Explore
Explore Validate
Validate Learn
Learn Western blot
Western blotAntibody data
- Antibody Data
- Antigen structure
- References [18]
- Comments [0]
- Validations
- Western blot [1]
- Immunocytochemistry [2]
- Immunohistochemistry [1]
- Other assay [6]
Submit
Validation data
Reference
Comment
Report error
- Product number
- PA1-309 - Provider product page

- Provider
- Invitrogen Antibodies
- Product name
- Estrogen Receptor alpha Polyclonal Antibody
- Antibody type
- Polyclonal
- Antigen
- Synthetic peptide
- Description
- PA1-309 detects estrogen receptor (ER) alpha from human, porcine and rat tissues and cells. PA1-309 has been successfully used in Western blot, immunofluoresence and immunohistochemistry procedures. By Western blot, this antibody detects an ~64 kDa protein representing ER alpha from MCF-7 cell extracts. Immunohistochemical staining of ER alpha in rat prostate with PA1-309 results in primarily nuclear staining. The PA1-309 immunogen is a synthetic peptide corresponding to residues N(21) E L E P L N R P Q L K(32) C of human ER alpha. This peptide sequence is completely conserved between human, rat, mouse, rabbit, sheep, porcine and bovine species. PA1-309 immunizing peptide (Cat. # PEP-037) is available for use in neutralization and control experiments. Recombinant ER alpha expressed in baculovirus system (Cat. # RP-310) can be purchased for control experiments in Western blot and gel shift assays.
- Reactivity
- Human, Rat, Porcine
- Host
- Rabbit
- Isotype
- IgG
- Vial size
- 50 μg
- Concentration
- 1 mg/mL
- Storage
- -20°C, Avoid Freeze/Thaw Cycles
Submitted references High levels estradiol affect blastocyst implantation and post-implantation development directly in mice.
CSNK1G2 differently sensitizes tamoxifen-induced decrease in PI3K/AKT/mTOR/S6K and ERK signaling according to the estrogen receptor existence in breast cancer cells.
Effects of the aryl hydrocarbon receptor agonist 3-methylcholanthrene on the 17β-estradiol regulated mRNA transcriptome of the rat uterus.
Amounts of an estrogen receptor β isoform increased in the theca of preovulatory follicles of sheep.
Expression of estrogen receptor alpha and beta in reactive astrocytes at the male rat hippocampus after status epilepticus.
Immunohistochemical localization of the estrogen receptor alpha (ERalpha) and progesterone receptor (PR) in the uterus of sika deer (Cervus nippon) during pregnancy.
ICAM-1 expression in vaginal cells as a potential biomarker for inflammatory response.
Nongenomic, endothelium-independent effects of estrogen on human coronary smooth muscle are mediated by type I (neuronal) NOS and PI3-kinase-Akt signaling.
Microvascular network remodeling in dura mater of ovariectomized pigs: role for angiopoietin-1 in estrogen-dependent control of vascular stability.
Estrogen receptor alpha mediates acute potassium channel stimulation in human coronary artery smooth muscle cells.
Estrogen receptors in the uterus and ovarian follicles of gilts treated with dihydrotestosterone.
Dose-dependent effects of 4-hydroxytamoxifen, the active metabolite of tamoxifen, on estrogen receptor-alpha expression in the rat uterus.
Cell cycle regulation of breast cancer cells through estrogen-induced activities of ERK and Akt protein kinases.
Attenuation of estrogenic effects by dihydrotestosterone in the pig uterus is associated with downregulation of the estrogen receptors.
Estrogen receptors alpha and beta in choroid plexus epithelial cells in Alzheimer's disease.
Exposure to the environmental estrogen bisphenol A differentially modulated estrogen receptor-alpha and -beta immunoreactivity and mRNA in male mouse testis.
Exposure to the environmental estrogen bisphenol A differentially modulated estrogen receptor-alpha and -beta immunoreactivity and mRNA in male mouse testis.
Strain differences in tamoxifen sensitivity of Sprague-Dawley and Fischer 344 rats.
Chang KT, Su YT, Tsai YR, Lan KC, Hsuuw YD, Kang HY, Chan WH, Huang FJ
Biomedical journal 2022 Feb;45(1):179-189
Biomedical journal 2022 Feb;45(1):179-189
CSNK1G2 differently sensitizes tamoxifen-induced decrease in PI3K/AKT/mTOR/S6K and ERK signaling according to the estrogen receptor existence in breast cancer cells.
Nguyen Hoang AT, Hoe KL, Lee SJ
PloS one 2021;16(4):e0246264
PloS one 2021;16(4):e0246264
Effects of the aryl hydrocarbon receptor agonist 3-methylcholanthrene on the 17β-estradiol regulated mRNA transcriptome of the rat uterus.
Helle J, Keiler AM, Zierau O, Dörfelt P, Vollmer G, Lehmann L, Chittur SV, Tenniswood M, Welsh J, Kretzschmar G
The Journal of steroid biochemistry and molecular biology 2017 Jul;171:133-143
The Journal of steroid biochemistry and molecular biology 2017 Jul;171:133-143
Amounts of an estrogen receptor β isoform increased in the theca of preovulatory follicles of sheep.
Cárdenas H, Pope WF
Animal reproduction science 2012 Apr;131(3-4):143-52
Animal reproduction science 2012 Apr;131(3-4):143-52
Expression of estrogen receptor alpha and beta in reactive astrocytes at the male rat hippocampus after status epilepticus.
Sakuma S, Tokuhara D, Hattori H, Matsuoka O, Yamano T
Neuropathology : official journal of the Japanese Society of Neuropathology 2009 Feb;29(1):55-62
Neuropathology : official journal of the Japanese Society of Neuropathology 2009 Feb;29(1):55-62
Immunohistochemical localization of the estrogen receptor alpha (ERalpha) and progesterone receptor (PR) in the uterus of sika deer (Cervus nippon) during pregnancy.
Yanagawa Y, Matsuura Y, Suzuki M, Katagiri S, Tsubota T
The Japanese journal of veterinary research 2008 Nov;56(3):139-49
The Japanese journal of veterinary research 2008 Nov;56(3):139-49
ICAM-1 expression in vaginal cells as a potential biomarker for inflammatory response.
Youn H, Hong K, Yoo JW, Lee CH
Biomarkers : biochemical indicators of exposure, response, and susceptibility to chemicals 2008 May;13(3):257-69
Biomarkers : biochemical indicators of exposure, response, and susceptibility to chemicals 2008 May;13(3):257-69
Nongenomic, endothelium-independent effects of estrogen on human coronary smooth muscle are mediated by type I (neuronal) NOS and PI3-kinase-Akt signaling.
Han G, Ma H, Chintala R, Miyake K, Fulton DJ, Barman SA, White RE
American journal of physiology. Heart and circulatory physiology 2007 Jul;293(1):H314-21
American journal of physiology. Heart and circulatory physiology 2007 Jul;293(1):H314-21
Microvascular network remodeling in dura mater of ovariectomized pigs: role for angiopoietin-1 in estrogen-dependent control of vascular stability.
Glinskii OV, Abraha TW, Turk JR, Rubin LJ, Huxley VH, Glinsky VV
American journal of physiology. Heart and circulatory physiology 2007 Aug;293(2):H1131-7
American journal of physiology. Heart and circulatory physiology 2007 Aug;293(2):H1131-7
Estrogen receptor alpha mediates acute potassium channel stimulation in human coronary artery smooth muscle cells.
Han G, Yu X, Lu L, Li S, Ma H, Zhu S, Cui X, White RE
The Journal of pharmacology and experimental therapeutics 2006 Mar;316(3):1025-30
The Journal of pharmacology and experimental therapeutics 2006 Mar;316(3):1025-30
Estrogen receptors in the uterus and ovarian follicles of gilts treated with dihydrotestosterone.
Cárdenas H, Pope WF
Domestic animal endocrinology 2005 Oct;29(3):523-33
Domestic animal endocrinology 2005 Oct;29(3):523-33
Dose-dependent effects of 4-hydroxytamoxifen, the active metabolite of tamoxifen, on estrogen receptor-alpha expression in the rat uterus.
Reed CA, Berndtson AK, Nephew KP
Anti-cancer drugs 2005 Jun;16(5):559-67
Anti-cancer drugs 2005 Jun;16(5):559-67
Cell cycle regulation of breast cancer cells through estrogen-induced activities of ERK and Akt protein kinases.
Geffroy N, Guédin A, Dacquet C, Lefebvre P
Molecular and cellular endocrinology 2005 Jun 15;237(1-2):11-23
Molecular and cellular endocrinology 2005 Jun 15;237(1-2):11-23
Attenuation of estrogenic effects by dihydrotestosterone in the pig uterus is associated with downregulation of the estrogen receptors.
Cárdenas H, Pope WF
Biology of reproduction 2004 Feb;70(2):297-302
Biology of reproduction 2004 Feb;70(2):297-302
Estrogen receptors alpha and beta in choroid plexus epithelial cells in Alzheimer's disease.
Hong-Goka BC, Chang FL
Neuroscience letters 2004 Apr 29;360(3):113-6
Neuroscience letters 2004 Apr 29;360(3):113-6
Exposure to the environmental estrogen bisphenol A differentially modulated estrogen receptor-alpha and -beta immunoreactivity and mRNA in male mouse testis.
Takao T, Nanamiya W, Nazarloo HP, Matsumoto R, Asaba K, Hashimoto K
Life sciences 2003 Jan 24;72(10):1159-69
Life sciences 2003 Jan 24;72(10):1159-69
Exposure to the environmental estrogen bisphenol A differentially modulated estrogen receptor-alpha and -beta immunoreactivity and mRNA in male mouse testis.
Takao T, Nanamiya W, Nazarloo HP, Matsumoto R, Asaba K, Hashimoto K
Life sciences 2003 Jan 24;72(10):1159-69
Life sciences 2003 Jan 24;72(10):1159-69
Strain differences in tamoxifen sensitivity of Sprague-Dawley and Fischer 344 rats.
Bailey JA, Nephew KP
Anti-cancer drugs 2002 Oct;13(9):939-47
Anti-cancer drugs 2002 Oct;13(9):939-47
No comments: Submit comment
Supportive validation
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
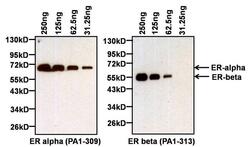
- Western blot analysis of Estrogen Receptor alpha (ER alpha) was performed by loading the indicated amounts of recombinant full-length human ER alpha (Product # A15674) and ER beta (Product # A15664) proteins, and 10 µL of PageRuler Prestained Protein Ladder (Product # 26616) per well onto a Novex® 4-20% Tris-Glycine polyacrylamide gel. Proteins were transferred to a PVDF membrane (Product # 88518) using the G2 Blotter (Product # 62288), and blocked with 5% milk in TBST for at least 1 hour at room temperature. ER alpha was detected at ~66 kD (left panel) using an ER alpha polyclonal antibody (Product # PA1-309) at a dilution of 1:1000 in blocking buffer overnight at 4C on a rocking platform, followed by an HRP-conjugated goat anti-rabbit IgG secondary antibody (Product # 31460) at a dilution of 1:40,000. Chemiluminescent detection was performed using SuperSignal West Pico (Product # 34080). The blot was then stripped with Restore Plus Western blot stripping buffer (Product # 46430) for at least 15 minutes at room temperature, re-blocked with 5% milk in TBST for at least 1 hour at room temperature, and re-probed with an ER beta polyclonal antibody (Product # PA1-313, right panel) at a dilution of 1:2000 overnight at 4C, followed by an HRP-conjugated goat anti-rabbit IgG secondary antibody (Product # 31460) at a dilution of 1:40,000. Chemiluminescent detection was again performed using SuperSignal West Pico (Product # 34080).
Supportive validation
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
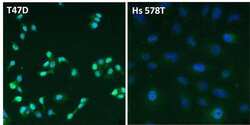
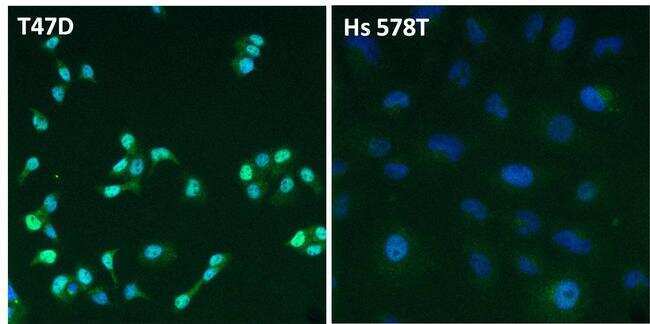
- Immunofluorescent analysis of Estrogen Receptor alpha (ER alpha, green) in T47D and Hs 578T breast cancer cells. The cells were fixed with 4% paraformaldehyde for 15 minutes at room temperature, permeabilized with 0.1% Triton X-100 for 15 minutes at room temperature, and blocked with 0.3% BSA in PBS for at least 30 minutes at room temperature. Cells were stained with an ER alpha polyclonal antibody (Product # PA1-309) at a dilution of 1:2000 in blocking buffer overnight at 4C, and then incubated with a DyLight 488-conjugated goat anti-rabbit IgG secondary antibody (Product # 35552) at a dilution of 1:500 for at least 1 hour at room temperature. Nuclei (blue) were stained with DAPI (Product # 62247). Images were taken on a Thermo Scientific ToxInsight Instrument at 20X magnification.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
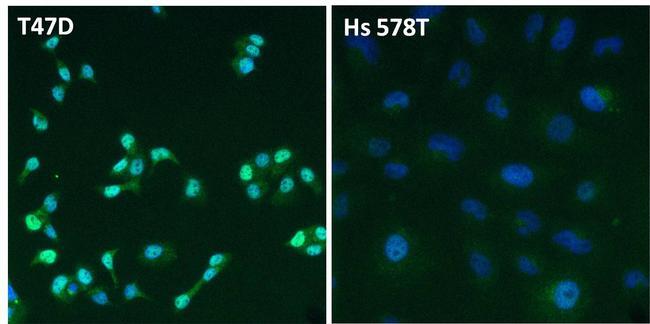
- Immunofluorescent analysis of Estrogen Receptor alpha (ER alpha, green) in T47D and Hs 578T breast cancer cells. The cells were fixed with 4% paraformaldehyde for 15 minutes at room temperature, permeabilized with 0.1% Triton X-100 for 15 minutes at room temperature, and blocked with 0.3% BSA in PBS for at least 30 minutes at room temperature. Cells were stained with an ER alpha polyclonal antibody (Product # PA1-309) at a dilution of 1:2000 in blocking buffer overnight at 4C, and then incubated with a DyLight 488-conjugated goat anti-rabbit IgG secondary antibody (Product # 35552) at a dilution of 1:500 for at least 1 hour at room temperature. Nuclei (blue) were stained with DAPI (Product # 62247). Images were taken on a Thermo Scientific ToxInsight Instrument at 20X magnification.
Supportive validation
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
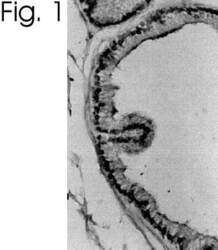
- Immunolocalization of ER alpha in rat prostate using Product # PA1-309.

Supportive validation
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
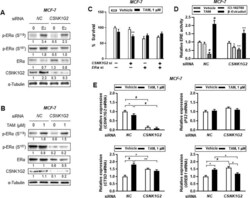
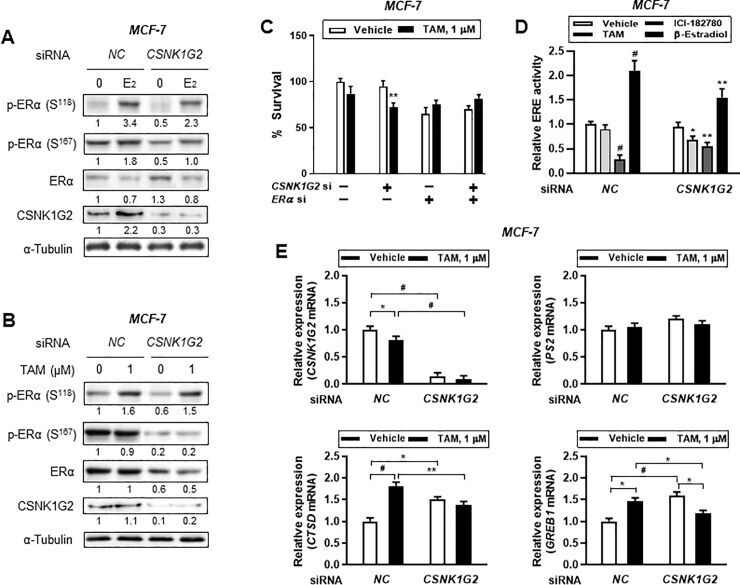
- Fig 4 Effect of CSNK1G2 on TAM-induced ER activity in ER + breast cancer cells. Immunoblots for phosphor-ER (p-ER) in CSNK1G2 siRNA-transfected breast cancer cells after treatment with (A) 10 nM E 2 or (B) 1 muM TAM for 24 h. Protein expression levels based on individual bands of phosphor form were compared to expression levels of alpha-tubulin and/or pan (total) protein. CSNK1G2 silencing efficiency was proved by western blotting for CSNK1G2. Each value under the blots indicates relative protein expression levels determined by densitometric analysis. (C) Cytotoxicity assay of CNSK1G2 -silenced ER + breast cancer cells. Cells transfected with different combinations of siRNA, such as NC siRNA alone, CSNK1G2 siRNA alone, ERalpha siRNA alone, or ERalpha plus CSNK1G2 siRNA, were treated with 1 muM TAM for 24 h; cellular toxicity was then measured using the MTT assay. (D) ERE-luciferase reporter assay showing a significant change in TAM-treated MCF-7 cells. CSNK1G2 knockdown by siRNA transfection in MCF-7 cells slightly reduced 10 nM estradiol, 1 muM ICI-182780, or 1 muM TAM-mediated ERalpha transcriptional activity. (E) Quantitative analysis for mRNA expression in MCF-7 cells. Knock-down cells with NC or CSNK1G2 siRNA were treated with vehicle or 1 muM of TAM for 24 h. Transcriptional patterns of diverse ER-responsive genes, PS2 , CTSD , and GREB1 , were analyzed by RT-qPCR in CSNK1G2 -silenced ER + breast cancer cells and were compared with those in NC siRNA-transfected control.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
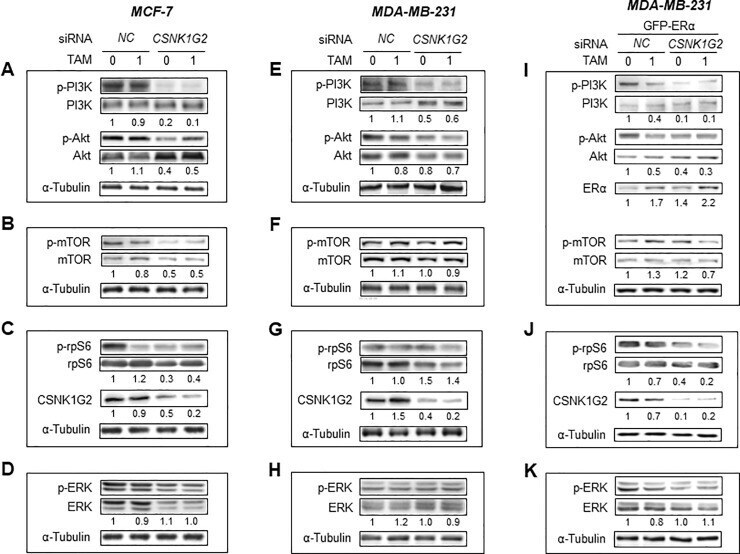
- Fig 5 CSNK1G2 effect on PI3K/AKT/mTOR/S6K and ERK signaling in human breast cancer cells. Immunoblots for PI3K/AKT/mTOR/S6K and ERK signaling-associated proteins in (A-D) MCF-7, ( E-H) MDA-MB-231, and (I-K) GFP-ERalpha-overexpressed MDA-MB-231 cells. Western blotting analysis from the breast cells transfected with control siRNA ( NC ) or CSNK1G2 siRNA ( CSNK1G2 ) were performed after treatment with vehicle or 1 muM TAM for 24 h. Protein expression levels based on individual bands of phosphor were compared to expression levels of alpha-tubulin and/or pan (total) protein. Each value under the blots indicates relative protein expression levels determined by densitometric analysis. Data are presented as mean (SD); n = 4.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
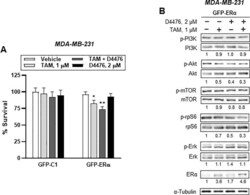
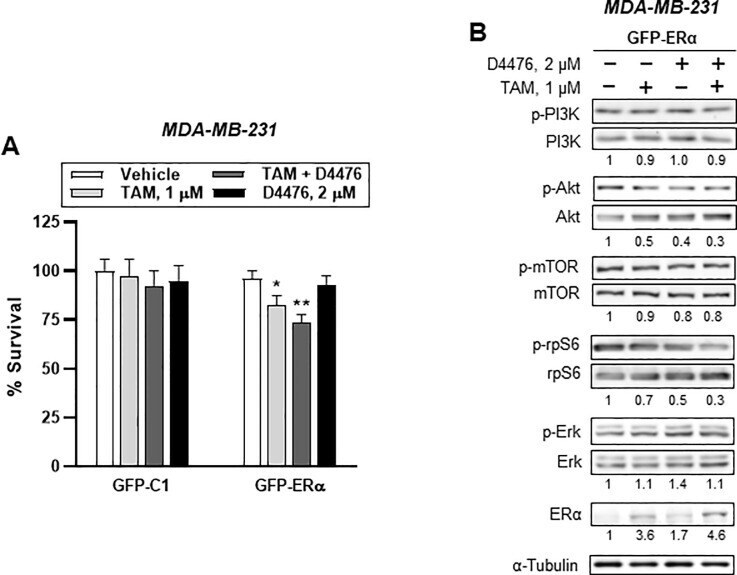
- Fig 9 GFP-ERalpha overexpression partly mimics PI3K/AKT/mTOR/S6K and ERK signaling in ER - breast cancer cells. (A) Cytotoxicity assay of either GFP-C1 or GFP-ERalpha-transfected ER - breast cancer cells. Bar denote relative percentage of % survival of transfected MDA-MB-231 cells after treatment with vehicle, 1 muM TAM, 2 muM D4476, and 1 muM TAM plus 2 muM D4476 for 24h. Cellular toxicity was measured using the MTT assay (mean (SD), n = 5; * P < 0.05, ** P < 0.01 vs . each represented counterpart). (B) Western blotting analysis from GFP-ERalpha-transfected ER - breast cells treated with vehicle or 2 muM D4476 together with 1 muM TAM for 24 h were performed. Immunoblots for PI3K/AKT/mTOR/S6K signaling-associated proteins in ER - MDA-MB-231 cells. Protein expression levels based on individual bands of phosphor were compared to expression levels of alpha-tubulin and/or pan (total) protein. Each value under the blots indicates relative protein expression levels determined by densitometric analysis (mean +- SEM, n = 4).
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
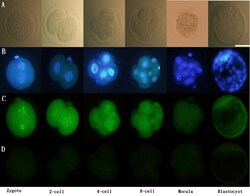
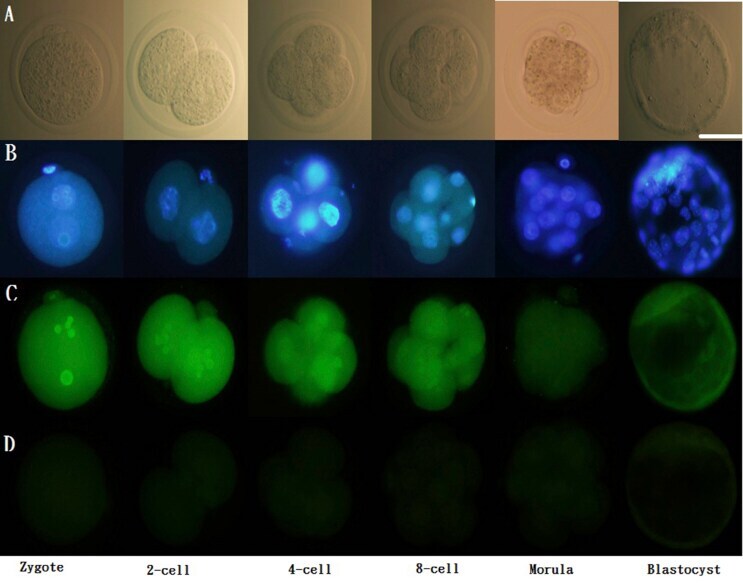
- Immunofluorescence of ERalpha in pre-implantation embryos. Scattered ERalpha expression was detected in the cytoplasm as well as the nucleus (co-stained with DAPI) of 1-cell stage embryos to blastocysts. ERalpha: green; DAPI: blue. Scale bar = 50 mum (A) Unstained; (B) stained with DAPI; (C) stained with primary and secondary antibodies for ERalpha; (D) stained with only secondary antibodies for ER alpha. Representative images from three experiments using biologically different samples (n >= 15) are shown. Fig. 1
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
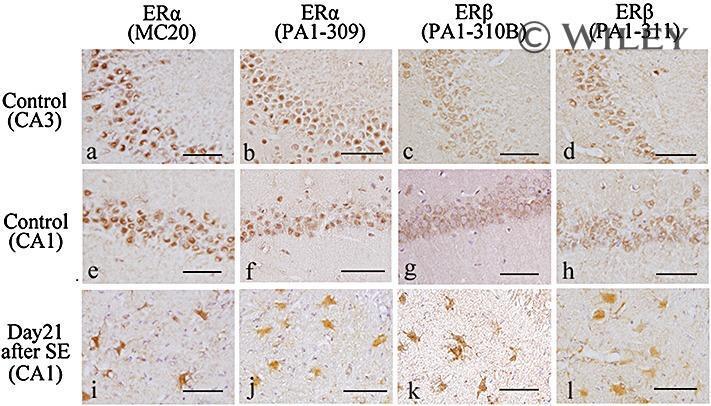
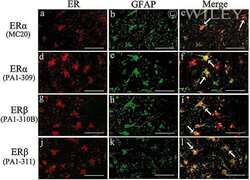
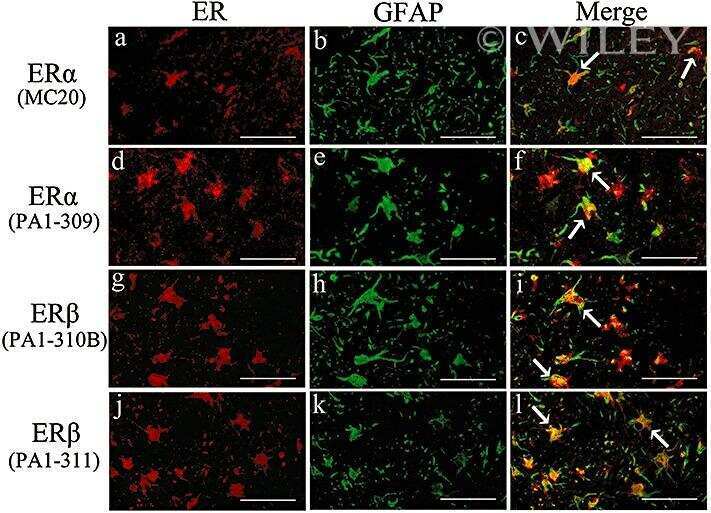
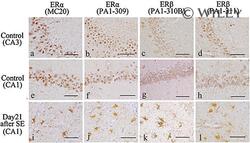
- 2 Distribution of estrogen receptor (ER)alpha and ERbeta immunoreactivity in the adult male rat hippocampus. (a,e) ERalpha immunoreactivity in the control rat hippocampus by anti-ERalpha antibody (MC-20): (a) CA3 region. ERalpha immunoreactivity was observed predominantly in the nucleus and to a lesser extent in the cytoplasm; (e) CA1 region. ERalpha was expressed predominantly in the nucleus and to a lesser extent in the cytoplasm. (b,f) ERalpha immunoreactivity in the control rat hippocampus by anti-ERalpha antibody (PA1-309): (b) Image of the CA3 region showing ERalpha-immunoreactivity predominantly in the nucleus and to a lesser extent in the cytoplasm; (f) Image of the CA1 region showing ERalpha immunoreactivity predominantly in the nucleus and to a lesser extent in the cytoplasm; (c,g) CA3 (c) and CA1 (g) of the control rats stained immunohistochemically with anti-ERbeta antibody (PA1-310B): (c) Image of the CA3 region of a showing ERbeta immunoreactivity mainly in the cytoplasm; (g) Image of the CA1 region. ERbeta was expressed in the cytoplasm. (d,h) CA3 (d) and CA1 (h) of control rats stained immunohistochemically with anti-ERbeta antibody (PA1-311): (d) Image of the CA3 region showing ERbeta immunoreactivity mainly in the cytoplasm; (h) Image of the CA1 region. ERbeta was expressed in the cytoplasm. (i-l) Images of the CA1 region with neuronal loss in the rats on day 21 after status epilepticus stained immunohistochemically with anti-ERalpha antibody (MC-20),