Explore
Explore Validate
Validate Learn
Learn Western blot
Western blot Immunocytochemistry
ImmunocytochemistryAntibody data
- Antibody Data
- Antigen structure
- References [19]
- Comments [0]
- Validations
- Immunocytochemistry [2]
- Immunohistochemistry [1]
- Other assay [3]
Submit
Validation data
Reference
Comment
Report error
- Product number
- PA1-308 - Provider product page

- Provider
- Invitrogen Antibodies
- Product name
- Estrogen Receptor alpha Polyclonal Antibody
- Antibody type
- Polyclonal
- Antigen
- Synthetic peptide
- Description
- PA1-308 has been successfully used in Western blot, immunofluorescence and immunohistochemistry procedures. By Western blot, this antibody detects an ~64 kDa protein representing endogenous levels of ER alpha from MCF-7 cell extracts. Immunohistochemical staining of ER alpha in rat prostate with PA1-308 results in primarily nuclear staining. The PA1-308 immunogen is a synthetic peptide corresponding to residues M(1) T M T L H T K A S G M A L L H Q I Q G N(21) C of human ER alpha. This peptide sequence is completely conserved between human, rat and mouse and is highly conserved between sheep, porcine, camel, bovine and chicken ER alpha. PA1-308 immunizing peptide (Cat. # PEP-036) is available for use in neutralization and control experiments. Recombinant ER alpha expressed in baculovirus system (Cat. # RP-310) can be purchased for control experiments in Western blot and gel shift assays.
- Reactivity
- Human, Rat
- Host
- Rabbit
- Isotype
- IgG
- Vial size
- 50 μg
- Concentration
- 1 mg/mL
- Storage
- -20°C, Avoid Freeze/Thaw Cycles
Submitted references Nitric Oxide Regulates Estrus Cycle Dependent Colonic Motility in Mice.
A Sexually Dimorphic Area of the Dorsal Hypothalamus in Mice and Common Marmosets.
Characterization of bladder sensory neurons in the context of myelination, receptors for pain modulators, and acute responses to bladder inflammation.
Expression and localization of aromatase P450AROM, estrogen receptor-α, and estrogen receptor-β in the developing fetal bovine frontal cortex.
Differential effects of steroids on the synthesis of polyunsaturated fatty acids by human neuroblastoma cells.
Estrogen increases survival in an orthotopic model of glioblastoma.
Pretreatment with phytoestrogen-rich plant decreases breast tumor incidence and exhibits lower profile of mammary ERalpha and ERbeta.
Phytosterol Pygeum africanum regulates prostate cancer in vitro and in vivo.
Genistein affects HER2 protein concentration, activation, and promoter regulation in BT-474 human breast cancer cells.
Androgen and estrogen receptor-mediated mechanisms of testosterone action in male rat pelvic autonomic ganglia.
Apoptotic and anti-proliferative effects of 17beta-estradiol and 17beta-estradiol-like compounds in the Hep3B cell line.
Phytoestrogens in common herbs regulate prostate cancer cell growth in vitro.
Rapid actions of estradiol on cyclic amp response-element binding protein phosphorylation in dorsal root ganglion neurons.
Multiple forms of estrogen receptor-alpha in cerebral blood vessels: regulation by estrogen.
Liganded androgen receptor interaction with beta-catenin: nuclear co-localization and modulation of transcriptional activity in neuronal cells.
17-Beta-estradiol regulates expression of genes that function in macrophage activation and cholesterol homeostasis.
17-Beta-estradiol regulates expression of genes that function in macrophage activation and cholesterol homeostasis.
Sex difference in septal neurons projecting axons to midbrain central gray in rats: a combined double retrograde tracing and ER-immunohistochemical study.
Cancer-associated myofibroblasts possess various factors to promote endometrial tumor progression.
Balasuriya GK, Nugapitiya SS, Hill-Yardin EL, Bornstein JC
Frontiers in neuroscience 2021;15:647555
Frontiers in neuroscience 2021;15:647555
A Sexually Dimorphic Area of the Dorsal Hypothalamus in Mice and Common Marmosets.
Moe Y, Kyi-Tha-Thu C, Tanaka T, Ito H, Yahashi S, Matsuda KI, Kawata M, Katsuura G, Iwashige F, Sakata I, Akune A, Inui A, Sakai T, Ogawa S, Tsukahara S
Endocrinology 2016 Dec;157(12):4817-4828
Endocrinology 2016 Dec;157(12):4817-4828
Characterization of bladder sensory neurons in the context of myelination, receptors for pain modulators, and acute responses to bladder inflammation.
Forrest SL, Osborne PB, Keast JR
Frontiers in neuroscience 2013;7:206
Frontiers in neuroscience 2013;7:206
Expression and localization of aromatase P450AROM, estrogen receptor-α, and estrogen receptor-β in the developing fetal bovine frontal cortex.
Peruffo A, Giacomello M, Montelli S, Corain L, Cozzi B
General and comparative endocrinology 2011 Jun 1;172(2):211-7
General and comparative endocrinology 2011 Jun 1;172(2):211-7
Differential effects of steroids on the synthesis of polyunsaturated fatty acids by human neuroblastoma cells.
Extier A, Perruchot MH, Baudry C, Guesnet P, Lavialle M, Alessandri JM
Neurochemistry international 2009 Sep;55(5):295-301
Neurochemistry international 2009 Sep;55(5):295-301
Estrogen increases survival in an orthotopic model of glioblastoma.
Barone TA, Gorski JW, Greenberg SJ, Plunkett RJ
Journal of neuro-oncology 2009 Oct;95(1):37-48
Journal of neuro-oncology 2009 Oct;95(1):37-48
Pretreatment with phytoestrogen-rich plant decreases breast tumor incidence and exhibits lower profile of mammary ERalpha and ERbeta.
Cherdshewasart W, Panriansaen R, Picha P
Maturitas 2007 Oct 20;58(2):174-81
Maturitas 2007 Oct 20;58(2):174-81
Phytosterol Pygeum africanum regulates prostate cancer in vitro and in vivo.
Shenouda NS, Sakla MS, Newton LG, Besch-Williford C, Greenberg NM, MacDonald RS, Lubahn DB
Endocrine 2007 Feb;31(1):72-81
Endocrine 2007 Feb;31(1):72-81
Genistein affects HER2 protein concentration, activation, and promoter regulation in BT-474 human breast cancer cells.
Sakla MS, Shenouda NS, Ansell PJ, Macdonald RS, Lubahn DB
Endocrine 2007 Aug;32(1):69-78
Endocrine 2007 Aug;32(1):69-78
Androgen and estrogen receptor-mediated mechanisms of testosterone action in male rat pelvic autonomic ganglia.
Purves-Tyson TD, Arshi MS, Handelsman DJ, Cheng Y, Keast JR
Neuroscience 2007 Aug 10;148(1):92-104
Neuroscience 2007 Aug 10;148(1):92-104
Apoptotic and anti-proliferative effects of 17beta-estradiol and 17beta-estradiol-like compounds in the Hep3B cell line.
Huang EJ, Wu CC, Huang HP, Liu JY, Lin CS, Chang YZ, Lin JA, Lin JG, Chen LM, Lee SD, Kuo WW, Huang CY
Molecular and cellular biochemistry 2006 Oct;290(1-2):1-7
Molecular and cellular biochemistry 2006 Oct;290(1-2):1-7
Phytoestrogens in common herbs regulate prostate cancer cell growth in vitro.
Shenouda NS, Zhou C, Browning JD, Ansell PJ, Sakla MS, Lubahn DB, Macdonald RS
Nutrition and cancer 2004;49(2):200-8
Nutrition and cancer 2004;49(2):200-8
Rapid actions of estradiol on cyclic amp response-element binding protein phosphorylation in dorsal root ganglion neurons.
Purves-Tyson TD, Keast JR
Neuroscience 2004;129(3):629-37
Neuroscience 2004;129(3):629-37
Multiple forms of estrogen receptor-alpha in cerebral blood vessels: regulation by estrogen.
Stirone C, Duckles SP, Krause DN
American journal of physiology. Endocrinology and metabolism 2003 Jan;284(1):E184-92
American journal of physiology. Endocrinology and metabolism 2003 Jan;284(1):E184-92
Liganded androgen receptor interaction with beta-catenin: nuclear co-localization and modulation of transcriptional activity in neuronal cells.
Pawlowski JE, Ertel JR, Allen MP, Xu M, Butler C, Wilson EM, Wierman ME
The Journal of biological chemistry 2002 Jun 7;277(23):20702-10
The Journal of biological chemistry 2002 Jun 7;277(23):20702-10
17-Beta-estradiol regulates expression of genes that function in macrophage activation and cholesterol homeostasis.
Kramer PR, Wray S
The Journal of steroid biochemistry and molecular biology 2002 Jul;81(3):203-16
The Journal of steroid biochemistry and molecular biology 2002 Jul;81(3):203-16
17-Beta-estradiol regulates expression of genes that function in macrophage activation and cholesterol homeostasis.
Kramer PR, Wray S
The Journal of steroid biochemistry and molecular biology 2002 Jul;81(3):203-16
The Journal of steroid biochemistry and molecular biology 2002 Jul;81(3):203-16
Sex difference in septal neurons projecting axons to midbrain central gray in rats: a combined double retrograde tracing and ER-immunohistochemical study.
Tsukahara S, Yamanouchi K
Endocrinology 2002 Jan;143(1):285-94
Endocrinology 2002 Jan;143(1):285-94
Cancer-associated myofibroblasts possess various factors to promote endometrial tumor progression.
Orimo A, Tomioka Y, Shimizu Y, Sato M, Oigawa S, Kamata K, Nogi Y, Inoue S, Takahashi M, Hata T, Muramatsu M
Clinical cancer research : an official journal of the American Association for Cancer Research 2001 Oct;7(10):3097-105
Clinical cancer research : an official journal of the American Association for Cancer Research 2001 Oct;7(10):3097-105
No comments: Submit comment
Supportive validation
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
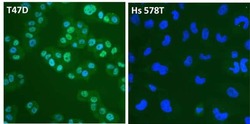
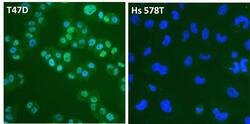
- Experimental details
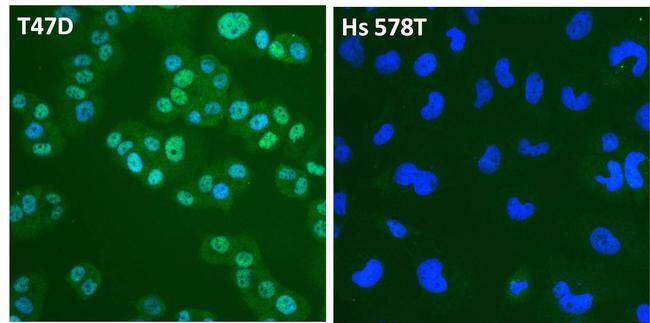
- Immunofluorescent analysis of Estrogen Receptor alpha (ER alpha, green) in T47D and Hs 578T breast cancer cells. The cells were fixed with 4% paraformaldehyde for 15 minutes at room temperature, permeabilized with 0.1% Triton X-100 for 15 minutes at room temperature, and blocked with 0.3% BSA in PBS for at least 30 minutes at room temperature. Cells were stained with an ER alpha polyclonal antibody (Product # PA1-308) at a dilution of 1:4000 in blocking buffer overnight at 4C, and then incubated with a DyLight 488-conjugated goat anti-rabbit IgG secondary antibody (Product # 35552) at a dilution of 1:500 for at least 1 hour at room temperature. Nuclei (blue) were stained with DAPI (Product # 62247). Images were taken on a Thermo Scientific ToxInsight Instrument at 20X magnification.
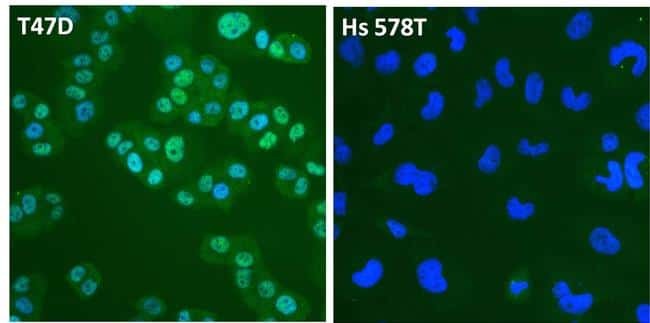
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- Immunofluorescent analysis of Estrogen Receptor alpha (ER alpha, green) in T47D and Hs 578T breast cancer cells. The cells were fixed with 4% paraformaldehyde for 15 minutes at room temperature, permeabilized with 0.1% Triton X-100 for 15 minutes at room temperature, and blocked with 0.3% BSA in PBS for at least 30 minutes at room temperature. Cells were stained with an ER alpha polyclonal antibody (Product # PA1-308) at a dilution of 1:4000 in blocking buffer overnight at 4C, and then incubated with a DyLight 488-conjugated goat anti-rabbit IgG secondary antibody (Product # 35552) at a dilution of 1:500 for at least 1 hour at room temperature. Nuclei (blue) were stained with DAPI (Product # 62247). Images were taken on a Thermo Scientific ToxInsight Instrument at 20X magnification.
Supportive validation
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
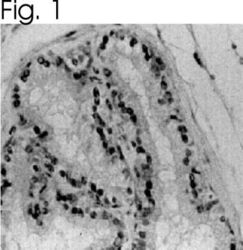
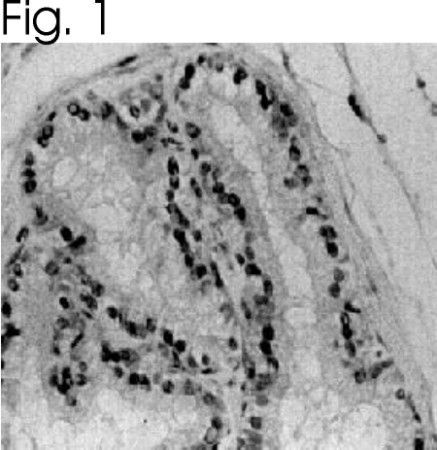
- Immunolocalization of ER alpha in rat prostate using Product # PA1-308.
Supportive validation
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
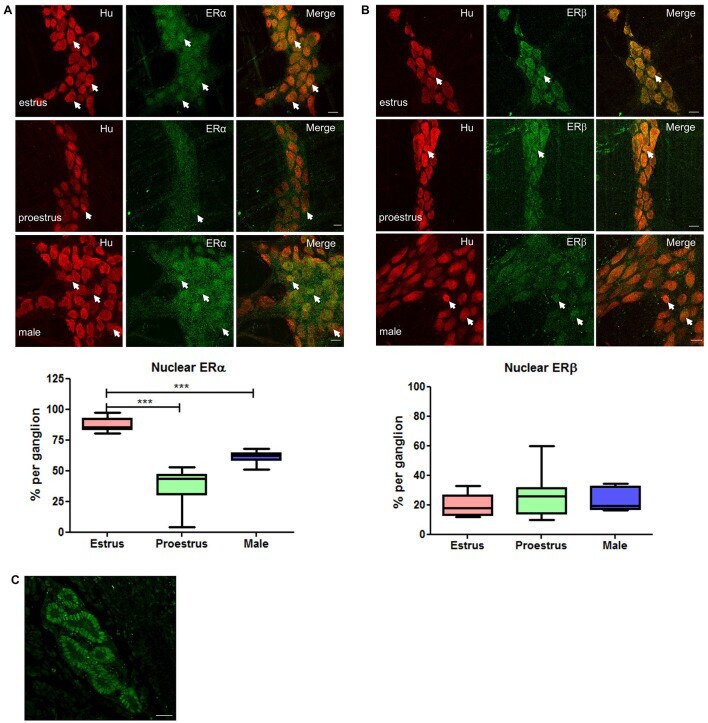
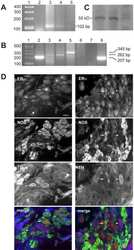
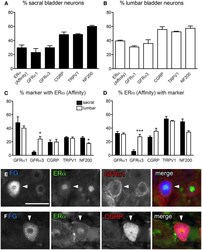
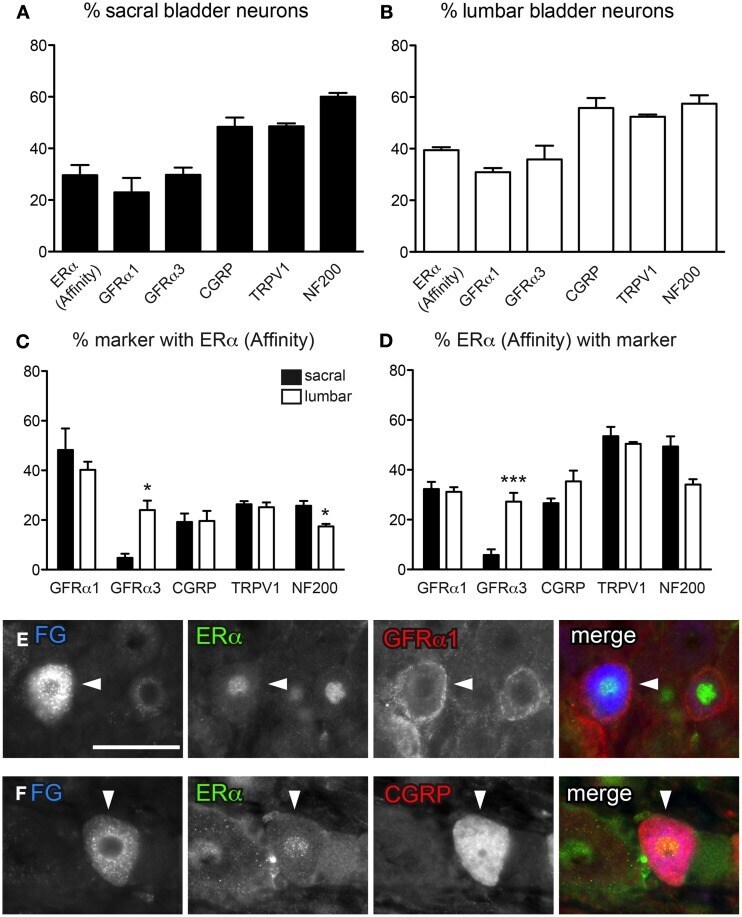
- Figure 2 Distribution of ERalpha and GFRs in bladder sensory neurons . This figure illustrates immunostaining for ERalpha using the antibody purchased from Affinity Bioreagents. (A, B) The proportion of sacral (A) and lumbar (B) bladder-projecting neurons containing ERalpha-, GFRalpha1, GFRalpha3-, CGRP-, TRPV1-, and NF200-immunoreactivity (IR). (C) The proportion of GFRalpha1-, GFRalpha3-, CGRP-, TRPV1-, and NF200-IR bladder sensory neurons containing ERalpha-IR. A higher proportion of GFRalpha3-IR neurons contained ERalpha-IR in lumbar vs. sacral DRGs ( * P = 0.01), whereas a higher proportion of NF200-IR neurons contained ERalpha-IR in sacral vs. lumbar DRGS ( * P = 0.03). (D) The proportion of ERalpha-IR bladder sensory neurons containing each marker. A higher proportion of ERalpha-IR neurons contained GFRalpha3-IR in lumbar vs. sacral DRGs ( *** P < 0.001). Data represents the mean +- s.e.m. ( n = 5), analyzed by a paired t -test on arcsine-transformed data. (E) Co-expression of ERalpha- and GFRalpha1-IR in a FluoroGold (FG)-labeled neuron. (F) Co-expression of ERalpha- and CGRP-IR in a FG-labeled neuron. Images are shown as monochrome image sets and the merged colorized image. The color of each label in the merged image is indicated by the color of the text in the relevant monochrome image. Scale bar in panel (E) represents 50 mum and applies to all images. Arrowheads indicate examples of retrogradely labeled neurons and their relative location in matching panels.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
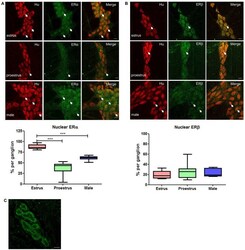
- FIGURE 3 Whole mount preparations of myenteric plexus expressing pan neuronal marker Hu in red and ERalpha/ERbeta in green. Significantly higher numbers of myenteric neurons expressed ERalpha in the nucleus during estrus while having some cytoplasmic expression compared to proestrus females and male myenteric neurons expressing less nuclear and cytoplasmic ERalpha. ERbeta expression was more cytoplasmic in all tissue samples and showed no significant difference in expression between groups. Scale bar = 20 mum ( n = 3, One-way ANOVA, *** p < 0.001). (A) Localization of ERalpha in the myenteric plexus preparations of estrus, proestrus, and male colon: ERalpha labeling was observed in more than 90% of myenteric neurons in both male and female mice. Images of representative myenteric ganglia, demonstrating neurons stained for Hu, ERalpha, and merged images of both Hu and ERalpha. During estrus stage, ERalpha labeling is predominantly nuclear while during proestrus and in male mice both nuclear and cytoplasmic labeling was observed (arrows indicate nuclear labeling of ERalpha). Scale bars = 20 mum. Graph shows quantified data ( n = 3) where there was significantly higher nuclear localization observed during estrus stage compared to both proestrus and male preparations (*** p < 0.001). (B) Localization of ERbeta in the myenteric plexus preparations of estrus, proestrus, and male colon: localization of the estrogen receptor beta (ERbeta) is unchanged during estrus. ERbeta labeling w