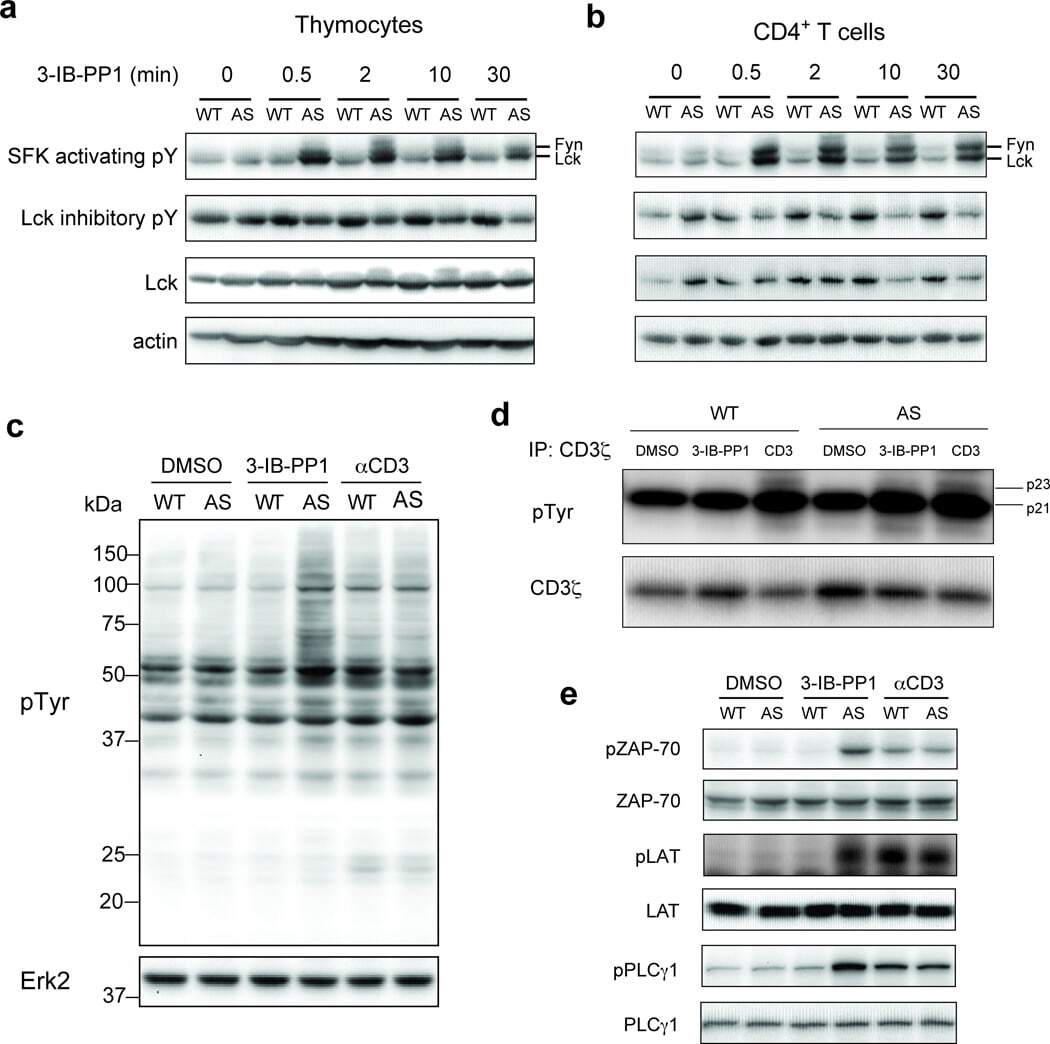
- Figure 1 Inhibition of Csk AS in primary murine T cells induces hyperactivation of SFKs and phosphorylation of TCR proximal signaling molecules. ( a ) Wild-type (WT) or Csk AS (AS) thymocytes or ( b ) peripheral CD4 + T cells were treated for the indicated times with 10 uM 3-IB-PP1 and analyzed by immunoblotting. ( c ) Wild-type (WT) or Csk AS (AS) thymocytes were treated for 3 min with vehicle (DMSO), 3-IB-PP1 or anti-CD3epsilon and analyzed by immunoblotting. ( d ) Wild-type (WT) or Csk AS (AS) thymocytes were treated for 3 min with vehicle (DMSO), 10uM 3-IB-PP1 or 20ug/mL anti-CD3epsilon, then lysed and immunoprecipitated with anti-CD3 zeta antibody. Immunoprecipitates were analyzed by immunoblotting. ( e ) Wild-type (WT) or Csk AS (AS) thymocytes were treated for 3 min with vehicle (DMSO), 10uM 3-IB-PP1 or 20ug/mL anti-CD3epsilon and analyzed by immunoblotting. All data are representative of 3 independent experiments.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
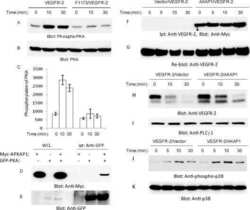
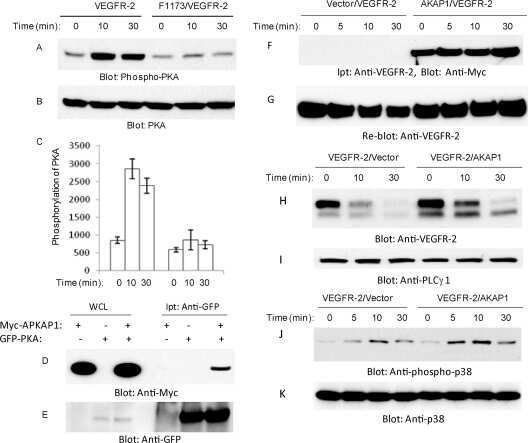
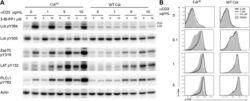
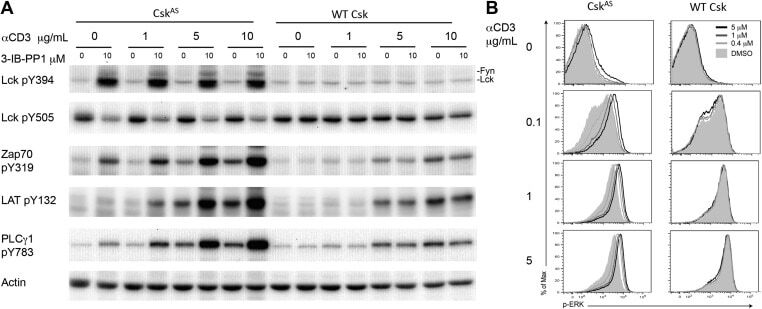
- Figure 1. Inhibiting Csk increases the magnitude of ligand-induced TCR signaling and reduces the threshold for TCR activation. ( A ) Purified Csk AS (AS) or wildtype (WT) CD4 + T cells stimulated for 2 min with 1 mug/ml, 5 mug/ml or 10 mug/ml anti-CD3epsilon antibody in the presence of DMSO or 10 muM 3-IB-PP1 were analyzed by immunoblotting for the phosphorylation of the activation loop tyrosine of Lck and Fyn (Src pY416 antibody) and the inhibitory tyrosine of Lck (Lck pY505), phosphorylated ZAP-70 (ZAP70 pY319), LAT (LAT pY132) and PLC-gamma1 (PLCgamma1 pY783), as well as total actin (loading control). Data are representative of at least three independent experiments. ( B ) Purified total Csk AS (AS) or wildtype (WT) T cells stimulated with the indicated dose of anti-CD3epsilon antibody for 2 min in the presence of DMSO or 5 muM, 1 muM or 0.4 muM 3-IB-PP1 were analyzed for phosphorylated ERK (p-ERK) by phosphoflow. Histograms were gated on CD4 + cells. Data are representative of at least three independent experiments for AS cells and two independent experiments for WT cells. DOI: http://dx.doi.org/ Figure 1--figure supplement 1. Inhibiting Csk during TCR stimulation prolongs TCR signals. ( A ) Purified Csk AS CD4 + T cells stimulated for 2, 5 or 10 min with anti-CD3epsilon antibody in the presence of DMSO or 10 muM 3-IB-PP1 were analyzed by immunoblotting for the phosphorylated ZAP-70, LAT and PLC-gamma1 as well as total GAPDH (loading control). Data are representative of a
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
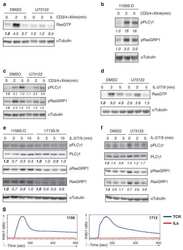
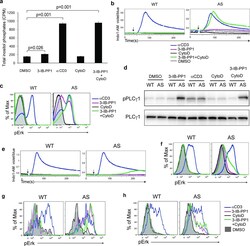
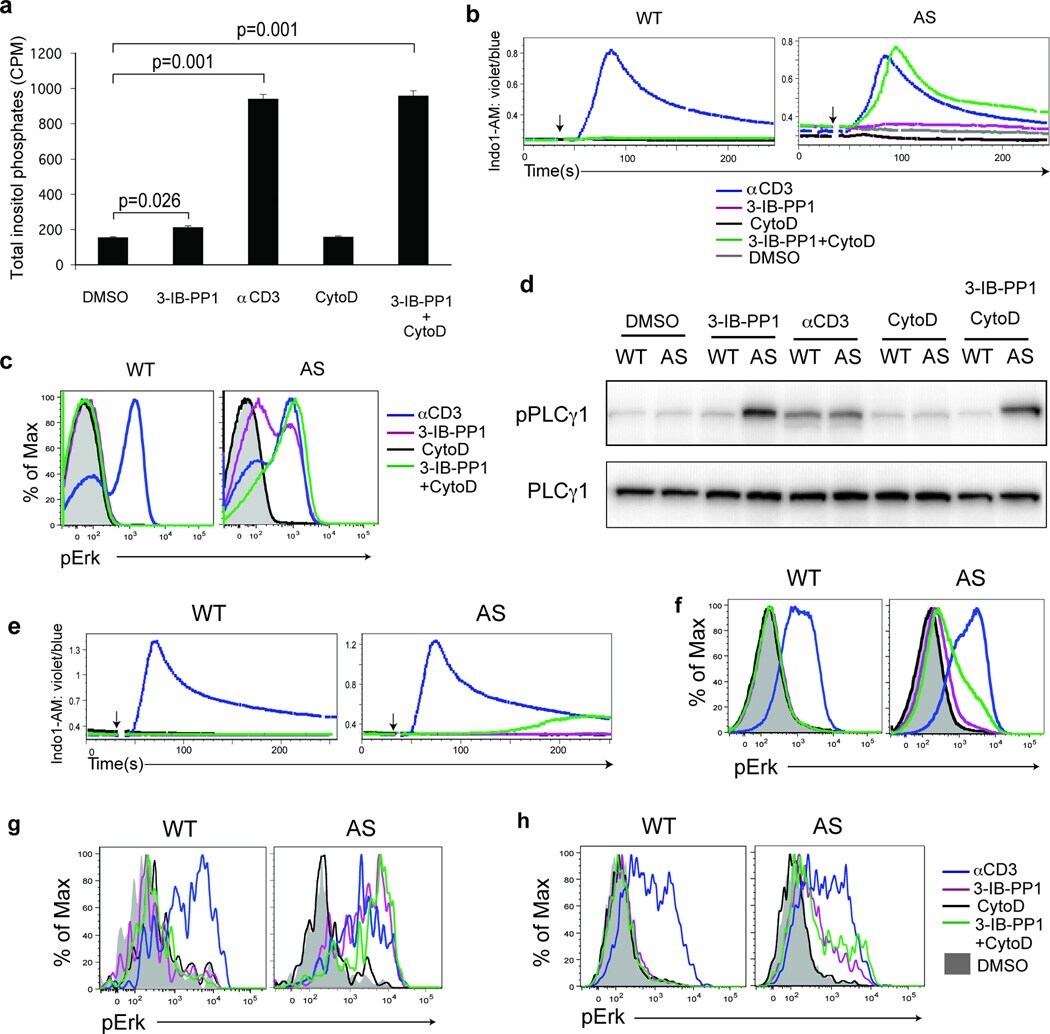
- Figure 3 Calcium flux and Erk phosphorylation are restored by altering the actin cytoskeleton following Csk AS inhibition. ( a ) Csk AS (AS) thymocytes were stimulated with vehicle (DMSO), 10uM 3-IB-PP1, 20ug/mL anti-CD3epsilon, 10uM cytochalasin D (CytoD) alone or with 10uM 3-IB-PP1 for 5 min, and total inositol phosphate content was measured (mean, +- S.E.M., n = 3 (technical replicates), P -values from paired two-tailed student's t -test). ( b ) Wild-type (WT) or Csk AS (AS) thymocytes were stimulated with vehicle (DMSO: gray), 10uM 3-IB-PP1 (magenta), 20ug/mL anti-CD3epsilon (blue), 10uM cytochalasin D (CytoD: black) alone or with 10uM 3-IB-PP1(green). Ratiometric assessment of intracellular calcium of CD4 + CD8 + thymocytes over time is shown. ( c ) Wild-type (WT) or Csk AS (AS) thymocytes were stimulated for 2 min as in b , then analyzed for pErk content. Histograms are gated on CD4 + CD8 + thymocytes. ( d ) Wild-type (WT) or Csk AS (AS) thymocytes were treated for 3 min as in a and analyzed by immunoblotting. ( e ) Wild-type (WT) or Csk AS (AS) splenocytes were stimulated as in b . Ratiometric assessment of intracellular calcium of CD4+ splenocytes over time is shown. ( f ) Wild-type (WT) or Csk AS (AS) splenocytes were stimulated for 2 min as in b , then analyzed for pErk content. Histograms are gated on CD4 + splenocytes. Shaded histogram is DMSO control. ( g ) Wild-type (WT) or Csk AS (AS) thymocytes or ( h ) purified T cells were stimulated for 2 min as in b ,
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
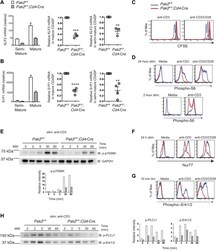
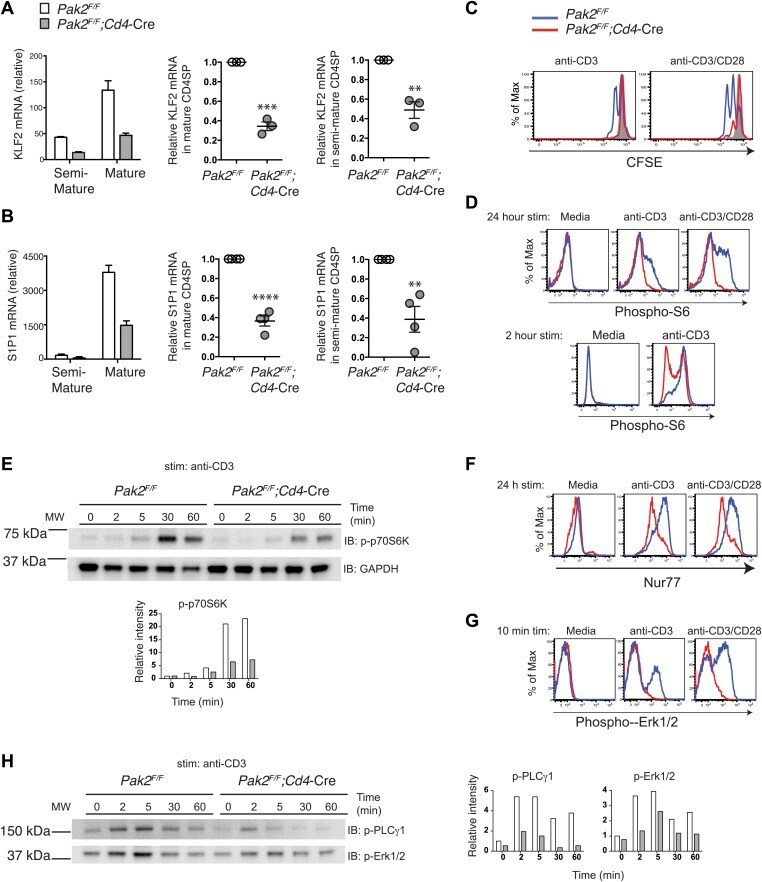
- Figure 7. Pak2 plays a key role in functional maturation of CD4SP thymocytes. ( A ) Defects in mRNA expression of KLF2 in mature CD4SP thymocytes from Pak2 F/F ;Cd4 -Cre mice. KLF2 mRNA levels in semi-mature and mature CD4SP thymocytes relative to Pak2 F/F DP thymocytes (left panel, mean +- SD of triplicates, results are representative of three independent experiment); KLF2 mRNA levels in mature Pak2 F/F ;Cd4 -Cre CD4SP thymocytes relative to mature Pak2 F/F CD4SP thymocytes (middle panel, mean +- SEM; each dot represents one mouse, n = 3); KLF2 mRNA levels in semi-mature Pak2 F/F ;Cd4 -Cre CD4SP thymocytes relative to semi-mature Pak2 F/F CD4SP thymocytes (right panel, mean +- SEM; each dot represents one mouse, n = 3). ***, p=0.0001; **, 0.001
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
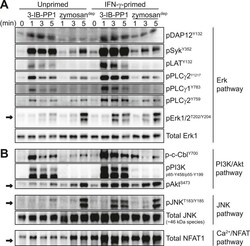
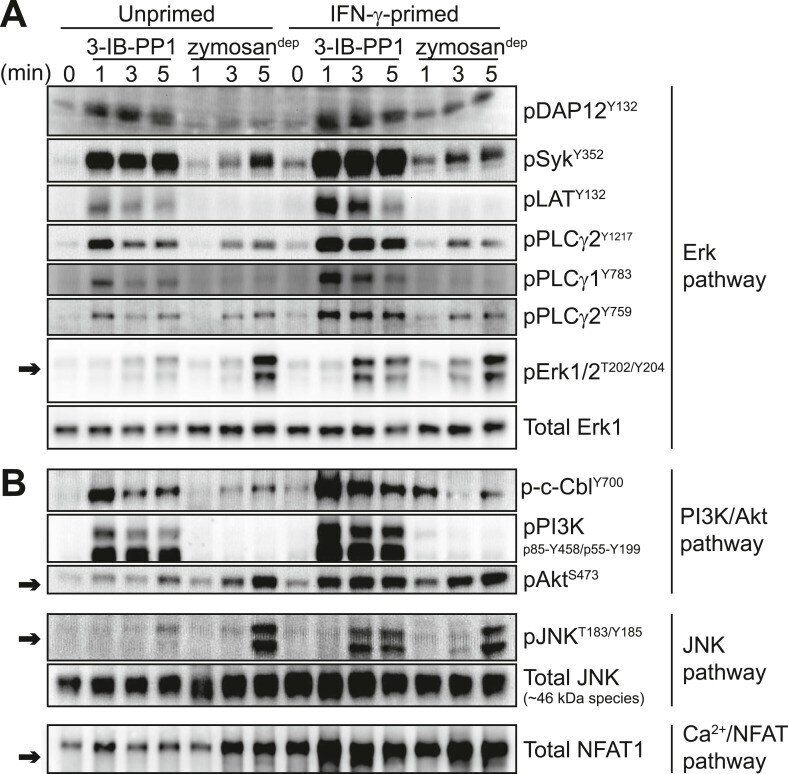
- Figure 5. Inflammatory priming is required for SFK activation to produce downstream signaling in the absence of receptor clustering. ( A ) Csk AS BMDMs were incubated 12-16 hr in non-priming medium or in medium containing 25 U/ml IFN-gamma. Signal transduction after 3-IB-PP1 or zymosan dep treatment was assessed by immunoblotting with antibodies specific to activating phosphorylation sites of DAP12, Syk, LAT, PLCgamma2, PLCgamma1, and Erk (arrow). Total Erk1 is shown as a loading control. ( B ) Signal transduction in the PI3K/Akt and JNK pathways was assessed by immunoblotting with antibodies specific for phosphorylated c-Cbl, PI3K, Akt (arrow), and JNK (arrow). Total JNK is shown as a loading control. NFAT1 activation was assessed by the faster migration of cellular NFAT1 upon dephosphorylation. See Figure 5--figure supplement 1 for further characterization of signaling in primed and unprimed cells. DOI: http://dx.doi.org/ Figure 5--figure supplement 1. Further characterization of signaling in primed and unprimed BMDMs. ( A ) Treatment of IFN-gamma-primed and unprimed Csk AS BMDMs with 3-IB-PP1 led to activating phosphorylation of the cytoskeleton-associated SFK substrates HS1, Vav, and FAK. ( B ) Comparison of background Erk phosphorylation in unprimed and IFN-gamma-primed Csk AS BMDMs with pulse-spin only. DOI: http://dx.doi.org/
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
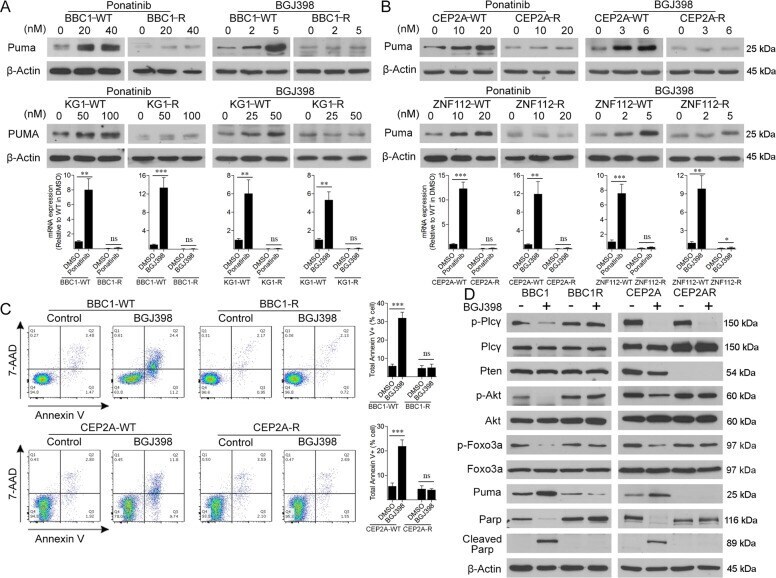
- Fig. 2 Imparied Puma activation contributes to TKI resitance SCLL. Western blot analysis of PUMA protein levels in the murine SCLL BBC1 cell line ( A , above) or human KG1 cell line, after treatment at the indicated concentrations of either the ponatinib or BGJ398 FGFR1 inhibitors, show a dose dependent increase in the parental cells, while no changes are seen in the resistant cells that carry FGFR1 mutations in the inhibitor binding site. The same effect is seen in in cell lines CEP2A and ZNF112, which are resistant due to Pten deletions ( B , above). RT-PCR analysis in all four cell lines shows that transcription levels of PUMA increases significantly in the cells treated ( A , B , below) with the FGFR1 inhibitors at the maximum concentrations shown in A and B (above). Flow cytometric analysis of BBC1 and CEP2A cells shows an increase in 7AAD/Annexin V+ cells in the parental cells treated with BGJ398 at the IC50, but no changes are observed in the resistant cells ( C ). Western blot analysis ( D ) of signaling intermediates demonstrates consistent Akt activation and Puma suppression in the resistant cells (see text) and phosphorylation of Foxo3a is suppressed in parental, but not in the resistant cells treated with BGJ398. Student's t -test was used for statistical comparison between groups. ** p < 0.01; *** p < 0.001; ns not significant.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
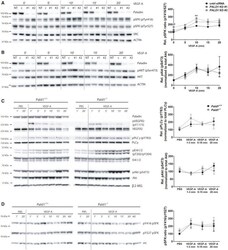
- Experimental details
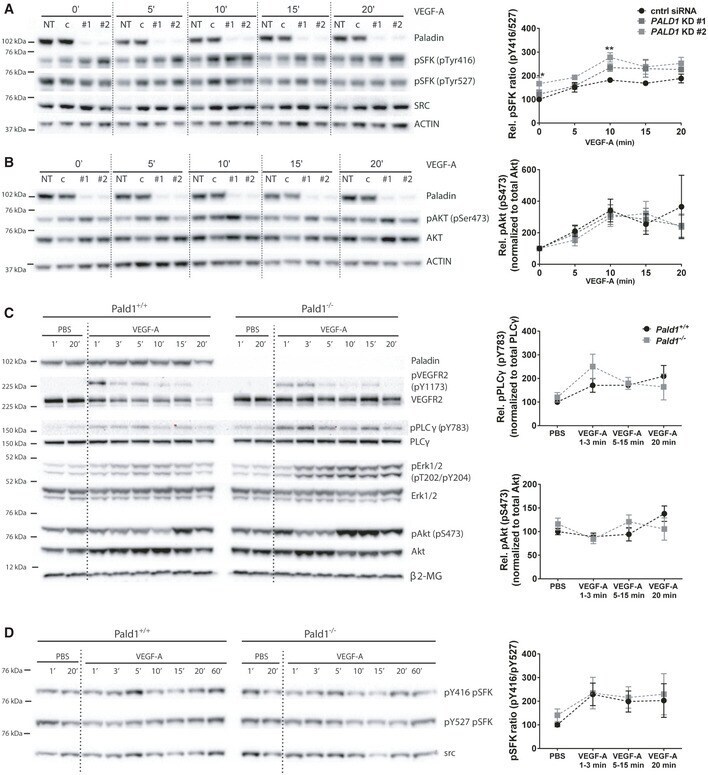
- Figure EV3 VEGF-A/VEGFR2 downstream signalling in vitro and in vivo A, B HDMEC, non-transfected (NT) or transfected with non-targeting (c/cntrl) or PALD1 -targeting siRNA (KD #1, KD #2), were treated with VEGF-A for the indicated time periods. Immunoblots of cell lysates to determine protein levels of ACTIN, Paladin, pTyr416, and pTyr527 of Src family kinases (SFK) and total Src (A), pSer473 AKT and total AKT (B), are shown (left) and their quantification (right). Mean +- SEM, two-way ANOVA, n = 3 biological replicates. C Immunoblotting of total heart lysates from Pald1 +/+ and Pald1 -/- mice, tail vein injected with VEGF-A or PBS for the indicated time points, for Paladin, phosphorylated and total levels of VEGFR2, phospholipase Cgamma (PLCgamma), Erk1/2, and Akt, and beta2-microglobulin (beta2-MG, loading control). Part of the same blot is also shown in Fig 3 . Quantification of pY783 PLCgamma normalized to PLCgamma ( n = 4 biological replicates) and pS473 Akt normalized to Akt ( n = 5 biological replicates), Mean +- SEM, two-way ANOVA. SSSS D Pald1 +/+ and Pald1 -/- mice were tail vein injected with VEGF-A or PBS for the indicated time periods. Heart lysates were blotted to determine the phosphorylated and total levels of Src family kinases (SFK). pY416 SFK normalized to pY527 SFK. Mean +- SEM, two-way ANOVA, n = 5 biological replicates. Data information: * P < 0.05, ** P < 0.01. Source data are available online for this figure.