Explore
Explore Validate
Validate Learn
Learn Western blot
Western blot ELISA
ELISA Immunohistochemistry
ImmunohistochemistryAntibody data
- Antibody Data
- Antigen structure
- References [61]
- Comments [0]
- Validations
- Immunohistochemistry [1]
- Other assay [42]
Submit
Validation data
Reference
Comment
Report error
- Product number
- 13-0200 - Provider product page

- Provider
- Invitrogen Antibodies
- Product name
- Amyloid Precursor Protein Monoclonal Antibody (LN27)
- Antibody type
- Monoclonal
- Antigen
- Other
- Reactivity
- Human
- Host
- Mouse
- Isotype
- IgG
- Antibody clone number
- LN27
- Vial size
- 100 μg
- Concentration
- 0.5 mg/mL
- Storage
- -20°C
Submitted references HIV-1 promotes ubiquitination of the amyloidogenic C-terminal fragment of APP to support viral replication.
PLD3 affects axonal spheroids and network defects in Alzheimer's disease.
Analysis of the intracellular traffic of IgG in the context of Down syndrome (trisomy 21).
Overexpression of Kinesin Superfamily Motor Proteins in Alzheimer's Disease.
HIV-1 counteracts an innate restriction by amyloid precursor protein resulting in neurodegeneration.
Disruption of endolysosomal trafficking pathways in glioma cells by methuosis-inducing indole-based chalcones.
Axonal motor protein KIF5A and associated cargo deficits in multiple sclerosis lesional and normal-appearing white matter.
TREM2 Haplodeficiency in Mice and Humans Impairs the Microglia Barrier Function Leading to Decreased Amyloid Compaction and Severe Axonal Dystrophy.
Attenuation of β-Amyloid Deposition and Neurotoxicity by Chemogenetic Modulation of Neural Activity.
Sodium Chloride Increases Aβ Levels by Suppressing Aβ Clearance in Cultured Cells.
At the centre of neuronal, synaptic and axonal pathology in murine prion disease: degeneration of neuroanatomically linked thalamic and brainstem nuclei.
TRPC6 specifically interacts with APP to inhibit its cleavage by γ-secretase and reduce Aβ production.
Human neural stem cells alleviate Alzheimer-like pathology in a mouse model.
Increasing Membrane Cholesterol Level Increases the Amyloidogenic Peptide by Enhancing the Expression of Phospholipase C.
Neuroprotective maraviroc monotherapy in simian immunodeficiency virus-infected macaques: reduced replicating and latent SIV in the brain.
Modulation of lipid kinase PI4KIIα activity and lipid raft association of presenilin 1 underlies γ-secretase inhibition by ginsenoside (20S)-Rg3.
Involvement of receptor tyrosine kinase Tyro3 in amyloidogenic APP processing and β-amyloid deposition in Alzheimer's disease models.
Kinesin light chain 1 suppression impairs human embryonic stem cell neural differentiation and amyloid precursor protein metabolism.
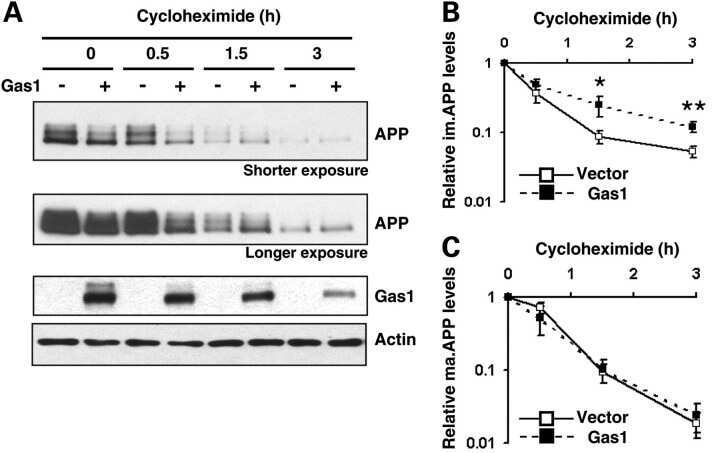
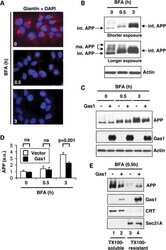
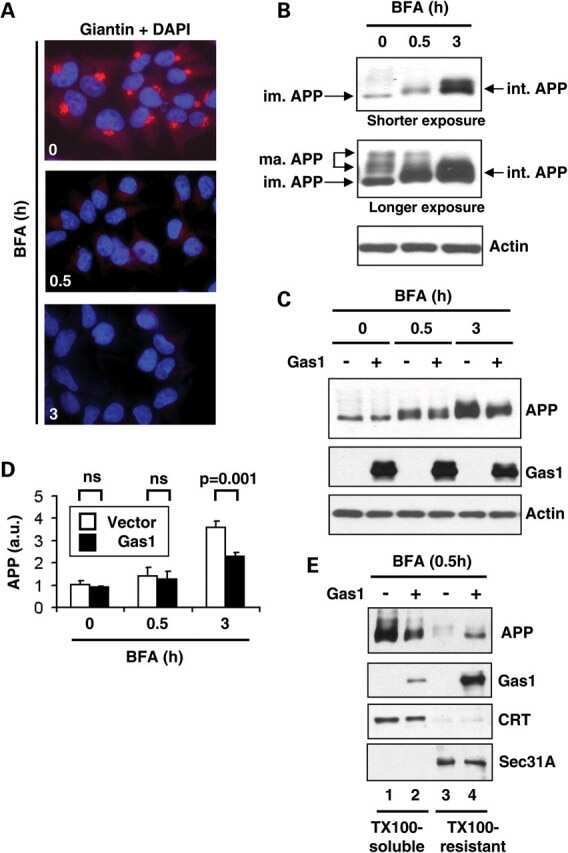
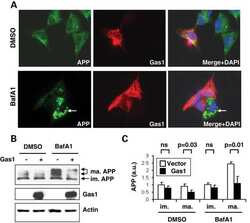
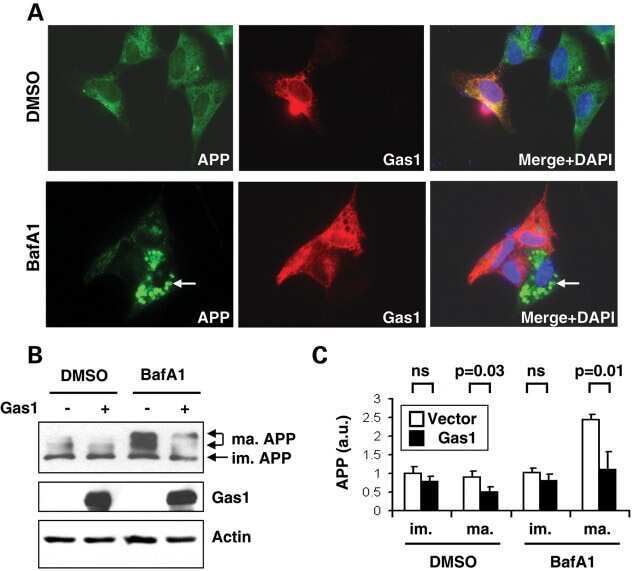
Gas1 interferes with AβPP trafficking by facilitating the accumulation of immature AβPP in endoplasmic reticulum-associated raft subdomains.
Intracellular distribution of amyloid beta peptide and its relationship to the lysosomal system.
Hyperpolarization-activated cyclic nucleotide gated channels: a potential molecular link between epileptic seizures and Aβ generation in Alzheimer's disease.
Phagocytosis of neuronal debris by microglia is associated with neuronal damage in multiple sclerosis.
Growth arrest-specific 1 binds to and controls the maturation and processing of the amyloid-beta precursor protein.
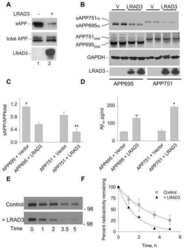
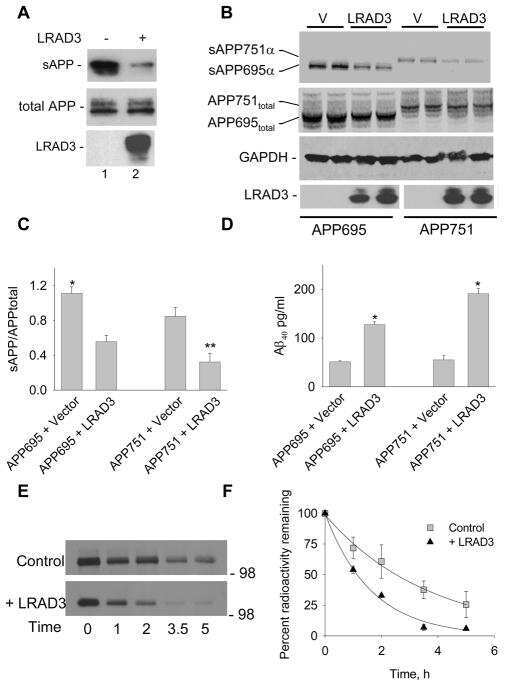
LRAD3, a novel low-density lipoprotein receptor family member that modulates amyloid precursor protein trafficking.
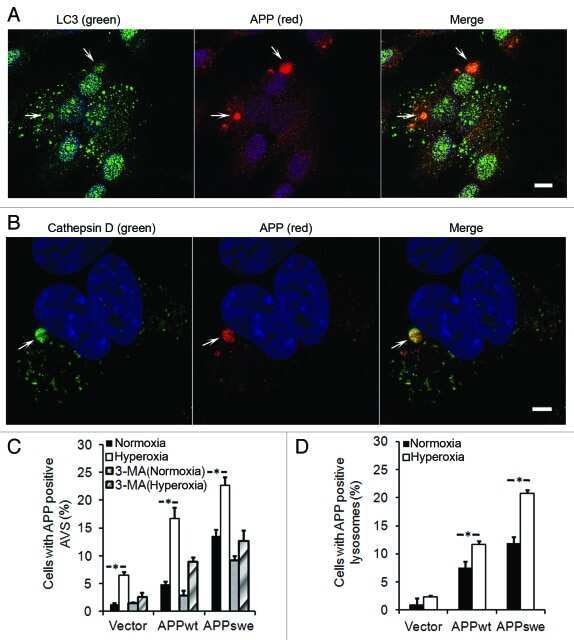
Macroautophagy-generated increase of lysosomal amyloid β-protein mediates oxidant-induced apoptosis of cultured neuroblastoma cells.
Macroautophagy-generated increase of lysosomal amyloid β-protein mediates oxidant-induced apoptosis of cultured neuroblastoma cells.
AMP-activated protein kinase signaling activation by resveratrol modulates amyloid-beta peptide metabolism.
Identification of novel N-terminal fragments of amyloid precursor protein in cerebrospinal fluid.
Alphab-crystallin is a target for adaptive immune responses and a trigger of innate responses in preactive multiple sclerosis lesions.
Early-onset and robust amyloid pathology in a new homozygous mouse model of Alzheimer's disease.
Natural host genetic resistance to lentiviral CNS disease: a neuroprotective MHC class I allele in SIV-infected macaques.
Autosomal dominant sensory ataxia: a neuroaxonal dystrophy.
Dual-specificity tyrosine(Y)-phosphorylation regulated kinase 1A-mediated phosphorylation of amyloid precursor protein: evidence for a functional link between Down syndrome and Alzheimer's disease.
CNI-1493 inhibits Abeta production, plaque formation, and cognitive deterioration in an animal model of Alzheimer's disease.
Amyloid precursor protein and Presenilin1 interact with the adaptor GRB2 and modulate ERK 1,2 signaling.
Calpain inhibitor MDL-28170 reduces the functional and structural deterioration of corpus callosum following fluid percussion injury.
Expression of APP pathway mRNAs and proteins in Alzheimer's disease.
A preliminary neuropathological study of Japanese encephalitis in humans and a mouse model.
L-glutamate and phorbol ester stimulate the release of secretory amyloid precursor protein from rat cortical synaptosomes.
14-3-3 protein in CSF: an early predictor of SIV CNS disease.
Fatal familial insomnia with an unusual prion protein deposition pattern: an autopsy report with an experimental transmission study.
Contributors to white matter damage in the frontal lobe in Alzheimer's disease.
Decreased levels of soluble amyloid beta-protein precursor and beta-amyloid protein in cerebrospinal fluid of patients with systemic lupus erythematosus.
Progressive cognitive impairment and anxiety induction in the absence of plaque deposition in C57BL/6 inbred mice expressing transgenic amyloid precursor protein.
Fe65 is not involved in the platelet-derived growth factor-induced processing of Alzheimer's amyloid precursor protein, which activates its caspase-directed cleavage.
Ultrastructural and behavioural changes precede amyloid deposition in a transgenic model of Alzheimer's disease.
Impact mechanics and axonal injury in a sheep model.
Novel cadherin-related membrane proteins, Alcadeins, enhance the X11-like protein-mediated stabilization of amyloid beta-protein precursor metabolism.
Central nervous system correlates of behavioral deficits following simian immunodeficiency virus infection.
Disruption of corticocortical connections ameliorates amyloid burden in terminal fields in a transgenic model of Abeta amyloidosis.
Accumulation of beta-amyloid precursor protein in axons correlates with CNS expression of SIV gp41.
The protease inhibitor, MG132, blocks maturation of the amyloid precursor protein Swedish mutant preventing cleavage by beta-Secretase.
Endosomal sorting of amyloid precursor protein-P-selectin chimeras influences secretase processing.
Human bleomycin hydrolase regulates the secretion of amyloid precursor protein.
Sporadic inclusion body myositis correlates with increased expression and cross-linking by transglutaminases 1 and 2.
Sporadic inclusion body myositis correlates with increased expression and cross-linking by transglutaminases 1 and 2.
Interaction of a neuron-specific protein containing PDZ domains with Alzheimer's amyloid precursor protein.
Neurofibrillary lesions in experimental aluminum-induced encephalopathy and Alzheimer's disease share immunoreactivity for amyloid precursor protein, A beta, alpha 1-antichymotrypsin and ubiquitin-protein conjugates.
Neurofibrillary lesions in experimental aluminum-induced encephalopathy and Alzheimer's disease share immunoreactivity for amyloid precursor protein, A beta, alpha 1-antichymotrypsin and ubiquitin-protein conjugates.
A novel protein, amyloid precursor-like protein 2, is present in human brain, cerebrospinal fluid and conditioned media.
Markers of axonal injury in post mortem human brain.
Gu F, Boisjoli M, Naghavi MH
Nature communications 2023 Jul 15;14(1):4227
Nature communications 2023 Jul 15;14(1):4227
PLD3 affects axonal spheroids and network defects in Alzheimer's disease.
Yuan P, Zhang M, Tong L, Morse TM, McDougal RA, Ding H, Chan D, Cai Y, Grutzendler J
Nature 2022 Dec;612(7939):328-337
Nature 2022 Dec;612(7939):328-337
Analysis of the intracellular traffic of IgG in the context of Down syndrome (trisomy 21).
Cejas RB, Tamaño-Blanco M, Blanco JG
Scientific reports 2021 May 26;11(1):10981
Scientific reports 2021 May 26;11(1):10981
Overexpression of Kinesin Superfamily Motor Proteins in Alzheimer's Disease.
Hares K, Miners JS, Cook AJ, Rice C, Scolding N, Love S, Wilkins A
Journal of Alzheimer's disease : JAD 2017;60(4):1511-1524
Journal of Alzheimer's disease : JAD 2017;60(4):1511-1524
HIV-1 counteracts an innate restriction by amyloid precursor protein resulting in neurodegeneration.
Chai Q, Jovasevic V, Malikov V, Sabo Y, Morham S, Walsh D, Naghavi MH
Nature communications 2017 Nov 15;8(1):1522
Nature communications 2017 Nov 15;8(1):1522
Disruption of endolysosomal trafficking pathways in glioma cells by methuosis-inducing indole-based chalcones.
Mbah NE, Overmeyer JH, Maltese WA
Cell biology and toxicology 2017 Jun;33(3):263-282
Cell biology and toxicology 2017 Jun;33(3):263-282
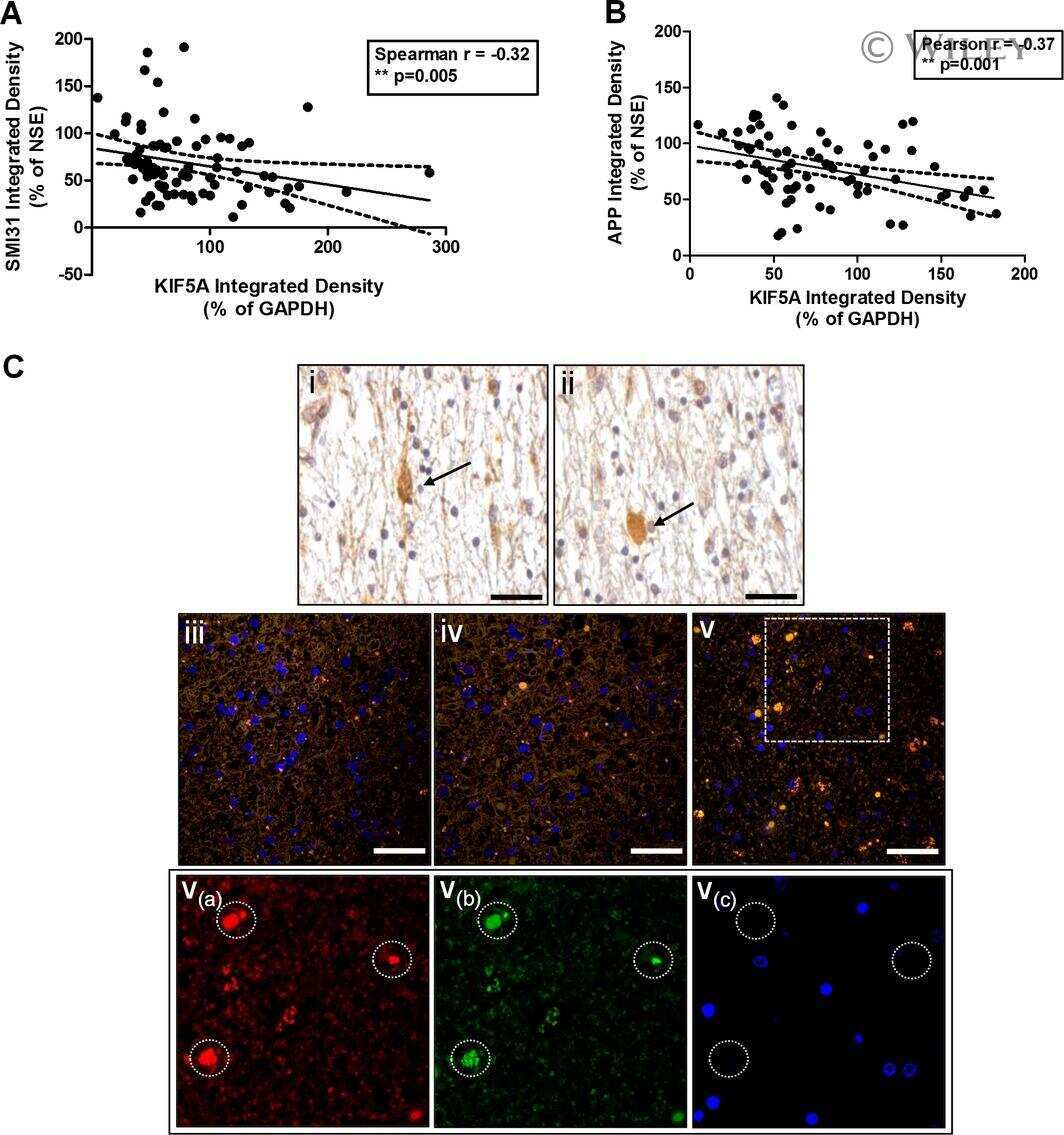
Axonal motor protein KIF5A and associated cargo deficits in multiple sclerosis lesional and normal-appearing white matter.
Hares K, Redondo J, Kemp K, Rice C, Scolding N, Wilkins A
Neuropathology and applied neurobiology 2017 Apr;43(3):227-241
Neuropathology and applied neurobiology 2017 Apr;43(3):227-241
TREM2 Haplodeficiency in Mice and Humans Impairs the Microglia Barrier Function Leading to Decreased Amyloid Compaction and Severe Axonal Dystrophy.
Yuan P, Condello C, Keene CD, Wang Y, Bird TD, Paul SM, Luo W, Colonna M, Baddeley D, Grutzendler J
Neuron 2016 May 18;90(4):724-39
Neuron 2016 May 18;90(4):724-39
Attenuation of β-Amyloid Deposition and Neurotoxicity by Chemogenetic Modulation of Neural Activity.
Yuan P, Grutzendler J
The Journal of neuroscience : the official journal of the Society for Neuroscience 2016 Jan 13;36(2):632-41
The Journal of neuroscience : the official journal of the Society for Neuroscience 2016 Jan 13;36(2):632-41
Sodium Chloride Increases Aβ Levels by Suppressing Aβ Clearance in Cultured Cells.
Cheng XJ, Gao Y, Zhao YW, Cheng XD
PloS one 2015;10(6):e0130432
PloS one 2015;10(6):e0130432
At the centre of neuronal, synaptic and axonal pathology in murine prion disease: degeneration of neuroanatomically linked thalamic and brainstem nuclei.
Reis R, Hennessy E, Murray C, Griffin ÉW, Cunningham C
Neuropathology and applied neurobiology 2015 Oct;41(6):780-97
Neuropathology and applied neurobiology 2015 Oct;41(6):780-97
TRPC6 specifically interacts with APP to inhibit its cleavage by γ-secretase and reduce Aβ production.
Wang J, Lu R, Yang J, Li H, He Z, Jing N, Wang X, Wang Y
Nature communications 2015 Nov 19;6:8876
Nature communications 2015 Nov 19;6:8876
Human neural stem cells alleviate Alzheimer-like pathology in a mouse model.
Lee IS, Jung K, Kim IS, Lee H, Kim M, Yun S, Hwang K, Shin JE, Park KI
Molecular neurodegeneration 2015 Aug 21;10:38
Molecular neurodegeneration 2015 Aug 21;10:38
Increasing Membrane Cholesterol Level Increases the Amyloidogenic Peptide by Enhancing the Expression of Phospholipase C.
Chun YS, Oh HG, Park MK, Kim TW, Chung S
Journal of neurodegenerative diseases 2013;2013:407903
Journal of neurodegenerative diseases 2013;2013:407903
Neuroprotective maraviroc monotherapy in simian immunodeficiency virus-infected macaques: reduced replicating and latent SIV in the brain.
Kelly KM, Beck SE, Metcalf Pate KA, Queen SE, Dorsey JL, Adams RJ, Avery LB, Hubbard W, Tarwater PM, Mankowski JL
AIDS (London, England) 2013 Nov 28;27(18):F21-8
AIDS (London, England) 2013 Nov 28;27(18):F21-8
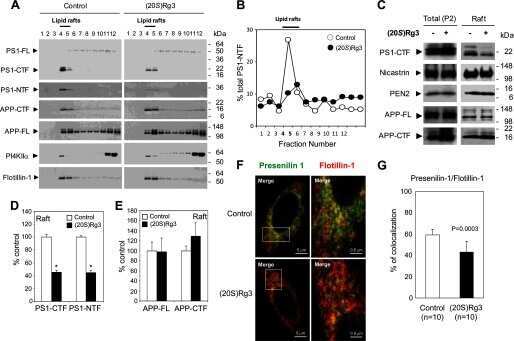
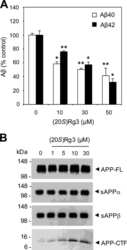
Modulation of lipid kinase PI4KIIα activity and lipid raft association of presenilin 1 underlies γ-secretase inhibition by ginsenoside (20S)-Rg3.
Kang MS, Baek SH, Chun YS, Moore AZ, Landman N, Berman D, Yang HO, Morishima-Kawashima M, Osawa S, Funamoto S, Ihara Y, Di Paolo G, Park JH, Chung S, Kim TW
The Journal of biological chemistry 2013 Jul 19;288(29):20868-20882
The Journal of biological chemistry 2013 Jul 19;288(29):20868-20882
Involvement of receptor tyrosine kinase Tyro3 in amyloidogenic APP processing and β-amyloid deposition in Alzheimer's disease models.
Zheng Y, Wang Q, Xiao B, Lu Q, Wang Y, Wang X
PloS one 2012;7(6):e39035
PloS one 2012;7(6):e39035
Kinesin light chain 1 suppression impairs human embryonic stem cell neural differentiation and amyloid precursor protein metabolism.
Killian RL, Flippin JD, Herrera CM, Almenar-Queralt A, Goldstein LS
PloS one 2012;7(1):e29755
PloS one 2012;7(1):e29755
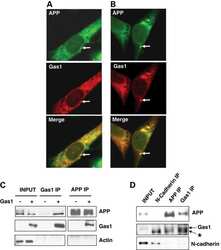
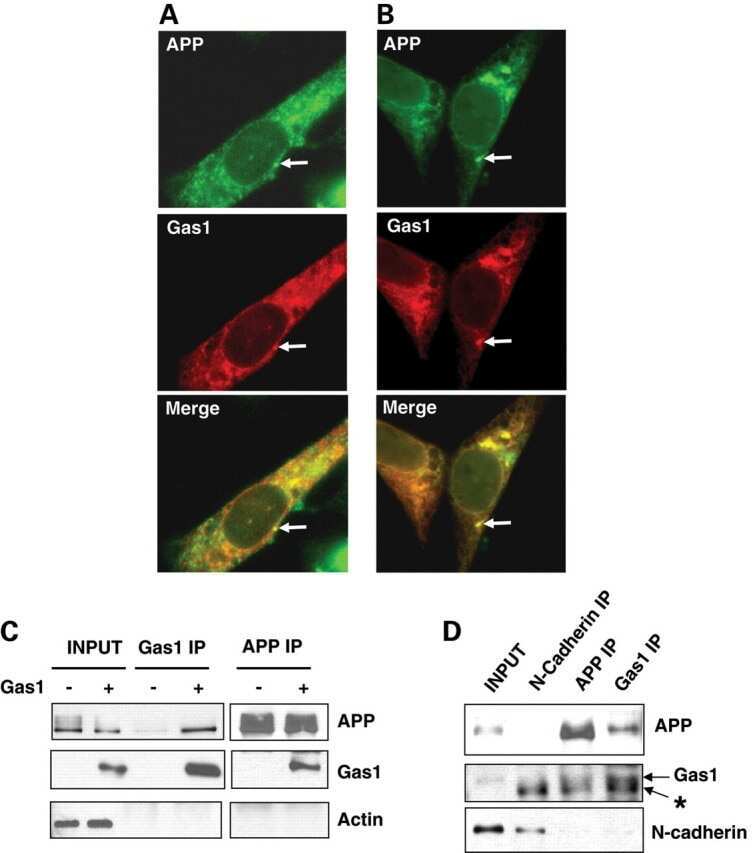
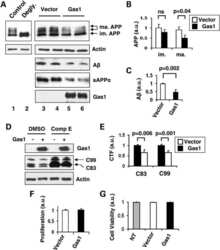
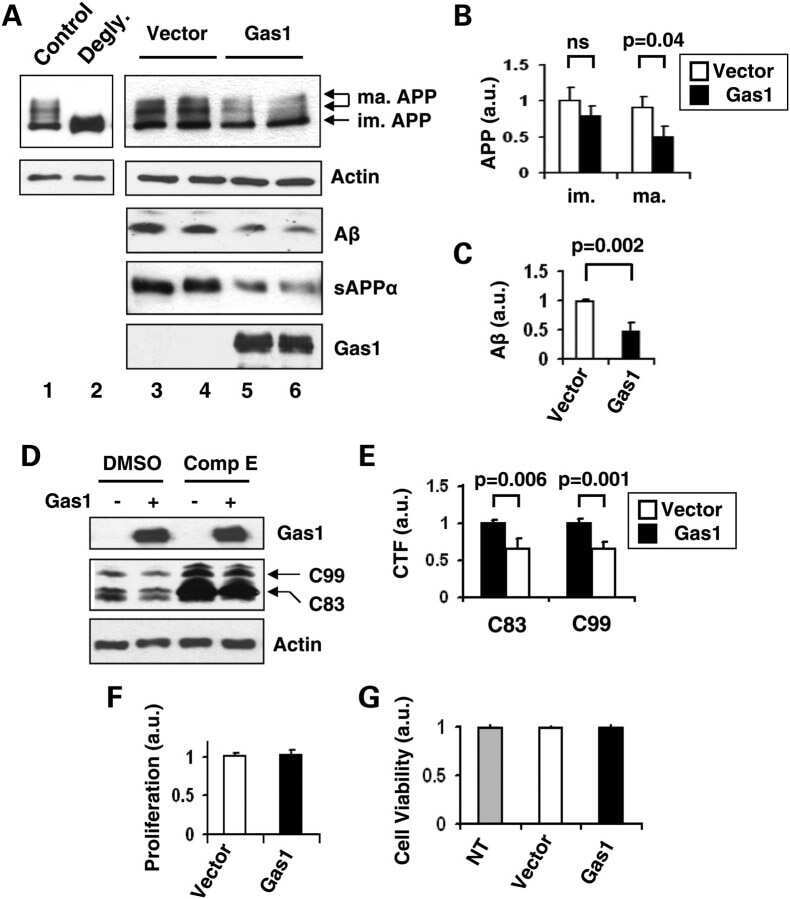
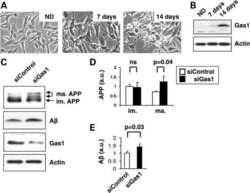
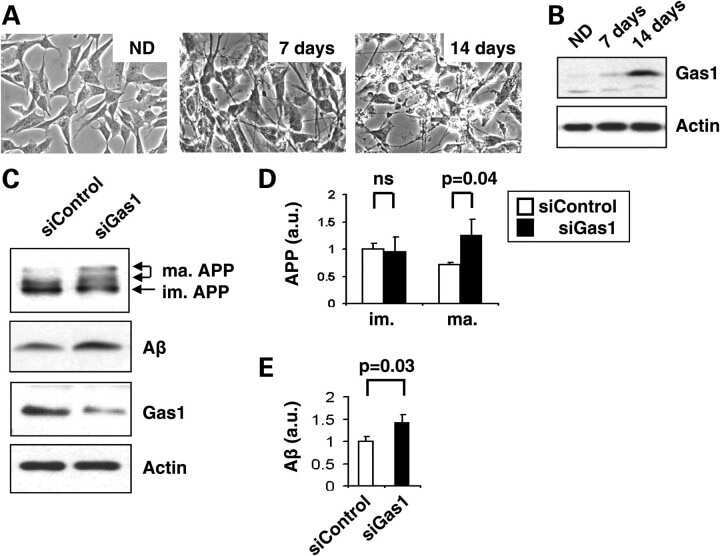
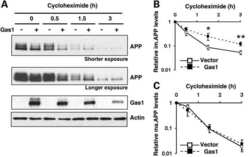
Gas1 interferes with AβPP trafficking by facilitating the accumulation of immature AβPP in endoplasmic reticulum-associated raft subdomains.
Chapuis J, Vingtdeux V, Capiralla H, Davies P, Marambaud P
Journal of Alzheimer's disease : JAD 2012;28(1):127-35
Journal of Alzheimer's disease : JAD 2012;28(1):127-35
Intracellular distribution of amyloid beta peptide and its relationship to the lysosomal system.
Zheng L, Cedazo-Minguez A, Hallbeck M, Jerhammar F, Marcusson J, Terman A
Translational neurodegeneration 2012 Sep 26;1(1):19
Translational neurodegeneration 2012 Sep 26;1(1):19
Hyperpolarization-activated cyclic nucleotide gated channels: a potential molecular link between epileptic seizures and Aβ generation in Alzheimer's disease.
Saito Y, Inoue T, Zhu G, Kimura N, Okada M, Nishimura M, Kimura N, Murayama S, Kaneko S, Shigemoto R, Imoto K, Suzuki T
Molecular neurodegeneration 2012 Oct 3;7:50
Molecular neurodegeneration 2012 Oct 3;7:50
Phagocytosis of neuronal debris by microglia is associated with neuronal damage in multiple sclerosis.
Huizinga R, van der Star BJ, Kipp M, Jong R, Gerritsen W, Clarner T, Puentes F, Dijkstra CD, van der Valk P, Amor S
Glia 2012 Mar;60(3):422-31
Glia 2012 Mar;60(3):422-31
Growth arrest-specific 1 binds to and controls the maturation and processing of the amyloid-beta precursor protein.
Chapuis J, Vingtdeux V, Campagne F, Davies P, Marambaud P
Human molecular genetics 2011 May 15;20(10):2026-36
Human molecular genetics 2011 May 15;20(10):2026-36
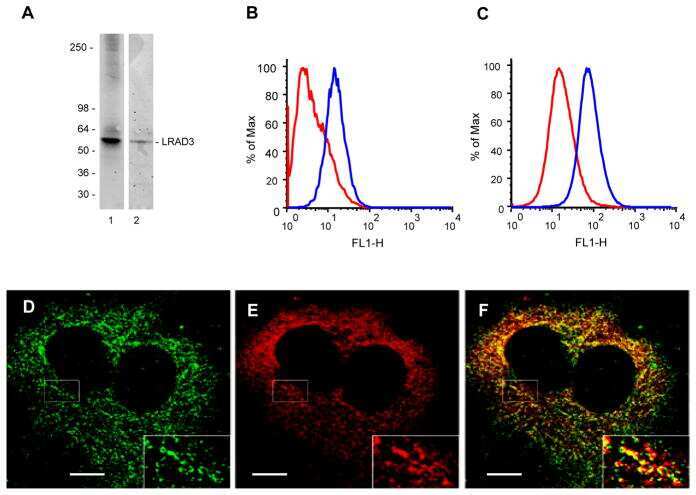
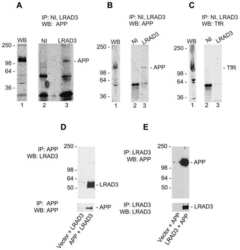
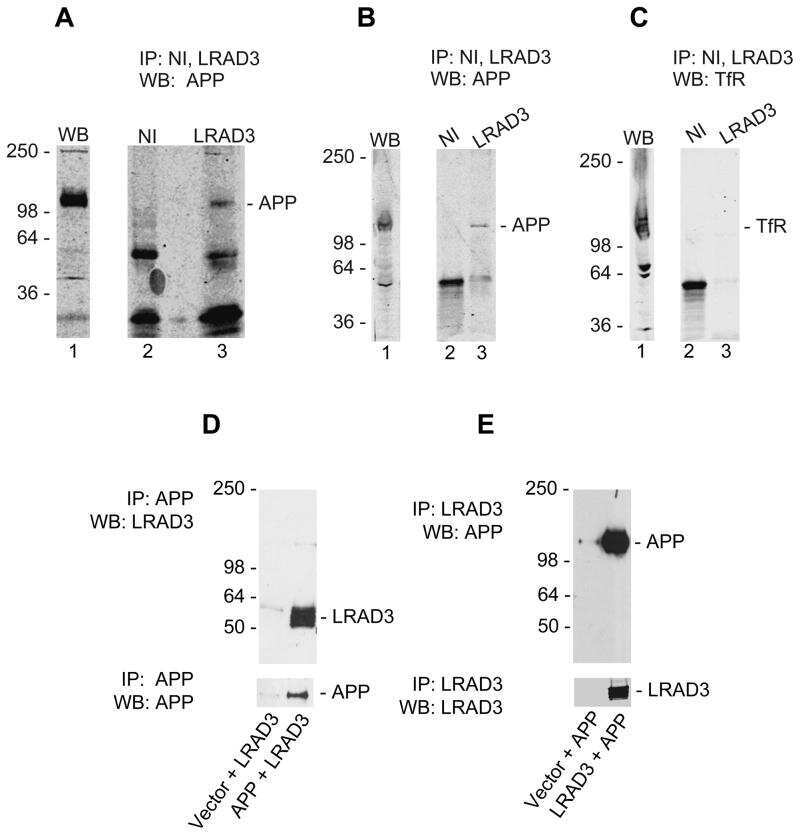
LRAD3, a novel low-density lipoprotein receptor family member that modulates amyloid precursor protein trafficking.
Ranganathan S, Noyes NC, Migliorini M, Winkles JA, Battey FD, Hyman BT, Smith E, Yepes M, Mikhailenko I, Strickland DK
The Journal of neuroscience : the official journal of the Society for Neuroscience 2011 Jul 27;31(30):10836-46
The Journal of neuroscience : the official journal of the Society for Neuroscience 2011 Jul 27;31(30):10836-46
Macroautophagy-generated increase of lysosomal amyloid β-protein mediates oxidant-induced apoptosis of cultured neuroblastoma cells.
Zheng L, Terman A, Hallbeck M, Dehvari N, Cowburn RF, Benedikz E, Kågedal K, Cedazo-Minguez A, Marcusson J
Autophagy 2011 Dec;7(12):1528-45
Autophagy 2011 Dec;7(12):1528-45
Macroautophagy-generated increase of lysosomal amyloid β-protein mediates oxidant-induced apoptosis of cultured neuroblastoma cells.
Zheng L, Terman A, Hallbeck M, Dehvari N, Cowburn RF, Benedikz E, Kågedal K, Cedazo-Minguez A, Marcusson J
Autophagy 2011 Dec;7(12):1528-45
Autophagy 2011 Dec;7(12):1528-45
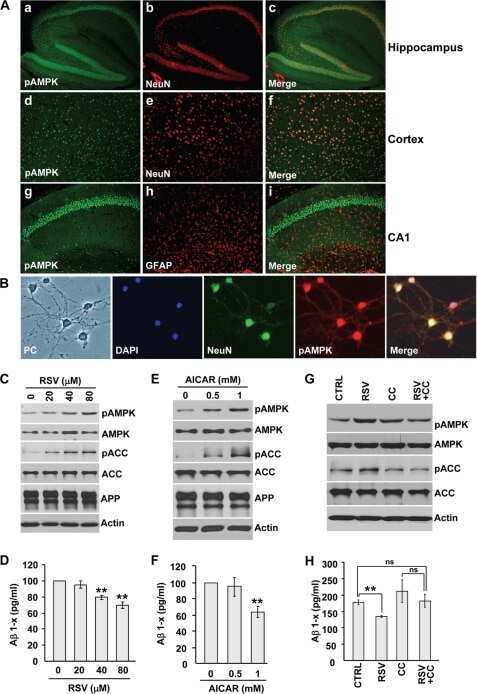
AMP-activated protein kinase signaling activation by resveratrol modulates amyloid-beta peptide metabolism.
Vingtdeux V, Giliberto L, Zhao H, Chandakkar P, Wu Q, Simon JE, Janle EM, Lobo J, Ferruzzi MG, Davies P, Marambaud P
The Journal of biological chemistry 2010 Mar 19;285(12):9100-13
The Journal of biological chemistry 2010 Mar 19;285(12):9100-13
Identification of novel N-terminal fragments of amyloid precursor protein in cerebrospinal fluid.
Portelius E, Brinkmalm G, Tran A, Andreasson U, Zetterberg H, Westman-Brinkmalm A, Blennow K, Ohrfelt A
Experimental neurology 2010 Jun;223(2):351-8
Experimental neurology 2010 Jun;223(2):351-8
Alphab-crystallin is a target for adaptive immune responses and a trigger of innate responses in preactive multiple sclerosis lesions.
van Noort JM, Bsibsi M, Gerritsen WH, van der Valk P, Bajramovic JJ, Steinman L, Amor S
Journal of neuropathology and experimental neurology 2010 Jul;69(7):694-703
Journal of neuropathology and experimental neurology 2010 Jul;69(7):694-703
Early-onset and robust amyloid pathology in a new homozygous mouse model of Alzheimer's disease.
Willuweit A, Velden J, Godemann R, Manook A, Jetzek F, Tintrup H, Kauselmann G, Zevnik B, Henriksen G, Drzezga A, Pohlner J, Schoor M, Kemp JA, von der Kammer H
PloS one 2009 Nov 20;4(11):e7931
PloS one 2009 Nov 20;4(11):e7931
Natural host genetic resistance to lentiviral CNS disease: a neuroprotective MHC class I allele in SIV-infected macaques.
Mankowski JL, Queen SE, Fernandez CS, Tarwater PM, Karper JM, Adams RJ, Kent SJ
PloS one 2008;3(11):e3603
PloS one 2008;3(11):e3603
Autosomal dominant sensory ataxia: a neuroaxonal dystrophy.
Moeller JJ, Macaulay RJ, Valdmanis PN, Weston LE, Rouleau GA, Dupré N
Acta neuropathologica 2008 Sep;116(3):331-6
Acta neuropathologica 2008 Sep;116(3):331-6
Dual-specificity tyrosine(Y)-phosphorylation regulated kinase 1A-mediated phosphorylation of amyloid precursor protein: evidence for a functional link between Down syndrome and Alzheimer's disease.
Ryoo SR, Cho HJ, Lee HW, Jeong HK, Radnaabazar C, Kim YS, Kim MJ, Son MY, Seo H, Chung SH, Song WJ
Journal of neurochemistry 2008 Mar;104(5):1333-44
Journal of neurochemistry 2008 Mar;104(5):1333-44
CNI-1493 inhibits Abeta production, plaque formation, and cognitive deterioration in an animal model of Alzheimer's disease.
Bacher M, Dodel R, Aljabari B, Keyvani K, Marambaud P, Kayed R, Glabe C, Goertz N, Hoppmann A, Sachser N, Klotsche J, Schnell S, Lewejohann L, Al-Abed Y
The Journal of experimental medicine 2008 Jul 7;205(7):1593-9
The Journal of experimental medicine 2008 Jul 7;205(7):1593-9
Amyloid precursor protein and Presenilin1 interact with the adaptor GRB2 and modulate ERK 1,2 signaling.
Nizzari M, Venezia V, Repetto E, Caorsi V, Magrassi R, Gagliani MC, Carlo P, Florio T, Schettini G, Tacchetti C, Russo T, Diaspro A, Russo C
The Journal of biological chemistry 2007 May 4;282(18):13833-44
The Journal of biological chemistry 2007 May 4;282(18):13833-44
Calpain inhibitor MDL-28170 reduces the functional and structural deterioration of corpus callosum following fluid percussion injury.
Ai J, Liu E, Wang J, Chen Y, Yu J, Baker AJ
Journal of neurotrauma 2007 Jun;24(6):960-78
Journal of neurotrauma 2007 Jun;24(6):960-78
Expression of APP pathway mRNAs and proteins in Alzheimer's disease.
Matsui T, Ingelsson M, Fukumoto H, Ramasamy K, Kowa H, Frosch MP, Irizarry MC, Hyman BT
Brain research 2007 Aug 3;1161:116-23
Brain research 2007 Aug 3;1161:116-23
A preliminary neuropathological study of Japanese encephalitis in humans and a mouse model.
German AC, Myint KS, Mai NT, Pomeroy I, Phu NH, Tzartos J, Winter P, Collett J, Farrar J, Barrett A, Kipar A, Esiri MM, Solomon T
Transactions of the Royal Society of Tropical Medicine and Hygiene 2006 Dec;100(12):1135-45
Transactions of the Royal Society of Tropical Medicine and Hygiene 2006 Dec;100(12):1135-45
L-glutamate and phorbol ester stimulate the release of secretory amyloid precursor protein from rat cortical synaptosomes.
Kirazov L, Kirazovi E, Schliebs R
Acta biologica Hungarica 2005;56(3-4):177-83
Acta biologica Hungarica 2005;56(3-4):177-83
14-3-3 protein in CSF: an early predictor of SIV CNS disease.
Helke KL, Queen SE, Tarwater PM, Turchan-Cholewo J, Nath A, Zink MC, Irani DN, Mankowski JL
Journal of neuropathology and experimental neurology 2005 Mar;64(3):202-8
Journal of neuropathology and experimental neurology 2005 Mar;64(3):202-8
Fatal familial insomnia with an unusual prion protein deposition pattern: an autopsy report with an experimental transmission study.
Sasaki K, Doh-ura K, Wakisaka Y, Tomoda H, Iwaki T
Neuropathology and applied neurobiology 2005 Feb;31(1):80-7
Neuropathology and applied neurobiology 2005 Feb;31(1):80-7
Contributors to white matter damage in the frontal lobe in Alzheimer's disease.
Chalmers K, Wilcock G, Love S
Neuropathology and applied neurobiology 2005 Dec;31(6):623-31
Neuropathology and applied neurobiology 2005 Dec;31(6):623-31
Decreased levels of soluble amyloid beta-protein precursor and beta-amyloid protein in cerebrospinal fluid of patients with systemic lupus erythematosus.
Trysberg E, Höglund K, Svenungsson E, Blennow K, Tarkowski A
Arthritis research & therapy 2004;6(2):R129-36
Arthritis research & therapy 2004;6(2):R129-36
Progressive cognitive impairment and anxiety induction in the absence of plaque deposition in C57BL/6 inbred mice expressing transgenic amyloid precursor protein.
Lee KW, Lee SH, Kim H, Song JS, Yang SD, Paik SG, Han PL
Journal of neuroscience research 2004 May 15;76(4):572-80
Journal of neuroscience research 2004 May 15;76(4):572-80
Fe65 is not involved in the platelet-derived growth factor-induced processing of Alzheimer's amyloid precursor protein, which activates its caspase-directed cleavage.
Zambrano N, Gianni D, Bruni P, Passaro F, Telese F, Russo T
The Journal of biological chemistry 2004 Apr 16;279(16):16161-9
The Journal of biological chemistry 2004 Apr 16;279(16):16161-9
Ultrastructural and behavioural changes precede amyloid deposition in a transgenic model of Alzheimer's disease.
Richardson JC, Kendal CE, Anderson R, Priest F, Gower E, Soden P, Gray R, Topps S, Howlett DR, Lavender D, Clarke NJ, Barnes JC, Haworth R, Stewart MG, Rupniak HT
Neuroscience 2003;122(1):213-28
Neuroscience 2003;122(1):213-28
Impact mechanics and axonal injury in a sheep model.
Anderson RW, Brown CJ, Blumbergs PC, McLean AJ, Jones NR
Journal of neurotrauma 2003 Oct;20(10):961-74
Journal of neurotrauma 2003 Oct;20(10):961-74
Novel cadherin-related membrane proteins, Alcadeins, enhance the X11-like protein-mediated stabilization of amyloid beta-protein precursor metabolism.
Araki Y, Tomita S, Yamaguchi H, Miyagi N, Sumioka A, Kirino Y, Suzuki T
The Journal of biological chemistry 2003 Dec 5;278(49):49448-58
The Journal of biological chemistry 2003 Dec 5;278(49):49448-58
Central nervous system correlates of behavioral deficits following simian immunodeficiency virus infection.
Weed MR, Hienz RD, Brady JV, Adams RJ, Mankowski JL, Clements JE, Zink MC
Journal of neurovirology 2003 Aug;9(4):452-64
Journal of neurovirology 2003 Aug;9(4):452-64
Disruption of corticocortical connections ameliorates amyloid burden in terminal fields in a transgenic model of Abeta amyloidosis.
Sheng JG, Price DL, Koliatsos VE
The Journal of neuroscience : the official journal of the Society for Neuroscience 2002 Nov 15;22(22):9794-9
The Journal of neuroscience : the official journal of the Society for Neuroscience 2002 Nov 15;22(22):9794-9
Accumulation of beta-amyloid precursor protein in axons correlates with CNS expression of SIV gp41.
Mankowski JL, Queen SE, Tarwater PM, Fox KJ, Perry VH
Journal of neuropathology and experimental neurology 2002 Jan;61(1):85-90
Journal of neuropathology and experimental neurology 2002 Jan;61(1):85-90
The protease inhibitor, MG132, blocks maturation of the amyloid precursor protein Swedish mutant preventing cleavage by beta-Secretase.
Steinhilb ML, Turner RS, Gaut JR
The Journal of biological chemistry 2001 Feb 9;276(6):4476-84
The Journal of biological chemistry 2001 Feb 9;276(6):4476-84
Endosomal sorting of amyloid precursor protein-P-selectin chimeras influences secretase processing.
Daugherty BL, Green SA
Traffic (Copenhagen, Denmark) 2001 Dec;2(12):908-16
Traffic (Copenhagen, Denmark) 2001 Dec;2(12):908-16
Human bleomycin hydrolase regulates the secretion of amyloid precursor protein.
Lefterov IM, Koldamova RP, Lazo JS
FASEB journal : official publication of the Federation of American Societies for Experimental Biology 2000 Sep;14(12):1837-47
FASEB journal : official publication of the Federation of American Societies for Experimental Biology 2000 Sep;14(12):1837-47
Sporadic inclusion body myositis correlates with increased expression and cross-linking by transglutaminases 1 and 2.
Choi YC, Park GT, Kim TS, Sunwoo IN, Steinert PM, Kim SY
The Journal of biological chemistry 2000 Mar 24;275(12):8703-10
The Journal of biological chemistry 2000 Mar 24;275(12):8703-10
Sporadic inclusion body myositis correlates with increased expression and cross-linking by transglutaminases 1 and 2.
Choi YC, Park GT, Kim TS, Sunwoo IN, Steinert PM, Kim SY
The Journal of biological chemistry 2000 Mar 24;275(12):8703-10
The Journal of biological chemistry 2000 Mar 24;275(12):8703-10
Interaction of a neuron-specific protein containing PDZ domains with Alzheimer's amyloid precursor protein.
Tomita S, Ozaki T, Taru H, Oguchi S, Takeda S, Yagi Y, Sakiyama S, Kirino Y, Suzuki T
The Journal of biological chemistry 1999 Jan 22;274(4):2243-54
The Journal of biological chemistry 1999 Jan 22;274(4):2243-54
Neurofibrillary lesions in experimental aluminum-induced encephalopathy and Alzheimer's disease share immunoreactivity for amyloid precursor protein, A beta, alpha 1-antichymotrypsin and ubiquitin-protein conjugates.
Huang Y, Herman MM, Liu J, Katsetos CD, Wills MR, Savory J
Brain research 1997 Oct 17;771(2):213-20
Brain research 1997 Oct 17;771(2):213-20
Neurofibrillary lesions in experimental aluminum-induced encephalopathy and Alzheimer's disease share immunoreactivity for amyloid precursor protein, A beta, alpha 1-antichymotrypsin and ubiquitin-protein conjugates.
Huang Y, Herman MM, Liu J, Katsetos CD, Wills MR, Savory J
Brain research 1997 Oct 17;771(2):213-20
Brain research 1997 Oct 17;771(2):213-20
A novel protein, amyloid precursor-like protein 2, is present in human brain, cerebrospinal fluid and conditioned media.
Webster MT, Groome N, Francis PT, Pearce BR, Sherriff FE, Thinakaran G, Felsenstein KM, Wasco W, Tanzi RE, Bowen DM
The Biochemical journal 1995 Aug 15;310 ( Pt 1)(Pt 1):95-9
The Biochemical journal 1995 Aug 15;310 ( Pt 1)(Pt 1):95-9
Markers of axonal injury in post mortem human brain.
Sherriff FE, Bridges LR, Gentleman SM, Sivaloganathan S, Wilson S
Acta neuropathologica 1994;88(5):433-9
Acta neuropathologica 1994;88(5):433-9
No comments: Submit comment
Supportive validation
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
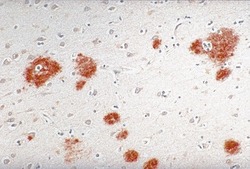
- Mouse anti-Beta-Amyloid Precursor Protein (LN27) stained Alzheimer's disease.
Supportive validation
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
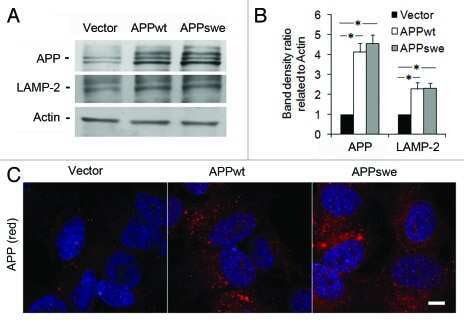
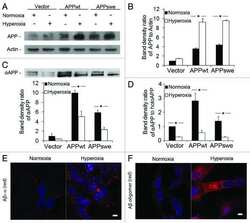
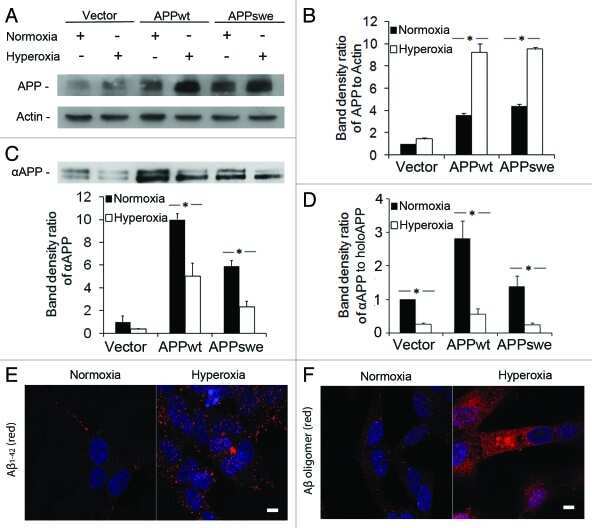
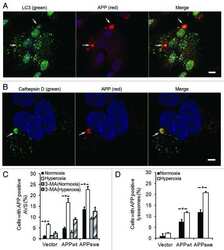
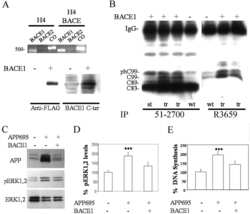
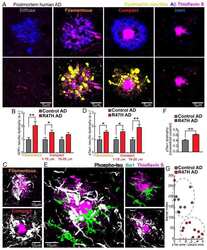
- Figure 1 SIV infection induced encephalitis, increased CNS CD68 expression, and axonal accumulation of APP. SIV-infected pig-tailed macaques that developed encephalitis had multifocal perivascular accumulations of infiltrating macrophages and multinucleate giant cells (A, arrows denote giant cells, * denotes blood vessel lumen, hematoxylin and eosin stain, bar = 50 uM), increased CNS CD68 expression reflecting activation of macrophages and microglia in subcortical white matter shown by immunohistochemical staining with an anti-CD68 antibody (B, bar = 50 uM), and accumulation of APP in axons (arrows) in the corpus callosum (C, bar = 100 uM).
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
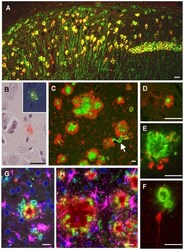
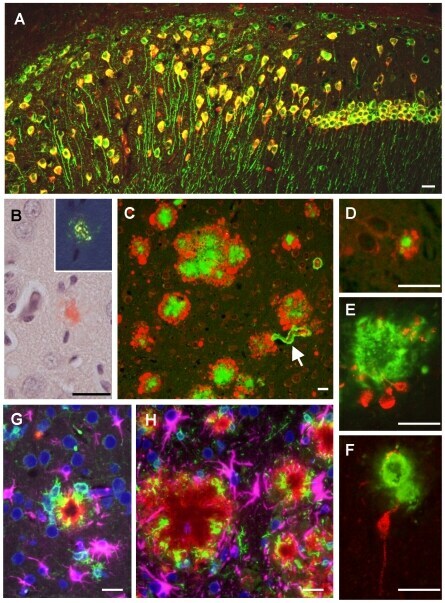
- Figure 1 Amyloid pathology and glial inflammation in ARTE10 mouse brain. A , Double-immunofluorescent colocalization of the transgenic human APP ( red , human-specific antibody against the N-terminal ectodomain) with the neuron-specific somato-dendritic marker MAP2 ( green ) in the subiculum and the CA1 region of the hippocampus. The transgene is amply expressed in most (>80%) neurons ( yellow merge colour), whereas non-neuronal cells show no signals beyond background. B , Congo-red stained plaque core ( red ) in the frontal cortex of a hemizygous ARTE10 mouse at 5 months of age. The green bi-refringence in polarized light (inset) is diagnostic of amyloid. C, D , Thioflavin-S positive plaque cores ( green ), each encircled by a sphere (corona) of dilated, strongly ubiquitin-positive dystrophic neurites ( red ) in the subiculum of a homozygous ARTE10 mouse at 3 months of age (D) and in the frontal cortex of a 19 months old hemizygous mouse (C), the latter with concomitant amyloid angiopathy (arrow). E, F , AT8-positive hyperphosphorylated tau ( red ) in dystrophic neurites adjacent to plaque cores ( green : Abeta40 and Abeta42) in homozygous ARTE10 mice at 12 (E) and 8 (F) months of age. G, H , Triple-immunofluorescent demonstration of plaque-associated mixed glial inflammation: Activated microglia ( green , AIF1/Iba1) and reactive astroglia ( magenta , GFAP) colonizing the periphery of amyloid cores ( red : Abeta/6E10) in the subicula of a homozygous 5 months (G) and a hemizy
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
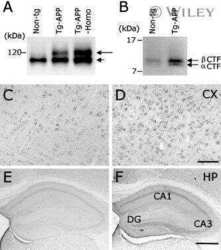
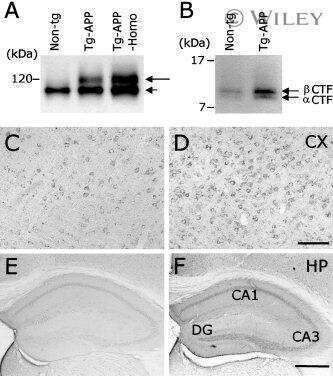
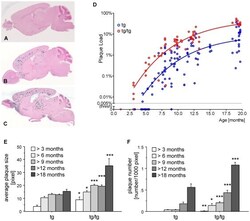
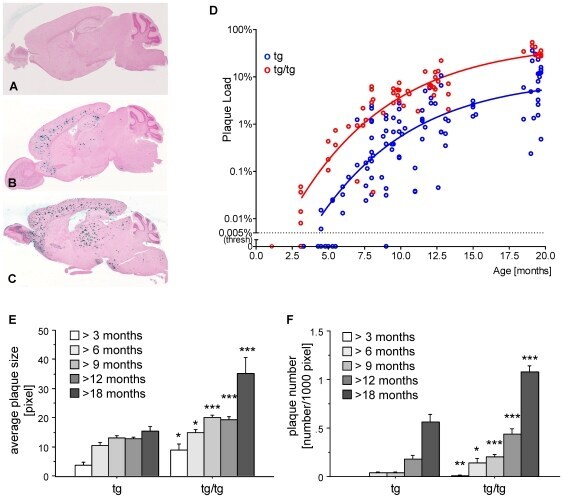
- Figure 2 Onset and development of plaques in ARTE10 mice. A-C , Digital micrographs of parasagittal forebrain sections from homozygous transgenic mice immunohistochemically stained for beta-amyloid plaques ( dark blue-green ) at 3 (A), 8 (B), and 13 months of age (C). D-F Plaques are quantified using Acapella(tm) plaque recognition software in regions of interest on stained mouse brain sections. D , Plaque load is expressed as percentage of plaque area per total area of neocortex and hippocampus across ages in homozygous ( red ) versus hemizygous mice ( blue ). Each symbol represents one animal. The average size (E) and number of plaques per area (F) progressively increase with age in hemizygous (tg) and homozygous (tg/tg) ARTE10 mice. Data are expressed as means + SEM; * p
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
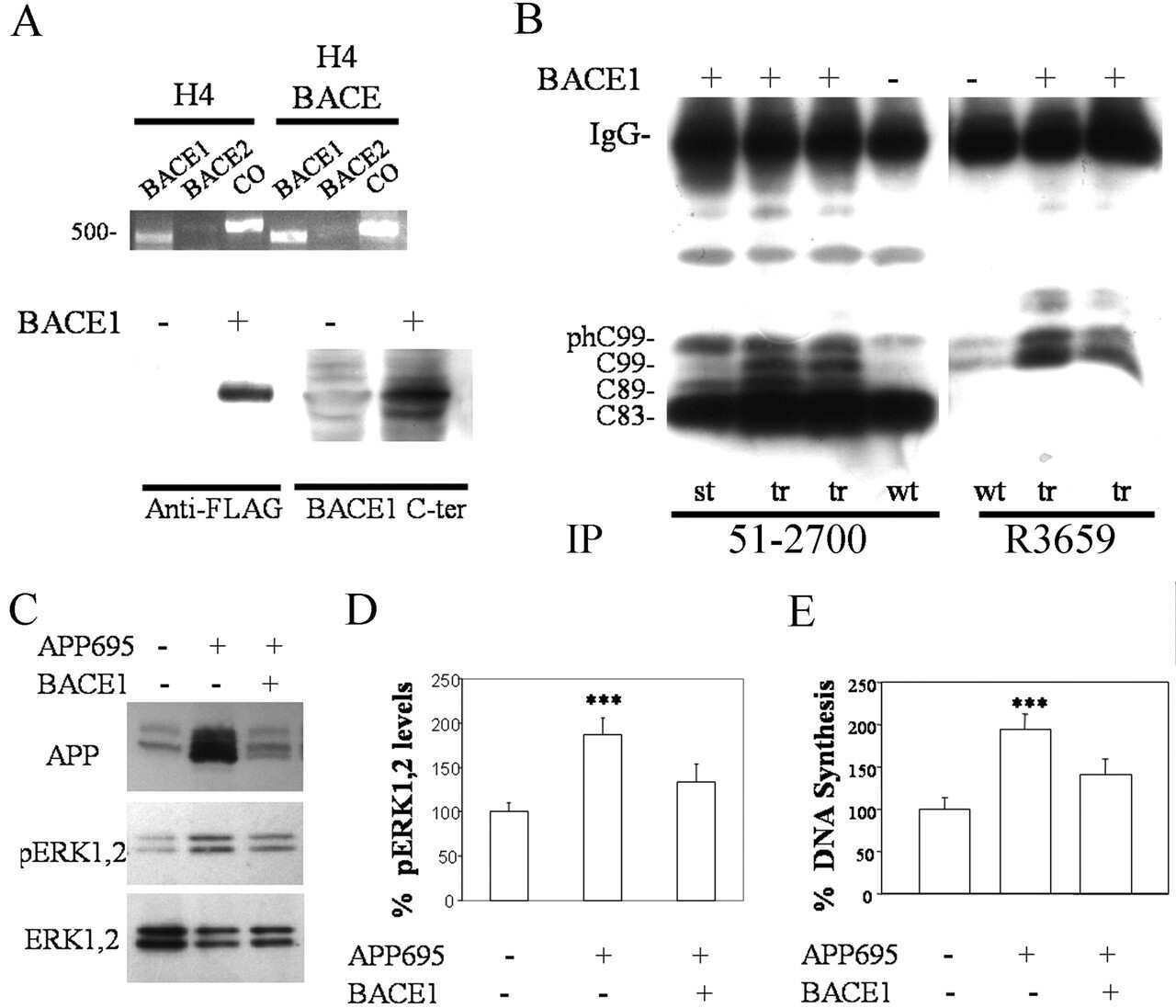
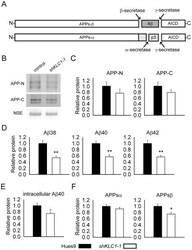
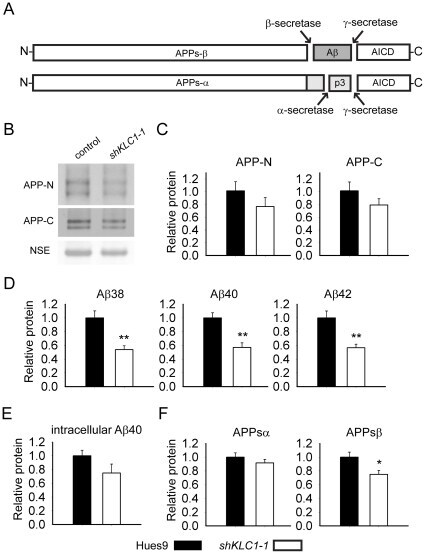
- Figure 4 Human neural cultures with reduced KLC1 exhibit altered APP metabolism. (A) APP proteolytic processing by either beta-and gamma-secretases or alpha- and gamma-secretases produces sAPPbeta and Abeta (shaded dark grey) or sAPPalpha and p3 fragments, respectively. (B-C) PA6 feeder neural differentiation cultures were harvested after seven weeks and equal protein from control and shKLC1-1 cultures were analyzed using Western blots. (B) Representative immunoblots for full length APP in control and shKLC1-1 neural differentiation lysates. Results for both amino (APP-N; LN27) and carboxy terminal (APP-C) antibodies are shown. The APP carboxyl terminal cleavage fragments were not reliably detected. (C) Quantification of full length APP levels relative to NSE. (D) Levels of extracellular human Abeta peptides 38, 40 or 42 amino acids in length detected in media conditioned by control or shKLC1-1 hESC co-cultured with PA6 feeder cells for seven weeks. Human Abeta was not detected from PA6 feeder only cultures. (E) Levels of Triton X-100 soluble intracellular human Abeta-40 in control or shKLC1-1 PA6 feeder differentiation cultures aged seven weeks. Intracellular Abeta peptides 38 or 42 amino acids long were not detectable. (F) Levels of human extracellular sAPPalpha and sAPPbeta were detected in media conditioned by control or shKLC1-1 PA6 feeder cocultures aged in vitro for seven weeks. Based on n = 6 each line; *p
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
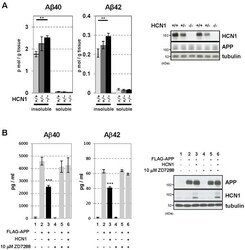
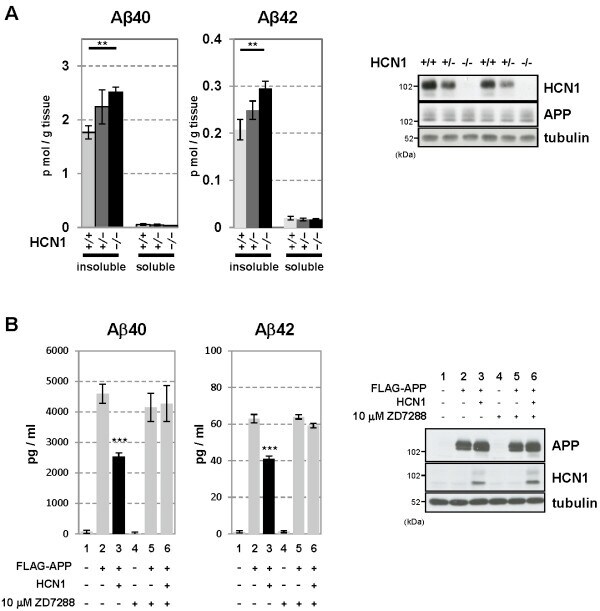
- Figure 4 Functional deficits in the HCN1 channel facilitate Abeta generation. ( A ) Quantification of endogenous murine Abeta40 and Abeta42 in the cortex of 4-month-old HCN1 +/+ , HCN1 +/- , and HCN1 -/- mice by using the sELISA system (left panels). Statistical analysis was performed using the two-tailed Mann-Whitney U -test (average +- SEM, n = 5, **p < 0.01). ( B ) Quantification of Abeta40 and Abeta42 secreted into the culture medium by N2a cells by using the sELISA system. FLAG-APP or FLAG-APP and HCN1 were transiently overexpressed in the cells with (+) or without (-) 10 muM ZD7288 (left panels). Statistical analysis was performed using one-way analysis of variance followed by Tukey's multiple comparison test (average +- SEM, n = 4, ***p < 0.001). Expression levels of APP and HCN1 was confirmed by immunoblotting (right panels in A and B ).
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
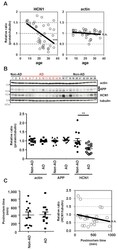
- Figure 6 HCN1 levels decrease in a age- and AD-dependent manner. ( A ) Quantification of the amount of HCN1 and actin in the brain (superior temporal gyrus) of 4-37-year-old cynomolgus monkeys by immunoblotting. Statistical analysis was performed using the two-tailed Pearson's correlation coefficient (HCN1: n = 39, r = -0.5363, ***p < 0.001; actin: n = 39, r = -0.1981, p > 0.05). ( B ) Quantification of the amount of HCN1, APP, actin, and tubulin in age-matched AD and non-AD postmortem brains (superior temporal gyrus) by immunoblotting. Statistical analysis was performed using the two-tailed Mann-Whitney U -test (non-AD: n = 13, AD: n = 12, **p < 0.01). ( C ) Comparison of postmortem time between non-AD and AD samples. Statistical analysis was performed using the two-tailed Mann-Whitney U -test (non-AD: n = 13, AD: n = 12, p = 0.8066). The correlation between postmortem time and HCN1 level was analyzed. Statistical analysis was performed using the two-tailed Pearson's correlation coefficient (n = 39, r = -0.2416, p = 0.2446). Descriptions of all subjects are provided in Table 1 .
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
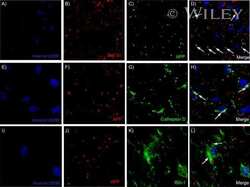
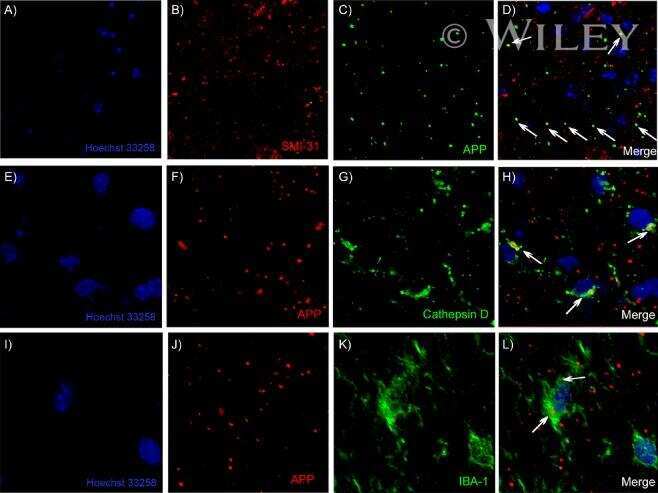
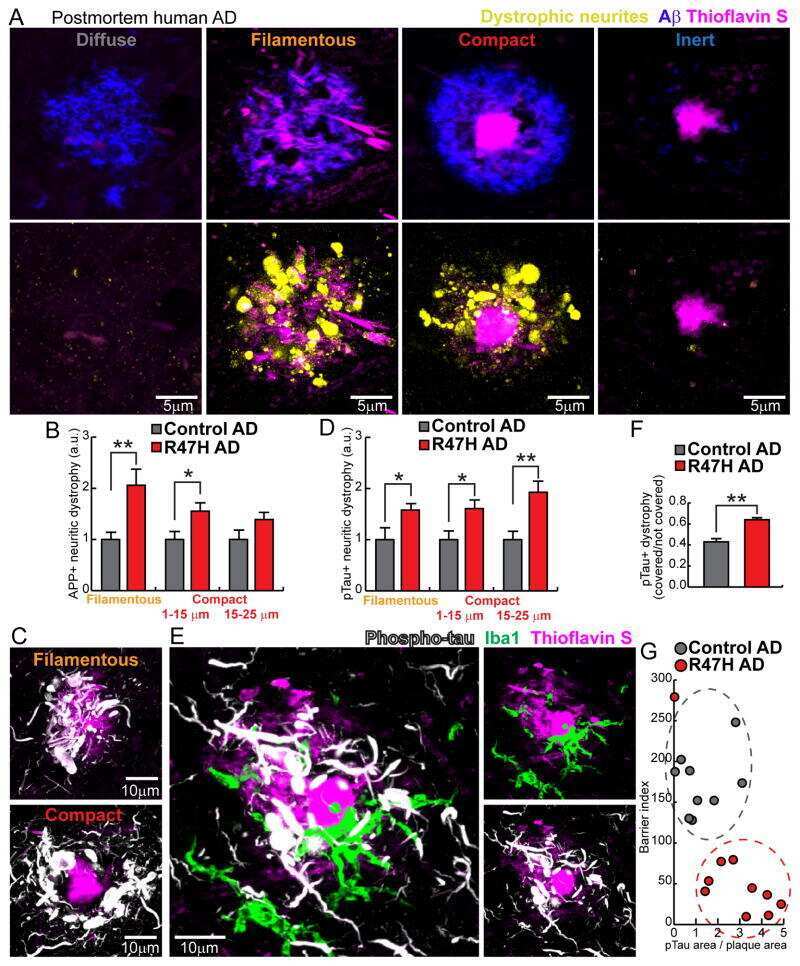
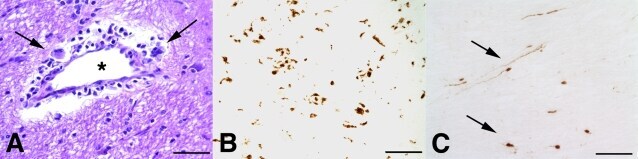
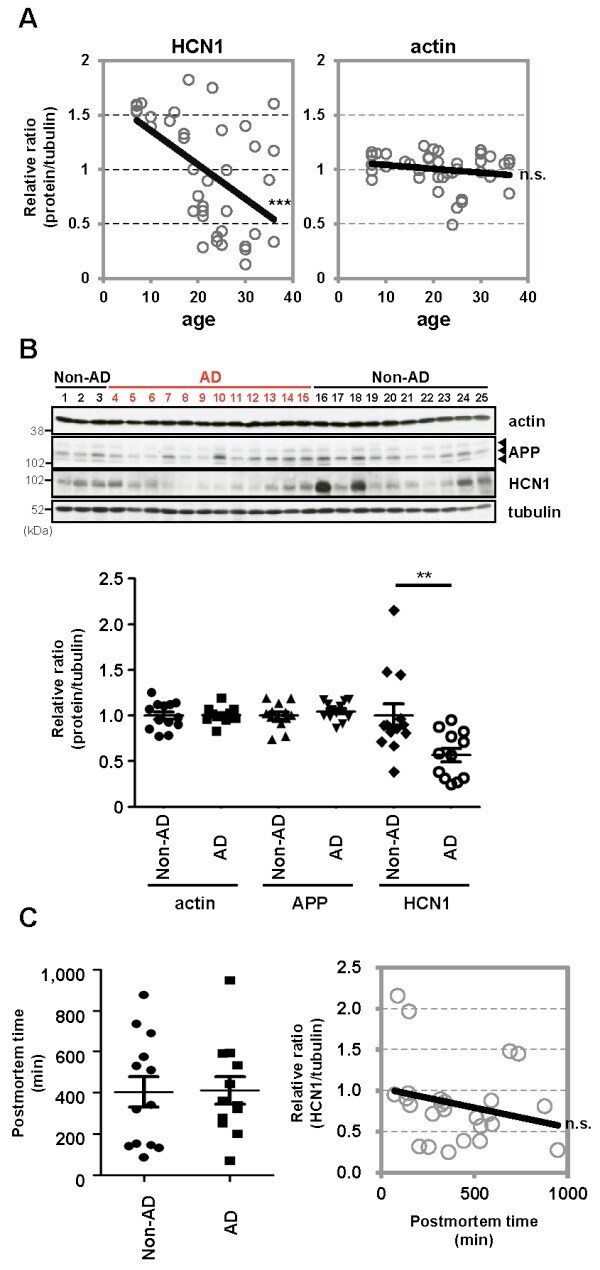
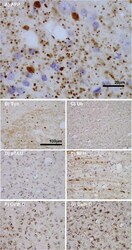
- Figure 3 Features of thalamic pathology. (A) High power (100x objective) photo-micrographs of thalamic APP pathology shows large numbers of 1-2 mum diameter positive elements as well as smaller numbers of much larger spheroids (>6 mum). For comparative purposes, a number of other pathological features were examined for similarities and these were photographed at 40x. (B) Sy38 shows large spheroids of similar size to APP and significant overall loss, but no small deposits. (C) Ubiquitin shows large numbers of small 'dot-like' deposits that are similar in size but very much fewer that those observed with APP . (D) Phospho tau (AT8) labelling shows frequent apparently neuritic labelling. (E) SMI -32, neurofilament H labelling showed strong spheroid labelling in axons emerging from the medial lemniscus but much fewer spheroids in the grey matter. (F) Cathepsin D showed typical lysosomal labelling of small spherical structures in a perinuclear location, predominantly visible in neurons of the PO in NBH animals. (G) Cathepsin D in ME7 animals showed predominantly glial localization and consisted of larger structures or accumulations in perinuclear but also in some glial processes. figure 2015 British Neuropathological Society
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
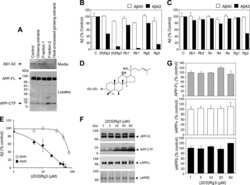
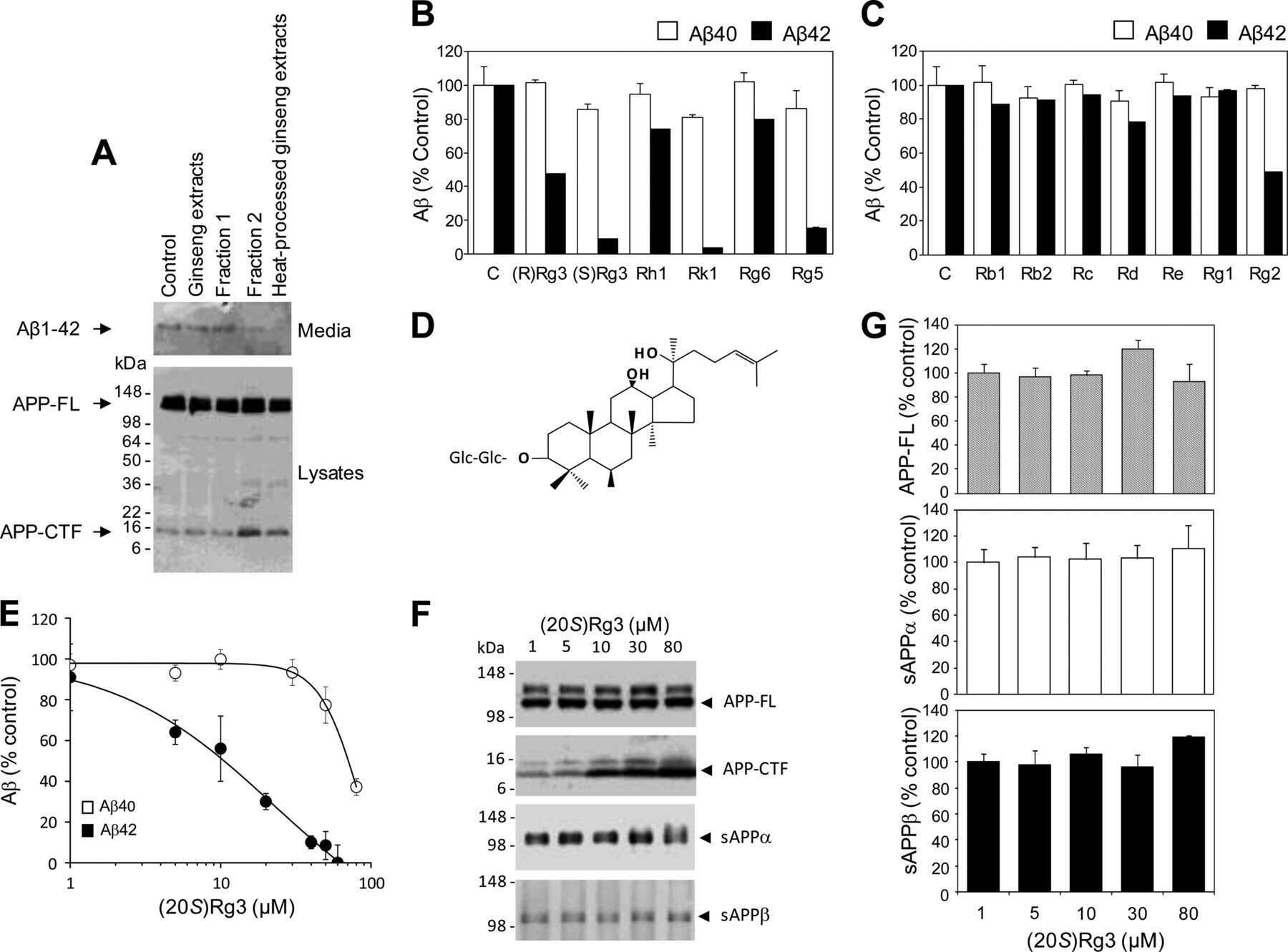
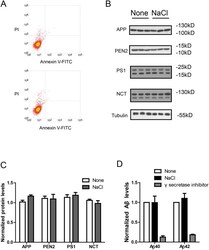
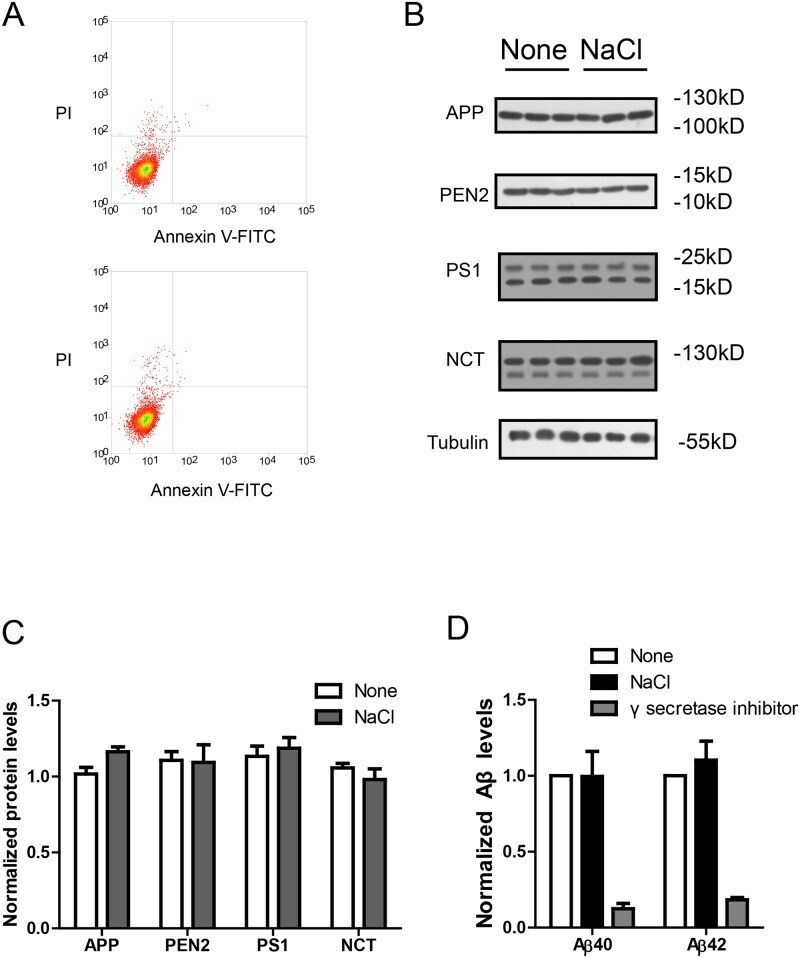
- Fig 2 NaCl treatment did not affect gamma secretase. (A) FACS analysis showed the Annexin V-FITC/ PI staining of HEK293 cells treated with normal (up) or NaCl (low) medium. Western blots (B) and their quantification (C) showed the protein levels of APP and gamma secretase components (PS1, NCT, PEN2) in the lysate of HEK293 cells overexpressing APP after normal or NaCl treatment (n = 3). (D) Enzyme activity assay showed the gamma secretase activity of HEK293 cell lysate after treatment with normal, NaCl medium or gamma secretase inhibitor-L685,458 (n = 3). Data were presented as means +- s.e.m. of indicated numbers of independent experiments.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
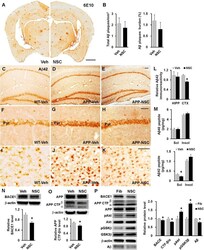
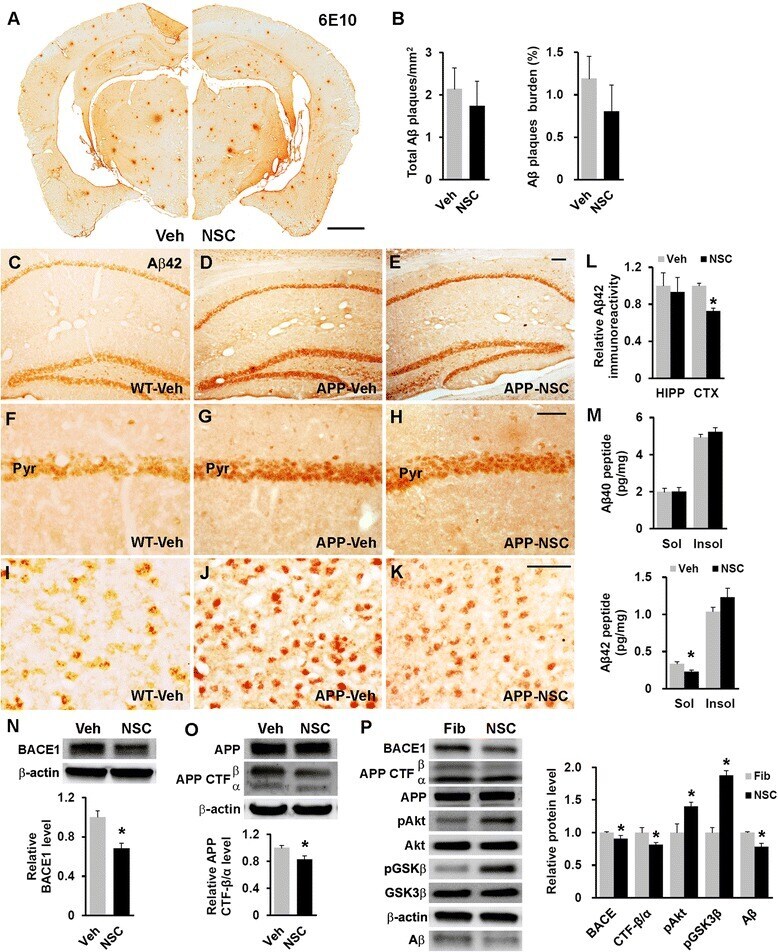
- Fig. 4 hNSC transplantation decreases brain intracellular soluble Abeta42 levels by regulating BACE1 in NSE/APPsw transgenic mice. a Photomicrographs of Abeta plaque (6E10) staining in hNSC-injected (NSC) and vehicle-injected (Veh) transgenic mice. Scale bar, 1 mm. b The number of Abeta plaques per unit area (left) and the percent area of the Abeta plaque load (right) between hNSC graft (NSC) and vehicle injection (Veh; n = 3 per group). c - l Representative images of Abeta42 immunostaining in the brains of vehicle-injected wild-type mice (WT-Veh; c ), and of vehicle-injected (APP-Veh; d ) and hNSC-injected (APP-NSC; e ) transgenic mice. Abeta42 immunostaining of each group in the pyramidal cell layer of the hippocampal CA1 region (HIPP; f - h ) and the posterior parietal cortex (CTX; i - k ). The relative levels of Abeta42 immunostaining in vehicle-injected (Veh) and hNSC-injected (NSC) transgenic mice ( n = 4 per group; l ). Scale bar, 100 mum ( e and h ) and 50 mum ( k ). m Levels of detergent-soluble (Sol) Abeta40/42 and detergent-insoluble (Insol) Abeta40/42 in the brains of hNSC-injected (NSC) and vehicle-injected (Veh) transgenic mice using ELISA kits ( n = 4 per group). n and o Relative levels of BACE1 (NSC, n = 6; Veh, n = 5; n ) and APP CTF-beta/CTF-alpha (NSC, n = 4; Veh, n = 3; o ) using western blot analyses in brains of hNSC-injected (NSC) and vehicle-injected (Veh) transgenic mice. p APPsw-expressing SK-N-MC cells treated with hNSC-derived (NSC) and fibroblast-
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
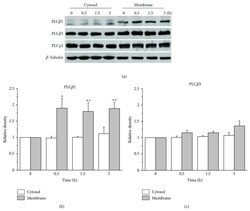
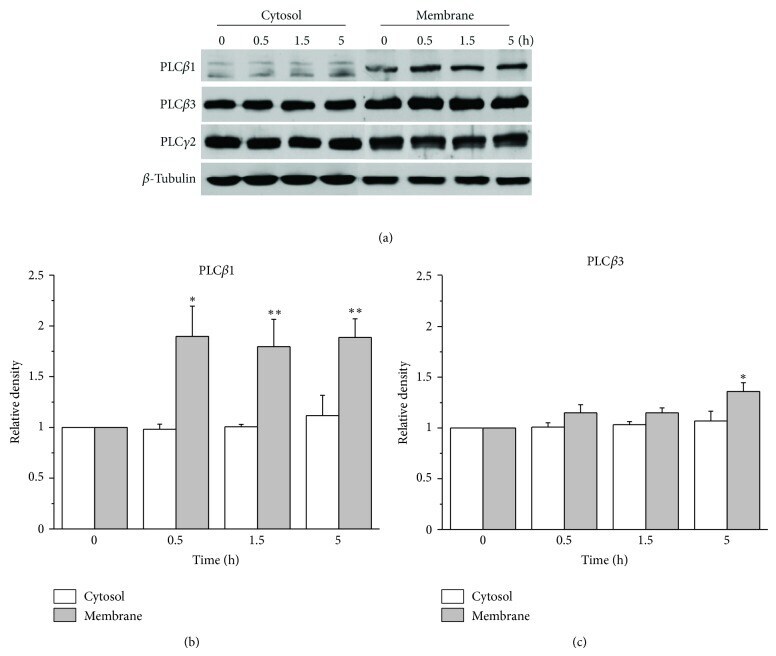
- Figure 3 Augmentation of membrane cholesterol levels increased PLC beta 1 and PLC beta 3 expressions. (a) Representative western blotting results showed specific increase of PLC beta 1 and PLC beta 3 expressions from APP-transfected HeLa cells. Cells were incubated with 75 mu M water-soluble cholesterol for the indicated times. Membrane and cytosol fractions were obtained as described in Materials and Methods. Similar results were obtained from 5 different experiments. Note that PLC beta 1 and PLC beta 3 expressions were increased in time-dependent manner by cholesterol from membrane fraction but not from cytosol fraction. In contrast, PLC gamma 2 expression was not changed by cholesterol. beta -tubulin was used to confirm the amount of proteins loaded. (b, c) Bars correspond to the densitometric analysis of PLC beta 1 and PLC beta 3 expressions from membrane and cytosol fractions ( n = 5). * P < 0.05; ** P < 0.01.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
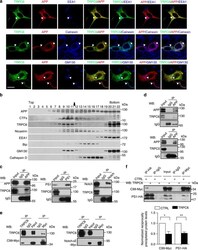
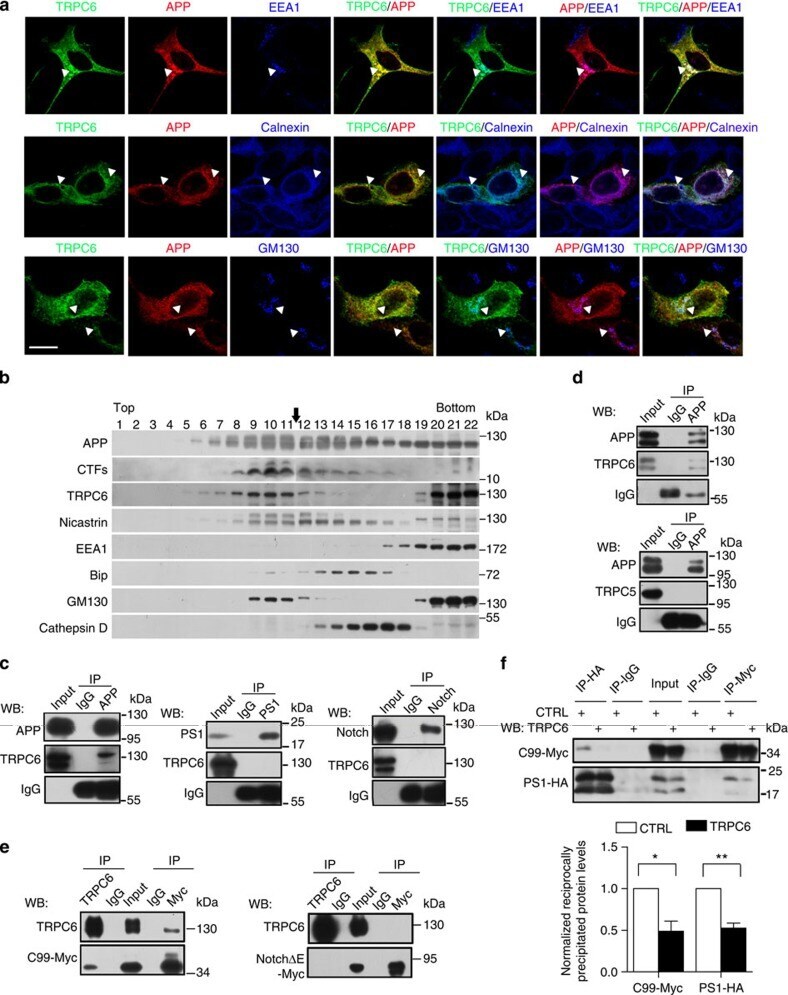
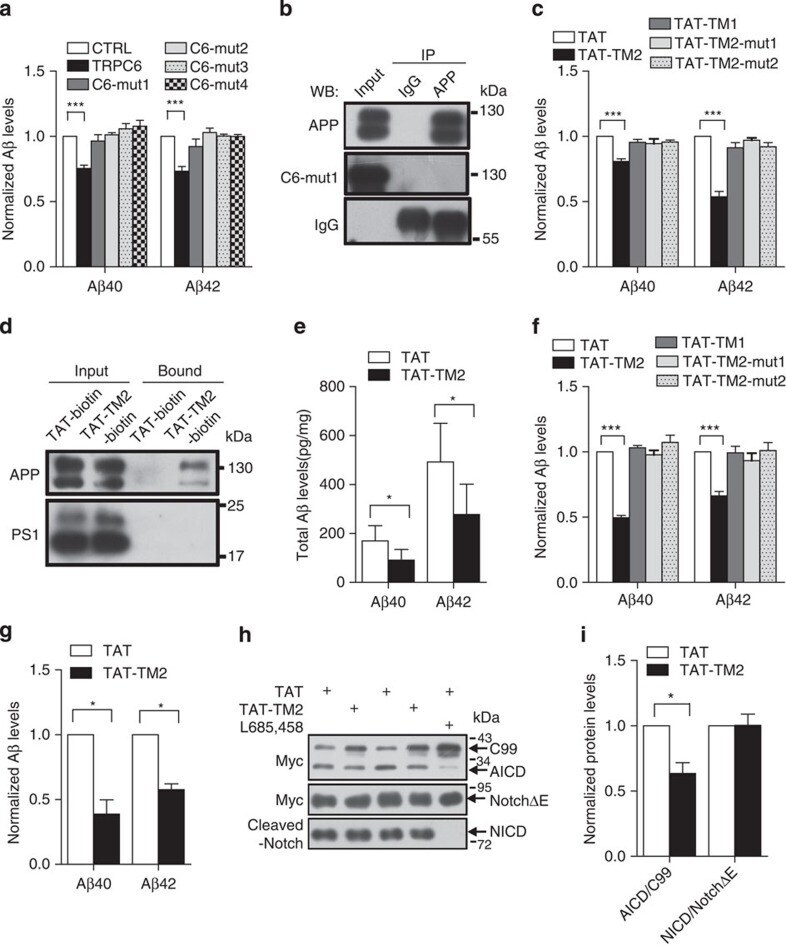
- Figure 3 TRPC6 interacted with APP (C99). ( a ) Immunocytochemical analysis of HEK293 cells transfected with APP-Myc and TRPC6-HA for 3 days. EEA1, Calnexin and GM130 are markers for early endosome, ER and Golgi, respectively. Arrow heads indicated the co-localized signals. Scale bar, 20 mum. ( b ) Immunoblots of indicated proteins after fractionation of mouse brain lysates. Bip and Cathepsin D are markers for ER and lysosome, respectively. Arrow indicated alignment of two gels with 11 lanes running in parallel. ( c ) Wild-type mouse brain lysates precipitated with the antibody against APP, PS1 or Notch, and immunoblotted with indicated antibodies. ( d ) Lysates of HEK293APP stable cells transfected with TRPC5 or 6 for 2 days precipitated with APP antibody, and immunoblotted with indicated antibodies. ( e ) Lysates of HEK293TRPC6 stable cells transfected with C99-Myc or NotchDeltaE-Myc for 1 day, treated with 10 muM L685,458 for 12 h and then precipitated with Myc or TRPC6 antibody, and immunoblotted with indicated antibodies. ( f ) Lysates of HEK293C99-Myc stable cells transfected with PS1-HA together with YFP or TRPC6 for 2 days and precipitated with Myc or HA antibody, and immunoblotted with indicated antibodies. Lower, quantification of the reciprocal precipitated protein levels of C99 and PS1 ( n =4). CTRL, transfection with YFP. Data were presented as means+-s.e.m. of indicated numbers of independent experiments. Two-tailed Student's t -test was performed. * P
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
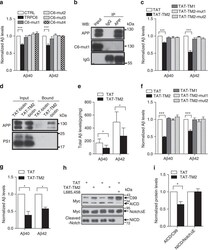
- Figure 4 TAT-TM2 reduced Abeta levels in vitro and in vivo . ( a ) ELISA examination of Abeta levels in the medium of HEK293APP cells transfected with TRPC6 or its mutants within second transmembrane domain (TM2) for 2 days ( n =3-6). ( b ) Lysates of HEK293APP stable cells transfected with C6-mut1 for 2 days precipitated with APP antibody and immunoblotted with indicated antibodies. ( c ) The Abeta levels in the medium of HEK293APP cells treated with 2 muM indicated peptides for 12 h ( n =3-6). ( d ) Lysates of HEK293APP stable cells incubated with 5 muM TAT-biotin or TAT-TM2-bitoin for 6 h precipitated with avidin beads and immunoblotted with indicated antibodies. ( e ) ELISA examination in a blinded manner of Abeta levels in the brain lysates of 5 month male APP/PS1 mice intra-peritoneally injected with 200 mul 2.5 mM TAT or TAT-TM2 for 3 h ( n =8 mice). ( f ) The Abeta levels generated in the in vitro purified C99 cleavage assay treated with 2 muM of indicated peptides for 2 h ( n =4). ( g ) Abeta levels in the medium of COS7C99 stable cells treated with 5 muM TAT-TM2 for 12 h ( n =3-4). ( h ) Immunoblots of AICD in HEK293C99 cells treated with 5 muM TAT-TM2 for 12 h or NICD in HEK293APP cells transfected with NotchE-myc for 1 day and then treated with 5 muM TAT-TM2 for 12 h. ( i ) Quantification of the ratio of AICD/C99 or NICD/NotchDeltaE ( n =3-4). CTRL, transfection with YFP . Data were presented as means+-s.e.m. of indicated numbers of independent experiments. Two-ta
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
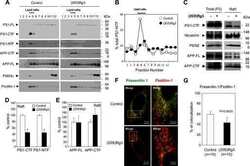
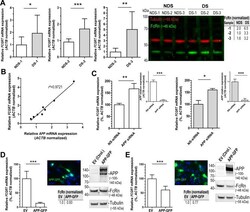
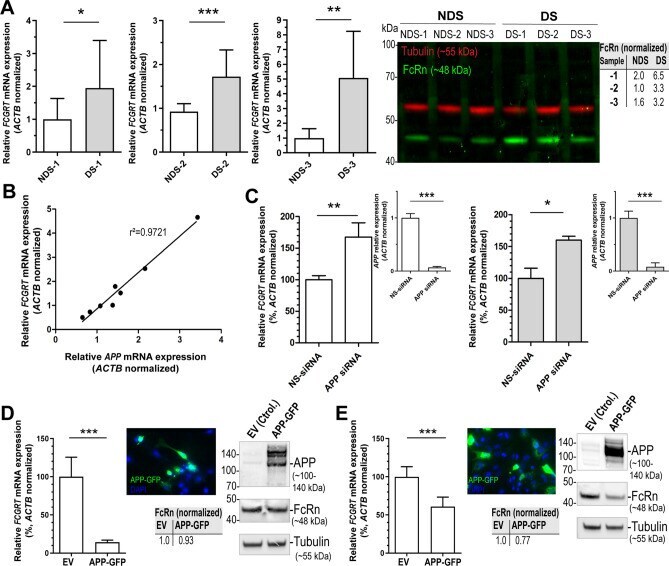
- Figure 5 FcRn expression in trisomic fibroblasts and impact of APP on FCGRT expression. ( A ) FCGRT mRNA relative fold expression (left), and FcRn protein expression (right) in trisomic (DS-1, -2, and -3) and diploid fibroblasts (NDS-1, -2, and -3) from sex and age-matched donors. Each bar represents the mean +- SD from two separate measurements performed in quadruplicate. Upper right panel: densitometric analysis of FcRn bands normalized to tubulin. ( B ) Linear regression analysis of FCGRT versus APP mRNA relative expression in diploid and trisomic fibroblasts (n = 8 cell lines). Each point represents the mean from two separate measurements performed in quadruplicate. ( C ) FCGRT mRNA relative expression in diploid (left panel, line AG07095, NDS-2) and trisomic fibroblasts (right panel, line AG06922, DS-2) analyzed 96 h post-transfection with a non-sense siRNA (NS-siRNA, control) or a siRNA pool against APP ( APP siRNA). ( D , E ) FCGRT mRNA relative expression (left panel), APP-GFP expression detected by fluorescence microscopy (upper middle panel), and FcRn and APP protein expression detected by immunoblotting (right panel) in AG07095 (NDS-2. D ) and HepG2 ( E ). Lower middle panel: densitometric analysis of FcRn bands normalized to tubulin. Cells were analyzed 48 h post-transfection with an empty vector (EV, control) or an APP-GFP plasmid. Each bar represents the mean +- SD from at least two measurements performed in quadruplicate. ***P < 0.001, **P < 0.01, *P < 0.05, St
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
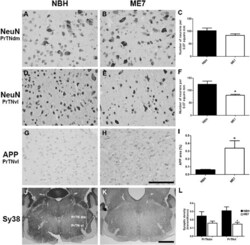
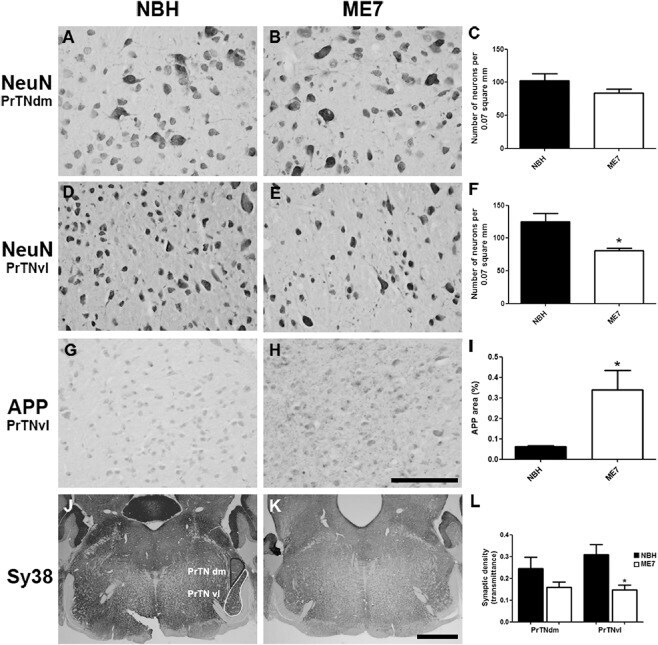
- Figure 6 Neuronal, APP and synaptic pathology in the principal trigeminal nucleus. (A-F) Micrographs illustrating NeuN labelling in PrTNdm and PrTNvl of NBH (A,D) and ME7 (B,E) animals, respectively. Masks used to quantify the number of neurons per area, generated in ImageJ software, are shown in supplemental material. ME7 induced significant neuronal loss in PrTNvl (F) but not in PrTNdm (C). * t -test, P < 0.01. n = 7 ME7 , n = 6 NBH . Scale bar: 100 mum. (G-I) Immunolabelling for APP in the PrTN of NBH (G) and ME7 (H). Greyscale images were treated to differentiate brown and blue labelling (see methods) and then thresholded in ImageJ software in order to quantifty APP deposits only. The percentage of APP -labelled area in the PrTN of NBH and ME7 animals ( n = 4 in each group) is shown in (I). * t -test, P < 0.05. (J-L) Immunolabelling for Sy38 in PrTN of NBH (J) and ME7 animals ( K ). The regions of PrTN : ventrolateral part (in white) and dorsomedial part (in black) are delineated in (J). Contrast indices are shown for synaptophysin transmittance measurements in PrTNvl and PrTNdm of NBH (J) and ME7 ( K ) groups. Statistically significant reduction of synaptic density was found in PrTNvl , but not in PrTNdm (L). * t -test, P < 0.01. PrTNVL , principal trigeminal nucleus, ventrolateral part; PrTNdm, principal trigeminal nucleus, dorsomedial part. n = 10 ME7 and 6 NBH . Scale bar: 1 mm. All data are shown as mean +- SEM . figure 2015 British Neuropathological Society
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
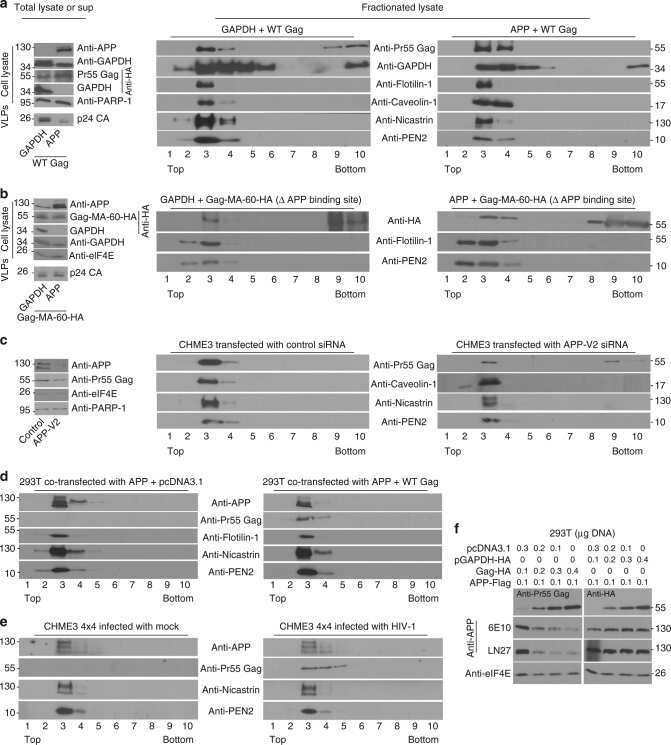
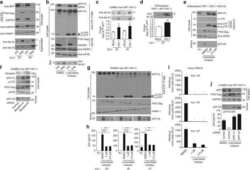
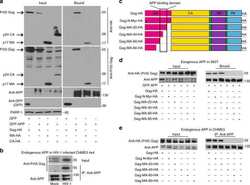
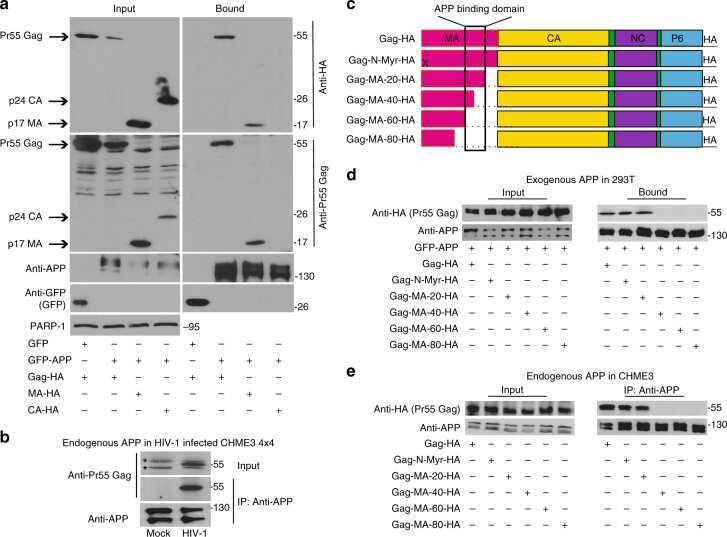
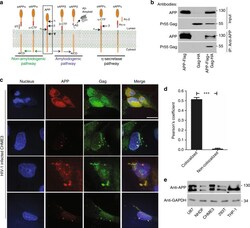
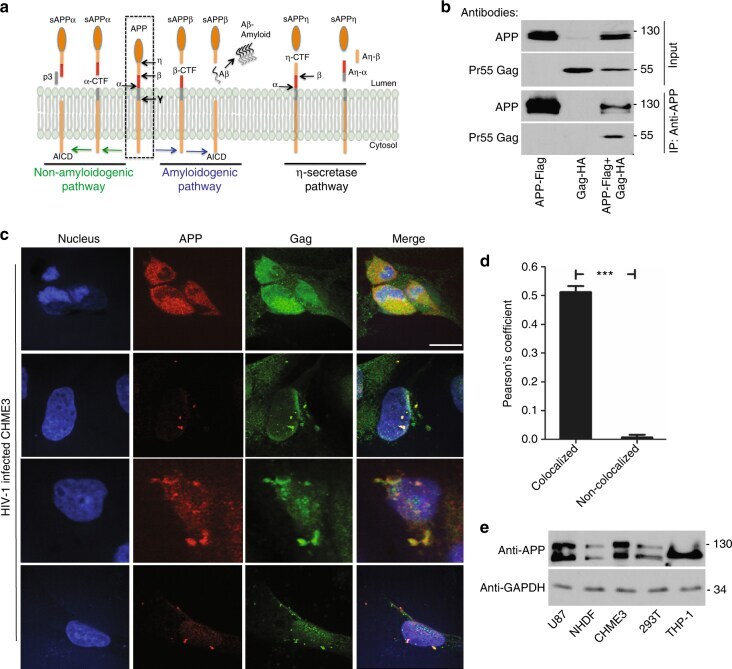
- Fig. 1 APP is highly expressed in macrophages and microglia and binds HIV-1 Gag. a APP processing through amyloidogenic, non-amyloidogenic and eta-secretase pathways involves alpha-, beta-, gamma- and eta-secretases. The Abeta peptide resulting in toxic amyloid oligomers and plaques is generated by sequential cleavages by beta- and gamma-secretases via the amyloidogenic pathway (central). b Human APP 770 (APP-Flag) binds HIV-1 Gag (Gag-HA) in anti-APP co-IP from transfected 293T cells. c Endogenous APP and Gag colocalize in CHME3 cells infected with HIV-1 carrying vesicular stomatitis virus G (VSV-G) envelope glycoprotein at 16 h post infection (h.p.i). Nuclei were stained with Hoechst (blue). All images were obtained using a 100x oil objective of a spinning disk confocal microscope. Scale bar = 10 mum. d Quantification of APP and Gag as determined by Pearson's coefficient in at least 10 random fields of view from samples as in c , shown as Mean +- SEM. The Pearson's correlation coefficients were r = 0.512 +- 0.02. e Endogenous APP levels in glioblastoma (U87), normal human dermal fibroblasts (NHDF), microglia (CHME3), 293T and differentiated THP-1 cells. Molecular weight markers (in kDa) are shown to the right of WBs
- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
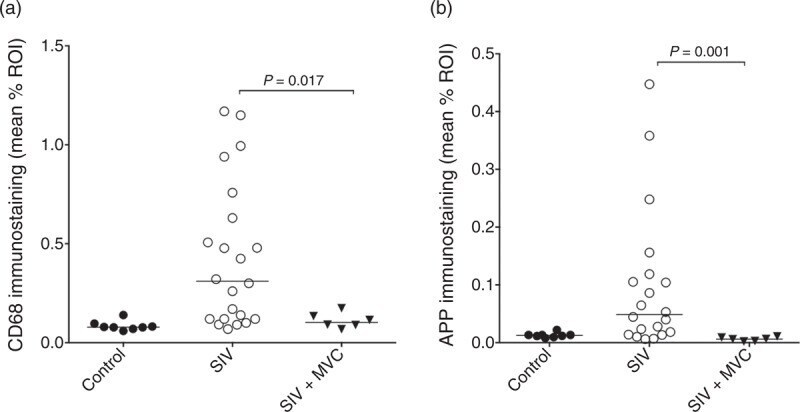
- Fig. 4 Treating SIV-infected macaques with maraviroc (MVC) significantly reduced CD68 immunostaining (a) in the basal ganglia compared with untreated SIV-infected macaques (open circles) to levels equivalent to uninfected macaques (closed circles). SIV-infected macaques treated with MVC had significantly reduced APP immunostaining in the corpus callosum compared with untreated SIV-infected macaques (circles) to levels unchanged from untreated SIV-infected macaques. Bars represent median values; Mann-Whitney test.