Explore
Explore Validate
Validate Learn
Learn Western blot
Western blot Immunocytochemistry
Immunocytochemistry Immunoprecipitation
ImmunoprecipitationAntibody data
- Antibody Data
- Antigen structure
- References [31]
- Comments [0]
- Validations
- Immunocytochemistry [6]
- Immunohistochemistry [1]
- Other assay [9]
Submit
Validation data
Reference
Comment
Report error
- Product number
- MA1-516 - Provider product page

- Provider
- Invitrogen Antibodies
- Product name
- HIF1A Monoclonal Antibody (mgc3)
- Antibody type
- Monoclonal
- Antigen
- Other
- Description
- MA1-516 detects hypoxia-inducible factor 1 alpha (HIF-1 alpha) from human, non-human primate, bovine, mouse and porcine cells. This antibody does not cross-react with ARNT or the related HIF-2 alpha. MA1-516 has been successfully used in Western blot, immunofluorescence, immunoprecipitation, immunocytochemistry, and gel shift procedures. By Western blot, this antibody detects an ~116 kDa protein representing HIF-1 alpha after hypoxic induction in COS cells. Immunofluorescence staining of HIF-1 alpha in COS-7 cells with MA1-516 yields nuclear staining after exposing cells to 1% oxygen for 4 hours. In gel shift assays, MA1-516 has been successfully used only with mouse HIF-1 alpha. Antibodies to this protein (and modification) were previously sold as part of a Thermo Scientific Cellomics High Content Screening Kit. This replacement antibody is now recommended for researchers who need an antibody for high content cell based assays. It has been thoroughly tested and validated for cellular immunofluorescence (IF) applications. Further optimization including the selection of the most appropriate fluorescent Dylight conjugated secondary antibody may have to be performed for your high content assay.
- Reactivity
- Human, Mouse, Bovine, Porcine
- Host
- Mouse
- Isotype
- IgG
- Antibody clone number
- mgc3
- Vial size
- 100 μL
- Concentration
- 1 mg/mL
- Storage
- -20°C, Avoid Freeze/Thaw Cycles
Submitted references Physiological Effects of Intermittent Passive Exposure to Hypobaric Hypoxia and Cold in Rats.
The HIF1α/JMY pathway promotes glioblastoma stem-like cell invasiveness after irradiation.
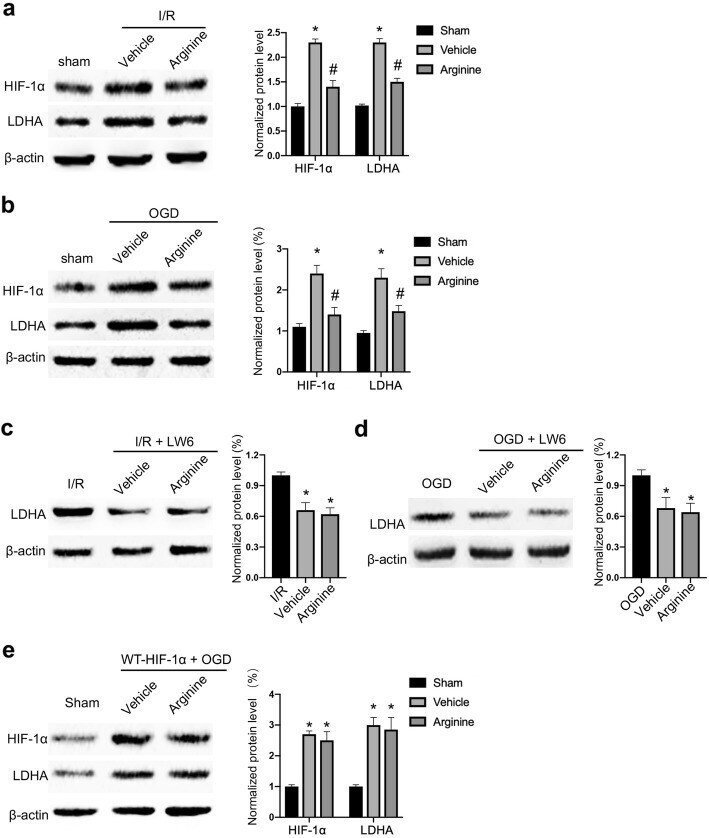
Arginine is neuroprotective through suppressing HIF-1α/LDHA-mediated inflammatory response after cerebral ischemia/reperfusion injury.
Shmt2: A Stat3 Signaling New Player in Prostate Cancer Energy Metabolism.
Molecular imaging HDACs class IIa expression-activity and pharmacologic inhibition in intracerebral glioma models in rats using PET/CT/(MRI) with [(18)F]TFAHA.
An Integrated Score and Nomogram Combining Clinical and Immunohistochemistry Factors to Predict High ISUP Grade Clear Cell Renal Cell Carcinoma.
Mixing and delivery of multiple controlled oxygen environments to a single multiwell culture plate.
STAT3, a Hub Protein of Cellular Signaling Pathways, Is Triggered by β-Hexaclorocyclohexane.
Innate immune memory in the brain shapes neurological disease hallmarks.
Identification of cardiac progenitors that survive in the ischemic human heart after ventricular myocyte death.
pVHL-mediated degradation of HIF-2α regulates estrogen receptor α expression in normoxic breast cancer cells.
A common functional regulatory variant at a type 2 diabetes locus upregulates ARAP1 expression in the pancreatic beta cell.
STUB1/CHIP is required for HIF1A degradation by chaperone-mediated autophagy.
Human hair follicle stem/progenitor cells express hypoxia markers.
Hypoxia-inducible factor and vascular endothelial growth factor are targets of dietary soy during acute stroke in female rats.
Hypoxia-inducible factor-1β (HIF-1β) is upregulated in a HIF-1α-dependent manner in 518A2 human melanoma cells under hypoxic conditions.
Disrupting galectin-1 interactions with N-glycans suppresses hypoxia-driven angiogenesis and tumorigenesis in Kaposi's sarcoma.
Soluble FLT1 binds lipid microdomains in podocytes to control cell morphology and glomerular barrier function.
Up-regulation of glyceraldehyde-3-phosphate dehydrogenase gene expression by HIF-1 activity depending on Sp1 in hypoxic breast cancer cells.
Hypoxia enhances transcriptional activity of androgen receptor through hypoxia-inducible factor-1α in a low androgen environment.
Hypoxia preconditioning protection of corneal stromal cells requires HIF1alpha but not VEGF.
Human placental hypoxia-inducible factor-1alpha expression correlates with clinical outcomes in chronic hypoxia in vivo.
Hypoxia upregulates carcinoembryonic antigen expression in cancer cells.
Hypoxia preconditioning protects corneal stromal cells against induced apoptosis.
Dynamic HIF1A regulation during human placental development.
Role of oxygen and vascular development in epithelial branching morphogenesis of the developing mouse lung.
Experimental angiogenesis of arterial vasa vasorum.
Angiogenic potential of prostate carcinoma cells overexpressing bcl-2.
Epolones induce erythropoietin expression via hypoxia-inducible factor-1 alpha activation.
General applicability of chicken egg yolk antibodies: the performance of IgY immunoglobulins raised against the hypoxia-inducible factor 1alpha.
Oxygen-regulated erythropoietin gene expression is dependent on a CpG methylation-free hypoxia-inducible factor-1 DNA-binding site.
Santocildes G, Viscor G, Pagès T, Ramos-Romero S, Torres JL, Torrella JR
Frontiers in physiology 2021;12:673095
Frontiers in physiology 2021;12:673095
The HIF1α/JMY pathway promotes glioblastoma stem-like cell invasiveness after irradiation.
Gauthier LR, Saati M, Bensalah-Pigeon H, Ben M'Barek K, Gitton-Quent O, Bertrand R, Busso D, Mouthon MA, Collura A, Junier MP, Chneiweiss H, Pineda JR, Boussin FD
Scientific reports 2020 Oct 30;10(1):18742
Scientific reports 2020 Oct 30;10(1):18742
Arginine is neuroprotective through suppressing HIF-1α/LDHA-mediated inflammatory response after cerebral ischemia/reperfusion injury.
Chen SF, Pan MX, Tang JC, Cheng J, Zhao D, Zhang Y, Liao HB, Liu R, Zhuang Y, Zhang ZF, Chen J, Lei RX, Li SF, Li HT, Wang ZF, Wan Q
Molecular brain 2020 Apr 22;13(1):63
Molecular brain 2020 Apr 22;13(1):63
Shmt2: A Stat3 Signaling New Player in Prostate Cancer Energy Metabolism.
Marrocco I, Altieri F, Rubini E, Paglia G, Chichiarelli S, Giamogante F, Macone A, Perugia G, Magliocca FM, Gurtner A, Maras B, Ragno R, Patsilinakos A, Manganaro R, Eufemi M
Cells 2019 Sep 6;8(9)
Cells 2019 Sep 6;8(9)
Molecular imaging HDACs class IIa expression-activity and pharmacologic inhibition in intracerebral glioma models in rats using PET/CT/(MRI) with [(18)F]TFAHA.
Laws MT, Bonomi RE, Kamal S, Gelovani DJ, Llaniguez J, Potukutchi S, Lu X, Mangner T, Gelovani JG
Scientific reports 2019 Mar 5;9(1):3595
Scientific reports 2019 Mar 5;9(1):3595
An Integrated Score and Nomogram Combining Clinical and Immunohistochemistry Factors to Predict High ISUP Grade Clear Cell Renal Cell Carcinoma.
Wu J, Xu WH, Wei Y, Qu YY, Zhang HL, Ye DW
Frontiers in oncology 2018;8:634
Frontiers in oncology 2018;8:634
Mixing and delivery of multiple controlled oxygen environments to a single multiwell culture plate.
Yao M, Sattler T, Rabbani ZN, Pulliam T, Walker G, Gamcsik MP
American journal of physiology. Cell physiology 2018 Nov 1;315(5):C766-C775
American journal of physiology. Cell physiology 2018 Nov 1;315(5):C766-C775
STAT3, a Hub Protein of Cellular Signaling Pathways, Is Triggered by β-Hexaclorocyclohexane.
Rubini E, Altieri F, Chichiarelli S, Giamogante F, Carissimi S, Paglia G, Macone A, Eufemi M
International journal of molecular sciences 2018 Jul 20;19(7)
International journal of molecular sciences 2018 Jul 20;19(7)
Innate immune memory in the brain shapes neurological disease hallmarks.
Wendeln AC, Degenhardt K, Kaurani L, Gertig M, Ulas T, Jain G, Wagner J, Häsler LM, Wild K, Skodras A, Blank T, Staszewski O, Datta M, Centeno TP, Capece V, Islam MR, Kerimoglu C, Staufenbiel M, Schultze JL, Beyer M, Prinz M, Jucker M, Fischer A, Neher JJ
Nature 2018 Apr;556(7701):332-338
Nature 2018 Apr;556(7701):332-338
Identification of cardiac progenitors that survive in the ischemic human heart after ventricular myocyte death.
Omatsu-Kanbe M, Nozuchi N, Nishino Y, Mukaisho KI, Sugihara H, Matsuura H
Scientific reports 2017 Jan 25;7:41318
Scientific reports 2017 Jan 25;7:41318
pVHL-mediated degradation of HIF-2α regulates estrogen receptor α expression in normoxic breast cancer cells.
Higashimura Y, Kitakaze T, Harada N, Inui H, Nakano Y, Yamaji R
FEBS letters 2016 Aug;590(16):2690-9
FEBS letters 2016 Aug;590(16):2690-9
A common functional regulatory variant at a type 2 diabetes locus upregulates ARAP1 expression in the pancreatic beta cell.
Kulzer JR, Stitzel ML, Morken MA, Huyghe JR, Fuchsberger C, Kuusisto J, Laakso M, Boehnke M, Collins FS, Mohlke KL
American journal of human genetics 2014 Feb 6;94(2):186-97
American journal of human genetics 2014 Feb 6;94(2):186-97
STUB1/CHIP is required for HIF1A degradation by chaperone-mediated autophagy.
Ferreira JV, Fôfo H, Bejarano E, Bento CF, Ramalho JS, Girão H, Pereira P
Autophagy 2013 Sep;9(9):1349-66
Autophagy 2013 Sep;9(9):1349-66
Human hair follicle stem/progenitor cells express hypoxia markers.
Rathman-Josserand M, Genty G, Lecardonnel J, Chabane S, Cousson A, François Michelet J, Bernard BA
The Journal of investigative dermatology 2013 Aug;133(8):2094-7
The Journal of investigative dermatology 2013 Aug;133(8):2094-7
Hypoxia-inducible factor and vascular endothelial growth factor are targets of dietary soy during acute stroke in female rats.
Ma Y, Lovekamp-Swan T, Bekele W, Dohi A, Schreihofer DA
Endocrinology 2013 Apr;154(4):1589-97
Endocrinology 2013 Apr;154(4):1589-97
Hypoxia-inducible factor-1β (HIF-1β) is upregulated in a HIF-1α-dependent manner in 518A2 human melanoma cells under hypoxic conditions.
Mandl M, Kapeller B, Lieber R, Macfelda K
Biochemical and biophysical research communications 2013 Apr 26;434(1):166-72
Biochemical and biophysical research communications 2013 Apr 26;434(1):166-72
Disrupting galectin-1 interactions with N-glycans suppresses hypoxia-driven angiogenesis and tumorigenesis in Kaposi's sarcoma.
Croci DO, Salatino M, Rubinstein N, Cerliani JP, Cavallin LE, Leung HJ, Ouyang J, Ilarregui JM, Toscano MA, Domaica CI, Croci MC, Shipp MA, Mesri EA, Albini A, Rabinovich GA
The Journal of experimental medicine 2012 Oct 22;209(11):1985-2000
The Journal of experimental medicine 2012 Oct 22;209(11):1985-2000
Soluble FLT1 binds lipid microdomains in podocytes to control cell morphology and glomerular barrier function.
Jin J, Sison K, Li C, Tian R, Wnuk M, Sung HK, Jeansson M, Zhang C, Tucholska M, Jones N, Kerjaschki D, Shibuya M, Fantus IG, Nagy A, Gerber HP, Ferrara N, Pawson T, Quaggin SE
Cell 2012 Oct 12;151(2):384-99
Cell 2012 Oct 12;151(2):384-99
Up-regulation of glyceraldehyde-3-phosphate dehydrogenase gene expression by HIF-1 activity depending on Sp1 in hypoxic breast cancer cells.
Higashimura Y, Nakajima Y, Yamaji R, Harada N, Shibasaki F, Nakano Y, Inui H
Archives of biochemistry and biophysics 2011 May 1;509(1):1-8
Archives of biochemistry and biophysics 2011 May 1;509(1):1-8
Hypoxia enhances transcriptional activity of androgen receptor through hypoxia-inducible factor-1α in a low androgen environment.
Mitani T, Yamaji R, Higashimura Y, Harada N, Nakano Y, Inui H
The Journal of steroid biochemistry and molecular biology 2011 Jan;123(1-2):58-64
The Journal of steroid biochemistry and molecular biology 2011 Jan;123(1-2):58-64
Hypoxia preconditioning protection of corneal stromal cells requires HIF1alpha but not VEGF.
Xing D, Bonanno JA
Molecular vision 2009 May 18;15:1020-7
Molecular vision 2009 May 18;15:1020-7
Human placental hypoxia-inducible factor-1alpha expression correlates with clinical outcomes in chronic hypoxia in vivo.
Zamudio S, Wu Y, Ietta F, Rolfo A, Cross A, Wheeler T, Post M, Illsley NP, Caniggia I
The American journal of pathology 2007 Jun;170(6):2171-9
The American journal of pathology 2007 Jun;170(6):2171-9
Hypoxia upregulates carcinoembryonic antigen expression in cancer cells.
Kokkonen N, Ulibarri IF, Kauppila A, Luosujärvi H, Rivinoja A, Pospiech H, Kellokumpu I, Kellokumpu S
International journal of cancer 2007 Dec 1;121(11):2443-50
International journal of cancer 2007 Dec 1;121(11):2443-50
Hypoxia preconditioning protects corneal stromal cells against induced apoptosis.
Xing D, Sun X, Li J, Cui M, Tan-Allen K, Bonanno JA
Experimental eye research 2006 May;82(5):780-7
Experimental eye research 2006 May;82(5):780-7
Dynamic HIF1A regulation during human placental development.
Ietta F, Wu Y, Winter J, Xu J, Wang J, Post M, Caniggia I
Biology of reproduction 2006 Jul;75(1):112-21
Biology of reproduction 2006 Jul;75(1):112-21
Role of oxygen and vascular development in epithelial branching morphogenesis of the developing mouse lung.
van Tuyl M, Liu J, Wang J, Kuliszewski M, Tibboel D, Post M
American journal of physiology. Lung cellular and molecular physiology 2005 Jan;288(1):L167-78
American journal of physiology. Lung cellular and molecular physiology 2005 Jan;288(1):L167-78
Experimental angiogenesis of arterial vasa vasorum.
Bayer IM, Caniggia I, Adamson SL, Langille BL
Cell and tissue research 2002 Mar;307(3):303-13
Cell and tissue research 2002 Mar;307(3):303-13
Angiogenic potential of prostate carcinoma cells overexpressing bcl-2.
Fernandez A, Udagawa T, Schwesinger C, Beecken W, Achilles-Gerte E, McDonnell T, D'Amato R
Journal of the National Cancer Institute 2001 Feb 7;93(3):208-13
Journal of the National Cancer Institute 2001 Feb 7;93(3):208-13
Epolones induce erythropoietin expression via hypoxia-inducible factor-1 alpha activation.
Wanner RM, Spielmann P, Stroka DM, Camenisch G, Camenisch I, Scheid A, Houck DR, Bauer C, Gassmann M, Wenger RH
Blood 2000 Aug 15;96(4):1558-65
Blood 2000 Aug 15;96(4):1558-65
General applicability of chicken egg yolk antibodies: the performance of IgY immunoglobulins raised against the hypoxia-inducible factor 1alpha.
Camenisch G, Tini M, Chilov D, Kvietikova I, Srinivas V, Caro J, Spielmann P, Wenger RH, Gassmann M
FASEB journal : official publication of the Federation of American Societies for Experimental Biology 1999 Jan;13(1):81-8
FASEB journal : official publication of the Federation of American Societies for Experimental Biology 1999 Jan;13(1):81-8
Oxygen-regulated erythropoietin gene expression is dependent on a CpG methylation-free hypoxia-inducible factor-1 DNA-binding site.
Wenger RH, Kvietikova I, Rolfs A, Camenisch G, Gassmann M
European journal of biochemistry 1998 May 1;253(3):771-7
European journal of biochemistry 1998 May 1;253(3):771-7
No comments: Submit comment
Supportive validation
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
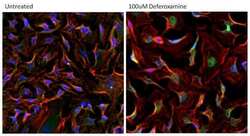
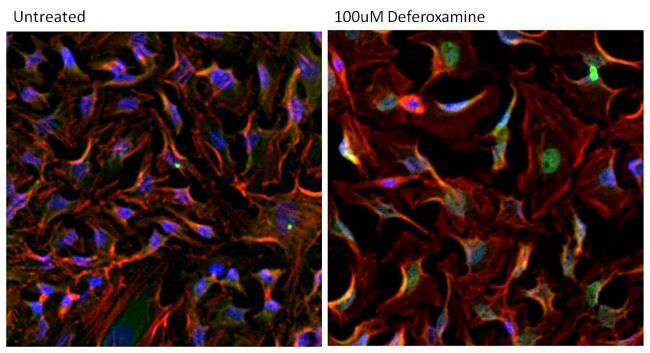
- Immunofluorescent analysis of HIF-1 alpha (green) in HeLa cells either left untreated (left panel) or treated with 100uM Deferoxamine mesylate for ~16 hours (right panel). Formalin-fixed cells were permeabilized with 0.1% Triton X-100 in TBS for 15 minutes at room temperature and blocked with 0.3% BSA for 15 minutes at room temperature. Cells were probed with a HIF-1 alpha monoclonal antibody (Product # MA1-516) at a dilution of 1:100 for at least 1 hour at room temperature, washed with PBS, and incubated with a DyLight 488-conjugated goat anti-mouse IgG secondary antibody (Product # 35502) at a dilution of 1:500 for 30 minutes at room temperature. F-actin (red) was stained with DyLight 594 Phalloidin (Product # 21836) and nuclei (blue) were stained with Hoechst 33342 dye (Product # 62249). Images were taken on a Thermo Scientific ArrayScan Instrument at 20X magnification.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
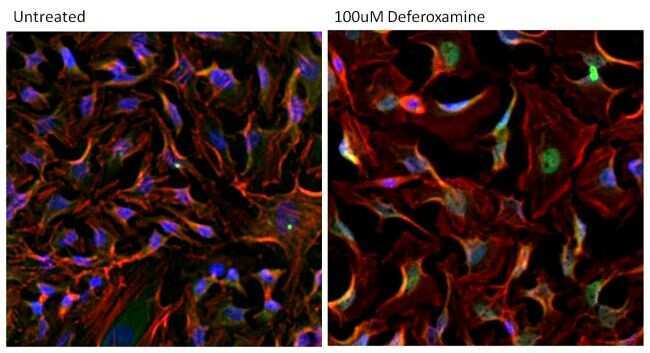
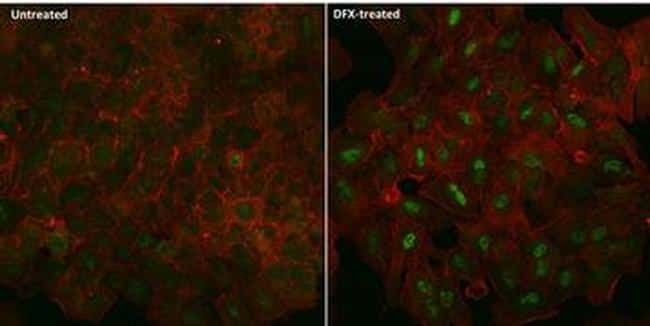
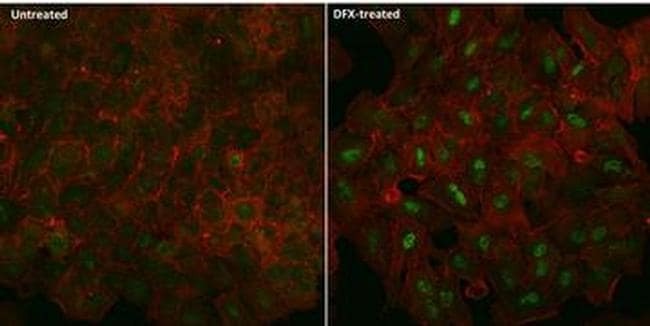
- Immunofluorescent analysis of HIF-1 alpha (green) in A549 cells either left untreated (left panel) or treated with 100uM desferrioxamine (DFX) for ~16 hours (right panel). The cells were fixed with 4% Paraformaldehyde for 15 min at room temp, permeabilized with 0.1% Triton X-100 in TBS for 15 minutes at room temperature and blocked with 0.3% BSA for 15 minutes at room temperature. Cells were probed with a HIF-1 alpha monoclonal antibody (Product # MA1-516) at a dilution of 1:100 overnight at 4C, washed with PBS, and incubated with an Alexa Fluor 488-conjugated goat anti-mouse IgG secondary antibody (Product # R37120) at a dilution of 1:500 for 2 hours at room temperature. F-actin (red) was stained with Alexa Fluor 647 Phalloidin (Product # A22287) and nuclei (blue) were stained with DAPI (Product # P-36931). Images were taken on a Zeiss LSM 710 confocal microscope.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
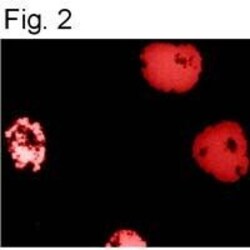
- Immunofluorescence staining of HIF-1 Alpha using Product # MA1-516.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
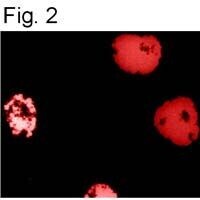
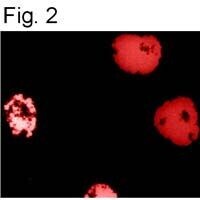
- Immunofluorescence staining of HIF-1 Alpha using Product # MA1-516.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
- Immunofluorescent analysis of HIF-1 alpha (green) in HeLa cells either left untreated (left panel) or treated with 100uM Deferoxamine mesylate for ~16 hours (right panel). Formalin-fixed cells were permeabilized with 0.1% Triton X-100 in TBS for 15 minutes at room temperature and blocked with 0.3% BSA for 15 minutes at room temperature. Cells were probed with a HIF-1 alpha monoclonal antibody (Product # MA1-516) at a dilution of 1:100 for at least 1 hour at room temperature, washed with PBS, and incubated with a DyLight 488-conjugated goat anti-mouse IgG secondary antibody (Product # 35502) at a dilution of 1:500 for 30 minutes at room temperature. F-actin (red) was stained with DyLight 594 Phalloidin (Product # 21836) and nuclei (blue) were stained with Hoechst 33342 dye (Product # 62249). Images were taken on a Thermo Scientific ArrayScan Instrument at 20X magnification.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
- Immunofluorescent analysis of HIF-1 alpha (green) in A549 cells either left untreated (left panel) or treated with 100uM desferrioxamine (DFX) for ~16 hours (right panel). The cells were fixed with 4% Paraformaldehyde for 15 min at room temp, permeabilized with 0.1% Triton X-100 in TBS for 15 minutes at room temperature and blocked with 0.3% BSA for 15 minutes at room temperature. Cells were probed with a HIF-1 alpha monoclonal antibody (Product # MA1-516) at a dilution of 1:100 overnight at 4C, washed with PBS, and incubated with an Alexa Fluor 488-conjugated goat anti-mouse IgG secondary antibody (Product # R37120) at a dilution of 1:500 for 2 hours at room temperature. F-actin (red) was stained with Alexa Fluor 647 Phalloidin (Product # A22287) and nuclei (blue) were stained with DAPI (Product # P-36931). Images were taken on a Zeiss LSM 710 confocal microscope.
Supportive validation
- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
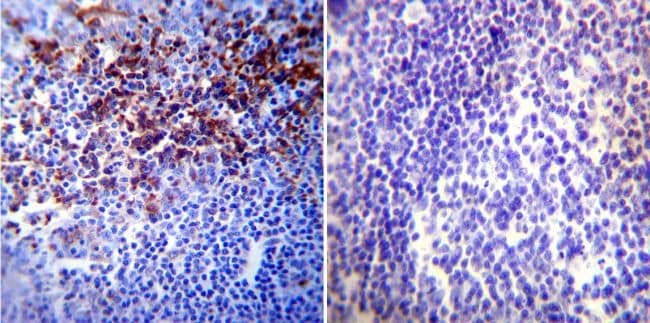
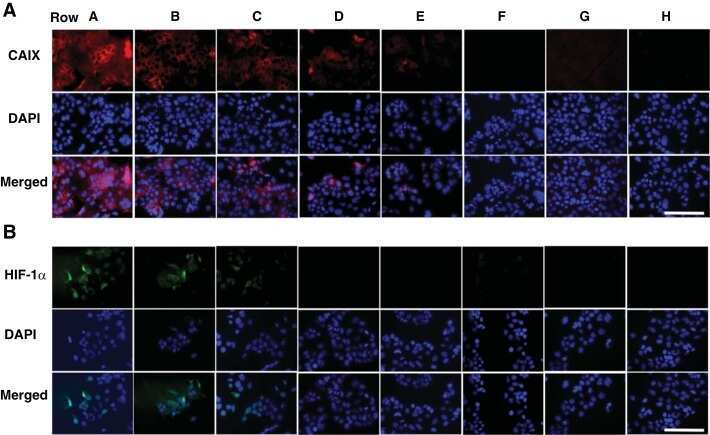
- Immunohistochemistry was performed on normal biopsies of deparaffinized Human tonsil tissue. To expose target proteins, heat induced antigen retrieval was performed using 10mM sodium citrate (pH6.0) buffer, microwaved for 8-15 minutes. Following antigen retrieval tissues were blocked in 3% BSA-PBS for 30 minutes at room temperature. Tissues were then probed at a dilution of 1:20 with a mouse monoclonal antibody recognizing HIF-1 alpha (Product # MA1-516) or without primary antibody (negative control) overnight at 4°C in a humidified chamber. Tissues were washed extensively with PBST and endogenous peroxidase activity was quenched with a peroxidase suppressor. Detection was performed using a biotin-conjugated secondary antibody and SA-HRP, followed by colorimetric detection using DAB. Tissues were counterstained with hematoxylin and prepped for mounting.
Supportive validation
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
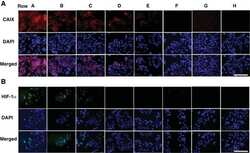
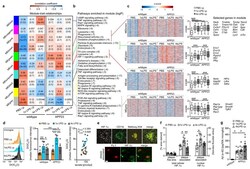
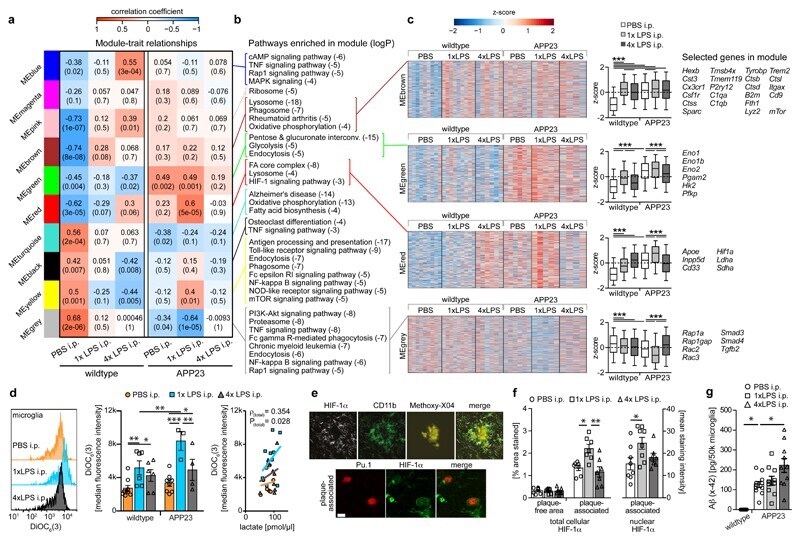
- Figure 5 Microglial gene expression and function 6 months after immune stimulation. a, Weighted gene correlation network analysis (top: correlation coefficient; bottom: P-value; n=9,9,6,6,5,4 animals). b , Selected KEGG pathways enriched in modules. c , Heatmaps of genes within modules, z-scores (boxplot whiskers: 5-95 th percentile; n=1601,990,949,3543 genes in modules) and selected genes. d , Microglial mitochondrial membrane potential (left/middle; n=9,6,6,8,3,4 animals) and Pearson's correlation with lactate release (right; n=11,10,10 animals). e , Staining for top : HIF-1alpha, microglia (CD11b) and amyloid plaques (Methoxy-X04) and bottom : HIF-1alpha and microglial nuclei (Pu.1; single confocal plane) in brain sections from 9-month-old animals. Scale bars: 20/5 um (top/bottom). f , Total cellular (n=7,7,7 animals) and nuclear (n=8,8,7 animals) HIF-1alpha staining intensity. g , Microglial Abeta content (n=5,11,10,10 animals). */**/*** P
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
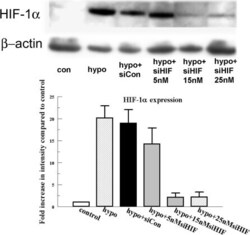
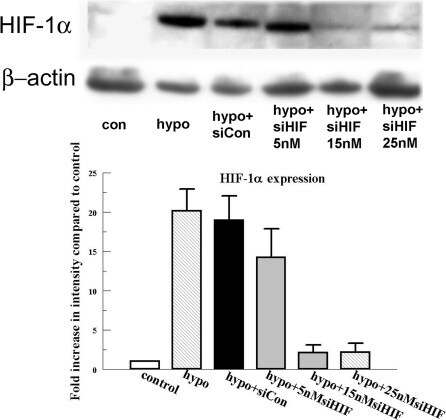
- Figure 2 Effect of HIF-1alpha siRNA on HIF-1alpha protein expression. Cells were transfected with 15 and 25 nM HIF-1alpha siRNA and an siRNA non targeting control as indicated for 6 h. Twenty-four hours after transfection, cells were exposed to 4 h of hypoxia. Protein was collected immediately after treatment and blotted for HIF-1alpha. Bar graph represents the band intensity relative to the control. Error bars represent the standard error of the mean (n=3). beta-actin is detected as loading control.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
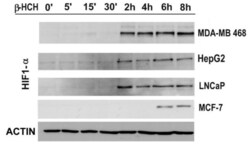
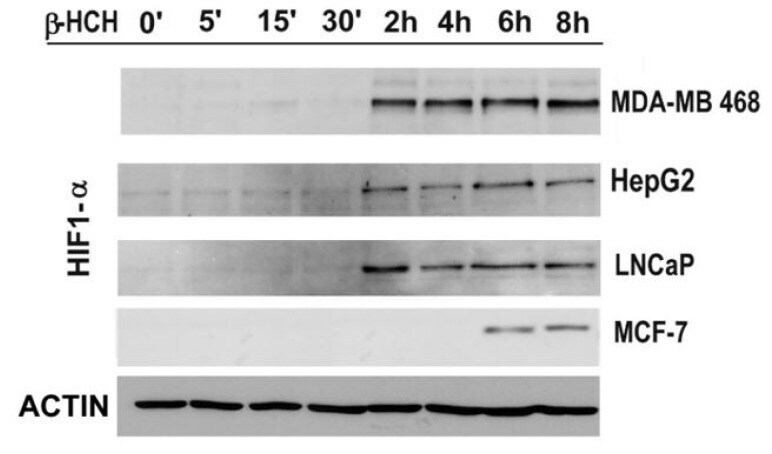
- Figure 6 The expression levels of HIF-1alpha were detected by immunoblot analysis in a time-course experiment performed on MDA-MB 468, HepG2, LNCaP, and MCF-7 cells treated with beta-HCH (10 muM). Actin was used as normalization protein.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
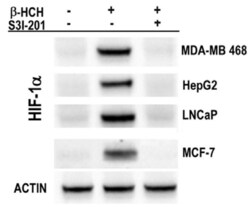
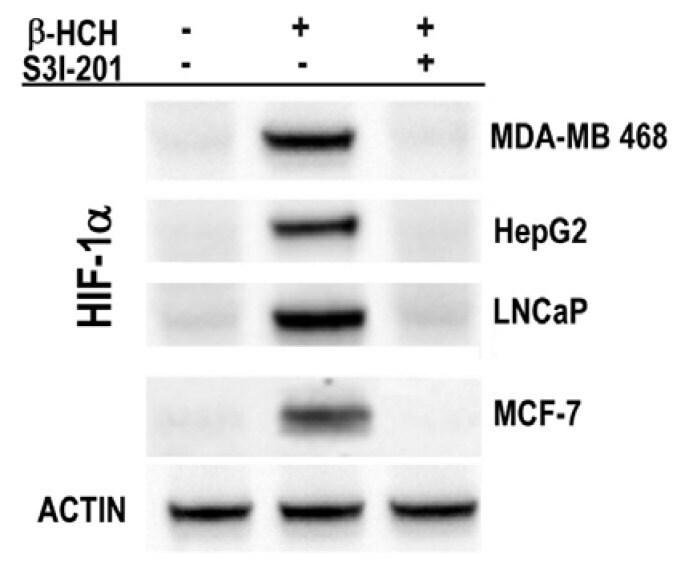
- Figure 7 Immunoblot analysis of HIF-1alpha expression in MDA-MB 468, HepG2, LNCaP, and MCF-7 cells after 8 h treatment with 10 muM beta-HCH. Analysis was performed after 8 h treatment with 10 muM beta-HCH, as well as on cells pre-incubated with a specific STAT3 inhibitor (S3I-201) before beta-HCH treatment. Actin was used as normalization protein.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
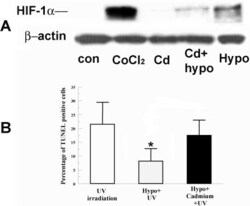
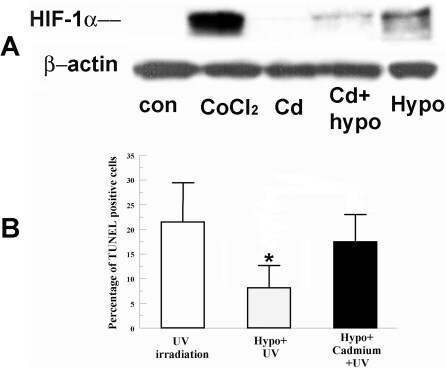
- Figure 1 Effect of Cadmium on HIF-1alpha accumulation and hypoxia preconditioning protection. A : Cells were incubated under normoxia with 200 muM CoCl 2 (as a positive control), 5 muM CdCl 2 alone, hypoxia (0.5% O 2 ) alone, or hypoxia with CdCl 2 for 4 h. Whole cell lysates were collected immediately after treatment, separated on SDS-PAGE gel and blotted for HIF-1alpha. beta-actin was detected as a loading control. B : Cells were pretreated with hypoxia with or without CdCl 2 for 4 h and irradiated 2 min with UV. Four hours after irradiation, cells were fixed and stained for TUNEL. Bar graph shows percentage of TUNEL positive cells in indicated groups. Error bars represent standard error of the mean (n=3); the asterisk indicates significantly different from UV control (p
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
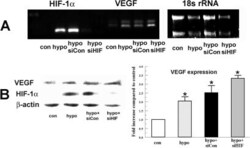
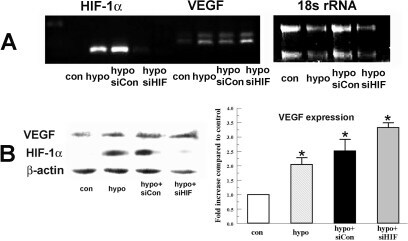
- Figure 5 Effect of HIF-1alpha siRNA on hypoxia-induced VEGF expression. Bovine stromal cells were transfected with HIF-1alpha siRNA or non-targeting siRNA control for 6 h. Twenty-four hours after transfection cells were preconditioned with hypoxia for 4 h. A : Total RNA was collected immediately after treatment. Image shows RT-PCR analysis for HIF-1alpha and VEGF . 18s rRNA was detected as an internal control. B : Protein analysis of VEGF and HIF-1alpha. Whole cell lysates were collected immediately after treatment, separated by SDS PAGE and probed for VEGF and HIF-1alpha. beta-actin was detected as a loading control. Bar graph shows VEGF expression in indicated groups relative to control. Error bars represent standard error of the mean (n=3).
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
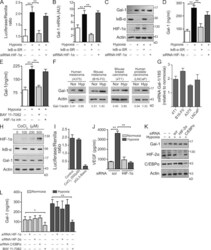
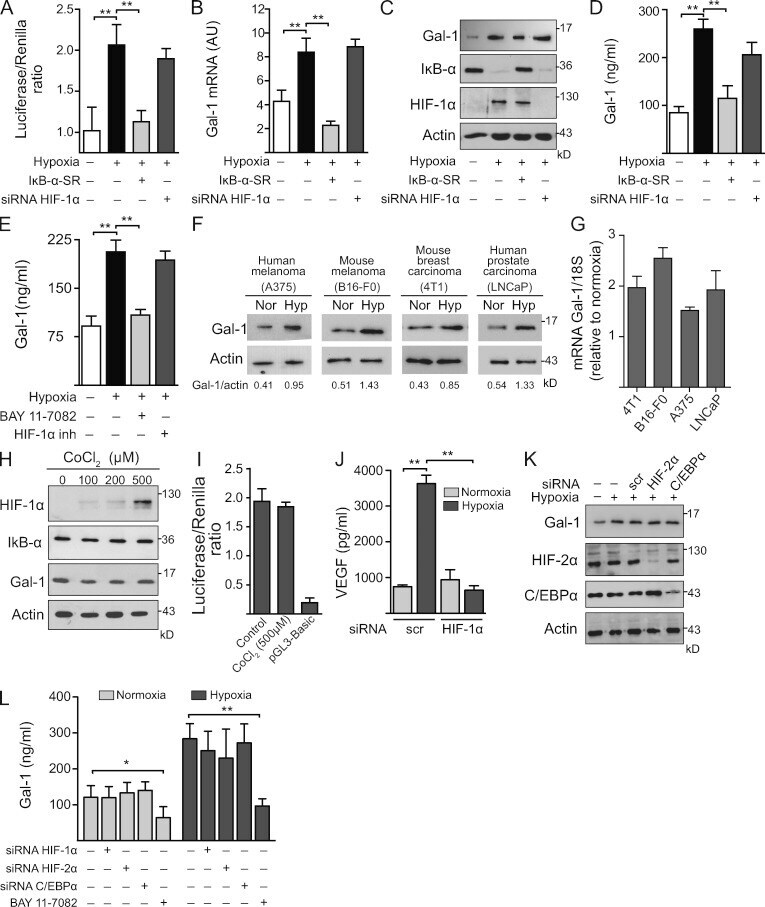
- Figure 3. Hypoxia controls Gal-1 expression in KS through HIF-independent, NF-kappaB-dependent mechanisms. (A-D) Expression of Gal-1 in KS cells transfected with or without HIF-1alpha siRNA or a super-repressor form of IkappaB-alpha (IkappaB-alpha-SR) and incubated under hypoxia or normoxia. (A) Promoter activity. (B) qRT-PCR of Gal-1 mRNA relative to RN18S1. AU, arbitrary units. Data are the mean +- SEM of five (A) or three (B) independent experiments. (C) Immunoblot of Gal-1, IkappaB-alpha, HIF-1alpha, and actin. Data are representative of four independent experiments. (D) ELISA of Gal-1 secretion. Data are the mean +- SEM of three independent experiments. (E) ELISA of Gal-1 secretion by KS cells cultured under hypoxic or normoxic conditions in the presence or absence of HIF-1alpha or NF-kappaB inhibitors. Data are the mean +- SEM of three independent experiments. (F and G) Immunoblot (F) and qRT-PCR (G) of Gal-1 expression induced by hypoxia (Hyp) in human and mouse melanoma (A375 and B16-F0), mouse breast carcinoma (4T1), and human prostate carcinoma (LNCaP) cell lines. Data are representative (F) or are the mean +- SEM (G) of three independent experiments. (H) Western blot of HIF-1alpha, IkappaB-alpha, Gal-1, and actin upon treatment of KS cells with CoCl 2 (chemical activator of HIF-1alpha). Data are representative of four experiments. (I) Gal-1 promoter activity upon treatment of KS cells with CoCl 2 . Modulation of pGL3-Gal-1-Luciferase activity relative to Renilla ex
- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
- Fig. 3 Arginine inhibits HIF-1alpha to reduce LDHA expression after cerebral I/R injury. a Western blotting shows that HIF-1alpha and LDHA are raised in MCAO rats and the increase is attenuated by arginine administration (n = 6 in each group, *p < 0.05 versus sham, # p < 0.05 versus vehicle, two-way ANOVA test). b Western blotting shows that HIF-1alpha and LDHA are raised in in OGD microglia and the increase is attenuated by arginine administration (n = 6 in each group, *p < 0.05 versus sham, # p < 0.05 versus vehicle, two-way ANOVA test). c Western blotting shows that administration of the HIF-1alpha inhibitor, LW6 decreases the protein level of LDHA in MCAO rats and occludes the effect of arginine (n = 6 in each group, *p < 0.05 versus vehicle, one-way ANOVA test). d Western blotting shows that administration of LW6 decreases the level of LDHA in microglia and occludes the effect of arginine (n = 6 in each group, *p < 0.05 versus OGD, one-way ANOVA test). e Western blotting shows that transfection of WT-HIF-1alpha abolishes arginine-induced LDHA reduction in OGD BV-2 cells (n = 6 in each group, *p < 0.05 versus OGD, one-way ANOVA test)